
Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation
by
 , , , and
, , , and
Alexey S. Kiryushkin
1,* ,
,
Elena L. Ilina
1 ,
,
Elizaveta D. Guseva
1,
Katharina Pawlowski
2,* and
and
Kirill N. Demchenko
1,*
1
Laboratory of Cellular and Molecular Mechanisms of Plant Development, Komarov Botanical Institute, Russian Academy of Sciences, 197376 Saint Petersburg, Russia
2
Department of Ecology, Environment and Plant Sciences, Stockholm University, 10691 Stockholm, Sweden
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(1), 51; https://doi.org/10.3390/plants11010051
Submission received: 26 November 2021
/
Revised: 15 December 2021
/
Accepted: 20 December 2021
/
Published: 24 December 2021
(This article belongs to the Special Issue Plant Genome Editing: Progress, Achievements, Applications and Future Prospects)
Abstract
:CRISPR/Cas-mediated genome editing is a powerful tool of plant functional genomics. Hairy root transformation is a rapid and convenient approach for obtaining transgenic roots. When combined, these techniques represent a fast and effective means of studying gene function. In this review, we outline the current state of the art reached by the combination of these approaches over seven years. Additionally, we discuss the origins of different Agrobacterium rhizogenes strains that are widely used for hairy root transformation; the components of CRISPR/Cas vectors, such as the promoters that drive Cas or gRNA expression, the types of Cas nuclease, and selectable and screenable markers; and the application of CRISPR/Cas genome editing in hairy roots. The modification of the already known vector pKSE401 with the addition of the rice translational enhancer OsMac3 and the gene encoding the fluorescent protein DsRed1 is also described.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Interest in the manipulation of genes in the whole plant or its organs has increased since 1983 when several studies first reported the generation of transgenic plants [1]. Hairy root transformation represents a convenient means of studying gene function in plants. This approach is based on the ability of the natural infectious agent, Agrobacterium rhizogenes (Rhizobium rhizogenes) [2], to infect wounded plants, resulting in the development of numerous roots from the wounded site, known as “hairy root disease”. Although different A. rhizogenes strains commonly used for hairy root transformation have been described or reviewed [3,4,5], no report to date has provided a clear description of the relationships among the different A. rhizogenes strains. The nomenclature of related strains can vary greatly due to the transfer of A. rhizogenes strains between laboratories, leading to the erroneous designation of A. rhizogenes strains used in specific studies. Accordingly, a detailed description of A. rhizogenes strain nomenclature is provided here, as is the relationship among the different strains.
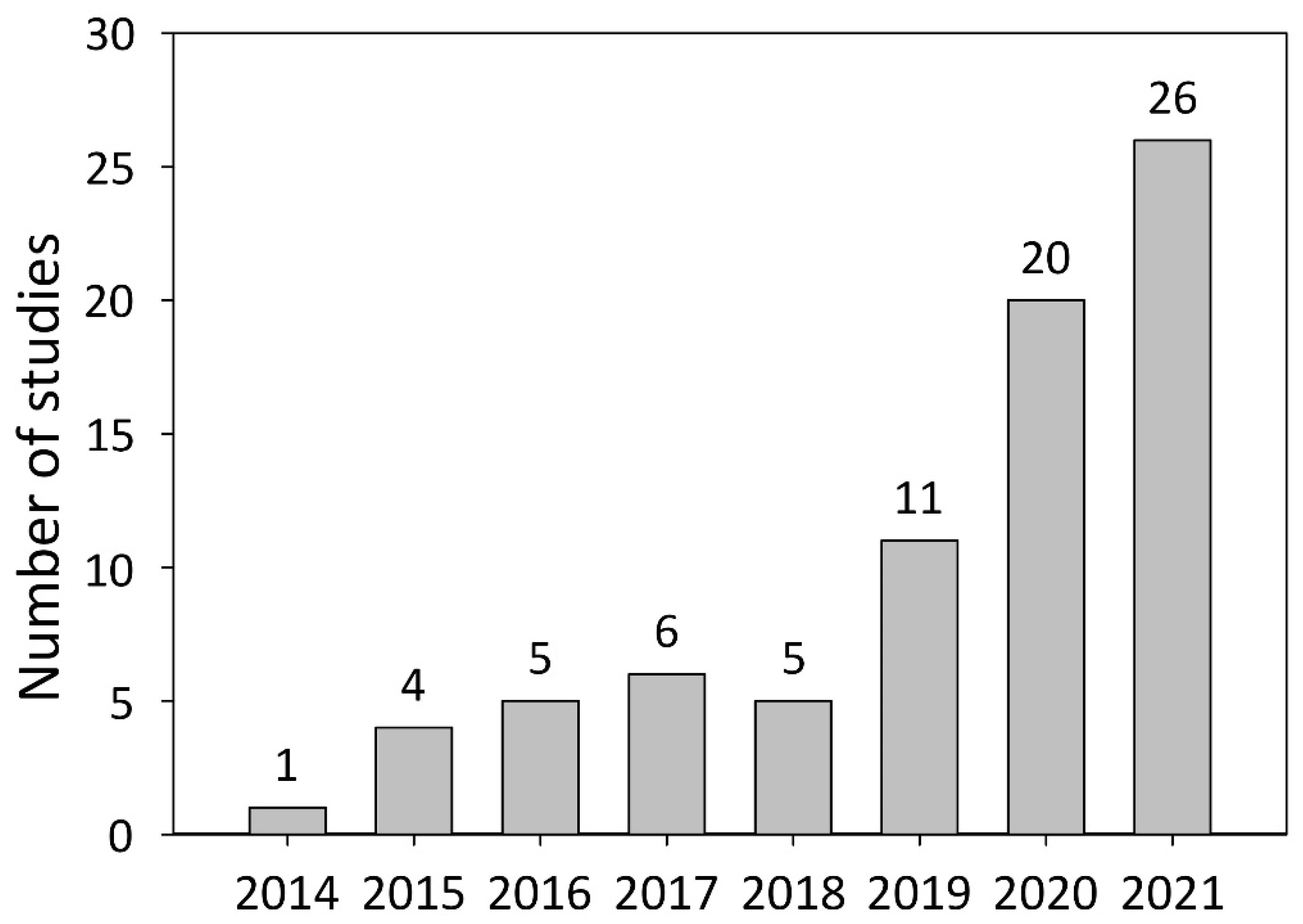
DNA modification systems such as meganucleases, zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and clustered regularly interspaced short palindromic repeats (CRISPR)-associated (Cas) nucleases are frequently used in studies on plant functional genomics. Meganucleases, ZFNs, and TALENs were the predominantly used DNA modification systems in plant research before the advent of the CRISPR/Cas era [1,6]. In 2013, it was shown that CRISPR/Cas-mediated genome editing could be used to investigate gene function in both plant protoplasts and whole plants [7]. The combination of CRISPR/Cas-mediated genome editing and the hairy root transformation approach was reported one year later [8]. Meanwhile, the number of studies combining these two techniques has grown to 78, involving 26 different plant species.
Here, we outline the general rules applicable to CRISPR/Cas vector construction. Accordingly, a part of this review focuses on the principles of CRISPR/Cas vector design. During the construction of CRISPR/Cas vectors, attention should be paid to the following four parts: which promoters to use to drive the expression of genes encoding different Cas nucleases; which Cas-based system to use; the design and construction of guide RNAs (gRNAs) and the assessment of their efficiency; the choice of genes encoding markers for the identification of transgenic roots.
A variety of constitutive promoters, such as the 35S promoter of the Cauliflower Mosaic Virus (p35S) and its variants (2xp35S, 2xp35SΩ, p35SPPDK), are frequently used to drive Cas expression. Other promoters used for CRISPR/Cas-based genome editing include strong promoters, such as pUbi or pActin, as well as inducible or organ- and tissue-specific promoters. The choice of Cas nuclease utilized for genome editing has also varied. Cas nucleases fall into the following three large groups based on their mode of action: those that can introduce double-stranded DNA breaks; those that introduce single-stranded breaks; those that do not introduce breaks. In addition to describing different gene-editing systems, we discuss their use in hairy root transformation, importantly including the choice of marker for the reliable identification of transgenic hairy roots. These markers are commonly divided into two large groups—selectable and screenable (visually) [9]—and we evaluate the advantages and disadvantages of each. New tricks using known markers and several novel markers with potential for future application are also reviewed, as are the research fields where genome editing by hairy root transformation is useful.
2. Agrobacterium rhizogenes: A Historical View of the Widely Used Strains and Their Nomenclature
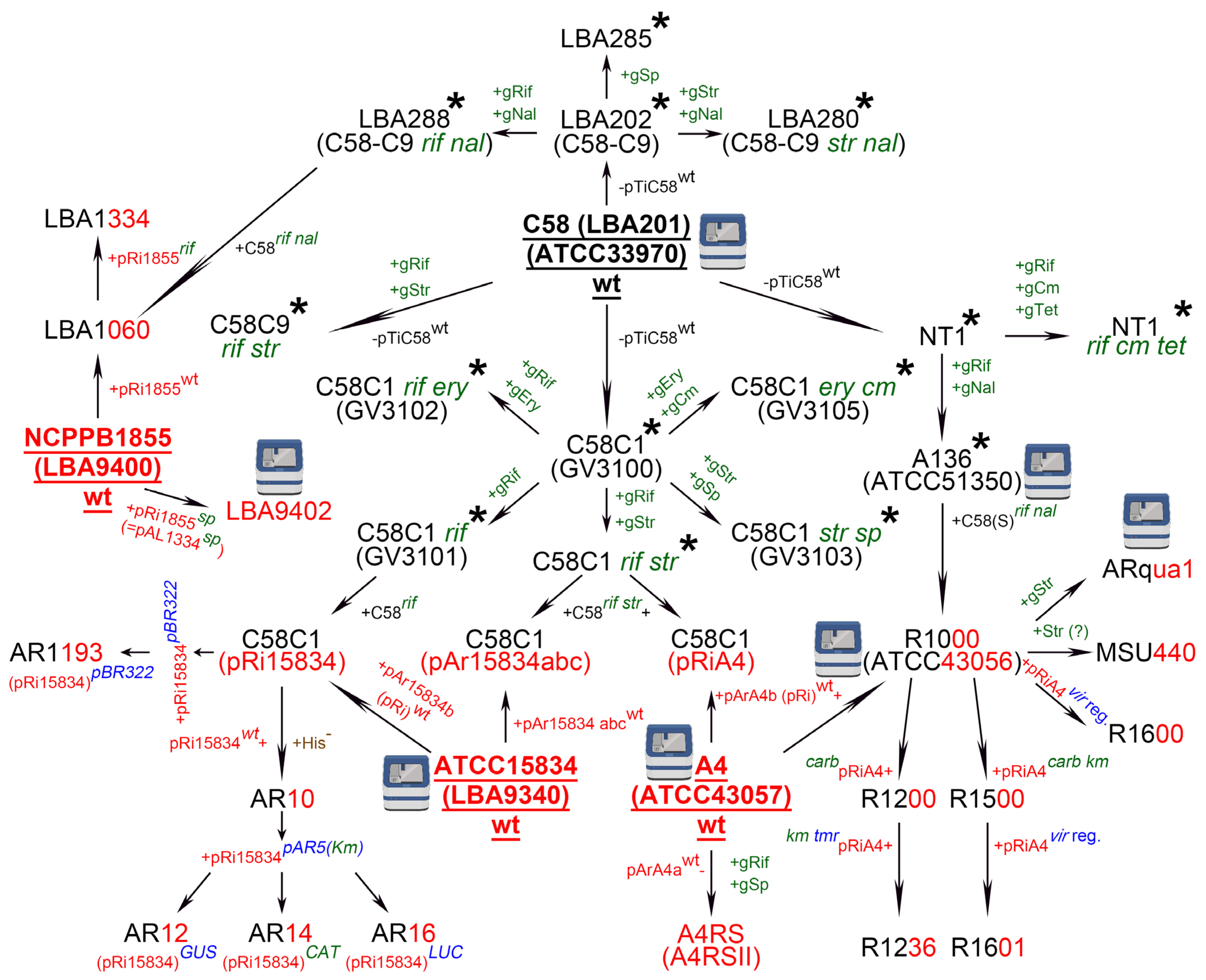
The hairy root transformation approach is based on the infection of wounded plants by the naturally existing soil-borne Gram-negative bacterium, A. rhizogenes. Although more than 20 strains of A. rhizogenes (wild-type or generated via genome engineering) [4] and more than 100 plant species are currently available for hairy root transformation [3,5,10], only 14 A. rhizogenes strains and 26 plant species have been used in genome editing experiments to date (Table S1). Reviews of A. rhizogenes strains are generally restricted to the classification of strains according to the opines they synthesize [4,11]. Here, we have replaced the usual list of A. rhizogenes strains with a scheme describing the relationships among them, similar to that recently presented for A. tumefaciens strains [12].
The 15,834 strain from the American Type Culture Collection (ATCC—also known as ATCC15834, 15834, or AR15834) [13] (Figure 1) was one of the first isolated wild-type A. rhizogenes strains to be widely used for hairy root transformation. The strain was mentioned for the first time in the 1980s by Frank F. White and Eugene Nester while investigating the virulence of root-inducing (Ri) and tumor-inducing (Ti) plasmids (pRi and pTi) [14,15]; however, ATCC15834 was likely isolated earlier, probably in 1971. ATCC15834 was deposited at the ATCC by Richard D. Durbin, who had obtained the strain from Peter A. Ark [13]. Durbin described a highly virulent A. rhizogenes strain that was originally isolated from rose by P.A. Ark [16] and we assume that this was the strain that was later designated as ATCC15834. The ATCC15834 strain harbors three large plasmids, namely, pAr15834a, b, and c (also designated as pAr15834abc) [14] (Figure 1). Because pAr15834b represents the pRi that is responsible for hairy root induction, it is often designated as pRi15834 [14] (Figure 1). The draft genome sequence of ATCC15834 was published for the first time in 2014, based on Illumina sequencing [17], and then again in 2020 based on combined Illumina and Oxford Nanopore sequencing [13] (Figure 1).
Another wild-type A. rhizogenes strain is A4, also designated as ATCC43057 [18] (Figure 1). Presumably, similarly to ATCC15834, this strain was isolated by P.A. Ark from naturally infected roses exhibiting a hairy root phenotype [19]. Again, as with ATCC15834, the A4 strain contains three large plasmids, pArA4a, b, and c, one of which, pArA4b, also called pRiA4, is responsible for hairy root induction [20] (Figure 1). The A4 draft genome was published first in November 2020 by the ATCC based on a combination of Oxford Nanopore with Illumina sequencing [18] and then independently in May 2021 by a research group from the Academia Sinica (Taipei, Taiwan) [21] (Figure 1). In 1987, Frank F. White proposed that A4 might be identical to strain ATCC15834 [22]; however, this possibility has not been tested to date. A derivative of A4 that is resistant to rifampicin and spectinomycin (named A4RS, based on the initials of the antibiotics) and lacks pArA4a [20] (Figure 1) is frequently used for hairy root transformation [23,24,25]. Some studies have used a slightly different designation for this strain, i.e., A4RSII [26,27] (Figure 1).
Two wild-type A. rhizogenes strains from the National Collection of Plant Pathogenic Bacteria (NCPPB), NCPPB2659 (commonly known as K599, not included in Figure 1) [28] and NCPPB1855 [29] (Figure 1), also deserve mention. K599 was isolated from cucumber exhibiting hairy root disease symptoms [30] and has been widely used for hairy root transformation in legumes such as Arachis hypogea, Glycine max, Glycine soja, and Vigna unguiculata (Table S1). Although pRi from K599 was sequenced in 2007 [31], the draft genome of this strain was only published much later, first in 2016 [32], and then in 2021 [33]. Strain NCPPB1855 originates from Rosa sp. [34] and is also known as LBA9400 [35] (Figure 1). This strain gave rise to a rifampicin-resistant derivative, LBA9402 [35], the genome of which was sequenced in 2021. This was the first fully assembled A. rhizogenes genome [34] (Figure 1). The abbreviation LBA stands for Lugdunum Batavorum Agrobacterium [36]. Lugdunum Batavorum is the (erroneous) Latin name for Leiden in the Netherlands. Agrobacterium strains with names beginning with LBA were either isolated at Leiden University or transferred to its strain collection from other laboratories. For instance, ATCC15834 has the designation LBA9340 in the Leiden collection [35] (Figure 1).
Several A. rhizogenes strains currently used for hairy root transformation [4,5] are transconjugants. To understand the relationships between transconjugant and wild-type A. rhizogenes strains, we partially review the history of different A. rhizogenes strains here. Many transconjugant A. rhizogenes strains are closely related to the A. tumefaciens strain C58 and have the C58 chromosomal background along with a pRi from the wild-type A. rhizogenes strains described above (Figure 1). Accordingly, understanding the history of A. rhizogenes strains also requires a review of the data on A. tumefaciens strains. A. tumefaciens strain C58 was originally isolated in 1958 by Robert Dickey (Cornell University, Ithaca, NY, USA) from a cherry gall (C designates cherry and 58 the year of collection). The draft C58 genome was published twice independently in 2001 [37,38] (Figure 1). A comparison of these sequencing data in 2013 showed that the C58 strain originating in Eugene Nester’s laboratory (Washington University, Seattle, WA, USA) was present as two isolates [39]. The first sequenced isolate was the C58 strain stored in the laboratory of Eugene Nester at the University of Washington. The second sequenced isolate also originated from Nester’s laboratory and was deposited at the ATCC in 1981 (ATCC33970) (Figure 1), subcultured three times by the ATCC and once at Monsanto Company, and then sequenced in 2001. The genomes of the two compared C58 isolates showed 30 true differences, including two deletions [39].
The history of the transconjugant A. rhizogenes strains began in the 1970s when it was found that A. tumefaciens C58 lost its pTi plasmid when grown at 37 °C [40,41], leading to the generation of A. tumefaciens strains cured of pTi. Three research groups (one in the USA, one in Belgium, and one in the Netherlands) independently isolated several C58 strains lacking pTi. Eugene Nester’s group (USA) cured the C58 strain (obtained from R. H. Hamilton at Pennsylvania State University) of its pTi using the method of Hamilton and Fall [40] and named the resulting strain NT1 [42] (Figure 1). In the same study, strain A136, which is resistant to rifampicin and nalidixic acid, was isolated from NT1. This strain was later deposited at the ATCC under the number 51350 and its genome sequence was published in 2020 [43]. There is also a derivative of NT1 that is resistant to rifampicin, chloramphenicol, and tetracycline (NT1 rif cm tet) [19] (Figure 1).
A series of C58 strains cured of the pTi was generated in a collaboration between Ghent University, the Vrije Universiteit Brussel (both in Belgium), and the University of Leiden (Netherlands) [12,41,44]. A C58 strain originally obtained from Milton P. Gordon (University of Washington) [41] gave rise to several cured strains (Figure 1), one of which was named C58C1 [44] (with C indicating that it was cured of the pTi) [12] (Figure 1). There are at least five avirulent derivatives of C58C1, each with a different antibiotic resistance profile, namely, resistance to rifampicin and streptomycin (C58C1 rif str) [44]; rifampicin and erythromycin (C58C1 rif ery) [45]; streptomycin and spectinomycin (C58C1 str sp) [46]; rifampicin (C58C1 rif); erythromycin and chloramphenicol (C58C1 ery cm) [47] (Figure 1). In subsequent studies, C58C1 and its antibiotic-resistant derivatives were given new designations beginning with the letters GV (standing for Ghent University and the Vrije Universiteit Brussel [12]), as follows: C58C1, without any antibiotic resistance, was designated GV3100 [47,48]; C58C1 rif was designated GV3101 [47]; C58C1 rif ery was designated GV3102 [46,47]; C58C1 str sp was designated GV3103 [46,47]; and C58C1 ery cm was designated GV3105 [47] (Figure 1). However, we did not find a GV designation for C58C1 rif str from the study of Larebeke et al. [44], and thus this strain does not have a GV name in our scheme (Figure 1). An equivalent of C58C1 rif str described by Larebeke et al. in 1975 [44] was another strain, C58C9, newly isolated by Tempé et al. two years after Larebeke’s study, in 1977 [49]. Because antibiotic resistances of C58C9 are the same as those of C58C1 rif str [44], we designated this strain C58C9 rif str (Figure 1).
Another A. tumefaciens strain, C58-C9, was isolated by the group of Jeff Schell (Leiden University, Netherlands) [50,51] in a manner analogous to that of C58C1 (GV3100). C58-C9, which is cured of the pTi, originated from wild-type A. tumefaciens C58 [41], and was renamed LBA201 according to Leiden University nomenclature [52] (Figure 1). Given that the chromosomal background of C58-C9 was shown to be characteristic of that of A. tumefaciens strain LBA202 [53], and that LBA202 represents C58 cured of its pTi [36,54], we propose that C58-C9 and LBA202 are synonyms for the same A. tumefaciens strain which is designated as LBA202 (C58-C9) in Figure 1. Three spontaneous derivatives of LBA202 (C58-C9) were subsequently obtained. One was resistant to nalidixic acid and streptomycin (LBA280 or C58-C9 str nal); one to nalidixic acid and rifampicin (LBA288 or C58-C9 rif nal) [53,54]; and one to spectinomycin (LBA285) [53] (Figure 1).
Somewhat confusingly, Tempé et al. reported the isolation of a C58C9 strain [49]. The C58-C9 strain from Leiden University has no antibiotic resistance [50,51,53], whereas C58C9 from Tempé et al. is resistant to rifampicin and streptomycin [49] (Figure 1). C58-C9 without antibiotic resistance was reported by both Bomhoff et al. [50] and Ledeboer et al. [51] in 1976, while C58C9 rif str was reported by Tempé et al. in 1977 [49]. Therefore, the C58C9 rif str strain isolated by Tempé et al. [49] could have been a derivative of either LBA202 (C58-C9) or the original wild-type A. tumefaciens C58 [41]. In our scheme, we have placed the C58C9 rif str strain independently from LBA202 (C58-C9) based on the data provided by Tempé et al. [49] (Figure 1).
As both LBA288 (C58-C9 rif nal) from the Leiden collection [53,54] and A136 from the ATCC (ATCC51350) [43] are derivatives of C58 cured of the pTi, and both are resistant to rifampicin and nalidixic acid, they should have the same chromosomal background (Figure 1). However, the sequence of A136 with the chromosomal background of C58 [also named Seattle C58, C58(S)] is slightly different from that of the previously sequenced “Ghent/Leiden C58C” chromosome of nopaline catabolizing, plasmid-cured Agrobacterium strains [55].
A. tumefaciens strain A136, with resistance to rifampicin and nalidixic acid in the C58(S) chromosomal background, and the wild-type A. rhizogenes strain A4 gave rise to A. rhizogenes R1000 [56] (Figure 1). Some studies state that the R1000 pRi is derived from strain A4T [56,57,58,59,60]; however, we did not find any studies describing the isolation of strain A4T. In a personal communication, Frank F. White (University of Florida, Gainesville, Florida, USA) also confirmed that R1000 has the C58(S) background from A136 and the pRi from A4, not A4T (Figure 1). Nevertheless, in several studies, A4T appears as a separate strain used for hairy root transformation [61,62]. Thus, it remains unclear whether A4 and A4T are the same or separate strains (T in A4T might merely indicate that A4 is a type strain), and a comparative analysis of the genome and plasmid sequences of A4, A4T, A136, and R1000 may be required to resolve the uncertainty with this nomenclature. R1000 is also known under its strain collection name ATCC43056 [63] (Figure 1), and in one study as 43056 [64]. The R1000 genome was published by ATCC in April 2021 [63], with the results showing a 96.62% average nucleotide identity (ANI) between the genomes of R1000 (ATCC43056) and C58 (ATCC33970) and 98.48% ANI between the genomes of R1000 and A136 (ATCC51350) [65]. These findings show that R1000 has the A. tumefaciens chromosomal background.
Six strains—R1200, R1236, R1500, R1600, ARqua1, and MSU440—are derived from R1000 (Figure 1). R1200 was obtained by the insertion of a Tn3 transposon conveying carbenicillin resistance (CarbR) into the rol region of R1000 pRiA4 [57] (Figure 1). The cytokinin synthesis locus tmr from pTiA6NC, which encodes an isopentenyl transferase, together with Tn5, which carries a kanamycin resistance (KmR) gene, were subsequently inserted into pRiA4 of R1200 via homologous recombination, yielding R1236 that carries a CarbR gene, a KmR gene, and the tmr locus [57] (Figure 1). R1200 has been widely used for hairy root induction in different species, including belladonna (Atropa belladonna) [66], maroon cucumber (Cucumis anguria) [67], and Tartary buckwheat (Fagopyrum tataricum) [68].
R1500 was generated by the insertion of a chimeric KmR gene into a defective Tn3 transposon (pET23) carrying the CarbR gene, followed by the insertion of the Tn3 transposon into pRiA4 via homologous recombination [69] (Figure 1). R1600 and R1601 were constructed via the modification of, respectively, the R1000 and R1500 strains with the pTVK291 cosmid harboring part of the vir region of pTiBO342 (conferring the supervirulent phenotype of A. tumefaciens A348) [70] (Figure 1). ARqua1 is a streptomycin-resistant derivative of R1000 [71] (Figure 1). The abbreviation ‘AR’ in the ARqua1 strain stands for A. rhizogenes and ‘qua’ for its creator, Hans Joachim Quandt (Inge Broer, personal communication). ARqua1 is also resistant to spectinomycin, because resistance to streptomycin normally provides cross-resistance to spectinomycin (our data, unpublished). The genome of ARqua1 with pRiA4 was sequenced in 2020 [60] (Figure 1).
