
Effect of Elevated CO2 on Biomolecules’ Accumulation in Caraway (Carum carvi L.) Plants at Different Developmental Stages

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
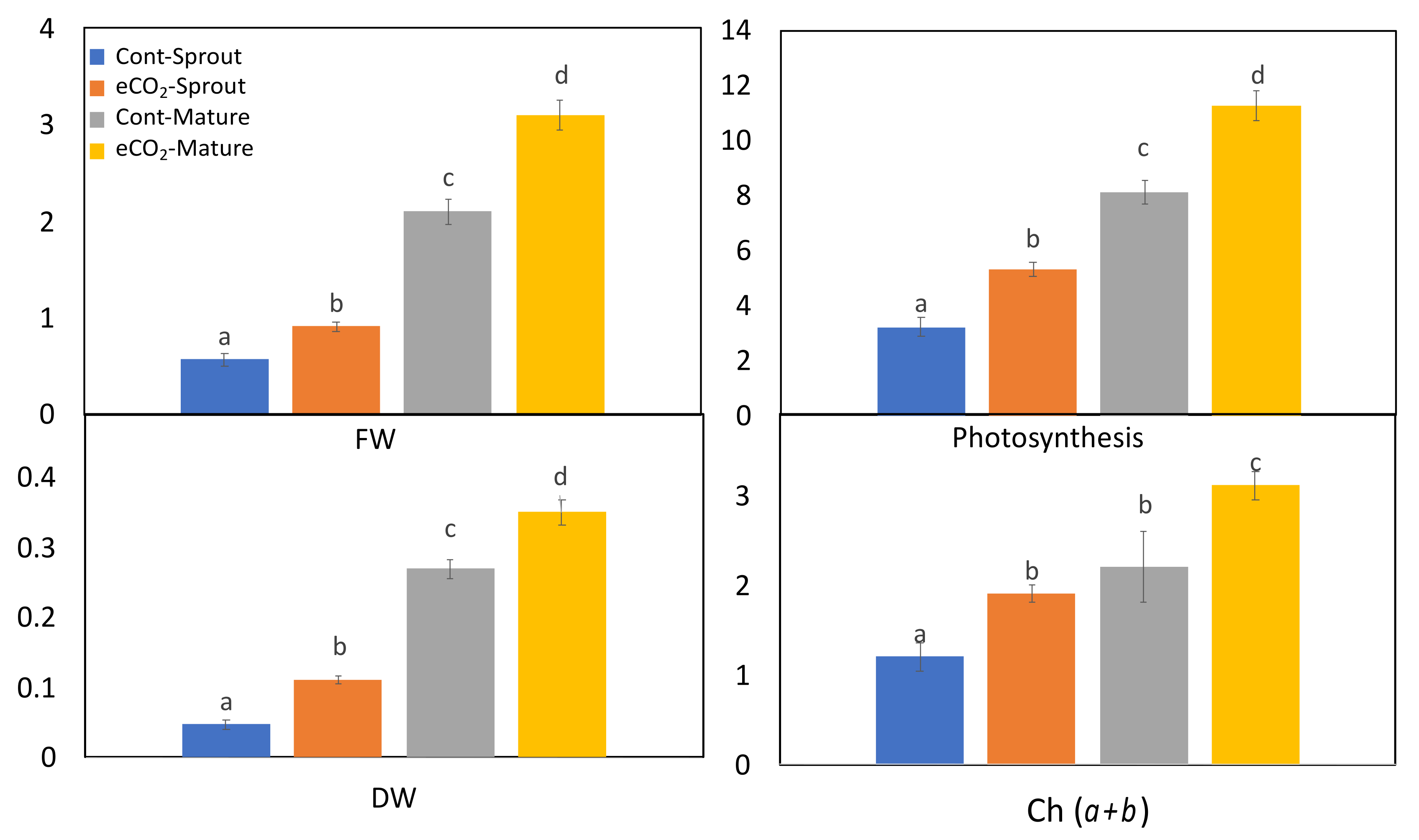
2.1. Growth and Biomass Production of Caraway Sprouts and Mature Plants as Affected by eCO2