The A. rhizogenes MSU440 strain was first mentioned in a study by Sonti et al. [72], where the authors stated that the MSU440 strain was obtained from Chris R. Somerville and contains the pRi from the A4 strain [72]. The abbreviation ‘MSU’ stands for the Michigan State University (Chris R. Somerville, personal communication). The origin of the MSU440 chromosomal background was not mentioned in Sonti et al. MSU440 was derived from R1000 and, therefore, has the pRi from the A. rhizogenes A4 strain and the C58 chromosomal background from the A. tumefaciens A136 strain (Chris R. Somerville, personal communication) (Figure 1). MSU440 may also be resistant to streptomycin [73] (Figure 1).
The chromosomal background of A. tumefaciens C58C1 rif str together with the pRi from A. rhizogenes A4 or pAr15834abc from ATCC15834 gave rise to, respectively, the C58C1 (pRiA4) and C58C1 (pAr15834abc) strains [74] (Figure 1). Besides C58C1 (pAr15834abc), a rifampicin-resistant C58C1 strain containing only pAr15834b (pRi15834) also exists [75,76,77], which we have designated C58C1 (pRi15834) (Figure 1) based on the fact that two separate strains—C58C1 (pAr15834abc) rif str [74] and C58C1 (pRi15834) rif [75,76,77]—are described in the literature. However, we did not find any studies describing a transconjugation between the chromosomal background of C58C1 rif (GV3101) and the pRi from strain ATCC15834.
Similarly to De Saeger et al. [12], who highlighted that the nomenclature of some published A. tumefaciens strains is frequently confusing, incomplete, or misleading, we note with regret the same fact about A. rhizogenes strains that have the A. tumefaciens C58C1 chromosomal background. For instance, in some studies on hairy root transformation, the A. rhizogenes strain used was indicated only by its chromosomal background C58C1 (see Table S1 [63,69,73]). We believe that it would be more appropriate to also indicate the pRi that was introduced into the A. tumefaciens strain. Good examples are transconjugant A. rhizogenes strain names such as C58 (pRiARqua1) and C58C1 (pRiA4) (see Table S1 [6,70,71]).
C58C1 (pRi15834) rif gave rise to several derivatives, including AR1193 [77], AR10, AR12, AR14, and AR16 [75]. The letters “AR” in these strain names presumably also stand for “A. rhizogenes”. AR1193 was obtained by the integration of a fragment from pBR322 [78] into the wild-type pRi15834 TL-DNA from strain C58C1 (pRi15834) rif, thus forming the basis for further insertion of target genes into the pRi by homologous recombination [77] (Figure 1). The name pBR322 follows the standard rules of plasmid nomenclature, i.e., the “p” stands for plasmid and the “BR” for the names of its creators Francisco Bolivar and Raymond L. Rodriguez, while the numeric designation, 322, stands for the number of the vector in the collection [79]. Strain AR10 represents a histidine auxotrophic mutant (his−) of C58C1 (pRi15834) rif [75] (Figure 1). This mutant was isolated to improve the counterselection of A. rhizogenes during the removal of the strain from plants after inoculation. Normally, antibiotics such as cefotaxime, augmentin, and timentin are used to decontaminate plant tissue [80,81]. The histidine auxotrophic derivative of A. rhizogenes AR10 can be easily removed as it cannot survive in medium without histidine [75]. AR10 his− gave rise to the strains AR12, AR14, and AR16 [75] (Figure 1), which were obtained via homologous recombination between the wild-type pRi15834 plasmid of AR10 and the pBR322-based plasmid, pAR5, harboring genes encoding β-glucuronidase (GUS), chloramphenicol acetyltransferase (CAT), or luciferase (LUC), respectively (Figure 1). Thus, the AR12, AR14, and AR16 strains have modified T-DNA containing selectable (CAT) or screenable (GUS or LUC) marker genes that can be used for the identification of transgenic hairy roots [75]. Moreover, part of the pBR322 sequence that is transferred from pAR5 to pRi15834 can form the basis for further insertion of another target gene into pRi15834 by homologous recombination [82].
The transconjugant A. rhizogenes LBA1334 strain has the C58C9 chromosomal background with rifampicin resistance [83] and the pRi from NCPPB1855 that carries a spectinomycin resistance gene [83,84] (Figure 1). At least two A. tumefaciens strains, C58C9 rif str [49] and LBA288 (C58-C9 rif nal) [53,54], have the C58C9 background with rifampicin resistance (Figure 1), rendering it problematic to identify the original A. tumefaciens strain that provided the chromosomal background for the A. rhizogenes LBA1334 strain. LBA1334 was obtained in two steps (Paul J. J. Hooykaas, personal communication). First, A. rhizogenes strain LBA1060 was obtained via the transfer of the pRi of A. rhizogenes NCPPB1855 into the chromosomal background of the A. tumefaciens strain LBA288 (C58-C9 rif nal) [53,54] (not that of the A. tumefaciens C58C9 rif str strain obtained by Tempé et al. [49]) (Figure 1). In the second step, the pRi of NCPPB1855 in LBA1060 was modified by adding a spectinomycin resistance gene (the pRi was named pAL1334) [83,84] (Figure 1). The AL in the pAL1334 stands for Agrobacterium Leiden. Strain LBA1334 also carries a nalidixic acid resistance gene from the C58C9 chromosomal background of LBA288 (C58-C9 rif nal) [53,54] (Figure 1). LBA1334 might also display chloramphenicol resistance [85] (Figure 1) given that resistance to nalidixic acid normally provides cross-resistance to chloramphenicol (Paul J. J. Hooykaas, personal communication).
The A. rhizogenes strains described above have various levels of virulence reflected by their ability to induce hairy root formation in different plant species [3]. One way to increase hairy root transformation efficiency is to change the virulence of A. rhizogenes strains, as has been done for the R1600 and R1601 strains [70]. The initial strains, R1000 and R1500, from which R1600 and R1601 were isolated (Figure 1), could not induce hairy roots on hybrid poplar (Populus trichocarpa × deltoides). Enhancing the virulence of R1000 and R1500 by adding the pTVK291 cosmid that carries part of the vir region of pTiBO342 (conferring the supervirulent phenotype to the A. tumefaciens strain A348) permitted the generation of hybrid poplar hairy roots and the subsequent regeneration of shoots from those hairy roots [70]. A similar modification was carried out for A. tumefaciens, i.e., the stable transformation rate of tomato (Solanum lycopersicum) with the resulting strain was increased 3.6-fold compared with that for the original GV2260 strain [86].
CRISPR/Cas9-mediated genome editing of existing strains may represent an additional means of obtaining A. rhizogenes with higher virulence [4,5]. A prerequisite for successful genome editing of A. rhizogenes strains is the availability of a genome sequence, i.e., the sequence of the chromosome and the pRi. To date, the chromosomes and Ri plasmids of several A. rhizogenes strains—ATCC15834 [13,17], A4 [18,21], NCPPB2659 (K599) [31,32,33], LBA9402 [34], ARqua1 [60], and R1000 [63]—have been sequenced (Figure 1). CRISPR/Cas9-based editing was performed for several NCPPB2659 (K599) genes, namely cus, encoding cucumopine synthase, rolB, rolC, and orf13. Proof of concept that genomes of Agrobacterium strains could be successfully edited was obtained in that the induction of hairy root formation on carrot disks by K599 rolB and rolC mutants was negatively affected [33]. Comparative analysis of the already sequenced genomes of A. rhizogenes strains will help to identify the parts that most strongly affect their virulence and thus help to obtain new supervirulent A. rhizogenes strains.
3. Editing the Plant Genome in Transgenic Hairy Roots: Vector Components
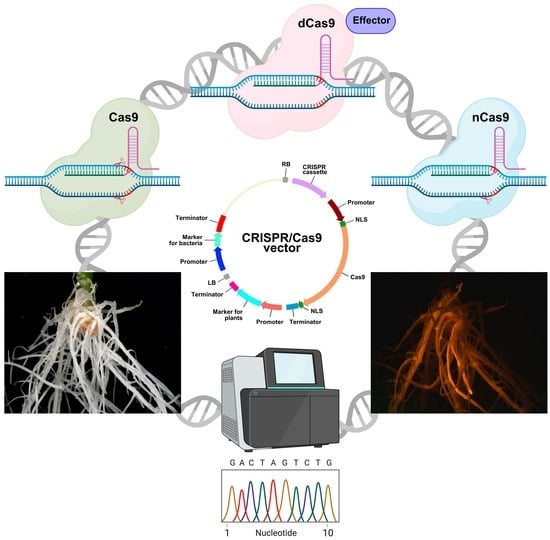
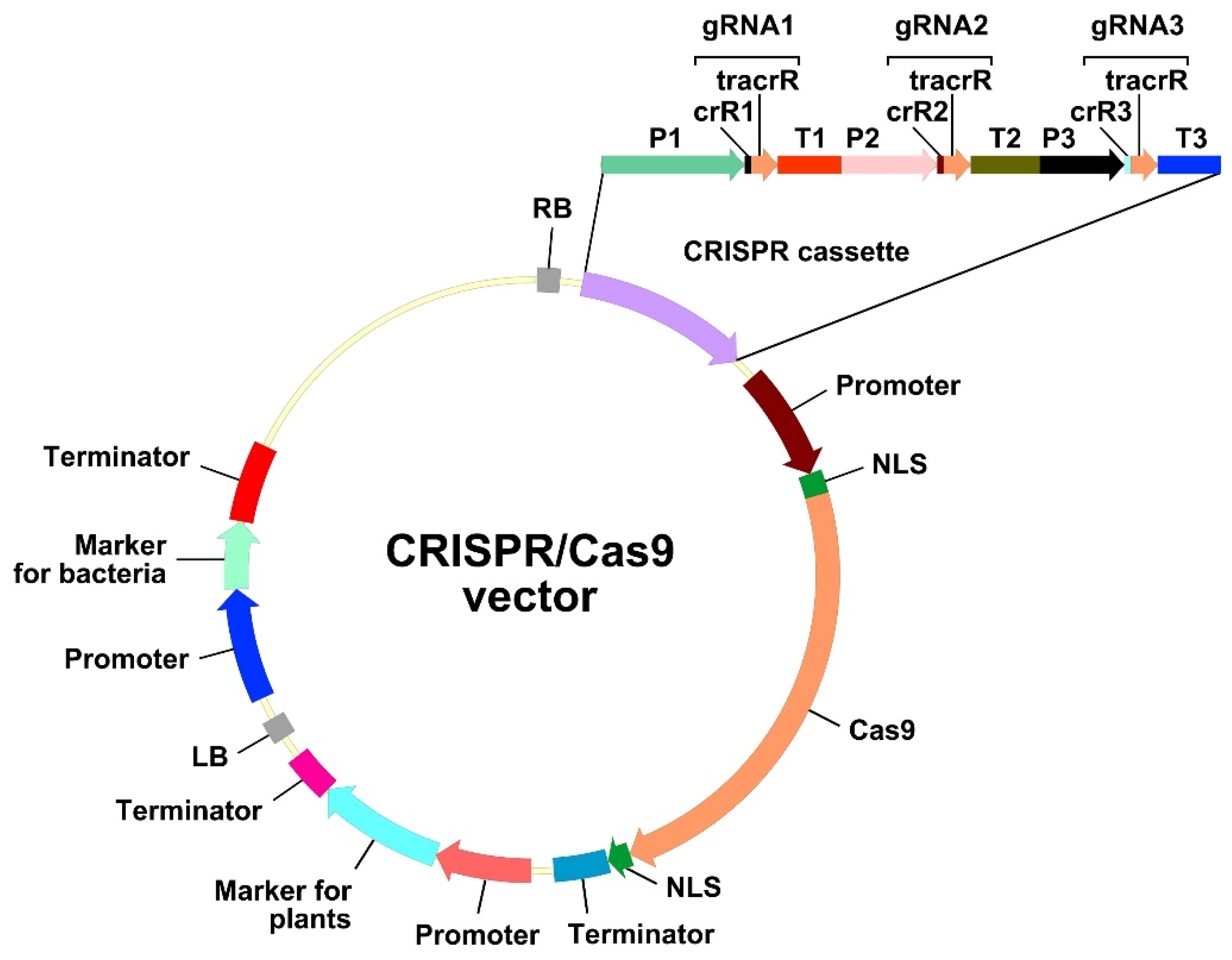
Hairy root transformation has long been used for the modification of plant traits, either because no protocols for stable transformation and regeneration were available, or because the targeted trait was only observed in roots [87]. Another reason for the use of hairy roots is that A. rhizogenes-mediated transformation leads to the quick regeneration of the transgenic biomass, which is necessary for the fast production of biomedical/pharmaceutical/industrial molecules of interest [10]. CRISPR/Cas-mediated genome editing can be performed in transgenic hairy roots for the same reasons. A construct for efficient genome editing and selection of hairy roots should contain three components, i.e., a cassette carrying the gene encoding the CRISPR-associated (Cas) nuclease, a cassette expressing the guide RNA (gRNA), and a cassette encoding a screenable or selectable marker (Figure 2).
3.1. Cassette for Cas Expression
The construction of a cassette for Cas expression in hairy roots depends on the aim of the study. Strong constitutive promoters are typically used to control Cas expression in plants. The Cauliflower Mosaic Virus (CaMV) 35S promoter (p35S) [88] is often (in 53 of the 78 studies that have employed CRISPR/Cas in hairy roots) used to drive expression of Cas9 (Table S1). Ubiquitin promoters (pUbi) from different species, including Arabidopsis, parsley (Petroselinum crispum), maize (Zea mays), rice (Oryza sativa), and soybean (G. max), are also popular choices; Cas9 expression was driven by pUbi in 19 of the 78 studies examined (Table S1). Other strong promoters have been used in individual cases. For instance, the promoter of the Arabidopsis actin2 gene (pAct2) was used in hairy roots of hybrid poplar (Populus tremula × alba). In other studies, the 35S enhancer was fused to the maize C4 pyruvate orthophosphate dikinase (C4PPDK) basal promoter (p35SPPDK) [89] to drive Cas9 expression in hairy roots of rubber dandelion (Taraxacum kok-saghyz), while the soybean pSCREAM M4 promoter (pM4), which drives the strong, constitutive expression of elongation factor 1A [90], was employed for Cas9 expression in soybean hairy roots. Additionally, in one instance, an organ-specific promoter (the promoter of the nodule-specific leghemoglobin b2 gene of the legume Lotus japonicus [pLjLb2]) was used to drive Cas9 expression in hairy roots of the same species (for pAct2, p35SPPDK, pM4 and pLjLb2 see Table S1 [25,46,52,66,84]).
Recent advances in plant genome editing have involved the construction of vectors in which tissue-specific [91] or inducible [92] promoters drive Cas9 expression. Although not all of these genome editing technologies have been applied to hairy roots, they undoubtedly have great potential in the field of root developmental biology. Hairy root transformation, combined with inducible CRISPR/Cas-mediated genome editing, should be used if the target gene has a pleiotropic effect on root development. For instance, the tomato SHOOT BORNE ROOTLETS (SBRL) gene is involved in the development of both adventitious and lateral roots [93]. In addition to having fewer lateral roots, tomato CRISPR/Cas sbrl mutants cannot develop adventitious roots if the main root is removed [93]. Consequently, investigating SBRL gene function during lateral root development using hairy root transformation with vectors constitutively expressing Cas9 will likely not be possible because the edited hairy roots will not develop. CRISPR/Cas-mediated tissue-specific knockout can be used in hairy roots for the same reason, namely, to avoid a pleiotropic effect. Another advantage of tissue-specific genome editing is that Cas9, expressed in a tissue-specific manner, can cause mutations in cells at the earliest stages of tissue or organ development. For instance, the Arabidopsis GATA23 promoter (pAtGATA23) [94] is active exclusively at the earliest stages of lateral root initiation (LRI) [91,95], and genes responsible for LRI in Arabidopsis and other higher plants can be edited at the earliest stages of LRI using Cas9 expression under the control of pAtGATA23 [91]. We have recently identified several genes expressed during the early stages of LRI in plants such as squash (Cucurbita pepo) and cucumber (Cucumis sativus) that display an alternative root branching mechanism [96], namely GATA24 and MEMBRANE-ASSOCIATED KINASE REGULATOR4 from squash (CpGATA24 and CpMAKR4) [97] and RAPID ALKALINIZATION FACTOR34 from cucumber (CsRALF34). Studies of the expression of pCpGATA24, pCpMAKR4 [97], and pCsRALF34 (Figure 3) promoter reporter fusions in the parental root tip have shown that, in a similar way to pAtGATA23, these promoters are active in the earliest stages of LRI, and therefore can be used for tissue-specific Cas9 expression in LRI studies involving Cucurbitaceae species.
Translational enhancers can be used to increase Cas9 translational efficiency, such as the enhancers from the Cowpea Mosaic Virus 5′- and 3′-untranslated regions (UTRs) employed for CRISPR/Cas9 genome editing in hairy roots of Abyssinian mustard (Brassica carinata). The CaMV 5′-UTR (omega enhancer) was applied in belladonna, the legume L. japonicus, potato (Solanum tuberosum), and tomato. The 5′-UTR of the Arabidopsis alcohol dehydrogenase (AtADH) gene (TAIR ID: AT1G77120) was used as a translational enhancer for genome editing in soybean hairy roots (for enhancers, see Table S1 [4,5,6,24,53,76]).
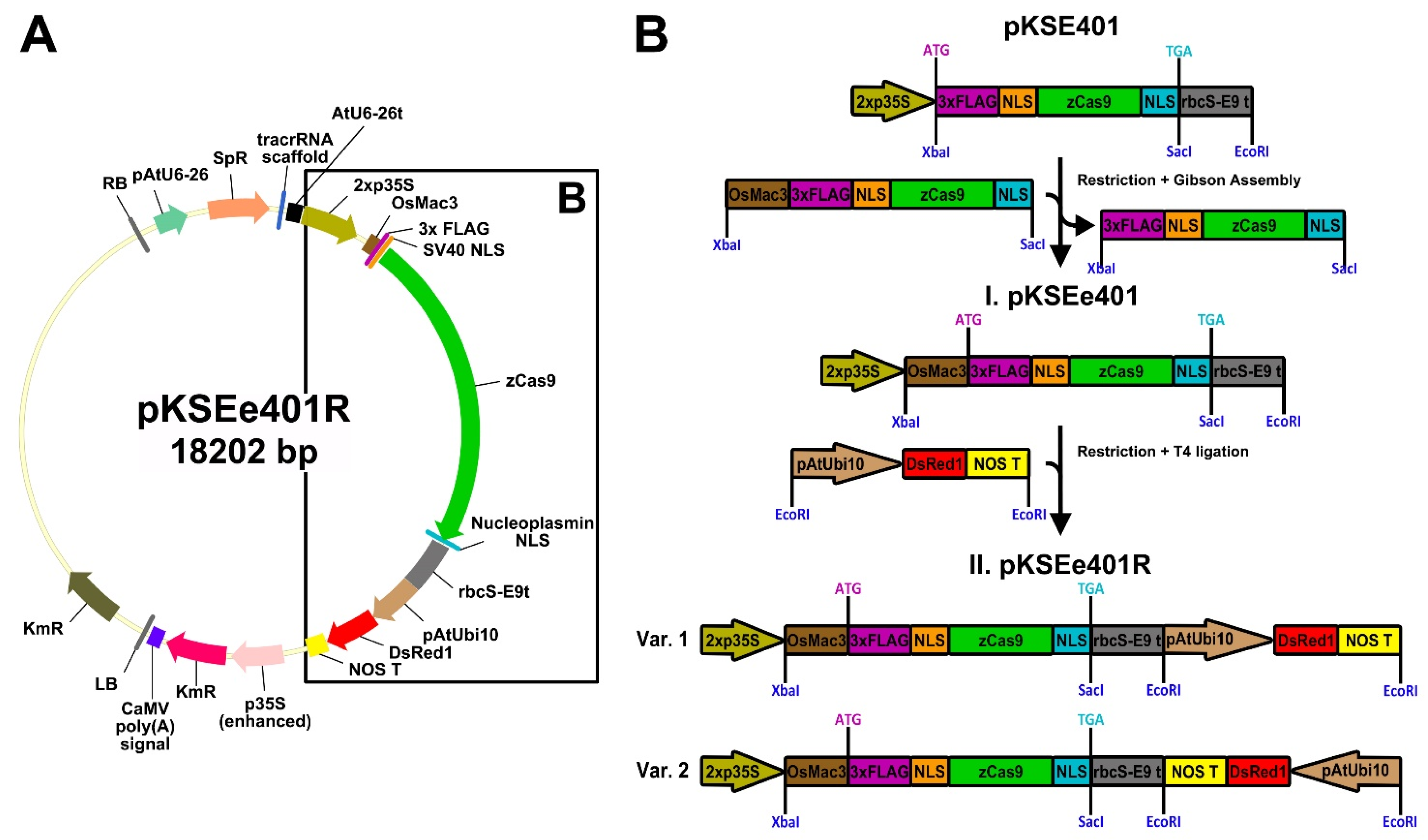
Another translational enhancer is the 5′-UTR of the Mac3 gene from japonica rice (OsMac3) [98]. An OsMac3 5′-UTR fragment (from −158 to −1 bp before the ATG) [99] was shown to exhibit sufficient activity as a translational enhancer [98] and can be used to improve genome editing in both monocots [99] and dicots [100]. The OsMac3 enhancer has been used in stable transformants. To enhance the translational efficiency of Cas9-based genome editing in hairy roots, we modified pKSE401 [101] to include the OsMac3 5′-UTR fragment (Figure 4A,B). The OsMac3 5′-UTR sequence was cloned from genomic DNA of japonica rice O. sativa cv. Flagman (Rice Research Institute, Krasnodar, Russian Federation), and then fused with the maize codon-optimized Cas9 (zCas9) from pKSE401 [101] via Gibson Assembly (Gibson Assembly® Master Mix, New England Biolabs, Ipswich, Massachusetts, USA) [102]. The resulting plasmid was named pKSEe401 (Figure 4B), with the “e4” in pKSEe401 representing enhanced zCas9, in contrast to the unenhanced zCas9 previously indicated by the digit 4 in pKSE401 [101].
The second important component of a CRISPR/Cas cassette is the Cas gene. As a rule, the Cas9 endonuclease of Streptococcus pyogenes and its modified versions are used for targeted gene editing; however, other Cas nucleases (e.g., SpRY or Cas12 versions) have also been employed [103,104,105]. For effective editing in plants, a Cas gene should be codon-optimized for use in dicots/monocots, or at least for the plant kingdom. However codon optimization of a Cas gene is not strictly required, because it does not guarantee 100% efficiency of genome editing. Moreover, there are examples where codon optimization was not performed, but editing was effective nonetheless. In 12 of the 78 studies examined, zCas9 was used for genome editing in dicots, including Madagaskar periwinkle (Catharanthus roseus), soybean, liquorice (Glycyrrhiza glabra), Chinese liquorice (G. uralensis), and the legume Medicago truncatula (Table S1). Furthermore, in 19 of the 78 evaluated studies, a Cas9 gene codon-optimized for humans was successfully used for hairy root-based genome editing (Table S1). The efficiency of genome editing can, however, be increased by introducing introns in the Cas9 gene [106]. In addition, nuclear localization signals (NLSs) (two are sufficient) must be attached to the Cas9 open reading frame (ORF) to ensure its nuclear targeting [107] (Figure 2).
Although there are several CRISPR/Cas technologies based on different Cas activities [104,108], the underlying molecular mechanism is the same in all cases. The two main components of such systems are the gRNA and the Cas nuclease. The gRNA consists of the following two parts: the crisprRNA (crRNA) and the trans-activating crRNA (tracrRNA). The gRNA binds to the target genomic DNA in front of a specific protospacer adjacent motif (PAM). The fate of the target genomic DNA sequence linked with the gRNA/Cas ribonucleoprotein complex depends on the type of Cas activity.
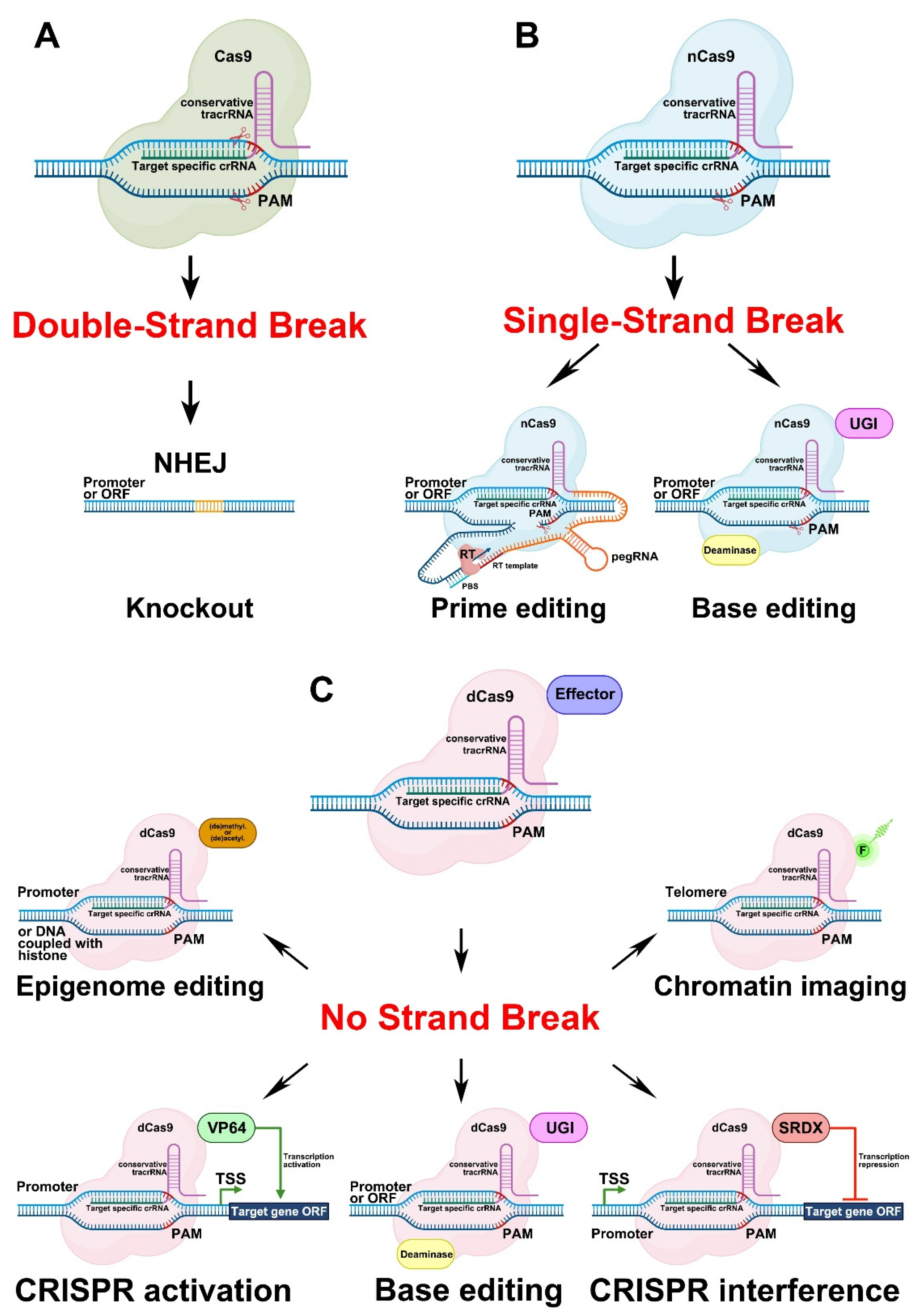
3.1.1. Genome Editing Based on Double-Strand Breaks in DNA Caused by Cas9 Activity
The most frequently used genome editing system involving Cas is based on the generation of double-strand breaks (DSBs) in the target DNA (Figure 5A). After the introduction of DSBs, non-homologous end joining (NHEJ), a DNA repair mechanism [109], leads to the development of different insertions or deletions (indels) in double-stranded DNA, thereby resulting in lesions in the ORFs (frameshifts) or promoter regulatory elements of target genes.
Genome editing via the introduction of DSBs in the ORFs is the most commonly employed of the CRISPR/Cas-based methods in hairy roots (it was applied in 76 of the 78 studies examined; Table S1). In contrast, two of the 78 reports examined do not concern changes in ORFs. One of these entailed large chromosomal deletions without ORF changes in soybean hairy roots (see Table S1 [37]), while the other involved the editing of a promoter region in tomato hairy roots (see Table S1 [77]). Here, a G-box element targeted by the transcription factors (TFs) MYC1/MYC2/GAME9 in the promoter of the tomato cholesterol biosynthesis gene STEROL C-5(6) DESATURASE 2 (C5-SD2) was successfully edited.
3.1.2. Genome Editing Based on Single-Strand Breaks in DNA Caused by Cas9 Activity
Another group of CRISPR/Cas approaches uses the nickase form of Cas9 (nCas9), which causes single-stranded breaks (SSBs) in DNA. This group comprises two types of genome editing systems—prime editing (PE) and base editing (Figure 5B).
The PE approach was introduced in 2019 for mammalian cells [110] and has been widely used in different fields of biology since its invention. The PE complex consists of two components, namely, a Cas nickase (nCas9) fused with a modified reverse transcriptase (RT) via a linker (Figure 5B). The PE gRNA (pegRNA), in turn, also comprises several parts fused into a single sequence—the crRNA, the tracrRNA, the linker sequence, the primer binding site (PBS), and the template for reverse transcription (Figure 5B). Directed by the pegRNA, nCas9 creates an SSB in front of a PAM in the target genomic DNA sequence. The PBS, which is complementary to the DNA strand, binds to the target sequence in front of the SSB. The nucleotide sequence behind the PBS is used as a template by the RT (Figure 5B). The new sequence formed by the RT can be reincorporated into the target DNA, which leads to mutations, or can be removed by exonuclease activity, which means that the target sequence remains unchanged [110,111,112]. Cases of PE have been reviewed for both transient (protoplasts) and stable plant transformation systems; however, the efficiency of this technology in plants was found to be relatively low [103].
Two classes of enzymes fused with nCas9 are applied for base editing, cytidine (CD) and adenine (AD) deaminases. Such fusions result in the development of the two types of base editors, cytosine (CBE) [113] and adenine (ABE) base editors [114] (Figure 5B). The CBE can change cytosine (C) to thymine (T) or guanine (G) to adenine (A) in the complementary DNA strand, while the ABE performs the opposite substitutions, i.e., T to C or A to G. Briefly, the base editing mechanism works as follows: nCas9, fused with an appropriate deaminase, is directed by the gRNA to the target DNA site and creates an SSB. Deaminase transforms the appropriate nucleotide on the DNA strand with the SSB. The CD transforms C to uracil (U) and U, in turn, is converted to T via DNA repair or replication [113]. To prevent the reversion of U to C via base excision repair, the uracil-DNA glycosylase inhibitor (UGI) domain is included in the CBE (Figure 5B). AD converts A to inosine (I), which is recognized as G by polymerases [114]. The edited strand with the changed nucleotide is then used as a template by the repair mechanism. The development of the base editing technology resulted in improvements in the CBEs [115,116,117,118,119] and ABEs [120,121] characteristics, such as reduced off-target activity, indel frequency, and nCas9 expression levels. Recently, double base editors with CD and AD activities were also developed, first for mammalian cells [122,123,124,125], and then for plants [126,127].
The applications of prime or base editing approaches have been reviewed for some plant species [103,128,129]. Prime and base editing are more accurate but less efficient than the repair of DSBs resulting from Cas9 activity. These approaches need to be optimized to enhance efficiency when used in plants [103,129], and may explain why no studies using prime or base editing on hairy roots have been published to date. Nevertheless, cases of prime or base editing have been reported for several species, including tomato, cotton (Gossypium hirsutum), Nicotiana benthamiana, potato, rapeseed (Brassica napus), and watermelon [103,128,129], for which hairy root transformation protocols are now available [8,130,131,132,133,134]. Besides prime or base editing, nCas9 can be used for gene knock-out, similar to how it was described for potato hairy roots (see Table S1 [79]).
3.1.3. Genome Editing Based on a Version of Cas9 That Causes no Strand Breaks
The catalytically inactive (dead) form of Cas9 (dCas9) has also become an important instrument for biologists. The effect of the dCas9-based system depends on the effector fused to the dCas9. Among effectors, there are activation (AD) or repressor (RD) domains, base editors, epigenome modificators as well as fluorescent proteins [135] (Figure 5C).
Similarly to nCas9, dCas9 can be fused with deaminases (Figure 5C), but the base editing efficiency of dCas9–deaminase systems is lower than that of systems involving nCas9 [113]. Nevertheless, base editing using dCas9 has also been reported for plants [136,137].
Another application of dCas9 is the creation of CRISPR activator (CRISPRa). The principle underlying the activity of CRISPRa is based on DNA binding by an AD fused to dCas9 and directed to the target sequence by the gRNA, which leads to the activation of the expression of the target gene (Figure 5C). The most commonly used AD for fusion with dCas9 is derived from the herpes simplex viral protein 16 (VP16) (UniProt ID—P06492) [138]. The AD represents a tetrameric repeat of the minimal VP16 AD (DALDDFDLDML) separated by glycine–serine linkers (GS), termed VP64 ([DALDDFDLDML]-GS-[DALDDFDLDML]-GS-[DALDDFDLDML]-GS-[DALDDFDLDML]) [139].
The CRISPR interference technology (CRISPRi) is based on the principle, that gRNA/dCas9 ribonucleoprotein (RNP) complex interfere with RNA Polymerase II following block of transcription initiation or elongation. The dCas without any repressor domain (RD) was effective for bacteria, but not for yeast and mammalian cells [140]. Apparently, it is less effective for plants too; therefore, to improve repression efficiency of CRISPRi in plants the additional RD started to use. The RD originates from investigations of the repressor activity of a 12-aa sequence (LDLDLELRLGFA) from the Arabidopsis SUPERMAN protein (TAIR ID: AT3G23130) [141]. The fusion of these 12 aa from SUPERMAN (SUPERMAN Repression Domain X [SRDX]) to the C-terminus of other TFs turned them into strong repressors [142].
The CRISPRa and the CRISPRi systems, based on fusions of VP64 or SRDX with dCas9, respectively (Figure 5C), were adapted for plants in 2015 [143,144]; however, compared with the use of CRISPR knockout systems, activator or repressor systems are still rarely used in plant science [145,146,147,148]. Besides VP64 and SRDX, several other ADs [149] and RDs [150] are still awaiting co-application with CRISPR technology or were only rarely used, such as EDLL [143,147], TAL AD (TAD) [143], and modified ERF2 (ERF2m) [145].
dCas9 can also be used for epigenome editing as well as for chromosome visualization (Figure 5C). In the former, dCas9 can be directly fused to different DNA methyltransferases/demethylases or histone acetyltransferases/deacetylases (Figure 5C) [151]. Several studies on epigenome editing in plants have been reviewed previously [151]. Direct fusions of dCas9 with fluorescent proteins have been used for chromosome imaging (Figure 5C) [152,153,154].
In addition, the dCas9-based systems can be classified according to the strategies used to recruit effectors for improving the efficiency of dCas9-based system. The greatest variety of approaches was reviewed for CRISPRa and CRISPRi [135,140]. Reviewed strategies can be extrapolated to other effectors (e.g., base editors, epigenome modificators as well as fluorescent proteins). Multiple copies of the same effector, or different effectors linked in tandem can be used. Another approach is the so-called synergistic activation mediator (SAM) system, based on the co-expression of the dCas9 with modified gRNA and different effectors fused with special proteins that recruit effectors to the dCas9/gRNA RNP complex through their RNA-binding activity [155]. The SunTag system is based on the co-expression of epitope-tagged dCas9 and effectors fused to the corresponding antibody [156]. The effector is attracted to the dCas9/gRNA RNP complex trough the recognition of epitope by the antibody.
Despite the successful use of some effectors in hairy roots, e.g., of SRDX [157,158] or fluorescent proteins [97,159] outside the dCas9-based system, we note that neither these techniques, nor base and prime editing, has yet been commonly applied for hairy roots. Nevertheless, we found the one attempt to obtain carrot hairy roots harboring the SAM system for telomere imaging [154], but results about SAM efficiency in hairy roots were not reported, rendering it difficult to evaluate the efficiency of SAM system for telomere imaging in hairy roots.
3.2. gRNA: Design and Testing
The cassette expressing the gRNA in the CRISPR/Cas vector comprises the promoter of the small nucleolar RNA (snoRNA) gene, the gRNA sequence, and the snoRNA transcription terminator (Figure 2). The gRNA sequence, in turn, consists of two fused sequences, namely, the 17–20-bp crisprRNA (crRNA) and the 80-bp trans-activating crRNA (tracrRNA) (Figure 2). The crRNA is complementary to the target genomic DNA; thus, its sequence is variable and study-specific. The sequence of the tracrRNA is conserved and functions as a binding scaffold for a Cas nuclease; consequently, it is usually already incorporated in the CRISPR/Cas vector backbone.
snoRNA gene promoters are recognized by both RNA polymerase II (U-snoRNA classes U1, U2, U4, and U5) and RNA polymerase III (Pol III) (U-snoRNA classes U3 and U6) [160,161]. Given that transcribed gRNAs should remain in the nucleus to guide Cas9 to the target genomic DNA sequence, only snoRNA gene promoters recognized by Pol III are used in CRISPR/Cas systems. Different Pol III promoters are normally used for expression of several gRNAs in one CRISPR cassette (Figure 2). When assembling multiple gRNAs, the strategy of using different Pol III promoters greatly increases the size of the cloned CRISPR cassette. Meanwhile, vectors using a polycistronic gRNA strategy based on different processing systems have also been published. The expression of multiple gRNAs in a polycistronic cassette is driven using only one Pol III promoter. A single gRNA can be formed and later cleaved either by systems based on tRNA processing mechanisms [162] or by RNA endoribonuclease Csy4 from Pseudomonas aeruginosa (Csy4) [163]. The tRNA processing system, which exists in all living organisms, precisely cleaves at the end of a tRNA precursor sequence added to gRNA. Unlike the tRNA processing mechanism, the Csy4 activity does not occur naturally in plants. Therefore, its ORF must be fused to the Cas ORF via an ORF encoding a short, self-excising peptide, 2A. Despite the advantages of a polycistronic CRISPR cassette, such as the reduction of the size of constructed vector and an increase in the number of gRNAs, this strategy of genome editing has rarely been used on hairy roots (5 of the 78 studies examined), compared to the use of a CRISPR cassette with different Pol III promoters for expression of several gRNAs (for tRNA and Csy4 processing mechanisms see Table S1 [4,5,53,76,79]).
Active promoters of plant snoRNA genes were first characterized in Arabidopsis in the 1990s. Five U6 genes and promoters were identified—U6-1 (TAIR ID: AT3G14735), U6-26 (TAIR ID: AT3G13855), U6-29 (TAIR ID: AT5G46315) [160], U6-22 (GeneBank ID LR782545.1; position: plus strand, 13922611–13922735 bp), and U6-25 (GeneBank ID LR699773.1; position: minus strand, 16444343–16444219 bp) [161]; and three U3 genes, namely, U3a (GeneBank ID LR782546.1; position: minus strand, 22165827 to 22165610 bp), U3b (TAIR ID: AT5G53902), and U3c (GeneBank ID LR699773.1; position: plus strand, 484549 to 484765 bp) [161]. Studies describing snoRNA genes and their promoters in monocots, including wheat (Triticum aestivum) (TaU6 and TaU3) [164] and rice (OsU3) [165], appeared later.
An avalanche-like increase in plant genome sequencing data [166], together with the first reports on genome editing in several plants species, including Arabidopsis, N. benthamiana, rice, sorghum (Sorghum bicolor), and wheat in 2013 [7], piqued the interest of researchers in the search for species- or family-specific promoters of snoRNA genes to improve genome editing in different plant species. Promoter sequences of snoRNA U6 and U3 genes (from here on abbreviated as pU6 and pU3, respectively) were identified in the genomes of a broad range of plant species, followed by the testing of their applicability for genome editing in the corresponding species. Four pU6 sequences were identified in the cucumber genome [167]; three pU6s were identified in cotton [168]; two pU3s and two pU6s were identified in grapevine (Vitis vinifera) [169]; six pU6s and one pU3 were identified in maize [170]; one pU6 was identified in stiff brome (Brachypodium distachyon) [171]; one pU6 was identified in Douglas fir (Pseudotsuga menziesii), which was used for CRISPR/Cas9 editing in radiate pine (Pinus radiata) [172]; one pU6 was identified in liverwort (Marchantia polymorpha) [173]; and one pU6 was identified in spreading earth moss (Physcomitrium patens) [174], in addition to others. Despite the wide variety of pU6/pU3 sequences described, changing from a widely used pU6/pU3 (e.g., pAtU6, pMtU6, pGmU6, or pOsU3) to a species-specific pU6/pU3 version does not guarantee improved genome editing efficiency, similar to what has been observed regarding the uncertain effects of Cas codon optimization. Nevertheless, pU6/pU3 sequences should be at least class-specific (for monocots or dicots) or division-specific (for Bryophyta, Coniferophyta, or Marchantiophyta).
Recognition of the target DNA by Cas9 is directed by the variable part of the gRNA, the crRNA, which is study-specific. In the simplest approximation, the search for protospacers that can potentially be transcribed into crRNA, proceeds as follows: first, an appropriate PAM should be identified. The canonical PAM comprises the sequence 5′-NGG-3′ in either the sense or antisense DNA strand. An oligonucleotide sequence consisting of 17–20 bases adjacent to the 5′-end of the PAM serves as the protospacer, and it can be included in the CRISPR/Cas vector by fusion without any spacers in front of the 5′-end of the tracrRNA (Figure 2).
Although the 5′-NGG-3′ PAM is widely utilized, it is not the only PAM used for genome editing, but merely the target of a subset of CRISPR/Cas systems. Cas variants that recognize other PAMs exist. Different Cas types have been previously reviewed, such as the SpRY variant that recognizes 5′-NGD-3′ or 5′-NAN-3′ (D: A, G, or T); the XNG-Cas9 variant that specifically recognizes 5′-NG-3′ or 5′-NAN-3′; and the iSpyMacCas9 variant that detects the A-rich PAM 5′-NAA-3′ [103,104,105]. Several Cas nucleases that recognize non-canonical PAMs have been used for genome editing in hairy root systems (see Table S1 [37,77]). The Cpf1 (also known as Cas12) nuclease of Lachnospiraceae bacterium (LbCpf1) and its temperature-tolerant variant (ttLbCpf1), which displays extended target recognition in T-rich PAM sequences (such as 5′-TTTN-3′), were used both for precise ORF editing and the induction of large chromosomal deletions in soybean hairy roots (see Table S1 [37]). Cas9(VQR), recognizing the 5′-NGA-3′ PAM, was successfully utilized for genome editing in the promoter region of the tomato cholesterol biosynthesis gene C5-SD2 (see Table S1 [77]).
The knowledge accumulated on plant genome editing has indicated that, besides an appropriate PAM, several other factors are also important in gRNA design (reviewed in [175,176]). Nucleotides complementary to the crRNA sequence should be present in both DNA strands. The size of the crRNA can vary from 18 to 24 bp (19–20 bp are common). The crRNA sequence should not contain poly(T), which represents the transcription termination signal for RNA polymerase III. At least two crRNAs per target gene should be designed in case one is unsuitable. The crRNA should have a high GC content (50–70%) to increase crRNA-target DNA hybrid stability. The crRNA sequence should not pair with more than six nucleotides of the tracrRNA scaffold. The use of a specific first nucleotide (A for pU3 and G for pU6) after snoRNA gene promoters enhances the expression and stability of the gRNA. If multiple gRNAs are arranged in a single expression vector, the loss and rearrangement of gRNA components can be avoided by using different RNA polymerase III-dependent promoters. Target specificity is determined by the first 10 nucleotides upstream of the PAM in the crRNA sequence [175,176].
The design and testing of gRNAs can be performed both in silico and in planta. The rapid growth of plant genome editing-related data, combined with the development of bioinformatics algorithms, has led to the development of a substantial amount of software that takes into account the factors listed above and helps researchers limit the range of crRNA sequences to be analyzed. Some of these programs have been compared in several reviews [177,178,179]. The WeReview repository for CRISPR bioinformatics tools was created to facilitate the search for and comparison of in silico gRNA design tools [180]. Currently, 104 programs are available for this purpose. Because all the programs are combined in a pivot table on the website, the use of table filters allows researchers to choose the appropriate CRISPR tool [180]. We believe that the most convenient resources for assessing the whole variety of factors affecting editing efficiency are CRISPOR [181] and CRISPR-PLANT v2 [182]. These programs are connected with the greatest diversity of sequenced plant genomes and also evaluate most of the above-mentioned factors that influence crRNA efficiency.
Tools for assessing gRNA effectiveness in planta are also available. Cas9 nuclease activity and the ability of the crRNA to bind to its target DNA sequence can be evaluated on protoplasts before hairy root [183,184] or stable [185,186] transformation. Another attractive technique for in planta gRNA testing is the use of fluorescent protein recovery [187]. Here, the 20–50-bp target DNA sequence chosen for CRISPR/Cas9 genome editing is cloned directly into the ORF of yellow fluorescent protein (YFP), resulting in a frameshift. When leaf epidermis cells are co-bombarded with the resulting YFP vector and the customized CRISPR/Cas9 vector, a DSB is induced at the target site. The imperfect repair of the DSB via NHEJ can restore the YFP ORF, leading to the correct translation of the YFP transcript sequence and, consequently, to YFP fluorescence. To estimate the number of genome editing events that lead to YFP fluorescence recovery, an external control with stable fluorescence, such as an mCherry-expressing vector, can be co-transformed with the YFP and CRISPR/Cas9 vectors. The YFP/mCherry fluorescence ratio is indicative of the Cas9 cleavage activity for the designed gRNA [187]. We believe that the described system could be improved by the assembly of all the components (YFP, mCherry, and CRISPR/Cas9) into a single vector.
Another system based on the green fluorescent protein recovery for detecting genome editing events has recently been developed and tested in planta [188]. The main difference from the above-described system is that the biosensors were developed not only for CRISPR knockout system but also for prime and base editing as well as for CRISPRa. However, use of external control with stable fluorescence together with these biosensors was not reported, which may be a disadvantage of new biosensors, since stable fluorescent control helps to assess the efficiency of transformation.
Nevertheless, these systems have a wide range of applications, although one condition has to be strictly met, as follows: the plant species chosen for the test systems and the species for which the gRNA is designed should belong to different plant families, as the native genome target sequence might interfere with the sequence inserted into the vector with biosensor. Similarly, these systems should be used with care if the selected target DNA region(s) is/are conserved across plant families.
Finally, owing to the short time required for hairy root establishment as compared with the regeneration of stably transformed plants, the former system can be used not only for gene function studies but also for the rapid testing of genome editing efficiency. A test of genome editing efficiency using hairy root transformation before stable transformation has been conducted for rapeseed and soybean (see Table S3 [7,15,21,22,25,27,31,32,40,44,46]). Genome editing efficiency was tested using hairy root transformation before the regeneration of whole-genome-edited plants from individual hairy roots in chicory (Cichorium intybus), the legume M. truncatula, tobacco (Nicotiana tabacum), potato, and rubber dandelion (see Table S3 [9,60,62,79,84]).
3.3. Markers of Transgenicity Used in CRISPR/Cas Vectors: Old Players, New Tricks
The choice of a marker for a CRISPR/Cas vector is important to allow to distinguish between transgenic hairy roots and wild-type roots. There is a great variety of markers, each with its advantages and disadvantages. Since 1983 when the first transgenic plants were obtained [1], methods for the separation of transgenic from non-transgenic material have become well established. Markers for transgenicity of transformed plants can be divided into two large groups, namely, selectable and (visually) screenable markers [9]. The use of selectable markers is based on the principle that plants not carrying the transgene insert will be eliminated following the administration of antibiotics, herbicides, or other phytotoxic compounds, whereas, screenable markers allow the separation of transgenic plants from non-transgenic ones using procedures such as enzymatic reaction-mediated staining of transgenic plant tissues, fluorescence, or pigmentation of transgenic plant parts.
3.3.1. Selectable Markers
Selectable markers were first employed in 1983 (Table S2). The first was the nptII gene encoding neomycin phosphotransferase II (NPTII), which provides resistance against two closely related antibiotics, neomycin and kanamycin [189] (Table S2). Subsequent studies on selectable markers led to the identification of enzymes that could inactivate herbicides (Table S2) or nonantibiotic- or nonherbicide-containing phytotoxic compounds [190,191,192] (Table S2), aiming to avoid the introduction of antibiotic resistance genes into crops and, therefore, into the environment. For instance, some markers allow the selection of transgenic plants for their ability to metabolize endogenous carbohydrates (e.g., xylose, mannose) or for their viability on medium containing high concentrations of non-toxic carbohydrates that are normally detrimental to plants [190,191,192] (Table S2). By 2005, at least 17 selectable markers had been identified (Table S2), a number that has since expanded [190,191,192]. In 2021, two new selectable markers were identified—a new algal gene encoding phosphomannose isomerase, which allows regeneration on 0.6% mannose [193], and the GIBBERELLIC ACID-STIMULATED ARABIDOPSIS 6 (GASA6) gene, which allows regeneration on sugar-free medium [194].
Genes encoding enzymes endowing resistance to antibiotics, herbicides, or other phytotoxic compounds are often used as selectable markers for hairy root transformation. Antibiotic resistance genes were utilized in 20 of the 78 studies examined (Table S1), and the selectable bar herbicide resistance gene in another 24 studies (Table S1). Thus, enzymes providing resistance to antibiotics or herbicides were used as selectable markers in 44 of the evaluated studies (Table S1). However, while these selectable markers are convenient to use, they share one significant disadvantage. Chimeric roots or organs consisting of transgenic and non-transgenic cells or tissues will not be counterselected because non-transformed cells would be protected from the selective agent by the surrounding transformed cells. Chimeric transgenic hairy roots harboring a CRISPR/Cas T-DNA would consist of a mixture of cells with mutated and non-mutated alleles of the target gene, making it impossible to evaluate the effect of the mutation on target gene function [195].
3.3.2. β-Glucuronidase-Based Screenable Markers
A fundamentally new type of screenable marker, β-glucuronidase (GUS) [196] encoded by the uidA gene from Escherichia coli [197], was discovered in 1986 (Table S2). The GUS enzyme converts a soluble, colorless substrate (5-bromo-4-chloro-3-indolyl-β-D-glucuronic acid) into an insoluble, colored product (chloro-bromoindigo), which allowed researchers to visually discriminate transgenic from non-transgenic plants [198]. A GUS fusion system, in which GUS constructs also contained a selectable marker, was initially used to study gene expression [196,199,200]. Consequently, in these first studies, GUS could not be considered a screenable marker as transgenicity in transformed plants was detected based on the selectable marker. The first studies using GUS as a screenable marker were performed on maize in 1988 [201], two years after GUS was first used in plants [196] (Table S2). The GUS construct was delivered by particle bombardment to, and detected in, maize cell suspension culture [201]. In 1989, GUS was also used as the screenable marker for the identification of transgenic hairy roots in the legume Lotus corniculatus [75].
At this point, nine selectable markers were already known and were routinely used for the identification of transgenic plant material (Table S2); however, several of them could not be used for the transformation of some plant species. For instance, in cabbage (Brassica oleracea var. capitata), kanamycin and hygromycin were found to inhibit the growth of both transgenic and non-transgenic hairy roots [202]. Kanamycin was also reported to be toxic for not only the formation of transgenic hairy roots, but also for the regeneration of Rubus sp. plants from transgenic calli [203]. The antibiotics cefotaxime and carbenicillin similarly prevented the regeneration of Rubus sp. plants from calli [203]. Additionally, some plant species chosen for hairy root transformation had natural resistance against antibiotics, as was reported for Brussels sprout (B. oleracea var. gemmifera) [204]. Therefore, the use of GUS as a screenable marker for hairy root transformation became widespread [75,205,206,207]. Chimeric constructs combining GUS with selectable markers were also generated. Such bifunctional markers combined the enzymatic activity of GUS with the activity of antibiotic- or herbicide-inactivating enzymes. For instance, fusions between GUS and NPTII [208] or between GUS and phosphinothricin N-acetyltransferase [209], encoded by the bar gene, have been used for the selection of transgenic plants.
Despite the obvious advantages of using screenable markers instead of selectable markers for the detection of genome-edited hairy roots, GUS staining has not been frequently used for this purpose (it was used only in 6 of the 78 studies examined) when compared with antibiotic/herbicide resistance-related selectable markers (Table S1). However, GUS has disadvantages as a screenable marker for the identification of genome-edited hairy roots. Each root must be split into at least three parts, one for GUS staining to confirm transgenicity, one for the isolation of gDNA to confirm genome editing, and one for the evaluation of morphology or metabolite content. Furthermore, if the transgenic root is chimeric, partial staining may lead to wrong conclusions.
3.3.3. Pigment Biosynthesis-Based Screenable Markers
Another subgroup of screenable markers, discovered after GUS, is based on the staining of transgenic material with natural pigments directly synthesized in the transformed tissue in a process that the transgene regulates or participates in. In 1990, the maize leaf color gene, encoding a protein that regulates the anthocyanin biosynthesis pathway, was proposed as a screenable marker. The activity of this gene leads to the anthocyanin pigmentation of transformed cells in maize kernels [210] (Table S2). The first studies using anthocyanin pigmentation as a screenable marker were performed on monocots [210,211,212] or used genes cloned from monocots for dicot transformation [213,214]. The potential use of anthocyanin pigmentation in hairy root transformation experiments was evaluated in twelve plant species in 1998 [215]. In short, the use of anthocyanin pigmentation as a screenable marker has been used in plant science since the 1990s.
Further evaluation of anthocyanin pigmentation-based screenable markers showed that genes encoding proteins involved in the regulation of anthocyanin biosynthesis [216], including some of the earliest identified genes from maize [217,218,219] (Table S2), belong to two large gene families, one encoding basic helix–loop–helix (bHLH) TFs and the other myeloblastosis (MYB) TFs [216,220]. Several members of the MYB and bHLH gene families from different plant species have also been proposed to serve as screenable markers over the last few years. These include the Arabidopsis PRODUCTION OF ANTHOCYANIN1 (PAP1) gene, encoding the MYB75 TF [221]; sweet potato (Ipomoea batatas) MYB1 gene splice variants a and b [222]; the MYB10 gene from apple (Malus domestica cv. Red Field) [223]; the bHLH gene Delila from snapdragon (Antirrhinum majus) [224]; the MYBA1 gene from grapevine (V. vinifera cv. Merlot) [225]; Legume Anthocyanin Production 1 (LAP1), a member of the MYB gene family of the legume M. truncatula [226]; the rice COLORLESS1 gene (OsC1), an ortholog of maize C1 (ZmC1) encoding ZmMYB1 [227]; and the Anthocyanin2 (AN2), an rubber tree (Hevea brassilensis) ortholog of Arabidopsis PAP1 as well as of grapevine MYB1A [228]. Several of these newly described genes, including MYB75/PAP1 from Arabidopsis [229] and LAP1 from M. truncatula [226,230], have been used as screenable markers for hairy root transformation. Additionally, Arabidopsis MYB75/PAP1 was cloned into a CRISPR/Cas9 vector as a screenable marker for hairy root transformation of soybean (see Table S1 [26]), whereas OsC1 was used for stable transformation of rice by a CRISPR/Cas9 vector [227]. Preliminary tests on the efficiency of MYB75/PAP1 as a marker showed that purple/red staining in soybean transgenic roots was consistent with GUS staining. Purple-stained soybean hairy roots obtained by CRISPR/Cas9-mediated editing of Rfg1 were also efficiently detected [229]. It was also reported, that detection of genome-edited rice regenerants by OsC1 screenable marker was also successful [227]. The diversity of genes regulating anthocyanin biosynthesis [216] leads to a wide choice of potential screenable markers in addition to the six already proposed. Conversely, the diversity of factors involved in the regulation of anthocyanin biosynthesis renders these factors unsuitable as universal screenable markers.
Another candidate for a screenable marker, betalain, was proposed in 2020 [231]. Betalains are red-violet or yellow pigments that accumulate in flowers, fruits, and vegetative tissues of plants in most core families of the order Caryophyllales. If betalains are present in plants, they may replace anthocyanins [232]. Betalain is biosynthesized from tyrosine in a four-step process that involves three genes [231]. The first step is catalyzed by a P450 cytochrome oxygenase, CYP76AD1; the second step is catalyzed by L-DOPA 4,5-dioxygenase (DODA) or, alternatively, CYP76AD1; the third step involves a non-enzymatic, condensation reaction; finally the fourth step is catalyzed by a glucosyltransferase (GT). A construct called RUBY, containing the genes coding for these three proteins combined in a single ORF and separated by the ORF for a short, self-excising peptide, 2A, was proposed as a new screenable marker [231,233]. Arabidopsis and rice plants transformed with the RUBY construct exhibited the red betalain coloration, allowing to distinguish transgenic plants from wild-type plants or calli [231].
Although the idea of an expression system for multiple protein-coding genes based on the use of the 2A self-cleaving peptide seems very attractive, the system does not always function efficiently. Three events are possible during the translation of a transcript containing two ORFs linked by the 2A coding sequence, namely, the independent translation of the two transcripts via a skipping mechanism; the ribosome cannot translate the 2A peptide sequence and is detached from the transcript; and the production of a fusion protein without the 2A peptide [234]. In addition to failures of the 2A self-cleavage system, several other factors can affect the efficiency of the system in plants, including the length of the self-cleaving peptide sequence and the nucleotide composition of the sequences surrounding the 2A sequence. Several scientists believe that the cleavage efficiency should be confirmed by Western blotting [235]. These observations indicate that the use of the RUBY construct can lead to the underestimation of transformation efficiency resulting from failures of the self-cleaving process. We could not find any studies that compared the transformation efficiency of RUBY with that of other constitutively expressed screenable markers (e.g., GUS, fluorescent proteins) in parallel.
Another version of a screenable marker based on the same gene set was proposed in 2021. Here, the transcription of CYP76AD1, DODA, and GT was driven by a constitutive promoter (p35S). N. benthamiana leaves infiltrated with a mixture of Agrobacterium strains, each of which harbored one of three vectors expressing CYP76AD1, DODA, or GT, exhibited betalain-colored spots. In another study, CYP76AD1, DODA, and GT, each under the control of a tomato fruit-specific promoter (pE8), were cloned into a single vector. The system was effective even though the construct contained three copies of the same promoter. In tomato fruits, dark red betalain coloration could be clearly distinguished from their natural red color [236]. The first use of betalain pigmentation as a screenable marker for hairy roots involved the use of a promoter induced in response to arbuscular mycorrhizal (AM) colonization in the so-called MycoRed reporter system [237]. The MycoRed constructs contained three genes in tandem, i.e., CYP76AD1 under the control of the legume M. truncatula AM-responsive promoters of pMtPT4 (PHOSPHATE TRANSPORTER4) or pMtBCP1 (BLUE COPPER PROTEIN1), while expression of the DODA and GT genes was driven by p35S and pUbi10, respectively. The approach allowed in vivo real-time visualization of AM colonization in M. truncatula hairy roots because transgenic root parts colonized by Rhizophagus irregularis turned purple due to the local induction of betalain biosynthesis.
The use of independent CYP76AD1, DODA, and GT constructs under the control of different promoters [236] allows for better control of their expression levels compared with the version containing a single construct with 2A self-cleavage peptide ORFs [231]. Nevertheless, the use of a construct with three separate cassettes also has disadvantages. The three-cassette combination is long when compared with commonly used marker gene constructs; accordingly, the T-DNA transferred by Agrobacterium into the plant genome is also significantly longer, which leads to a reduction in transformation efficiency. It is theoretically possible to generate a construct containing all three betalain synthesis genes and a Cas9 gene, the expression of which would be driven by bidirectional promoters that drive gene expression in both upstream and downstream directions [238,239]. Two constitutive bidirectional promoters could be used in a CRISPR/Cas9 vector also containing the three betalain biosynthesis genes, allowing the expression of four genes under two promoters. The use of bidirectional promoters in CRISPR/Cas9 systems adapted for plants has been previously reported [240].
To conclude the overview of pigment biosynthesis-based screenable markers, we highlight their advantages and restrictions. As advantages, these markers are non-invasive, which represents a clear advantage over the use of GUS-staining technology. They can be visually screened and allow for the detection of chimerism, an option not available when using selectable markers. However, before these markers can be used, it is necessary to determine whether the hairy roots of a given plant species show natural red or purple pigmentation, such as the shikonin-containing hairy roots of the purple gromwell (Lithospermum erythrorhizon) [241,242] and Arnebia hispidissima [243]; the purple pigmented hairy roots of Egyptian henbane (Hyoscyamus muticus) [244]; the hairy roots of the purple coneflower (Echinacea purpurea) when cultured in the light [245]; the betalain-containing hairy roots of red beet (Beta vulgaris) [246,247]; the rutin-containing hairy roots of Tartary buckwheat [248]; and the hairy roots of black carrot (Daucus carota ssp. sativus var. atrorubens) when cultured in the light [249].
3.3.4. Fluorescent Protein-Based Screenable Markers
Fluorescent protein (FP)-based screenable markers were the last group of markers introduced for the detection of transgenic plants/tissue (Table S2). The discovery of the GFP from the jellyfish Aequorea victoria in 1992 [250] changed the world of screenable markers. However, the native gfp gene exhibited low levels of expression and, in plant cells, the encoded protein displayed low fluorescence. To improve gfp, modified versions of the gene were used, such as synthetic gfp (sgfp) [251], modified gfp (mgfp4) [252], and soluble-modified gfp (smgfp) [253]. The first report on the use of GFP as a screenable marker without the application of any additional already known markers (e.g., antibiotic- or herbicide resistance gene, or GUS) occurred in April 1995 [254] (Table S2). Protoplasts of sweet orange (Citrus sinensis) were transformed either with p35S-GFP alone or together with a p35S-GUS construct, with green fluorescence being detected in both cases [254]. The screening of whole Arabidopsis plants by GFP fluorescence and in the absence of a selectable marker was reported two years later [252], while the successful use of GFP for the detection of transgenic hairy roots of the legume L. japonicus was reported only in 2003 [255]. The range of FP-based screenable markers was expanded when a new red FP, DsRed1, was obtained from Discosoma sp. in 1999 [256] (Table S2). DsRed1 was first used as a screenable marker for the identification of transgenic tobacco protoplasts [257] (Table S2), but its application as a screenable marker for transgenic plants was gradually expanded, such as for the detection of transgenic Arabidopsis seeds [258]. The use of DsRed1 as a screenable marker in hairy root transformation experiments was demonstrated for the legumes M. truncatula [159] and soybean [259], as well as for Datisca glomerata, a plant with roots with a thick periderm [260].
FPs are often used as components of bifunctional markers. To expand the applicability of selectable markers, GFP was fused with several proteins, including aminoglycoside-(3″)(9)-adenylyltransferase [261], NPTII [262], blasticidin S deaminase [263], and phosphomannose isomerase [264]. Furthermore, GFP–GUS fusions are frequently used as screenable markers [265].
FP-based screenable markers seem ideal in the context of CRISPR/Cas9 genome editing combined with hairy root transformation. First, they can be used to distinguish chimeric from fully transgenic roots in a non-invasive manner. Second, FP-based fluorescence can be stronger than any intense natural pigmentation of obtained hairy roots, allowing even transgenic hairy roots with natural pigmentation to be easily identified. Finally, FP-based screenable markers have wide applicability in combination with CRISPR/Cas9 vectors. They not only allow the identification of transgenic hairy roots, which can have positive genome editing events, but can also be used to identify transgenic seeds [167,266,267,268,269,270,271] obtained from regenerants of the same hairy roots. The Cas9 nuclease-encoding gene must be removed from the genome of plants regenerated from explants to prevent non-specific genome editing in future generations. Accordingly, seeds obtained from regenerated plants will be selected for the absence of the screenable marker, i.e., only seeds that do not show fluorescence and, therefore, do not have a T-DNA insert with an active Cas gene will be selected.
Notably, the number of available FPs has continued to grow, and the palette is not limited to GFP and DsRed1. Researchers can choose from an appropriate spectrum of FP-based screenable markers using the FP database (FPbase) [272]. Overall, it is difficult to find disadvantages associated with FP-based screenable markers. The one drawback might be that expensive equipment such as a fluorescence stereomicroscope with an appropriate set of filters is required for the detection of fluorescence, which may explain why FP-based screenable markers were only used for genome editing using hairy roots in 23 of the 78 studies examined (Table S1).
3.3.5. New Strategies for the Insertion of a Marker Cassette into a CRISPR/Cas Vector
In addition to the type of marker used, strategies for the insertion of a marker cassette into the CRISPR/Cas vector backbone should also be considered in vector design. Two strategies are currently possible. The first one involves the insertion of a separate cassette consisting of a constitutive promoter, a marker gene, and a transcription terminator into the construct. This strategy has been successfully applied in most CRISPR/Cas9 studies using hairy root transformation (74 of the 78 studies examined) (Table S1). The second strategy involves the fusion of the Cas9 gene with the gene encoding the marker (selectable or screenable). Depending on the sequence between two ORFs, a Cas9 protein can be produced linked to marker protein via a linker, or both proteins can mature independently.
The fusion of Cas9 nuclease with a FP through a linker was used in some of the first genome editing studies in plants [7,273]. A GFP fused via a linker to a Cas9 codon-optimized for humans was used to detect transgenic, and thus genome-edited, N. benthamiana leaf cells. That this resulted in the successful editing of the phytoene desaturase gene [273] suggests that the fusion of GFP with Cas9 did not affect the nuclease activity of the latter. A similar system could be used as a screenable marker for hairy root transformation as the Cas9–FP fusion would be bifunctional, i.e., it could be used both for transgenic root detection and genome editing. However, we did not find any studies using this strategy for hairy roots. The application of this strategy must be precisely tested to ensure that this type of CRISPR/Cas-based system allows the separation of transgenic hairy roots from non-transgenic ones.
The independent maturation of the nuclease and marker protein was proposed by Osakabe et al. first for TALEN [274] and then for Cas9 nuclease [275]. Here, two ORFs are linked by a sequence encoding the self-excising peptide 2A [233]. The resulting chimeric protein is subsequently cleaved into a nuclease (e.g., Cas9) and a marker protein (e.g., FP). Since 2016, the use of this version of the nuclease–marker protein fusion has become widespread for stable plant transformation [91,275,276,277,278,279,280,281,282]. For instance, a Cas9-2A-GFP construct was utilized for genome editing using hairy root transformation (four of the 78 studies examined) in belladonna, potato, the legume L. japonicus, and tomato (see Table S1 [4,5,53,76]). Even though successful CRISPR/Cas9 editing of target genes was reported in these studies, the identification of transgenic hairy roots via GFP fluorescence was reported only for L. japonicus. For belladonna, transgenic roots were identified based on kanamycin resistance, while for tomato and potato, identification was based on direct PCR-based analysis (see Table S1 [4,5,53,76]). These observations suggest that the efficiency of transgenic hairy root detection through FPs derived from the Cas9-2A-FP chimeric protein varies among plant species, presumably due to the imperfection of the 2A self-cleavage, and requires preliminary analysis using an external control with stable fluorescence.
3.3.6. Construction of the CRISPR/Cas9 Vector with the DsRed1 Screenable Marker
CRISPR/Cas9 vectors carrying FP-based screenable markers are widely used both for the positive selection of transgenic hairy roots (Table S1) and the negative selection of seeds obtained from genome-edited plants [167,266,267,268,269,270,271]. One of the most popular CRISPR/Cas9 vector systems used for genome editing both in hairy roots and in stable transformants is pKSE401 and its relatives (e.g., pBSE401, pHSE40), which allow the simultaneous editing of up to four gene targets [101]. At least 7 of the 78 studies used pKSE401 or pHSE401 modified by the inclusion of several screenable markers. pKSE401 was modified either with GFP [167,267] or GUS [283,284,285], while pHSE401 was modified with GFP [167], AtMYB75/PAP1 (resulting in anthocyanin pigmentation) [229], or GUS [286]. However, no reports exist of pKSE401 or its derivatives being modified to include a red fluorescent protein-encoding gene.
We modified pKSE401 [101] with the gene encoding DsRed1 (Figure 4B) because its emission spectrum could be clearly separated from the green autofluorescence present in the roots of our model plant Cucumis sativus (Figure 6C,F). For this purpose, the pAtUbi10-DsRed1-NOS terminator cassette created by Limpens et al. [159] was amplified from the pK7GWIWGII-RedRoot vector [287,288] with minor modifications. The cassette is 2469 bp long and originally consists of 1523 bp of pAtUbi10 (from −1508 to +15 relative to the ATG of the AtUbi10 gene); 681 bp of the DsRed1 ORF; and 265 bp of the NOS terminator. We have previously shown that the shorter version of pAtUbi10 (636 bp; from −637 to −1 relative to the ATG of the AtUbi10 gene) from pCMU-NUCr (AddGene #61168) [289] can also drive the constitutive expression of an FP reporter in transgenic hairy roots [96]. Alignment of the two pAtUbi10 sequences indicated that they were 99.5% identical from position −637 to −1 (relative to the ATG of the AtUbi10 gene). Accordingly, we designed primers that would amplify approximately half of the AtUbi10 promoter from the original pAtUbi10-DsRed1-NOS cassette (652 instead of 1523 bp) and generated a pAtUbi10-DsRed1-NOS cassette containing the modified AtUbi10 promoter (from −637 to +15 relative to the ATG of the AtUbi10 gene). The 1598-bp PCR product (pAtUbi10-DsRed1-NOS terminator) was cloned into pKSE401 containing the zCas9 gene enhanced by the OsMac3 5′-UTR (Figure 4B), resulting in a pKSEe401R vector with the DsRed1 cassette in either tandem or reverse orientation relative to the Cas9 cassette (Figure 4B). The DsRed1 expression cassettes (pAtUbi10-DsRed1-NOS terminator) were sequenced and no nucleotide rearrangements or substitutions were observed compared with the original pAtUbi10-DsRed1-NOS sequence.
The resulting vectors were used to transform cucumber (Cucumis sativus cv. Kustovoy, Sortsemovoshch, St. Petersburg, Russian Federation) using A. rhizogenes strain R1000 as previously described [290]. In total, 54 composite plants with hairy roots were obtained (Figure 6A,D). No DsRed1 fluorescence was observed in 27 composite plants transformed with pKSEe401R containing the DsRed1 cassette in tandem orientation to the Cas9 cassette (Figure 6B). In contrast, DsRed1 fluorescence was detected in each of 27 composite plants with hairy roots harboring the DsRed1 cassette in reverse orientation to the Cas9 cassette (Figure 6E). This indicates that a CRISPR/Cas9 vector with a DsRed1 cassette in reverse orientation can be applied both for the identification of transgenic hairy roots and the selection of seeds from regenerants.
4. Possible Applications of CRISPR/Cas9 Genome Editing Using Hairy Root Transformation
An increasing number of studies have reported on the use of CRISPR/Cas9 genome editing since the first application in tomato hairy roots in 2014 [8] (Tables S1 and S3), with four to six studies per year being published between 2015 and 2019 (Figure 7). Altogether, 78 studies combining CRISPR/Cas9 technology and hairy root transformation were undertaken in 26 plant species (Figure 7; Tables S1 and S3).
Two main approaches were used in these studies. The first involved the detection of CRISPR/Cas9 genome editing events in hairy roots without further phenotyping (Table S3). Some of these studies aimed at testing the efficiency of the CRISPR/Cas9 genome editing approach using hairy root transformation for the different plant species. In another group of studies, phenotyping of edited roots was excluded as the main goal was to obtain whole genome-edited plants regenerated from individual edited hairy roots. In other cases, hairy root transformation was used as a system for the rapid testing of crRNA efficiency; gRNAs with maximal efficiency in the screening system were selected for use in stable transformation (Table S3). The second approach involved both genotyping and phenotyping of genome-edited hairy roots (Table S3). Investigations were divided into the following four subgroups according to the aims: root development or root function; root nodule symbiosis; resistance to biotic or abiotic stresses; or metabolic engineering (Table S3).
5. Conclusions and Perspectives
Successful genome editing using hairy root transformation requires several preconditions. First, the genome sequence of the target plant must be known. A large number of sequenced plant genomes [166,291] and plant genomic databases [292,293] are currently available, and the numbers keep growing [294]. Moreover, the absence of a published genome sequence is now less of an obstacle than previously as genome sequencing and assembly have become much cheaper and faster owing to the development of new sequencing technologies and assembly algorithms [295,296].
The second precondition for successful genome editing is a protocol for high-efficiency transformation by A. rhizogenes strains and the regeneration of hairy roots. To date, more than 100 plant species have been shown to form hairy roots after infection with A. rhizogenes [3,5,10]. While this list is impressive, it means that hairy root transformation protocols have yet to be developed for many plant species. When doing so, attention must be paid to the selection of A. rhizogenes strains, some of which can be highly virulent for certain species; the co-cultivation conditions of agrobacteria with a plant; the stage of plant development; and the type of explant. A well-adapted hairy root transformation protocol can increase the transformation and regeneration efficiency of transgenic roots by up to 70% [297]. Another way to improve the efficiency of a hairy root transformation protocol might be the generation of new supervirulent A. rhizogenes strains either via introducing additional virulence genes [70] or by genome editing [33].
The third precondition refers to the design of an appropriate CRISPR/Cas vector containing cassettes expressing the Cas gene, gRNAs, and transgenicity marker genes. Although different codon-optimized Cas genes together with numerous constitutive, inducible, or tissue-specific promoters have been used in plants, the essential components of a CRISPR/Cas vector, in our opinion, are custom-designed crRNA sequences and a cassette with an efficient selectable or screenable marker of transgenicity, as even codon-optimization of a Cas gene does not guarantee 100% genome editing efficiency. Additionally, whether the use of species-specific snoRNA U3/U6 gene promoters for gRNA expression affects editing efficiency remains controversial [298,299].
The fourth precondition is the proper design of the gRNA specifically of its variable part, the target-specific crRNA. It remains unknown why some crRNAs have higher mutation rates than others; however, the accumulation of experimental data has led to the development of bioinformatics algorithms that allow the assessment of some of the factors influencing the effectiveness of the chosen crRNA. Thus far, there are 104 programs for in silico crRNA design and analysis of its efficiency [180].
The choice of a marker-gene is an important consideration in the design of a CRISPR/Cas vector as the use of an unsuitable marker of transgenicity can lead to the so-called double chimerism problem. The transgenic material might be chimeric both with respect to both the insertion of the artificial T-DNA and the mutations introduced by the CRISPR/Cas system. The first chimerism-related event arises when the marker does not allow to distinguish between completely transgenic roots or calli and chimeric ones that consist of transgenic and non-transgenic tissues. T-DNA chimeras are best avoided by the using screenable markers (e.g., GUS, FPs, anthocyanin or betalain pigmentation). The second event arises because the mutations introduced in the genome by the CRISPR/Cas system are random. Multiple mutations in one or both alleles of the same gene can be introduced in an individual plant or hairy root, and this chimerism can be detected only by sequencing. Examples have been described for both stable [300,301] and hairy root [302,303] transformation.
Not all CRISPR/Cas techniques have been applied in hairy roots. CRISPR/Cas systems with Cas9, Cas9(VQR), LbCpf1, tt-LbCpf1, or nCas9D10A nucleases have only been used to knock out target genes. Other techniques, such as base editing, prime editing, CRISPRa, CRISPRi, epigenome modification, or chromatin imaging, have yet to be successfully combined with hairy root transformation.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/plants11010051/s1: Table S1: List of Agrobacterium strains and CRISPR/Cas vector components from studies using hairy root transformation; Table S2: Examples of the old-known conventional and nonconventional markers used for identification of transgenicity in transformed plants; Table S3: List of research tasks solved using CRISPR/Cas editing of different genes in hairy roots.
Author Contributions
A.S.K. conceived the idea. A.S.K. prepared the original review draft. A.S.K. and K.N.D. developed figures and tables. A.S.K., K.P. and K.N.D. organized figures and tables. Experimental data included in the review were obtained as follows: A.S.K. constructed pKSEe401R and tested it in cucumber hairy roots; E.D.G., E.L.I. and K.N.D. performed the construction of a pRALF34-promoter construct and the promoter localization experiments. A.S.K., E.L.I., K.P. and K.N.D. reviewed and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The Russian Foundation for Basic Research supported the pKSEe401R vector construction experiments (Grant No. 20-316-80046). The Russian Science Foundation supported the cucumber RALF34 identification and promoter localization experiments (Grant No. 20-16-00115).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All relevant data are within the paper and its Supplementary Materials files.
Acknowledgments
We are grateful to Marc van Montagu (Ghent University, Ghent, Belgium) and Csaba Koncz (Max Planck Institute for Plant Breeding Research, Cologne, Germany), for data about the origin of the GV abbreviation in A. tumefaciens strain names; Frank F. White (University of Florida, Gainesville, Florida, USA), for clarification of the process how the A. rhizogenes R1000 strain was isolated; Jens Stougaard (Aarhus University, Aarhus, Denmark), for information about the isolation of A. rhizogenes transconjugant strains C58C1 (pRi15834) rif and AR1193; Inge Broer (University of Rostock, Rostock, Germany), for helpful commentary about the origin of the A. rhizogenes ARqua1 strain name; Chris R. Somerville (University of California, Berkeley, California, USA), for data about the origin of the A. rhizogenes strain MSU440; Paul J.J. Hooykaas (Leiden University, Leiden, Netherlands), for explaining how the A. rhizogenes strain LBA1334 was isolated and for helpful discussions about the available genome assemblies of other A. rhizogenes strains; Tsubasa Shoji (Riken Center for Sustainable Resource Science, Tokyo, Japan) for providing information about selectable marker used for belladonna (Atropa belladonna) transgenic hairy root detection; Sylvestre Marillonnet (Leibniz Institute of Plant Biochemistry, Halle, Germany), for providing information about hairy root transformation of the Madagaskar periwinkle (Catharanthus roseus); Qiyan Jiang (Institute of Crop Sciences, Beijing, China) for providing details about lncRNA77580 genome editing in soybean (Glycine max) hairy roots; Mark Youles (The Sainsbury Laboratory SynBio, Norwich, United Kingdom) and Ruilian Han (Beijing Normal University, Beijing, China), for details concerning the construction of plasmid pICSL002218A; Uwe Ludewig (University of Hohenheim, Stuttgart, Germany), for information about plasmid construction for white lupin hairy root transformation; Vladislav V. Yemelyanov (Saint Petersburg State University, Saint Petersburg, Russian Federation), for providing japonica rice seedlings for gDNA extraction; Erik Limpens (Wageningen University, Wageningen, Netherlands), for providing plasmid pK7GWIWG2(II)-RedRoot; the BioRender Team (www.biorender.com, accessed on 29 November 2021), for the opportunity to create illustrations; and the Charlesworth Author Service Team (www.cwauthors.com, accessed on 29 November 2021) for editing a draft of this manuscript. We are grateful to all researchers whose studies were the basis of this review, and we apologize to researchers whose excellent studies were not cited in this review and its supplementary materials due to space limitations. This study was performed using equipment of the Core Facility of Cell and Molecular Technologies in Plant Science at the Komarov Botanical Institute (Saint Petersburg, Russian Federation) and the Core Facility of Genomic Technologies, Proteomics and Cell Biology at the Research Institute for Agricultural Microbiology (Saint Petersburg, Pushkin, Russian Federation).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| CRISPR | clustered regularly interspaced short palindromic repeats |
| Cas | CRISPR associated nuclease |
| crRNA | crisprRNA |
| tracrRNA | trans-activating crisprRNA |
| PAM | protospacer adjacent motif |
| ORF | open reading frame |
| TF | transcription factor |
| TSS | transcription start site |
References
- Somssich, M. A short history of plant transformation. Peer J. 2019, 7, 1–28. [Google Scholar] [CrossRef]
- Young, J.M.; Kuykendall, L.D.; Martínez-Romero, E.; Kerr, A.; Sawada, H. A revision of Rhizobium Frank 1889, with an emended description of the genus, and the inclusion of all species of Agrobacterium Conn 1942 and Allorhizobium undicola de Lajudie et al. 1998 as new combinations: Rhizobium radiobacter, R. rhizogenes, R. rubi, R. undicola and R. vitis. Int. J. Syst. Evol. Microbiol. 2001, 51, 89–103. [Google Scholar] [CrossRef]
- Porter, J.R.; Flores, H. Host range and implications of plant infection by Agrobacterium rhizogenes. Crit. Rev. Plant. Sci. 1991, 10, 387–421. [Google Scholar] [CrossRef]
- Bahramnejad, B.; Naji, M.; Bose, R.; Jha, S. A critical review on use of Agrobacterium rhizogenes and their associated binary vectors for plant transformation. Biotechnol. Adv. 2019, 37, 1–14. [Google Scholar] [CrossRef]
- Desmet, S.; Dhooghe, E.; De Keyser, E.; Van Huylenbroeck, J.; Müller, R.; Geelen, D.; Lütken, H. Rhizogenic agrobacteria as an innovative tool for plant breeding: Current achievements and limitations. Appl. Microbiol. Biotechnol. 2020, 104, 2435–2451. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, Z.; Iqbal, M.S.; Ahmad, A.; Memon, A.G.; Ansari, M.I. New prospects on the horizon: Genome editing to engineer plants for desirable traits. Curr. Plant Biol. 2020, 24, 1–12. [Google Scholar] [CrossRef]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Nekrasov, V. Plant genome editing made easy: Targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ron, M.; Kajala, K.; Pauluzzi, G.; Wang, D.; Reynoso, M.A.; Zumstein, K.; Garcha, J.; Winte, S.; Masson, H.; Inagaki, S.; et al. Hairy root transformation using Agrobacterium rhizogenes as a tool for exploring cell type-specific gene expression and function using tomato as a model. Plant Physiol. 2014, 166, 455–469. [Google Scholar] [CrossRef]
- Twyman, R.M.; Stöger, E.; Kohli, A.; Capell, T.; Christou, P. Selectable and screenable markers for rice transformation. In Testing for Genetic Manipulation in Plants, 1st ed.; Jackson, J.F., Linskens, H.F., Eds.; Springer: Berlin, Germany, 2002; pp. 1–17. [Google Scholar] [CrossRef]
- Dhiman, N.; Patial, V.; Bhattacharya, A. The current status and future applications of hairy root cultures. In Biotechnological Approaches for Medicinal and Aromatic Plants: Conservation, Genetic Improvement and Utilization, 1st ed.; Kumar, N., Ed.; Springer Science+Business Media LLC: Singapore, 2018; pp. 87–155. [Google Scholar] [CrossRef]
- Alok, A.; Kumar, J.; Upadhyay, S.K. Engineering in hairy roots using CRISPR/Cas9-mediated editing. In Hairy Roots: An Effective Tool of Plant Biotechnology; Srivastava, V., Mehrotra, S., Mishra, S., Eds.; Springer Science+Business Media LLC: Singapore, 2018; pp. 329–342. [Google Scholar] [CrossRef]
- De Saeger, J.; Park, J.; Chung, H.S.; Hernalsteens, J.-P.; Van Lijsebettens, M.; Inzé, D.; Van Montagu, M.; Depuydt, S. Agrobacterium strains and strain improvement: Present and outlook. Biotechnol. Adv. 2021, 107677, 1–25. [Google Scholar] [CrossRef]
- Agrobacterium rhizogenes (ATCC® 15834™). Available online: https://www.atcc.org/products/15834 (accessed on 15 December 2021).
- White, F.F.; Nester, E.W. Hairy root: Plasmid encodes virulence traits in Agrobacterium rhizogenes. J. Bacteriol. 1980, 141, 1134–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, F.F.; Nester, E.W. Relationship of plasmids responsible for hairy root and crown gall tumorigenicity. J. Bacteriol. 1980, 144, 710–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopkins, D.L.; Durbin, R.D. Induction of adventitious roots by culture filtrates of the hairy root bacterium, Agrobacterium rhizogenes. Cannadian J. Microbiol. 1971, 17, 1409–1412. [Google Scholar] [CrossRef]
- Kajala, K.; Coil, D.A.; Brady, S.M. Draft genome sequence of Rhizobium rhizogenes strain ATCC 15834. Genome Announc. 2014, 2, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrobacterium rhizogenes (ATCC® 43057™). Available online: https://www.atcc.org/products/43057 (accessed on 15 December 2021).
- Moore, L.; Warren, G.; Strobel, G. Involvement of a plasmid in the hairy root disease of plants caused by Agrobacterium rhizogenes. Plasmid 1979, 2, 617–626. [Google Scholar] [CrossRef]
- Jouanin, L.; Tourneur, J.; Tourneur, C.; Casse-Delbart, F. Restriction maps and homologies of the three plasmids of Agrobacterium rhizogenes strain A4. Plasmid 1986, 16, 124–134. [Google Scholar] [CrossRef]
- Cho, S.-T.; Lin, Y.-C.; Lai, E.-M.; Kuo, C.-H. Complete Genome Sequence of Agrobacterium rhizogenes A4 (EML492). Available online: https://www.ncbi.nlm.nih.gov/assembly/GCF_018138105.1 (accessed on 15 December 2021).
- White, F.F.; Sinkar, V.P. Molecular analysis of root induction by Agrobacterium rhizogenes. In Plant DNA Infectious Agents, 1st ed.; Hohn, T., Schell, J., Eds.; Springer: Vienna, Austria, 1987; pp. 149–177. [Google Scholar] [CrossRef]
- Alpizar, E.; Dechamp, E.; Espeout, S.; Royer, M.; Lecouls, A.C.; Nicole, M.; Bertrand, B.; Lashermes, P.; Etienne, H. Efficient production of Agrobacterium rhizogenes-transformed roots and composite plants for studying gene expression in coffee roots. Plant Cell. Rep. 2006, 25, 959–967. [Google Scholar] [CrossRef]
- Bonaldi, K.; Gherbi, H.; Franche, C.; Bastien, G.; Fardoux, J.; Barker, D.; Giraud, E.; Cartieaux, F. The Nod factor–independent symbiotic signaling pathway: Development of Agrobacterium rhizogenes–mediated transformation for the legume Aeschynomene indica. Mol. Plant-Microbe Interact. 2010, 23, 1537–1544. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Hu, G.; Dupas, A.; Medina, L.; Blandels, N.; San Clemente, H.; Ladouce, N.; Badawi, M.; Hernandez-Raquet, G.; Mounet, F.; et al. Implementing the CRISPR/Cas9 technology in Eucalyptus hairy roots using wood-related genes. Int. J. Mol. Sci. 2020, 21, 3408. [Google Scholar] [CrossRef] [PubMed]
- Pellegrineschi, A.; Damon, J.-P.; Valtorta, N.; Paillard, N.; Tapfer, D. Improvement of ornamental characters and fragrance production in lemon-scented Geranium through genetic transformation by Agrobacterium rhizogenes. Bio/Technology 1994, 12, 64–68. [Google Scholar] [CrossRef]
- Stiller, J.; Martirani, L.; Tuppale, S.; Chian, R.-J.; Chiurazzi, M.; Gresshoff, P.M. High frequency transformation and regeneration of transgenic plants in the model legume Lotus japonicus. J. Exp. Bot. 1997, 48, 1357–1365. [Google Scholar] [CrossRef] [Green Version]
- NCPPB No. 2659, Rhizobium radiobacter. Available online: http://ncppb.fera.defra.gov.uk/furtherinfo.cfm?ncppb_no=2659 (accessed on 15 December 2021).
- NCPPB No. 1855, Rhizobium rhizogenes. Available online: http://ncppb.fera.defra.gov.uk/furtherinfo.cfm?ncppb_no=1855 (accessed on 15 December 2021).
- Combard, A.; Brevet, J.; Borowski, D.; Cam, K.; Tempé, J. Physical map of the T-DNA region of Agrobacterium rhizogenes strain NCPPB2659. Plasmid 1987, 18, 70–75. [Google Scholar] [CrossRef]
- Mankin, S.L.; Hill, D.S.; Olhoft, P.M.; Toren, E.; Wenck, A.R.; Nea, L.; Xing, L.; Brown, J.A.; Fu, H.; Ireland, L.; et al. Disarming and sequencing of Agrobacterium rhizogenes strain K599 (NCPPB2659) plasmid pRi2659. In Vitro Cell. Dev. Biol. Plant 2007, 43, 521–535. [Google Scholar] [CrossRef]
- Valdes Franco, J.A.; Collier, R.; Wang, Y.; Huo, N.; Gu, Y.; Thilmony, R.; Thomson, J.G. Draft genome sequence of Agrobacterium rhizogenes strain NCPPB2659. Genome Announc. 2016, 4, 1–2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, S.D.; Karimi, M.; Impens, L.; Van Lerberge, E.; Coussens, G.; Aesaert, S.; Rombaut, D.; Holtappels, D.; Ibrahim, H.M.M.; Van Montagu, M.; et al. Efficient CRISPR-mediated base editing in Agrobacterium spp. Proc. Natl. Acad. Sci. USA 2021, 118, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hooykaas, M.J.G.; Hooykaas, P.J.J. The genome sequence of hairy root Rhizobium rhizogenes strain LBA9402: Bioinformatics analysis suggests the presence of a new opine system in the agropine Ri plasmid. MicrobiologyOpen 2021, 10, 1–18. [Google Scholar] [CrossRef]
- Costantino, P.; Hooykaas, P.J.J.; den Dulk-Ras, H.; Schilperoort, R.A. Tumor formation and rhizogenicity of Agrobacterium rhizogenes carrying Ti plasmids. Gene 1980, 11, 79–87. [Google Scholar] [CrossRef]
- Hooykaas, P.J.J.; Klapwijk, P.M.; Nuti, M.P.; Schilperoort, R.A.; Rörsch, A. Transfer of the Agrobacterium tumefaciens Ti plasmid to avirulent Agrobacteria and to Rhizobium ex planta. Microbiology 1977, 98, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Wood, D.W.; Setubal, J.C.; Kaul, R.; Monks, D.E.; Kitajima, J.P.; Okura, V.K.; Zhou, Y.; Chen, L.; Wood, G.E.; Almeida, N.F.; et al. The genome of the natural genetic engineer Agrobacterium tumefaciens C58. Science 2001, 294, 2317–2323. [Google Scholar] [CrossRef] [Green Version]
- Goodner, B.; Hinkle, G.; Gattung, S.; Miller, N.; Blanchard, M.; Qurollo, B.; Goldman, B.S.; Cao, Y.; Askenazi, M.; Halling, C.; et al. Genome sequence of the plant pathogen and biotechnology agent Agrobacterium tumefaciens C58. Science 2001, 294, 2323–2328. [Google Scholar] [CrossRef]
- Slater, S.; Setubal, J.C.; Goodner, B.; Houmiel, K.; Sun, J.; Kaul, R.; Goldman, B.S.; Farrand, S.K.; Almeida, N.; Burr, T.; et al. Reconciliation of sequence data and updated annotation of the genome of Agrobacterium tumefaciens C58, and distribution of a linear chromosome in the genus Agrobacterium. Appl. Environ. Microbiol. 2013, 79, 1414–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, R.H.; Fall, M.Z. The loss of tumor-initiating ability in Agrobacterium tumefaciens by incubation at high temperature. Experientia 1971, 27, 229–230. [Google Scholar] [CrossRef]
- Van Larebeke, N.; Engler, G.; Holsters, M.; Den Elsacker, S.V.; Zaenen, I.; Schilperoort, R.A.; Schell, J. Large plasmid in Agrobacterium tumefaciens essential for crown gall-inducing ability. Nature 1974, 252, 169–170. [Google Scholar] [CrossRef] [PubMed]
- Watson, B.; Currier, T.C.; Gordon, M.P.; Chilton, M.D.; Nester, E.W. Plasmid required for virulence of Agrobacterium tumefaciens. J. Bacteriol. 1975, 123, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhizobium radiobacter (ATCC® 51350™). Available online: https://www.atcc.org/products/51350 (accessed on 15 December 2021).
- Van Larebeke, N.; Genetello, C.H.; Schell, J.; Schilperoort, R.A.; Hermans, A.K.; Hernalsteens, J.P.; Van Montagu, M. Acquisition of tumour-inducing ability by non-oncogenic agrobacteria as a result of plasmid transfer. Nature 1975, 255, 742–743. [Google Scholar] [CrossRef]
- Genetello, C.; Van Larebeke, N.; Holsters, M.; De Picker, A.; Van Montagu, M.; Schell, J. Ti plasmids of Agrobacterium as conjugative plasmids. Nature 1977, 265, 561–563. [Google Scholar] [CrossRef]
- Petit, A.; Tempé, J.; Kerr, A.; Holsters, M.; Van Montagu, M.; Schell, J. Substrate induction of conjugative activity of Agrobacterium tumefaciens Ti plasmids. Nature 1978, 271, 570–572. [Google Scholar] [CrossRef]
- Holsters, M.; Silva, B.; Van Vliet, F.; Genetello, C.; De Block, M.; Dhaese, P.; Depicker, A.; Inzé, D.; Engler, G.; Villarroel, R.; et al. The functional organization of the nopaline A. tumefaciens plasmid pTiC58. Plasmid 1980, 3, 212–230. [Google Scholar] [CrossRef]
- Holsters, M.; de Waele, D.; Depicker, A.; Messens, E.; Van Montagu, M.; Schell, J. Transfection and transformation of Agrobacterium tumefaciens. Mol. Gen. Genet. 1978, 163, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Tempé, J.; Petit, A.; Holsters, M.; Van Montagu, M.; Schell, J. Thermosensitive step associated with transfer of the Ti plasmid during conjugation: Possible relation to transformation in crown gall. Proc. Natl. Acad. Sci. USA 1977, 74, 2848–2849. [Google Scholar] [CrossRef] [Green Version]
- Bomhoff, G.; Klapwijk, P.M.; Kester, H.C.M.; Schilperoort, R.A.; Hernalsteens, J.P.; Schell, J. Octopine and nopaline synthesis and breakdown genetically controlled by a plasmid of Agrobacterium tumefaciens. Mol. Gen. Genet. 1976, 145, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Ledeboer, A.M.; Krol, A.J.M.; Dons, J.J.M.; Spier, F.; Schilperoort, R.A.; Zaenen, I.; Van Larebeke, N.; Schell, J. On the isolation of TI-plasmid from Agrobacterium tumefaciens. Nucleic Acids Res. 1976, 3, 449–464. [Google Scholar] [CrossRef] [Green Version]
- Klapwijk, P.M.; de Jonge, A.J.R.; Schilperoort, R.A.; Rörsch, A. An enrichment technique for auxotrophs of Agrobacterium tumefaciens using a combination of carbenicillin and lysozyme. J. Gen. Microbiol. 1975, 91, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Hooykaas, P.J.J.; den Dulk-Ras, H.; Schilperoort, R.A. Molecular mechanism of Ti plasmid mobilization by R plasmids: Isolation of Ti plasmids with transposon-insertions in Agrobacterium tumefaciens. Plasmid 1980, 4, 64–75. [Google Scholar] [CrossRef]
- Hooykaas, P.J.; den Dulk-Ras, H.; Ooms, G.; Schilperoort, R.A. Interactions between octopine and nopaline plasmids in Agrobacterium tumefaciens. J. Bacteriol. 1980, 143, 1295–1306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fodor, A.; Makrai, L.; Vozik, D.; Olasz, F.; Kiss, J.; Klein, M.G.; Ghaffar, M.-A.B.-A.; Szabados, L.; El-Deen, A.N.; Fodor, L.; et al. Agrobacterium may be used as a suitable experimental system for genetic analysis of resistance to (at least Xenorhabdus budapestensis) antimicrobial peptide complexes. PeerJ 2018, 6, e26900v26901. [Google Scholar] [CrossRef]
- Owens, L.D.; Cress, D.E. Genotypic variability of soybean response to Agrobacterium strains harboring the Ti or Ri plasmids. Plant Physiol. 1985, 77, 87–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, F.F.; Taylor, B.H.; Huffman, G.A.; Gordon, M.P.; Nester, E.W. Molecular and genetic analysis of the transferred DNA regions of the root-inducing plasmid of Agrobacterium rhizogenes. J. Bacteriol. 1985, 164, 33–44. [Google Scholar] [CrossRef] [Green Version]
- McInnes, E.; Davey, M.R.; Mulligan, B.J.; Davies, K.; Sargent, A.W.; Morgan, A.J. Use of a disarmed Ri plasmid vector in analysis of transformed root induction. J. Exp. Bot. 1989, 40, 1135–1144. [Google Scholar] [CrossRef]
- Dommisse, E.M.; Leung, D.W.M.; Shaw, M.L.; Conner, A.J. Onion is a monocotyledonous host for Agrobacterium. Plant Sci. 1990, 69, 249–257. [Google Scholar] [CrossRef]
- Thompson, M.G.; Cruz-Morales, P.; Moore, W.M.; Pearson, A.N.; Keasling, J.D.; Scheller, H.V.; Shih, P.M. Draft genome sequence of Agrobacterium fabrum ARqua1. Microbiol. Resour. Announc. 2020, 9, 1–3. [Google Scholar] [CrossRef]
- Zhou, Y.; Neuhäuser, B.; Neumann, G.; Ludewig, U. LaALMT1 mediates malate release from phosphorus-deficient white lupin root tips and metal root to shoot translocation. Plant Cell. Environ. 2020, 43, 1691–1706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, L.; Jena, S.; Swain, S.S.; Sahoo, S.; Chand, P.K. Agrobacterium rhizogenes-mediated transformation of a multi-medicinal herb, Boerhaavia diffusa L.: Optimization of the process and anti-microbial activity against bacterial pathogens causing urinary tract infections. Front. Life Sci. 2013, 7, 197–209. [Google Scholar] [CrossRef]
- Rhizobium sp. (ATCC® 43056™). Available online: https://www.atcc.org/products/43056 (accessed on 15 December 2021).
- Irigoyen, S.; Ramasamy, M.; Pant, S.; Niraula, P.; Bedre, R.; Gurung, M.; Rossi, D.; Laughlin, C.; Gorman, Z.; Achor, D.; et al. Plant hairy roots enable high throughput identification of antimicrobials against Candidatus Liberibacter spp. Nat. Commun. 2020, 11, 1–14. [Google Scholar] [CrossRef]
- Genome of Rhizobium sp. (ATCC® 43056™). Available online: https://genomes.atcc.org/genomes/e86859d407384d1c?tab=related-genomes-tab (accessed on 15 December 2021).
- Yoshimatsu, K.; Jaziri, M.; Kamada, H.; Shimomura, K. Production of diploid and haploid transgenic Atropa belladonna plants: Morphological traits and tropane alkaloid production. Belg. J. Bot. 1997, 130, 38–46. [Google Scholar]
- Joseph Sahayarayan, J.; Udayakumar, R.; Arun, M.; Ganapathi, A.; Alwahibi, M.S.; Aldosari, N.S.; Morgan, A.M.A. Effect of different Agrobacterium rhizogenes strains for in-vitro hairy root induction, total phenolic, flavonoids contents, antibacterial and antioxidant activity of (Cucumis anguria L.). J. Biol. Sci. 2020, 27, 2972–2979. [Google Scholar] [CrossRef] [PubMed]
- Thwe, A.; Valan Arasu, M.; Li, X.; Park, C.H.; Kim, S.J.; Al-Dhabi, N.A.; Park, S.U. Effect of different Agrobacterium rhizogenes strains on hairy root induction and phenylpropanoid biosynthesis in tartary buckwheat (Fagopyrum tataricum Gaertn). Front. Microbiol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sinkar, V.P.; Pythoud, F.; White, F.F.; Nester, E.W.; Gordon, M.P. rolA locus of the Ri plasmid directs developmental abnormalities in transgenic tobacco plants. Genes Dev. 1988, 2, 688–697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pythoud, F.; Sinkar, V.P.; Nester, E.W.; Gordon, M.P. Increased virulence of Agrobacterium rhizogenes conferred by the vir region of pTiBo542: Application to genetic engineering of poplar. Nat. Biotechnol. 1987, 5, 1323–1327. [Google Scholar] [CrossRef]
- Quandt, H.-J.; Pühler, A.; Broer, I. Transgenic root nodules of Vicia hirsuta: A fast and efficient system for the study of gene expression in indeterminate-type nodules. Mol. Plant-Microbe Interact. 1993, 6, 699–706. [Google Scholar] [CrossRef]
- Sonti, R.V.; Chiurazzi, M.; Wong, D.; Davies, C.S.; Harlow, G.R.; Mount, D.W.; Signer, E.R. Arabidopsis mutants deficient in T-DNA integration. Proc. Natl. Acad. Sci. USA 1995, 92, 11786–11790. [Google Scholar] [CrossRef] [Green Version]
- MSU440 Competent Cells. Available online: https://www.lifeasible.com/?s=MSU440 (accessed on 15 December 2021).
- Petit, A.; David, C.; Dahl, G.A.; Ellis, J.G.; Guyon, P.; Casse-Delbart, F.; Tempé, J. Further extension of the opine concept: Plasmids in Agrobacterium rhizogenes cooperate for opine degradation. Mol. Gen. Genet. 1983, 190, 204–214. [Google Scholar] [CrossRef]
- Hansen, J.; Jørgensen, J.-E.; Stougaard, J.; Marcker, K.A. Hairy roots—A short cut to transgenic root nodules. Plant Cell. Rep. 1989, 8, 12–15. [Google Scholar] [CrossRef]
- Petit, A.; Stougaard, J.; Kühle, A.; Marcker, K.A.; Tempé, J. Transformation and regeneration of the legume Lotus corniculatus: A system for molecular studies of symbiotic nitrogen fixation. Mol. Gen. Genet. 1987, 207, 245–250. [Google Scholar] [CrossRef]
- Stougaard, J.; Abildsten, D.; Marcker, K.A. The Agrobacterium rhizogenes pRi TL-DNA segment as a gene vector system for transformation of plants. Mol. Gen. Genet. 1987, 207, 251–255. [Google Scholar] [CrossRef]
- Bolivar, F.; Rodriguez, R.L.; Greene, P.J.; Betlach, M.C.; Heyneker, H.L.; Boyer, H.W.; Crosa, J.H.; Falkow, S. Construction and characterization of new cloning vehicle. II. A multipurpose cloning system. Gene 1977, 2, 95–113. [Google Scholar] [CrossRef]
- Scitable. Available online: https://www.nature.com/scitable/popular-students-page/149/ (accessed on 15 December 2021).
- Opabode, J.T. Agrobacterium-mediated transformation of plants: Emerging factors that influence efficiency. Biotechnol. Mol. Biol. Rev. 2006, 1, 12–20. [Google Scholar] [CrossRef]
- Vergauwe, A.; Van Geldre, E.; Inzé, D.; Van Montagu, M.; Van den Eeckhout, E. The use of amoxicillin and ticarcillin in combination with a β-lactamase inhibitor as decontaminating agents in the Agrobacterium tumefaciens-mediated transformation of Artemisia annua L. J. Biotechnol. 1996, 52, 89–95. [Google Scholar] [CrossRef]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Nielsen, A.M.; Stougaard, J. Agrobacterium rhizogenes pRi TL-DNA integration system: A gene vector for Lotus japonicus transformation. In Lotus Japonicus Handbook, 1st ed.; Márquez, A.J., Ed.; Springer Science+Business Media LLC: Dordrecht, The Netherlands, 2005; pp. 285–287. [Google Scholar] [CrossRef]
- Díaz, C.L.; Melchers, L.S.; Hooykaas, P.J.J.; Lugtenberg, B.J.J.; Kijne, J.W. Root lectin as a determinant of host–plant specificity in the Rhizobium–legume symbiosis. Nature 1989, 338, 579–581. [Google Scholar] [CrossRef]
- Offringa, I.A.; Melchers, L.S.; Regensburg-Tuink, A.J.G.; Costantino, P.; Schilperoort, R.A.; Hooykaas, P.J.J. Complementation of Agrobacterium tumefaciens tumor-inducing aux mutants by genes from the TR-region of the Ri plasmid of Agrobacterium rhizogenes. Proc. Natl. Acad. Sci. USA 1986, 83, 6935–6939. [Google Scholar] [CrossRef] [Green Version]
- Díaz, C.L.; Grønlund, M.; Schlaman, H.R.M.; Spaink, H.P. Induction of hairy roots for symbiotic gene expression studies. In Lotus Japonicus Handbook, 1st ed.; Márquez, A.J., Ed.; Springer Science+Business Media LLC: Dordrecht, The Netherlands, 2005; pp. 261–277. [Google Scholar] [CrossRef]
- Nonaka, S.; Someya, T.; Kadota, Y.; Nakamura, K.; Ezura, H. Super-Agrobacterium ver. 4: Improving the transformation frequencies and genetic engineering possibilities for crop plants. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Christey, M.C. Use of Ri-mediated transformation for production of transgenic plants. In Vitro Cell. Dev. Biol. Plant 2001, 37, 687–700. [Google Scholar] [CrossRef]
- Amack, S.C.; Antunes, M.S. CaMV35S promoter–a plant biology and biotechnology workhorse in the era of synthetic biology. Curr. Plant Biol. 2020, 24, 1–9. [Google Scholar] [CrossRef]
- Sheen, J. Metabolic repression of transcription in higher plants. Plant Cell 1990, 2, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; McHale, L.K.; Finer, J.J. Isolation and characterization of “GmScream” promoters that regulate highly expressing soybean (Glycine max Merr.) genes. Plant Sci. 2015, 241, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decaestecker, W.; Andrade Buono, R.; Pfeiffer, M.L.; Vangheluwe, N.; Jourquin, J.; Karimi, M.; Van Isterdael, G.; Beeckman, T.; Nowack, M.K.; Jacobs, T.B.B. CRISPR-TSKO: A technique for efficient mutagenesis in specific cell types, tissues, or organs in Arabidopsis. Plant Cell 2019, 31, 2868–2887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Ye, L.; Ursache, R.; Mähönen, A.P. An inducible genome editing system for plants. Nat. Plants 2020, 6, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Omary, M.; Gil-Yarom, N.; Yahav, C.; Steiner, E.; Efroni, I. A conserved superlocus regulates above- and belowground root initiation. bioRxiv 2020. [Google Scholar] [CrossRef]
- De Rybel, B.; Vassileva, V.; Parizot, B.; Demeulenaere, M.; Grunewald, W.; Audenaert, D.; Van Campenhout, J.; Overvoorde, P.; Jansen, L.; Vanneste, S.; et al. A novel Aux/IAA28 signaling cascade activates GATA23-dependent specification of lateral root founder cell identity. Curr. Biol. 2010, 20, 1697–1706. [Google Scholar] [CrossRef]
- Vangheluwe, N.; Beeckman, T. Lateral root initiation and the analysis of gene function using genome editing with CRISPR in Arabidopsis. Genes 2021, 12, 884. [Google Scholar] [CrossRef]
- Ilina, E.L.; Kiryushkin, A.S.; Semenova, V.A.; Demchenko, N.P.; Pawlowski, K.; Demchenko, K.N. Lateral root initiation and formation within the parental root meristem of Cucurbita pepo: Is auxin a key player? Ann. Bot. 2018, 122, 873–888. [Google Scholar] [CrossRef] [Green Version]
- Kiryushkin, A.S.; Ilina, E.L.; Puchkova, V.A.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Lateral root initiation in the parental root meristem of cucurbits: Old players in a new position. Front. Plant Sci. 2019, 10, 1–16. [Google Scholar] [CrossRef]
- Aoki, H.; Teramura, H.; Schepetilnikov, M.; Ryabova, L.A.; Kusano, H.; Shimada, H. Enhanced translation of the downstream ORF attributed to a long 5’ untranslated region in the OsMac1 gene family members, OsMac2 and OsMac3. Plant Biotechnol. 2014, 31, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Onodera, H.; Shingu, S.; Ohnuma, M.; Horie, T.; Kihira, M.; Kusano, H.; Teramura, H.; Shimada, H. Establishment of a conditional TALEN system using the translational enhancer dMac3 and an inducible promoter activated by glucocorticoid treatment to increase the frequency of targeted mutagenesis in plants. PLoS ONE 2018, 13, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Kusano, H.; Ohnuma, M.; Mutsuro-Aoki, H.; Asahi, T.; Ichinosawa, D.; Onodera, H.; Asano, K.; Noda, T.; Horie, T.; Fukumoto, K.; et al. Establishment of a modified CRISPR/Cas9 system with increased mutagenesis frequency using the translational enhancer dMac3 and multiple guide RNAs in potato. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- Xing, H.-L.; Dong, L.; Wang, Z.-P.; Zhang, H.-Y.; Han, C.-Y.; Liu, B.; Wang, X.-C.; Chen, Q.-J. A CRISPR/Cas9 toolkit for multiplex genome editing in plants. BMC Plant Biol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, D.G.; Young, L.; Chuang, R.-Y.; Venter, J.C.; Hutchison, C.A.; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar] [CrossRef]
- Huang, T.-K.; Puchta, H. Novel CRISPR/Cas applications in plants: From prime editing to chromosome engineering. Transgenic Res. 2021, 30, 529–549. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.M.; Zhang, Y.; Yuan, G.; De, K.; Chen, J.-G.; Muchero, W.; Tuskan, G.A.; Qi, Y.; Yang, X. Construct design for CRISPR/Cas-based genome editing in plants. Trends Plant Sci. 2021, 26, 1133–1152. [Google Scholar] [CrossRef] [PubMed]
- Molla, K.A.; Sretenovic, S.; Bansal, K.C.; Qi, Y. Precise plant genome editing using base editors and prime editors. Nat. Plants 2021, 7, 1166–1187. [Google Scholar] [CrossRef]
- Grützner, R.; Martin, P.; Horn, C.; Mortensen, S.; Cram, E.J.; Lee-Parsons, C.W.T.; Stuttmann, J.; Marillonnet, S. High-efficiency genome editing in plants mediated by a Cas9 gene containing multiple introns. Plant Commun. 2021, 2, 1–15. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A.; et al. Multiplex genome engineering using CRISPR/Cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlson, C.K.S.; Mohd-Noor, S.N.; Nolte, N.; Tan, B.C. CRISPR/dCas9-based systems: Mechanisms and applications in plant sciences. Plants 2021, 10, 2055. [Google Scholar] [CrossRef] [PubMed]
- Moore, J.K.; Haber, J.E. Cell cycle and genetic requirements of two pathways of nonhomologous end-joining repair of double-strand breaks in Saccharomyces cerevisiae. Mol. Cell. Biol. 1996, 16, 2164–2173. [Google Scholar] [CrossRef] [Green Version]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef]
- Marzec, M.; Brąszewska-Zalewska, A.; Hensel, G. Prime editing: A new way for genome editing. Trends Cell Biol. 2020, 30, 257–259. [Google Scholar] [CrossRef]
- Marzec, M.; Hensel, G. Prime editing: Game changer for modifying plant genomes. Trends Plant Sci. 2020, 25, 722–724. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef] [Green Version]
- Gaudelli, N.M.; Komor, A.C.; Rees, H.A.; Packer, M.S.; Badran, A.H.; Bryson, D.I.; Liu, D.R. Programmable base editing of A•T to G•C in genomic DNA without DNA cleavage. Nature 2017, 551, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Nishida, K.; Arazoe, T.; Yachie, N.; Banno, S.; Kakimoto, M.; Tabata, M.; Mochizuki, M.; Miyabe, A.; Araki, M.; Hara, K.Y.; et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems. Science 2016, 353, 1–10. [Google Scholar] [CrossRef]
- Kim, Y.B.; Komor, A.C.; Levy, J.M.; Packer, M.S.; Zhao, K.T.; Liu, D.R. Increasing the genome-targeting scope and precision of base editing with engineered Cas9-cytidine deaminase fusions. Nat. Biotechnol. 2017, 35, 371–376. [Google Scholar] [CrossRef]
- Komor, A.C.; Zhao, K.T.; Packer, M.S.; Gaudelli, N.M.; Waterbury, A.L.; Koblan, L.W.; Kim, Y.B.; Badran, A.H.; Liu, D.R. Improved base excision repair inhibition and bacteriophage Mu Gam protein yields C:G-to-T:A base editors with higher efficiency and product purity. Sci. Adv. 2017, 3, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rees, H.A.; Komor, A.C.; Yeh, W.-H.; Caetano-Lopes, J.; Warman, M.; Edge, A.S.B.; Liu, D.R. Improving the DNA specificity and applicability of base editing through protein engineering and protein delivery. Nat. Commun. 2017, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Koblan, L.W.; Doman, J.L.; Wilson, C.; Levy, J.M.; Tay, T.; Newby, G.A.; Maianti, J.P.; Raguram, A.; Liu, D.R. Improving cytidine and adenine base editors by expression optimization and ancestral reconstruction. Nat. Biotechnol. 2018, 36, 843–846. [Google Scholar] [CrossRef]
- Richter, M.F.; Zhao, K.T.; Eton, E.; Lapinaite, A.; Newby, G.A.; Thuronyi, B.W.; Wilson, C.; Koblan, L.W.; Zeng, J.; Bauer, D.E.; et al. Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity. Nat. Biotechnol. 2020, 38, 883–891. [Google Scholar] [CrossRef]
- Gaudelli, N.M.; Lam, D.K.; Rees, H.A.; Solá-Esteves, N.M.; Barrera, L.A.; Born, D.A.; Edwards, A.; Gehrke, J.M.; Lee, S.-J.; Liquori, A.J.; et al. Directed evolution of adenine base editors with increased activity and therapeutic application. Nat. Biotechnol. 2020, 38, 892–900. [Google Scholar] [CrossRef]
- Grünewald, J.; Zhou, R.; Lareau, C.A.; Garcia, S.P.; Iyer, S.; Miller, B.R.; Langner, L.M.; Hsu, J.Y.; Aryee, M.J.; Joung, J.K. A dual-deaminase CRISPR base editor enables concurrent adenine and cytosine editing. Nat. Biotechnol. 2020, 38, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhu, B.; Chen, L.; Xie, L.; Yu, W.; Wang, Y.; Li, L.; Yin, S.; Yang, L.; Hu, H.; et al. Dual base editor catalyzes both cytosine and adenine base conversions in human cells. Nat. Biotechnol. 2020, 38, 856–860. [Google Scholar] [CrossRef] [PubMed]
- Sakata, R.C.; Ishiguro, S.; Mori, H.; Tanaka, M.; Tatsuno, K.; Ueda, H.; Yamamoto, S.; Seki, M.; Masuyama, N.; Nishida, K.; et al. Base editors for simultaneous introduction of C-to-T and A-to-G mutations. Nat. Biotechnol. 2020, 38, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Huang, X.; Wang, X.; Gou, S.; Liang, Y.; Chen, F.; Li, N.; Ouyang, Z.; Zhang, Q.; Ge, W.; et al. ACBE, a new base editor for simultaneous C-to-T and A-to-G substitutions in mammalian systems. BMC Biol. 2020, 18, 1–14. [Google Scholar] [CrossRef]
- Li, C.; Zhang, R.; Meng, X.; Chen, S.; Zong, Y.; Lu, C.; Qiu, J.-L.; Chen, Y.-H.; Li, J.; Gao, C. Targeted, random mutagenesis of plant genes with dual cytosine and adenine base editors. Nat. Biotechnol. 2020, 38, 875–882. [Google Scholar] [CrossRef]
- Xu, R.; Kong, F.; Qin, R.; Li, J.; Liu, X.; Wei, P. Development of an efficient plant dual cytosine and adenine editor. J. Integr. Plant Biol. 2021, 63, 1600–1605. [Google Scholar] [CrossRef]
- Bharat, S.S.; Li, S.; Li, J.; Yan, L.; Xia, L. Base editing in plants: Current status and challenges. Crop J. 2020, 8, 384–395. [Google Scholar] [CrossRef]
- Azameti, M.K.; Dauda, W.P. Base editing in plants: Applications, challenges, and future prospects. Front. Plant Sci. 2021, 12, 1–13. [Google Scholar] [CrossRef]
- Pant, S.R.; McNeece, B.T.; Sharma, K.; Nirula, P.M.; Jiang, J.; Harris, J.L.; Lawrence, G.W.; Klink, V.P. A plant transformation system designed for high throughput genomics in Gossypium hirsutum to study root–organism interactions. J. Plant Interact. 2015, 10, 11–20. [Google Scholar] [CrossRef] [Green Version]
- Sindarovska, Y.R.; Gerasymenko, I.M.; Sheludko, Y.V.; Komarnytskyy, I.K.; Bannikova, M.A.; Kuchuk, N.V. Transgenic plants regenerated from hairy roots of Nicotiana benthamiana: A promising host for transient expression of foreign proteins. Tsitologiya I Genet. 2005, 39, 9–14. [Google Scholar]
- Horn, P.; Santala, J.; Nielsen, S.L.; Hühns, M.; Broer, I.; Valkonen, J.P.T. Composite potato plants with transgenic roots on non-transgenic shoots: A model system for studying gene silencing in roots. Plant Cell Rep. 2014, 33, 1977–1992. [Google Scholar] [CrossRef]
- Menze, A.; Möllers, C. Transformation of Different Brassica napus Cultivars with Three Different Strains of Agrobacterium rhizogenes. In Proceedings of the 10th International Rapeseed Congress, Canberra, Australia, 26–29 September 1999. [Google Scholar]
- Kajikawa, M.; Morikawa, K.; Abe, Y.; Yokota, A.; Akashi, K. Establishment of a transgenic hairy root system in wild and domesticated watermelon (Citrullus lanatus) for studying root vigor under drought. Plant Cell Rep. 2010, 29, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Moradpour, M.; Abdulah, S.N.A. CRISPR/dCas9 platforms in plants: Strategies and applications beyond genome editing. Plant Biotechnol. J. 2020, 18, 32–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zong, Y.; Wang, Y.; Li, C.; Zhang, R.; Chen, K.; Ran, Y.; Qiu, J.-L.; Wang, D.; Gao, C. Precise base editing in rice, wheat and maize with a Cas9-cytidine deaminase fusion. Nat. Biotechnol. 2017, 35, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Kuang, Y.; Ren, B.; Wang, J.; Zhang, D.; Lin, H.; Yang, B.; Zhou, X.; Zhou, H. Highly efficient A·T to G·C base editing by Cas9n-guided tRNA adenosine deaminase in rice. Mol. Plant 2018, 11, 631–634. [Google Scholar] [CrossRef] [Green Version]
- Triezenberg, S.J.; Kingsbury, R.C.; McKnight, S.L. Functional dissection of VP16, the trans-activator of herpes simplex virus immediate early gene expression. Genes Dev. 1988, 2, 718–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beerli, R.R.; Segal, D.J.; Dreier, B.; Barbas, C.F. Toward controlling gene expression at will: Specific regulation of the erbB-2/HER-2 promoter by using polydactyl zinc finger proteins constructed from modular building blocks. Proc. Natl. Acad. Sci. USA 1998, 95, 14628–14633. [Google Scholar] [CrossRef] [Green Version]
- La Russa, M.F.; Qi, L.S. The new state of the art: Cas9 for gene activation and repression. Mol. Cell. Biol. 2015, 35, 3800–3809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiratsu, K.; Ohta, M.; Matsui, K.; Ohme-Takagi, M. The SUPERMAN protein is an active repressor whose carboxy-terminal repression domain is required for the development of normal flowers. FEBS Lett. 2002, 514, 351–354. [Google Scholar] [CrossRef] [Green Version]
- Hiratsu, K.; Mitsuda, N.; Matsui, K.; Ohme-Takagi, M. Identification of the minimal repression domain of SUPERMAN shows that the DLELRL hexapeptide is both necessary and sufficient for repression of transcription in Arabidopsis. Biochem. Biophys. Res. Commun. 2004, 321, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Piatek, A.; Ali, Z.; Baazim, H.; Li, L.; Abulfaraj, A.; Al-Shareef, S.; Aouida, M.; Mahfouz, M.M. RNA-guided transcriptional regulation in planta via synthetic dCas9-based transcription factors. Plant Biotechnol. J. 2015, 13, 578–589. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhang, D.; Baltes, N.J.; Paul, J.W.; Tang, X.; Zheng, X.; Voytas, D.F.; Hsieh, T.-F.; Zhang, Y.; Qi, Y. A CRISPR/Cas9 toolbox for multiplexed plant genome editing and transcriptional regulation. Plant Physiol. 2015, 169, 971–985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Zhang, D.; Xiong, X.; Yan, B.; Xie, W.; Sheen, J.; Li, J.-F. A potent Cas9-derived gene activator for plant and mammalian cells. Nat. Plants 2017, 3, 930–936. [Google Scholar] [CrossRef] [Green Version]
- Lowder, L.G.; Paul, J.W.; Qi, Y. Multiplexed transcriptional activation or repression in plants using CRISPR-dCas9-based systems. In Plant Gene Regulatory Networks: Methods and Protocols, 1st ed.; Kaufmann, K., Mueller-Roeber, B., Eds.; Springer Science+Business Media LLC: New York, NY, USA, 2017; pp. 167–184. [Google Scholar] [CrossRef]
- Lowder, L.G.; Zhou, J.; Zhang, Y.; Malzahn, A.; Zhong, Z.; Hsieh, T.-F.; Voytas, D.F.; Zhang, Y.; Qi, Y. Robust transcriptional activation in plants using multiplexed CRISPR-Act2.0 and mTALE-Act systems. Mol. Plant 2018, 11, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Neumann, M.; Duro, D.I.; Schmid, M. CRISPR-based tools for targeted transcriptional and epigenetic regulation in plants. PLoS ONE 2019, 14, 1–20. [Google Scholar] [CrossRef]
- Li, J.; Blue, R.; Zeitler, B.; Strange, T.L.; Pearl, J.R.; Huizinga, D.H.; Evans, S.; Gregory, P.D.; Urnov, F.D.; Petolino, J.F. Activation domains for controlling plant gene expression using designed transcription factors. Plant Biotechnol. J. 2013, 11, 671–680. [Google Scholar] [CrossRef]
- Ikeda, M.; Ohme-Takagi, M. A novel group of transcriptional repressors in Arabidopsis. Plant Cell. Physiol. 2009, 50, 970–975. [Google Scholar] [CrossRef] [Green Version]
- Selma, S.; Orzáez, D. Perspectives for epigenetic editing in crops. Transgenic Res. 2021, 30, 381–400. [Google Scholar] [CrossRef]
- Dreissig, S.; Schiml, S.; Schindele, P.; Weiss, O.; Rutten, T.; Schubert, V.; Gladilin, E.; Mette, M.F.; Puchta, H.; Houben, A. Live-cell CRISPR imaging in plants reveals dynamic telomere movements. Plant J. 2017, 91, 565–573. [Google Scholar] [CrossRef] [Green Version]
- Fujimoto, S.; Matsunaga, S. Visualization of chromatin loci with transiently expressed CRISPR/Cas9 in plants. Cytologia 2017, 82, 559–562. [Google Scholar] [CrossRef]
- Khosravi, S.; Schindele, P.; Gladilin, E.; Dunemann, F.; Rutten, T.; Puchta, H.; Houben, A. Application of aptamers improves CRISPR-based live imaging of plant telomeres. Front. Plant Sci. 2020, 11, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Konermann, S.; Brigham, M.D.; Trevino, A.E.; Joung, J.; Abudayyeh, O.O.; Barcena, C.; Hsu, P.D.; Habib, N.; Gootenberg, J.S.; Nishimasu, H.; et al. Genome-scale transcriptional activation by an engineered CRISPR-Cas9 complex. Nature 2015, 517, 583–588. [Google Scholar] [CrossRef] [Green Version]
- Tanenbaum, M.E.; Gilbert, L.A.; Qi, L.S.; Weissman, J.S.; Vale, R.D. A protein-tagging system for signal amplification in gene expression and fluorescence imaging. Cell 2014, 159, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Sun, Z.; Su, C.; Wang, Y.; Yan, Q.; Chen, J.; Ott, T.; Li, X. A GmNINa-miR172c-NNC1 regulatory network coordinates the nodulation and autoregulation of nodulation pathways in soybean. Mol. Plant 2019, 12, 1211–1226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, X.; Xie, C.; Ruan, Q.; Zhang, X.; Wu, C.; Han, B.; Qian, J.; Zhou, W.; Nützmann, H.-W.; Kai, G. The transcription factor OpWRKY2 positively regulates the biosynthesis of the anticancer drug camptothecin in Ophiorrhiza pumila. Hortic. Res. 2021, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Limpens, E.; Ramos, J.; Franken, C.; Raz, V.; Compaan, B.; Franssen, H.; Bisseling, T.; Geurts, R. RNA interference in Agrobacterium rhizogenes–transformed roots of Arabidopsis and Medicago truncatula. J. Exp. Bot. 2004, 55, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Waibel, F.; Filipowicz, W. U6 snRNA genes of Arabidopsis are transcribed by RNA polymerase III but contain the same two upstream promoter elements as RNA polymerase ll-transcribed U-snRNA genes. Nucleic Acids Res. 1990, 18, 3451–3458. [Google Scholar] [CrossRef] [PubMed]
- Marshallsay, C.; Kiss, T.; Filipowicz, W. Amplification of plant U3 and U6 snRNA gene sequences using primers specific for an upstream promoter element and conserved intragenic regions. Nucleic Acids Res. 1990, 18, 3459–3466. [Google Scholar] [CrossRef] [Green Version]
- Xie, K.; Minkenberg, B.; Yang, Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system. Proc. Natl. Acad. Sci. USA 2015, 112, 3570–3575. [Google Scholar] [CrossRef] [Green Version]
- Čermák, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.Y.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L.; et al. A multipurpose toolkit to enable advanced genome engineering in plants. Plant Cell 2017, 29, 1196–1217. [Google Scholar] [CrossRef] [Green Version]
- Marshallsay, C.; Connelly, S.; Filipowicz, W. Characterization of the U3 and U6 snRNA genes from wheat: U3 snRNA genes in monocot plants are transcribed by RNA polymerase III. Plant Mol. Biol. 1992, 19, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Qu, F.; Zhai, W.; Chen, H.; Zhu, L.-H.; Morris, T.J. Cloning, characterization and transient expression of the gene encoding a rice U3 small nuclear RNA. Gene 1996, 172, 217–220. [Google Scholar] [CrossRef]
- plaBiPD. Available online: https://www.plabipd.de/plant_genomes_pa.ep (accessed on 15 December 2021).
- Hu, B.; Li, D.; Liu, X.; Qi, J.; Gao, D.; Zhao, S.; Huang, S.; Sun, J.; Yang, L. Engineering non-transgenic gynoecious cucumber using an improved transformation protocol and optimized CRISPR/Cas9 system. Mol. Plant 2017, 10, 1575–1578. [Google Scholar] [CrossRef] [Green Version]
- Long, L.; Guo, D.-D.; Gao, W.; Yang, W.-W.; Hou, L.-P.; Ma, X.-N.; Miao, Y.-C.; Botella, J.R.; Song, C.-P. Optimization of CRISPR/Cas9 genome editing in cotton by improved sgRNA expression. Plant Methods 2018, 14, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 system for genome editing in grape by using grape promoters. Hortic. Res. 2021, 8, 1–12. [Google Scholar] [CrossRef]
- Qi, X.; Dong, L.; Liu, C.; Mao, L.; Liu, F.; Zhang, X.; Cheng, B.; Xie, C. Systematic identification of endogenous RNA polymerase III promoters for efficient RNA guide-based genome editing technologies in maize. Crop. J. 2018, 6, 314–320. [Google Scholar] [CrossRef]
- Alja, S.; Catalin, V.; Jennifer, B.; Karin, L.; John, V.; Markus, P.; Christian, S.H. Broad spectrum developmental role of Brachypodium AUX1. New Phytol. 2018, 219, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Charleson, P.; Lorelle, P.; Barbara, G.; Cathie, R.; Mathias, S.; Glenn, T. Genome editing with CRISPR/Cas9 in Pinus radiata (D. Don). BMC Plant Biol. 2021, 21, 1–9. [Google Scholar] [CrossRef]
- Sugano, S.S.; Shirakawa, M.; Takagi, J.; Matsuda, Y.; Shimada, T.; Hara-Nishimura, I.; Kohchi, T. CRISPR/Cas9-mediated targeted mutagenesis in the liverwort Marchantia polymorpha L. Plant Cell Physiol. 2014, 55, 475–481. [Google Scholar] [CrossRef]
- Collonnier, C.; Epert, A.; Mara, K.; Maclot, F.; Guyon-Debast, A.; Charlot, F.; White, C.; Schaefer, D.G.; Nogué, F. CRISPR-Cas9-mediated efficient directed mutagenesis and RAD51-dependent and RAD51-independent gene targeting in the moss Physcomitrella patens. Plant Biotechnol. J. 2017, 15, 122–131. [Google Scholar] [CrossRef]
- Liang, G.; Zhang, H.; Lou, D.; Yu, D. Selection of highly efficient sgRNAs for CRISPR/Cas9-based plant genome editing. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shan, S.; Soltis, P.S.; Soltis, D.E.; Yang, B. Considerations in adapting CRISPR/Cas9 in nongenetic model plant systems. Appl. Plant Sci. 2020, 8, 1–17. [Google Scholar] [CrossRef]
- Bradford, J.; Perrin, D. A benchmark of computational CRISPR-Cas9 guide design methods. PLOS Comput. Biol. 2019, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Sledzinski, P.; Nowaczyk, M.; Olejniczak, M. Computational tools and resources supporting CRISPR-Cas experiments. Cells 2020, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, Y.; Zhang, T. Computational approaches for effective CRISPR guide RNA design and evaluation. Comput. Struct. Biotechnol. J. 2020, 18, 35–44. [Google Scholar] [CrossRef]
- Torres-Perez, R.; Garcia-Martin, J.A.; Montoliu, L.; Oliveros, J.C.; Pazos, F. WeReview: CRISPR tools—Live repository of computational tools for assisting CRISPR/Cas experiments. Bioengineering 2019, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Haeussler, M.; Schönig, K.; Eckert, H.; Eschstruth, A.; Mianné, J.; Renaud, J.-B.; Schneider-Maunoury, S.; Shkumatava, A.; Teboul, L.; Kent, J.; et al. Evaluation of off-target and on-target scoring algorithms and integration into the guide RNA selection tool CRISPOR. Genome Biol. 2016, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Minkenberg, B.; Zhang, J.; Xie, K.; Yang, Y. CRISPR-PLANT v2: An online resource for highly specific guide RNA spacers based on improved off-target analysis. Plant Biotechnol. J. 2019, 17, 5–8. [Google Scholar] [CrossRef] [Green Version]
- Yuan, M.; Zhu, J.; Gong, L.; He, L.; Lee, C.; Han, S.; Chen, C.; He, G. Mutagenesis of FAD2 genes in peanut with CRISPR/Cas9 based gene editing. BMC Biotechnol. 2019, 19, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, L.; Tan, Q.; Fan, Q.; Zhu, H.; Hong, Z.; Zhang, Z.; Duanmu, D. Efficient inactivation of symbiotic nitrogen fixation related genes in Lotus japonicus using CRISPR/Cas9. Front. Plant Sci. 2016, 7, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Nadakuduti, S.S.; Starker, C.G.; Ko, D.K.; Jayakody, T.B.; Buell, C.R.; Voytas, D.F.; Douches, D.S. Evaluation of methods to assess in vivo activity of engineered genome-editing nucleases in protoplasts. Front. Plant Sci. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Gerasimova, S.V.; Hertig, C.; Korotkova, A.M.; Kolosovskaya, E.V.; Otto, I.; Hiekel, S.; Kochetov, A.V.; Khlestkina, E.K.; Kumlehn, J. Conversion of hulled into naked barley by Cas endonuclease-mediated knockout of the NUD gene. BMC Plant Biol. 2020, 20, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Budhagatapalli, N.; Schedel, S.; Gurushidze, M.; Pencs, S.; Hiekel, S.; Rutten, T.; Kusch, S.; Morbitzer, R.; Lahaye, T.; Panstruga, R.; et al. A simple test for the cleavage activity of customized endonucleases in plants. Plant Methods 2016, 12, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yuan, G.; Hassan, M.M.; Yao, T.; Lu, H.; Vergara, M.M.; Labbé, J.L.; Muchero, W.; Pan, C.; Chen, J.-G.; Tuskan, G.A.; et al. Plant-based biosensors for detecting CRISPR-mediated genome engineering. ACS Synth. Biol. 2021, 10, 3600–3603. [Google Scholar] [CrossRef] [PubMed]
- Herrera-Estrella, L.; De Block, M.; Messens, E.; Hernalsteens, J.-P.; Van Montagu, M.; Schell, J. Chimeric genes as dominant selectable markers in plant cells. EMBO J. 1983, 2, 987–995. [Google Scholar] [CrossRef]
- Sundar, I.K.; Sakthivel, N. Advances in selectable marker genes for plant transformation. J. Plant Physiol. 2008, 165, 1698–1716. [Google Scholar] [CrossRef]
- Rosellini, D. Selectable markers and reporter genes: A well furnished toolbox for plant science and genetic engineering. Crit. Rev. Plant Sci. 2012, 31, 401–453. [Google Scholar] [CrossRef]
- Breyer, D.; Kopertekh, L.; Reheul, D. Alternatives to antibiotic resistance marker genes for in vitro selection of genetically modified plants–scientific developments, current use, operational access and biosafety considerations. Crit. Rev. Plant Sci. 2014, 33, 286–330. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Huang, J. Characterization of an algal phosphomannose isomerase gene and its application as a selectable marker for genetic manipulation of tomato. Plant Divers. 2021, 43, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Gu, J.; Wang, X.; Zhang, S. A novel non-antibiotic selectable marker GASA6 for plant transformation. Plant Cell Tiss. Org. Cult. 2021, 148, 1–12. [Google Scholar] [CrossRef]
- Pena, L.; Cervera, M.; Juárez, J.; Ortega, C.; Pina, J.; Durán-Vila, N.; Navarro, L. High efficiency Agrobacterium-mediated transformation and regeneration of citrus. Plant Sci. 1995, 104, 183–191. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Burgess, S.M.; Hirsh, D. β-Glucuronidase from Escherichia coli as a gene-fusion marker. Proc. Natl. Acad. Sci. USA 1986, 83, 8447–8451. [Google Scholar] [CrossRef] [Green Version]
- Novel, G.; Novel, M. Mutants d’Escherichia coli K 12 affectés pour leur croissance sur méthyl-β-D-glucuronide: Localisation du gène de structure de la β-D-glucuronidase (uid A). Mol. Gen. Genet. 1973, 120, 319–335. [Google Scholar] [CrossRef]
- Cervera, M. Histochemical and fluorometric assays for uidA (GUS) gene detection. In Transgenic Plants: Methods and Protocols, 1st ed.; Peña, L., Ed.; Humana Press: Totowa, NJ, USA, 2004; pp. 203–213. [Google Scholar] [CrossRef]
- Jefferson, R.A. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep. 1987, 5, 387–405. [Google Scholar] [CrossRef]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 6, 3901–3907. [Google Scholar] [CrossRef]
- Klein, T.M.; Gradziel, T.; Fromm, M.E.; Sanford, J.C. Factors influencing gene delivery into Zea mays cells by high–velocity microprojectiles. Bio/Technology 1988, 6, 559–563. [Google Scholar] [CrossRef]
- Berthomieu, P.; Jouanin, L. Transformation of rapid cycling cabbage (Brassica oleracea var. capitata) with Agrobacterium rhizogenes. Plant Cell Rep. 1992, 11, 334–338. [Google Scholar] [CrossRef]
- Graham, J.; McNicol, R.J.; Kumar, A. Use of the GUS gene as a selectable marker for Agrobacterium-mediated transformation of Rubus. Plant Cell Tiss. Org. Cult. 1990, 20, 35–39. [Google Scholar] [CrossRef]
- Hosoki, T.; Kigo, T. Transformation of Brussels sprouts (Brassica oleracea var. gemmifera Zenk.) by Agrobacterium rhizogenes harboring a reporter β-glucuronidase gene. J. Jpn. Soc. Hortic. Sci. 1994, 63, 589–592. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.-G.; Stein, B.; Suzuki, H.; Verma, D.P.S. Expression of antisense nodulin-35 RNA in Vigna aconitifolia transgenic root nodules retards peroxisome development and affects nitrogen availability to the plant. Plant J. 1993, 3, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Boisson-Dernier, A.; Chabaud, M.; Garcia, F.; Bécard, G.; Rosenberg, C.; Barker, D.G. Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol. Plant-Microbe Interact. 2001, 14, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Puddephat, I.J.; Robinson, H.T.; Fenning, T.M.; Barbara, D.J.; Morton, A.; Pink, D.A.C. Recovery of phenotypically normal transgenic plants of Brassica oleracea upon Agrobacterium rhizogenes-mediated co-transformation and selection of transformed hairy roots by GUS assay. Mol. Breed. 2001, 7, 229–242. [Google Scholar] [CrossRef]
- Datla, R.S.S.; Hammerlindl, J.K.; Pelcher, L.E.; Crosby, W.L.; Selvaraj, G. A bifunctional fusion between β-glucuronidase and neomycin phosphotransferase: A broad-spectrum marker enzyme for plants. Gene 1991, 101, 239–246. [Google Scholar] [CrossRef]
- Kamo, K.; McElroy, D.; Chamberiain, D. Transforming embryogenic cell lines of Gladiolus with either a bar-uidA fusion gene or cobombardment. In Vitro Cell. Dev. Biol. Plant 2000, 36, 182–187. [Google Scholar] [CrossRef]
- Ludwig, S.R.; Bowen, B.; Beach, L.; Wessler, S.R. A regulatory gene as a novel visible marker for maize transformation. Science 1990, 247, 449–450. [Google Scholar] [CrossRef]
- Dhir, S.K.; Pajeau, M.E.; Frommn, M.E.; Fry, J.E. Anthocyanin genes as visual markers for wheat transformation. In Improvement of Cereal Quality by Genetic Engineering, 1st ed.; Henry, R.J., Ronalds, J.A., Eds.; Plenum Press: New York, NY, USA, 1994; pp. 71–75. [Google Scholar] [CrossRef]
- Chawla, H.S.; Cass, L.A.; Simmonds, J.A. Expression of anthocyanin pigmentation in wheat tissues transformed with anthocyanin regulatory genes. Curr. Sci. 1999, 76, 1365–1370. [Google Scholar]
- Lloyd, A.M.; Walbot, V.; Davis, R.W. Arabidopsis and Nicotiana anthocyanin production activated by maize regulators R and C1. Science 1992, 258, 1773–1775. [Google Scholar] [CrossRef]
- Quattrocchio, F.; Wing, J.F.; Leppen, H.; Mol, J.; Koes, R.E. Regulatory genes controlling anthocyanin pigmentation are functionally conserved among plant species and have distinct sets of target genes. Plant Cell 1993, 5, 1497–1512. [Google Scholar] [CrossRef]
- Damiani, F.; Paolocci, F.; Consonni, G.; Crea, F.; Tonelli, C.; Arcioni, S. A maize anthocyanin transactivator induces pigmentation in hairy roots of dicotyledonous species. Plant Cell. Rep. 1998, 17, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Khusnutdinov, E.; Sukhareva, A.; Panfilova, M.; Mikhaylova, E. Anthocyanin biosynthesis genes as model genes for genome editing in plants. Int. J. Mol. Sci. 2021, 22, 8752. [Google Scholar] [CrossRef] [PubMed]
- Paz-Ares, J.; Wienand, U.; Peterson, P.A.; Saedler, H. Molecular cloning of the c locus of Zea mays: A locus regulating the anthocyanin pathway. EMBO J. 1986, 5, 829–833. [Google Scholar] [CrossRef]
- Chandler, V.L.; Radicella, J.P.; Robbins, T.P.; Chen, J.; Turks, D. Two regulatory genes of the maize anthocyanin pathway are homologous: Isolation of B utilizing R genomic sequences. Plant Cell 1989, 1, 1175–1183. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, S.R.; Habera, L.F.; Dellaporta, S.L.; Wessler, S.R. Lc, a member of the maize R gene family responsible for tissue-specific anthocyanin production, encodes a protein similar to transcriptional activators and contains the myc-homology region. Proc. Natl. Acad. Sci. USA 1989, 86, 7092–7096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feller, A.; Machemer, K.; Braun, E.L.; Grotewold, E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011, 66, 94–116. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-L.; Zeng, H.-N.; Shi, M.-Z.; Xie, D.-Y. Development of tobacco callus cultures over expressing Arabidopsis PAP1/MYB75 transcription factor and characterization of anthocyanin biosynthesis. Planta 2008, 229, 37–51. [Google Scholar] [CrossRef]
- Kim, C.Y.; Ahn, Y.O.; Kim, S.H.; Kim, Y.-H.; Lee, H.-S.; Catanach, A.S.; Jacobs, J.M.E.; Conner, A.J.; Kwak, S.-S. The sweet potato IbMYB1 gene as a potential visible marker for sweet potato intragenic vector system. Physiol. Plant. 2010, 139, 229–240. [Google Scholar] [CrossRef]
- Kortstee, A.J.; Khan, S.A.; Helderman, C.; Trindade, L.M.; Wu, Y.; Visser, R.G.F.; Brendolise, C.; Allan, A.; Schouten, H.J.; Jacobsen, E. Anthocyanin production as a potential visual selection marker during plant transformation. Transgenic Res. 2011, 20, 1253–1264. [Google Scholar] [CrossRef] [Green Version]
- Naing, A.H.; Lim, K.B.; Kim, C.K. The usage of snapdragon Delila (Del) gene as a visible selection marker for the antibiotic-free transformation system. J. Plant Biol. 2015, 58, 110–116. [Google Scholar] [CrossRef]
- Kandel, R.; Bergey, D.R.; Dutt, M.; Sitther, V.; Li, Z.T.; Gray, D.J.; Dhekney, S.A. Evaluation of a grapevine-derived reporter gene system for precision breeding of Vitis. Plant Cell. Tiss. Org. Cult. 2016, 124, 599–609. [Google Scholar] [CrossRef]
- Zhang, S.; Kondorosi, É.; Kereszt, A. An anthocyanin marker for direct visualization of plant transformation and its use to study nitrogen-fixing nodule development. J. Plant Res. 2019, 132, 695–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Zhu, M.; Wu, J.; Ouyang, L.; Wang, R.; Sun, H.; Yan, L.; Wang, L.; Xu, M.; Zhan, H.; et al. Repurposing of anthocyanin biosynthesis for plant transformation and genome editing. Front. Plant Sci. 2020, 2, 1–10. [Google Scholar] [CrossRef]
- Huang, T.; Xin, S.; Fang, Y.; Chen, T.; Chang, J.; Ko, N.C.K.; Huang, H.; Hua, Y. Use of a novel R2R3-MYB transcriptional activator of anthocyanin biosynthesis as visual selection marker for rubber tree (Hevea brasiliensis) transformation. Ind. Crops Prod. 2021, 174, 1–11. [Google Scholar] [CrossRef]
- Fan, Y.; Wang, X.; Li, H.; Liu, S.; Jin, L.; Lyu, Y.; Shi, M.; Liu, S.; Yang, X.; Lyu, S. Anthocyanin, a novel and user-friendly reporter for convenient, non-destructive, low cost, directly visual selection of transgenic hairy roots in the study of rhizobia-legume symbiosis. Plant Methods 2020, 16, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.; Chen, K.; Su, Y.; Jiang, S.; Xu, P.; Murray, J.D. A root tip-specific expressing anthocyanin marker for direct identification of transgenic tissues by the naked eye in symbiotic studies. Plants 2021, 10, 605. [Google Scholar] [CrossRef]
- He, Y.; Zhang, T.; Sun, H.; Zhan, H.; Zhao, Y. A reporter for noninvasively monitoring gene expression and plant transformation. Hortic. Res. 2020, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sunnadeniya, R.; Bean, A.; Brown, M.; Akhavan, N.; Hatlestad, G.; Gonzalez, A.; Symonds, V.V.; Lloyd, A. Tyrosine hydroxylation in betalain pigment biosynthesis is performed by cytochrome P450 enzymes in beets (Beta vulgaris). PLoS ONE 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halpin, C.; Cooke, S.E.; Barakate, A.; Amrani, A.E.; Ryan, M.D. Self-processing 2A-polyproteins–a system for co-ordinate expression of multiple proteins in transgenic plants. Plant J. 1999, 17, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, O.; Wall, J.B.J.; Zheng, M.; Zhou, Y.; Wang, L.; Ruth Vaseghi, H.; Qian, L.; Liu, J. Systematic comparison of 2A peptides for cloning multi-genes in a polycistronic vector. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef]
- Khosla, A.; Rodriguez-Furlan, C.; Kapoor, S.; Van Norman, J.M.; Nelson, D.C. A series of dual-reporter vectors for ratiometric analysis of protein abundance in plants. Plant Direct 2020, 4, 1–14. [Google Scholar] [CrossRef]
- Grützner, R.; Schubert, R.; Horn, C.; Yang, C.; Vogt, T.; Marillonnet, S. Engineering betalain biosynthesis in tomato for high level betanin production in fruits. Front. Plant Sci. 2021, 12, 1–17. [Google Scholar] [CrossRef]
- Timoneda, A.; Yunusov, T.; Quan, C.; Gavrin, A.; Brockington, S.F.; Schornack, S. MycoRed: Betalain pigments enable in vivo real-time visualisation of arbuscular mycorrhizal colonisation. PLOS Biol. 2021, 19, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; He, Y.; Gan, S. Bidirectionalization of polar promoters in plants. Nat. Biotechnol. 2001, 19, 677–679. [Google Scholar] [CrossRef]
- Dey, N.; Sarkar, S.; Acharya, S.; Maiti, I.B. Synthetic promoters in planta. Planta 2015, 242, 1077–1094. [Google Scholar] [CrossRef]
- Ren, Q.; Zhong, Z.; Wang, Y.; You, Q.; Li, Q.; Yuan, M.; He, Y.; Qi, C.; Tang, X.; Zheng, X.; et al. Bidirectional promoter-based CRISPR/Cas9 systems for plant genome editing. Front. Plant Sci. 2019, 10, 1–11. [Google Scholar] [CrossRef]
- Shimomura, K.; Sudo, H.; Saga, H.; Kamada, H. Shikonin production and secretion by hairy root cultures of Lithospermum erythrorhizon. Plant Cell. Rep. 1991, 10, 282–285. [Google Scholar] [CrossRef]
- Brigham, L.A.; Michaels, P.J.; Flores, H.E. Cell-specific production and antimicrobial activity of naphthoquinones in roots of Lithospermum erythrorhizon. Plant Physiol. 1999, 119, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Pal, M. Induction of shikonin production in hairy root cultures of Arnebia hispidissima via Agrobacterium rhizogenes-mediated genetic transformation. J. Crop Sci. Biotechnol. 2010, 13, 99–106. [Google Scholar] [CrossRef]
- Moyano, E.; Jouhikainen, K.; Tammela, P.; Palazón, J.; Cusidó, R.M.; Piñol, M.T.; Teeri, T.H.; Oksman–Caldentey, K.M. Effect of pmt gene overexpression on tropane alkaloid production in transformed root cultures of Datura metel and Hyoscyamus muticus. J. Exp. Bot. 2003, 54, 203–211. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Tian, C.-L.; Murch, S.J.; Saxena, P.K.; Liu, C.-Z. Light-enhanced caffeic acid derivatives biosynthesis in hairy root cultures of Echinacea purpurea. Plant Cell. Rep. 2007, 26, 1367–1372. [Google Scholar] [CrossRef]
- Thimmaraju, R.; Venkatachalam, L.; Bhagyalakshmi, N. Morphometric and biochemical characterization of red beet (Beta vulgaris L.) hairy roots obtained after single and double transformations. Plant Cell. Rep. 2008, 27, 1039–1052. [Google Scholar] [CrossRef] [PubMed]
- Neelwarne, B. Red beet hairy root cultures. In Red Beet Biotechnology: Food and Pharmaceutical Applications, 1st ed.; Neelwarne, B., Ed.; Springer Science+Business Media LLC: Boston, MA, USA, 2013; pp. 199–249. [Google Scholar] [CrossRef]
- Thwe, A.A.; Kim, J.K.; Li, X.; Bok Kim, Y.; Romij Uddin, M.; Kim, S.J.; Suzuki, T.; Park, N.I.; Park, S.U. Metabolomic analysis and phenylpropanoid biosynthesis in hairy root culture of tartary buckwheat cultivars. PLoS ONE 2013, 8, 1–13. [Google Scholar] [CrossRef]
- Barba-Espín, G.; Chen, S.-T.; Agnolet, S.; Hegelund, J.N.; Stanstrup, J.; Christensen, J.H.; Müller, R.; Lütken, H. Ethephon-induced changes in antioxidants and phenolic compounds in anthocyanin-producing black carrot hairy root cultures. J. Exp. Bot. 2020, 71, 7030–7045. [Google Scholar] [CrossRef] [PubMed]
- Prasher, D.C.; Eckenrode, V.K.; Ward, W.W.; Prendergast, F.G.; Cormier, M.J. Primary structure of the Aequorea victoria green-fluorescent protein. Gene 1992, 111, 229–233. [Google Scholar] [CrossRef]
- Chiu, W.-l.; Niwa, Y.; Zeng, W.; Hirano, T.; Kobayashi, H.; Sheen, J. Engineered GFP as a vital reporter in plants. Curr. Biol. 1996, 6, 325–330. [Google Scholar] [CrossRef] [Green Version]
- Haseloff, J.; Siemering, K.R.; Prasher, D.C.; Hodge, S. Removal of a cryptic intron and subcellular localization of green fluorescent protein are required to mark transgenic Arabidopsis plants brightly. Proc. Natl. Acad. Sci. USA 1997, 94, 2122–2127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, S.J.; Vierstra, R.D. Soluble, highly fluorescent variants of green fluorescent protein (GFP) for use in higher plants. Plant Mol. Biol. 1998, 36, 521–528. [Google Scholar] [CrossRef]
- Niedz, R.P.; Sussman, M.R.; Satterlee, J.S. Green fluorescent protein: An in vivo reporter of plant gene expression. Plant Cell Rep. 1995, 14, 403–406. [Google Scholar] [CrossRef]
- Kumagai, H.; Kouchi, H. Gene silencing by expression of hairpin RNA in Lotus japonicus roots and root nodules. Mol. Plant-Microbe Interact. 2003, 16, 663–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matz, M.V.; Fradkov, A.F.; Labas, Y.A.; Savitsky, A.P.; Zaraisky, A.G.; Markelov, M.L.; Lukyanov, S.A. Fluorescent proteins from nonbioluminescent Anthozoa species. Nat. Biotechnol. 1999, 17, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Jach, G.; Binot, E.; Frings, S.; Luxa, K.; Schell, J. Use of red fluorescent protein from Discosoma sp. (dsRED) as a reporter for plant gene expression. Plant J. 2001, 28, 483–491. [Google Scholar] [CrossRef] [Green Version]
- Stuitje, A.R.; Verbree, E.C.; Van Der Linden, K.H.; Mietkiewska, E.M.; Nap, J.-P.; Kneppers, T.J.A. Seed-expressed fluorescent proteins as versatile tools for easy (co)transformation and high-throughput functional genomics in Arabidopsis. Plant Biotechnol. J. 2003, 1, 301–309. [Google Scholar] [CrossRef]
- Lin, M.-H.; Gresshoff, P.M.; Indrasumunar, A.; Ferguson, B.J. pHairyRed: A novel binary vector containing the DsRed2 reporter gene for visual selection of transgenic hairy roots. Mol. Plant 2011, 4, 537–545. [Google Scholar] [CrossRef]
- Markmann, K.; Giczey, G.; Parniske, M. Functional adaptation of a plant receptor- kinase paved the way for the evolution of intracellular root symbioses with bacteria. PLoS Biol. 2008, 6, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.S.; Maliga, P. Fluorescent antibiotic resistance marker for tracking plastid transformation in higher plants. Nat. Biotechnol. 1999, 17, 910–915. [Google Scholar] [CrossRef]
- Li, Z.; Jayasankar, S.; Gray, D.J. Expression of a bifunctional green fluorescent protein (GFP) fusion marker under the control of three constitutive promoters and enhanced derivatives in transgenic grape (Vitis vinifera). Plant Sci. 2001, 160, 877–887. [Google Scholar] [CrossRef]
- Ochiai-Fukuda, T.; Takahashi-Ando, N.; Ohsato, S.; Igawa, T.; Kadokura, K.; Hamamoto, H.; Nakasako, M.; Kudo, T.; Shibata, T.; Yamaguchi, I.; et al. A fluorescent antibiotic resistance marker for rapid production of transgenic rice plants. J. Biotechnol. 2006, 122, 521–527. [Google Scholar] [CrossRef]
- Dutt, M.; Lee, D.H.; Grosser, J.W. Bifunctional selection–reporter systems for genetic transformation of citrus: Mannose- and kanamycin-based systems. In Vitro Cell. Dev. Biol. Plant 2010, 46, 467–476. [Google Scholar] [CrossRef]
- Quaedvlieg, N.E.M.; Schlaman, H.R.M.; Admiraal, P.C.; Wijting, S.E.; Stougaard, J.; Spaink, H.P. Fusions between green fluorescent protein and β-glucuronidase as sensitive and vital bifunctional reporters in plants. Plant Mol. Biol. 1998, 38, 861–873. [Google Scholar] [CrossRef]
- Gao, X.; Chen, J.; Dai, X.; Zhang, D.; Zhao, Y. An effective strategy for reliably isolating heritable and Cas9-free Arabidopsis mutants generated by CRISPR/Cas9-mediated genome editing. Plant Physiol. 2016, 171, 1794–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, T.; Yu, X.; Yang, H.; Gao, Q.; Ji, H.; Wang, Y.; Yan, G.; Peng, Y.; Luo, H.; Liu, K.; et al. Development and validation of an effective CRISPR/Cas9 vector for efficiently isolating positive transformants and transgene-free mutants in a wide range of plant species. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Aliaga, F.N.; Zhang, C.; Presa, S.; Granell, A.; Alabadi, D.; Sadanandom, A.; Blazquez, M.A.; Minguet, E.G. Identification of transgene-free CRISPR edited plants of rice, tomato and Arabidopsis by monitoring DsRED fluorescence in dry seeds. Front. Plant Sci. 2019, 10, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Okada, A.; Arndell, T.; Borisjuk, N.; Sharma, N.; Watson-Haigh, N.S.; Tucker, E.J.; Baumann, U.; Langridge, P.; Whitford, R. CRISPR/Cas9-mediated knockout of Ms1 enables the rapid generation of male-sterile hexaploid wheat lines for use in hybrid seed production. Plant Biotechnol. J. 2019, 17, 1905–1913. [Google Scholar] [CrossRef] [Green Version]
- Ursache, R.; Fujita, S.; Tendon, V.D.; Geldner, N. Combined fluorescent seed selection and multiplex CRISPR/Cas9 assembly for fast generation of multiple Arabidopsis mutants. Plant Methods 2021, 17, 1–14. [Google Scholar] [CrossRef]
- Yan, Y.; Zhu, J.; Qi, X.; Cheng, B.; Liu, C.; Xie, C. Establishment of an efficient seed fluorescence reporter-assisted CRISPR/Cas9 gene editing in maize. J. Integr. Plant Biol. 2021, 63, 1671–1680. [Google Scholar] [CrossRef] [PubMed]
- Lambert, T.J. FPbase: A community-editable fluorescent protein database. Nat. Methods 2019, 16, 277–278. [Google Scholar] [CrossRef] [PubMed]
- Nekrasov, V.; Staskawicz, B.; Weigel, D.; Jones, J.D.G.; Kamoun, S. Targeted mutagenesis in the model plant Nicotiana benthamiana using Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 691–693. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, K.; Osakabe, Y.; Toki, S. Site-directed mutagenesis in Arabidopsis using custom-designed zinc finger nucleases. Proc. Natl. Acad. Sci. USA 2010, 107, 12034–12039. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Watanabe, T.; Sugano, S.S.; Ueta, R.; Ishihara, R.; Shinozaki, K.; Osakabe, K. Optimization of CRISPR-Cas9 genome editing to modify abiotic stress responses in plants. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ueta, R.; Abe, C.; Watanabe, T.; Sugano, S.S.; Ishihara, R.; Ezura, H.; Osakabe, Y.; Osakabe, K. Rapid breeding of parthenocarpic tomato plants using CRISPR/Cas9. Sci. Rep. 2017, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nishitani, C.; Hirai, N.; Komori, S.; Wada, M.; Okada, K.; Osakabe, K.; Yamamoto, T.; Osakabe, Y. Efficient genome editing in apple using a CRISPR/Cas9 system. Sci. Rep. 2016, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Osakabe, Y.; Liang, Z.; Ren, C.; Nishitani, C.; Osakabe, K.; Wada, M.; Komori, S.; Malnoy, M.; Velasco, R.; Poli, M.; et al. CRISPR–Cas9-mediated genome editing in apple and grapevine. Nat. Protoc. 2018, 13, 2844–2863. [Google Scholar] [CrossRef]
- Hashimoto, R.; Ueta, R.; Abe, C.; Osakabe, Y.; Osakabe, K. Efficient multiplex genome editing induces precise, and self-ligated type mutations in tomato plants. Front. Plant Sci. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F.; Suzuki, T.; Osakabe, Y.; Betsuyaku, S.; Kondo, Y.; Dohmae, N.; Fukuda, H.; Yamaguchi-Shinozaki, K.; Shinozaki, K. A small peptide modulates stomatal control via abscisic acid in long-distance signalling. Nature 2018, 556, 235–238. [Google Scholar] [CrossRef]
- Petersen, B.L.; Möller, S.R.; Mravec, J.; Jørgensen, B.; Christensen, M.; Liu, Y.; Wandall, H.H.; Bennett, E.P.; Yang, Z. Improved CRISPR/Cas9 gene editing by fluorescence activated cell sorting of green fluorescence protein tagged protoplasts. BMC Biotechnol. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hua, L.; Gupta, A.; Tricoli, D.; Edwards, K.J.; Yang, B.; Li, W. Development of an Agrobacterium-delivered CRISPR/Cas9 system for wheat genome editing. Plant Biotechnol. J. 2019, 17, 1623–1635. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Yang, S.; Liu, J.; Terecskei, K.; Ábrahám, E.; Gombár, A.; Domonkos, Á.; Szűcs, A.; Körmöczi, P.; Wang, T.; et al. Host-secreted antimicrobial peptide enforces symbiotic selectivity in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2017, 114, 6854–6859. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.; Wang, Q.; Fedorova, E.; Liu, J.; Qin, Q.; Zheng, Q.; Price, P.A.; Pan, H.; Wang, D.; Griffitts, J.S.; et al. Microsymbiont discrimination mediated by a host-secreted peptide in Medicago truncatula. Proc. Natl. Acad. Sci. USA 2017, 114, 6848–6853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Liu, J.; Li, H.; Yang, S.; Körmöczi, P.; Kereszt, A.; Zhu, H. Nodule-specific cysteine-rich peptides negatively regulate nitrogen-fixing symbiosis in a strain-specific manner in Medicago truncatula. Mol. Plant-Microbe Interact. 2018, 31, 240–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Yang, S.; Liu, J.; Zhu, H. Rj4, a gene controlling nodulation specificity in soybeans, encodes a thaumatin-like protein but not the one previously reported. Plant Physiol. 2016, 170, 26–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY™ vectors for Agrobacterium-mediated plant transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Limpens, E.; Mirabella, R.; Fedorova, E.; Franken, C.; Franssen, H.; Bisseling, T.; Geurts, R. Formation of organelle-like N2-fixing symbiosomes in legume root nodules is controlled by DMI2. Proc. Natl. Acad. Sci. USA 2005, 102, 10375–10380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, S.; Harrison, M.J. A set of fluorescent protein-based markers expressed from constitutive and arbuscular mycorrhiza-inducible promoters to label organelles, membranes and cytoskeletal elements in Medicago truncatula. Plant J. 2014, 80, 1151–1163. [Google Scholar] [CrossRef]
- Ilina, E.L.; Logachov, A.A.; Laplaze, L.; Demchenko, N.P.; Pawlowski, K.; Demchenko, K.N. Composite Cucurbita pepo plants with transgenic roots as a tool to study root development. Ann. Bot. 2012, 110, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Song, Y.; Li, X.; Chen, J.; Mo, L.; Zhang, X.; Lin, Z.; Zhang, L. Genome sequences of horticultural plants: Past, present, and future. Hortic. Res. 2019, 6, 11–23. [Google Scholar] [CrossRef] [Green Version]
- van Dijk, A.D.J. Plant Genomics Databases. Methods and Protocols, 1st ed.; Springer Science+Business Media LLC: New York, NY, USA, 2017; pp. 1–336. [Google Scholar]
- Chen, F.; Dong, W.; Zhang, J.; Guo, X.; Chen, J.; Wang, Z.; Lin, Z.; Tang, H.; Zhang, L. The sequenced angiosperm genomes and genome databases. Front. Plant Sci. 2018, 9, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Shang, L.; Zhu, Q.-H.; Fan, L.; Guo, L. Twenty years of plant genome sequencing: Achievements and challenges. Trends Plant Sci. 2021, 2207, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-W.; Harkess, A. A guide to sequence your favorite plant genomes. Appl. Plant Sci. 2018, 6, 1–7. [Google Scholar] [CrossRef]
- Paajanen, P.; Kettleborough, G.; López-Girona, E.; Giolai, M.; Heavens, D.; Baker, D.; Lister, A.; Cugliandolo, F.; Wilde, G.; Hein, I.; et al. A critical comparison of technologies for a plant genome sequencing project. GigaScience 2019, 8, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Wang, X.; Cao, L.; Ji, J.; Liu, T.; Duan, K. Highly efficient Agrobacterium rhizogenes-mediated hairy root transformation for gene functional and gene editing analysis in soybean. Plant Methods 2021, 17, 1–12. [Google Scholar] [CrossRef]
- Cai, Y.; Chen, L.; Liu, X.; Sun, S.; Wu, C.; Jiang, B.; Han, T.; Hou, W. CRISPR/Cas9-mediated genome editing in soybean hairy roots. PLoS ONE 2015, 10, 1–13. [Google Scholar] [CrossRef]
- Du, H.; Zeng, X.; Zhao, M.; Cui, X.; Wang, Q.; Yang, H.; Cheng, H.; Yu, D. Efficient targeted mutagenesis in soybean by TALENs and CRISPR/Cas9. J. Biotechnol. 2016, 217, 90–97. [Google Scholar] [CrossRef]
- Pan, C.; Ye, L.; Qin, L.; Liu, X.; He, Y.; Wang, J.; Chen, L.; Lu, G. CRISPR/Cas9-mediated efficient and heritable targeted mutagenesis in tomato plants in the first and later generations. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ding, L.; Chen, Y.; Ma, Y.; Wang, H.; Wei, J. Effective reduction in chimeric mutants of poplar trees produced by CRISPR/Cas9 through a second round of shoot regeneration. Plant Biotechnol. Rep. 2020, 14, 549–558. [Google Scholar] [CrossRef]
- Triozzi, P.; Schmidt, H.W.; Dervinis, C.; Kirst, M.; Conde, D. Simple, efficient and open-source CRISPR/Cas9 strategy for multi-site genome editing in Populus tremula × alba. Tree Physiol. 2021, 41, 2216–2227. [Google Scholar] [CrossRef] [PubMed]
- Hasebe, F.; Yuba, H.; Hashimoto, T.; Saito, K.; Funa, N.; Shoji, T. CRISPR/Cas9-mediated disruption of the PYRROLIDINE KETIDE SYNTHASE gene reduces the accumulation of tropane alkaloids in Atropa belladonna hairy roots. Biosci. Biotechnol. Biochem. 2021, 85, 2404–2409. [Google Scholar] [CrossRef]
Figure 1.
Relationships between Agrobacterium rhizogenes and Agrobacterium tumefaciens strains. A. rhizogenes strains are given in red, A. tumefaciens strains in black, transconjugants between A. rhizogenes and A. tumefaciens are given in both colors. The abbreviations of the four main ancestral wild type Agrobacterium strains are highlighted by bold print and underlining: A4, ATCC15834, NCPPB1855, C58. Arrows show the directions of isolation of derivaties. Parts of Agrobacterium strain abbreviations: AR—Agrobacterium rhizogenes; ARqua1—Agrobacterium rhizogenes strain obtained by H.J. Quandt; ATCC—American Type Culture Collection; A4RS—A4 derivative resistant to rifampicin and spectinomycin; C58—A. tumefaciens strain isolated from cherry gall in 1958; C58C1 or C58C9—C58 strain cured of Ti plasmid (pTi), the numeric designation 1 or 9 refers to a colony number; GV—Ghent University and Vrije Universiteit Brussel; LBA—Lugdunum Batavorum Agrobacterium; MSU—Michigan State University; NCPPB—National Collection of Plant Pathogenic Bacteria. Plus or minus indicates a transfer or loss of any component in the resulting Agrobacterium strain. The chromosomal background or the Ri plasmid transferred to a new A. rhizogenes strain are indicated next to the arrows in black or red, respectively. Antibiotic resistances or other modifications of the chromosomal background or of the Ri plasmids are designated as follows. C58 derivatives cured of the Ti plasmid are indicated with an asterisk. WT stands for wild type Agrobacterium strain or for its native plasmids (pTi or pRi). Antibiotic resistances appearing either in the chromosome (g) or as inserts in pRi are given in green; Carb—carbenicillin, Cm (CAT)—chloramphenicol, Ery—erythromycin, Km—kanamycin, Nal—nalidixic acid, Sp—spectinomycin, Str—streptomycin, Tet—tetracycline; a question mark denotes suggested resistance. Other modifications inserted into pRi (given in blue): pAR5—vector harboring genes encoding either β-glucuronidase (GUS) or chloramphenicol acetyltransferase (CAT) or luciferase (LUC), was integrated into wild type pRi15834 of the strain AR10 his− via homologous recombination; pBR322—part of the pBR322 sequence inserted into wild type pRi15834 of the strain C58C1 (pRi15834) via homologous recombination; tmr—cytokinin synthesis locus encoding an isopentenyl transferase; vir reg.—part of the vir region from pTiBO342 (conferring the supervirulent phenotype to A. tumefaciens A348). Other modifications (given in brown): His-—histidine auxotrophy acquired by random mutagenesis of C58C1 (pRi15834). The sequencer icon (created with BioRender) indicates Agrobacterium strains with sequenced genome.
Figure 1.
Relationships between Agrobacterium rhizogenes and Agrobacterium tumefaciens strains. A. rhizogenes strains are given in red, A. tumefaciens strains in black, transconjugants between A. rhizogenes and A. tumefaciens are given in both colors. The abbreviations of the four main ancestral wild type Agrobacterium strains are highlighted by bold print and underlining: A4, ATCC15834, NCPPB1855, C58. Arrows show the directions of isolation of derivaties. Parts of Agrobacterium strain abbreviations: AR—Agrobacterium rhizogenes; ARqua1—Agrobacterium rhizogenes strain obtained by H.J. Quandt; ATCC—American Type Culture Collection; A4RS—A4 derivative resistant to rifampicin and spectinomycin; C58—A. tumefaciens strain isolated from cherry gall in 1958; C58C1 or C58C9—C58 strain cured of Ti plasmid (pTi), the numeric designation 1 or 9 refers to a colony number; GV—Ghent University and Vrije Universiteit Brussel; LBA—Lugdunum Batavorum Agrobacterium; MSU—Michigan State University; NCPPB—National Collection of Plant Pathogenic Bacteria. Plus or minus indicates a transfer or loss of any component in the resulting Agrobacterium strain. The chromosomal background or the Ri plasmid transferred to a new A. rhizogenes strain are indicated next to the arrows in black or red, respectively. Antibiotic resistances or other modifications of the chromosomal background or of the Ri plasmids are designated as follows. C58 derivatives cured of the Ti plasmid are indicated with an asterisk. WT stands for wild type Agrobacterium strain or for its native plasmids (pTi or pRi). Antibiotic resistances appearing either in the chromosome (g) or as inserts in pRi are given in green; Carb—carbenicillin, Cm (CAT)—chloramphenicol, Ery—erythromycin, Km—kanamycin, Nal—nalidixic acid, Sp—spectinomycin, Str—streptomycin, Tet—tetracycline; a question mark denotes suggested resistance. Other modifications inserted into pRi (given in blue): pAR5—vector harboring genes encoding either β-glucuronidase (GUS) or chloramphenicol acetyltransferase (CAT) or luciferase (LUC), was integrated into wild type pRi15834 of the strain AR10 his− via homologous recombination; pBR322—part of the pBR322 sequence inserted into wild type pRi15834 of the strain C58C1 (pRi15834) via homologous recombination; tmr—cytokinin synthesis locus encoding an isopentenyl transferase; vir reg.—part of the vir region from pTiBO342 (conferring the supervirulent phenotype to A. tumefaciens A348). Other modifications (given in brown): His-—histidine auxotrophy acquired by random mutagenesis of C58C1 (pRi15834). The sequencer icon (created with BioRender) indicates Agrobacterium strains with sequenced genome.

Figure 2.
Map of a CRISPR/Cas9 vector. Vector components (clockwise): RB—right T-DNA border; CRISPR cassette containing P—promoter of the small nucleolar RNA (snoRNA) gene; gRNA—guide RNA; crR—crisprRNA (target specific); tracrR—trans-activating crisprRNA (conserved, used as a binding scaffold for Cas9); T—terminator of the snoRNA gene; NLS—nuclear localization signal; Cas9—CRISPR associated nuclease 9; LB—left T-DNA border.
Figure 2.
Map of a CRISPR/Cas9 vector. Vector components (clockwise): RB—right T-DNA border; CRISPR cassette containing P—promoter of the small nucleolar RNA (snoRNA) gene; gRNA—guide RNA; crR—crisprRNA (target specific); tracrR—trans-activating crisprRNA (conserved, used as a binding scaffold for Cas9); T—terminator of the snoRNA gene; NLS—nuclear localization signal; Cas9—CRISPR associated nuclease 9; LB—left T-DNA border.

Figure 3.
Localization of CsRALF34 expression in a Cucumis sativus root tip. Confocal laser scanning microscopy of vibratome longitudinal (A,B) and cross (C) sections of transgenic pCsRALF34::mNeonGreen-H2B roots. Green channel—fluorescence of mNeonGreen-H2B; magenta channel—cell walls are counter stained with SCRI Renaissance 2200. (A) An overview and (B) close-up of the parental root meristem shows the acropetal sequence of CsRALF34 promoter activity in protoxylem, pericycle and endodermis. CsRALF34 expression arises first in the protoxylem at a distance of 150 µm from the initial cells. (C) The establishment of activity in xylem, pericycle layers and endodermis on a cross section at a distance of 300 µm from the initial cells. Arrows indicate developing lateral root primordia. Scale bars: 100 µm in (A), and 40 μm in (B,C).
Figure 3.
Localization of CsRALF34 expression in a Cucumis sativus root tip. Confocal laser scanning microscopy of vibratome longitudinal (A,B) and cross (C) sections of transgenic pCsRALF34::mNeonGreen-H2B roots. Green channel—fluorescence of mNeonGreen-H2B; magenta channel—cell walls are counter stained with SCRI Renaissance 2200. (A) An overview and (B) close-up of the parental root meristem shows the acropetal sequence of CsRALF34 promoter activity in protoxylem, pericycle and endodermis. CsRALF34 expression arises first in the protoxylem at a distance of 150 µm from the initial cells. (C) The establishment of activity in xylem, pericycle layers and endodermis on a cross section at a distance of 300 µm from the initial cells. Arrows indicate developing lateral root primordia. Scale bars: 100 µm in (A), and 40 μm in (B,C).

Figure 4.
pKSEe401R plasmid. (A) Map of pKSEe401R; parts (clockwise): RB—right T-DNA border; pAtU6-26—promoter of the Arabidopsis small nucleolar RNA (snoRNA) U6-26 gene; SpR—gene encoding spectinomycin resistance; tracrRNA scaffold—trans-activating crisprRNA (conservative, used as a binding scaffold for Cas9); AtU6-26t—terminator of the Arabidopsis snoRNA U6-26 gene; p35S—35S promoter from the Cauliflower Mosaic Virus (CaMV); OsMac3—158 bp fragment of the OsMac3 5′-UTR; 3xFLAG—sequence encoding protein tag with the amino acid sequence DYKDHDGDYKDHDIDYKDDDDK; NLS—nuclear localization signal; SV40 NLS—NLS derived from Simian Virus 40 T antigen; zCas9—maize codon-optimized CRISPR associated nuclease 9; rbcS-E9 t—terminator from pea ribulose-1,5-bisphosphate carboxylase small subunit (rbcS) E9 gene; pAtUbi10—promoter from the Arabidopsis polyubiquitin 10 gene; DsRed1—gene encoding orange fluorescent protein DsRed1 from Discosoma sp.; NOS T—nopaline synthase terminator; KmR—gene encoding kanamycin resistance; CaMV poly(A) signal—polyadenylation signal from CaMV (often used as terminator sequence); LB—left T-DNA border. pKSEe401R nomenclature: p—plasmid; K—KmR; S—p35S; e4—zCas9 enhanced with OsMac3 5′ UTR; 01—pAtU6-26p; R—DsRed1. (B) pKSEe401R was constructed in two steps: (I) cloning of the 158 bp OsMac3 5′-UTR fragment into pKSE401 using Gibson Assembly resulted in pKSEe401; (II) insertion of the DsRed1 cassette into pKSEe401 using T4 ligation resulted in pKSEe401R with the DsRed1 cassette in either in tandem (Var. 1) or in reverse (Var. 2) orientation relative to the Cas9 cassette.
Figure 4.
pKSEe401R plasmid. (A) Map of pKSEe401R; parts (clockwise): RB—right T-DNA border; pAtU6-26—promoter of the Arabidopsis small nucleolar RNA (snoRNA) U6-26 gene; SpR—gene encoding spectinomycin resistance; tracrRNA scaffold—trans-activating crisprRNA (conservative, used as a binding scaffold for Cas9); AtU6-26t—terminator of the Arabidopsis snoRNA U6-26 gene; p35S—35S promoter from the Cauliflower Mosaic Virus (CaMV); OsMac3—158 bp fragment of the OsMac3 5′-UTR; 3xFLAG—sequence encoding protein tag with the amino acid sequence DYKDHDGDYKDHDIDYKDDDDK; NLS—nuclear localization signal; SV40 NLS—NLS derived from Simian Virus 40 T antigen; zCas9—maize codon-optimized CRISPR associated nuclease 9; rbcS-E9 t—terminator from pea ribulose-1,5-bisphosphate carboxylase small subunit (rbcS) E9 gene; pAtUbi10—promoter from the Arabidopsis polyubiquitin 10 gene; DsRed1—gene encoding orange fluorescent protein DsRed1 from Discosoma sp.; NOS T—nopaline synthase terminator; KmR—gene encoding kanamycin resistance; CaMV poly(A) signal—polyadenylation signal from CaMV (often used as terminator sequence); LB—left T-DNA border. pKSEe401R nomenclature: p—plasmid; K—KmR; S—p35S; e4—zCas9 enhanced with OsMac3 5′ UTR; 01—pAtU6-26p; R—DsRed1. (B) pKSEe401R was constructed in two steps: (I) cloning of the 158 bp OsMac3 5′-UTR fragment into pKSE401 using Gibson Assembly resulted in pKSEe401; (II) insertion of the DsRed1 cassette into pKSEe401 using T4 ligation resulted in pKSEe401R with the DsRed1 cassette in either in tandem (Var. 1) or in reverse (Var. 2) orientation relative to the Cas9 cassette.

Figure 5.
Types of the CRISPR/Cas9 systems. (A) System based on the Cas9 form producing a double-strand break, NHEJ—non-homologous end joining. (B) Approaches using nickase Cas9 (nCas9) activity resulting in a single-strand break, pegRNA—prime editing guide RNA comprised of a target specific crRNA, a conservative tracrRNA, a PBS—primer binding site, an RT template—RNA template for reverse transcription (direction of reverse transcription indicated by a dark blue arrow). The linker joining crRNA and tracrRNA with PBS and the RT template is indicated in orange. RT—reverse transcriptase; UGI—uracil-DNA glycosylase inhibitor domain. (C) Techniques using the catalytically inactive (dead) form of Cas9 (dCas9); types of effectors used in dCas9-based systems: (de)methyl.—methyltransferase or demethylase for epigenetic modifications of DNA; (de)acetyl.—acetyltransferase or deacetylase for epigenetic modifications of histones; F—fluorescent protein; VP64—tetrameric repeat of the minimal activation domain of herpes simplex viral protein 16; UGI—uracil-DNA glycosylase inhibitor domain; deaminase—cytidine or adenosine deaminase; SRDX—SUPERMAN Repression Domain X. For systems based on Cas9 and nCas9 activities, scissors indicated in red show the place where DNA strand breaks occur. Figures were created with BioRender.
Figure 5.
Types of the CRISPR/Cas9 systems. (A) System based on the Cas9 form producing a double-strand break, NHEJ—non-homologous end joining. (B) Approaches using nickase Cas9 (nCas9) activity resulting in a single-strand break, pegRNA—prime editing guide RNA comprised of a target specific crRNA, a conservative tracrRNA, a PBS—primer binding site, an RT template—RNA template for reverse transcription (direction of reverse transcription indicated by a dark blue arrow). The linker joining crRNA and tracrRNA with PBS and the RT template is indicated in orange. RT—reverse transcriptase; UGI—uracil-DNA glycosylase inhibitor domain. (C) Techniques using the catalytically inactive (dead) form of Cas9 (dCas9); types of effectors used in dCas9-based systems: (de)methyl.—methyltransferase or demethylase for epigenetic modifications of DNA; (de)acetyl.—acetyltransferase or deacetylase for epigenetic modifications of histones; F—fluorescent protein; VP64—tetrameric repeat of the minimal activation domain of herpes simplex viral protein 16; UGI—uracil-DNA glycosylase inhibitor domain; deaminase—cytidine or adenosine deaminase; SRDX—SUPERMAN Repression Domain X. For systems based on Cas9 and nCas9 activities, scissors indicated in red show the place where DNA strand breaks occur. Figures were created with BioRender.

Figure 6.
Cucumber (Cucumis sativus) hairy roots co-transformed with pKSEe401R with a DsRed1 cassette in tandem (A–C) or reverse (D–F) orientation relative to the Cas9 cassette. (B) DsRed1 fluorescence was absent when the fluorescent protein (FP) cassette was in tandem orientation relative to the Cas9 cassette; (E) DsRed1 fluorescence was visible when the FP cassette was in reverse orientation relative to the Cas9 cassette. (A,D) bright field illumination; (C,F) root autofluorescence. Scale bars denote 5 mm.
Figure 6.
Cucumber (Cucumis sativus) hairy roots co-transformed with pKSEe401R with a DsRed1 cassette in tandem (A–C) or reverse (D–F) orientation relative to the Cas9 cassette. (B) DsRed1 fluorescence was absent when the fluorescent protein (FP) cassette was in tandem orientation relative to the Cas9 cassette; (E) DsRed1 fluorescence was visible when the FP cassette was in reverse orientation relative to the Cas9 cassette. (A,D) bright field illumination; (C,F) root autofluorescence. Scale bars denote 5 mm.

Figure 7.
Number of studies on CRISPR/Cas9 genome editing in hairy roots (analyzed on the 13 December 2021). Histogram is based on the Google-search of studies presented in Table S1.
Figure 7.
Number of studies on CRISPR/Cas9 genome editing in hairy roots (analyzed on the 13 December 2021). Histogram is based on the Google-search of studies presented in Table S1.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kiryushkin, A.S.; Ilina, E.L.; Guseva, E.D.; Pawlowski, K.; Demchenko, K.N. Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation. Plants 2022, 11, 51. https://doi.org/10.3390/plants11010051
AMA Style
Kiryushkin AS, Ilina EL, Guseva ED, Pawlowski K, Demchenko KN. Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation. Plants. 2022; 11(1):51. https://doi.org/10.3390/plants11010051
Chicago/Turabian StyleKiryushkin, Alexey S., Elena L. Ilina, Elizaveta D. Guseva, Katharina Pawlowski, and Kirill N. Demchenko. 2022. "Hairy CRISPR: Genome Editing in Plants Using Hairy Root Transformation" Plants 11, no. 1: 51. https://doi.org/10.3390/plants11010051
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.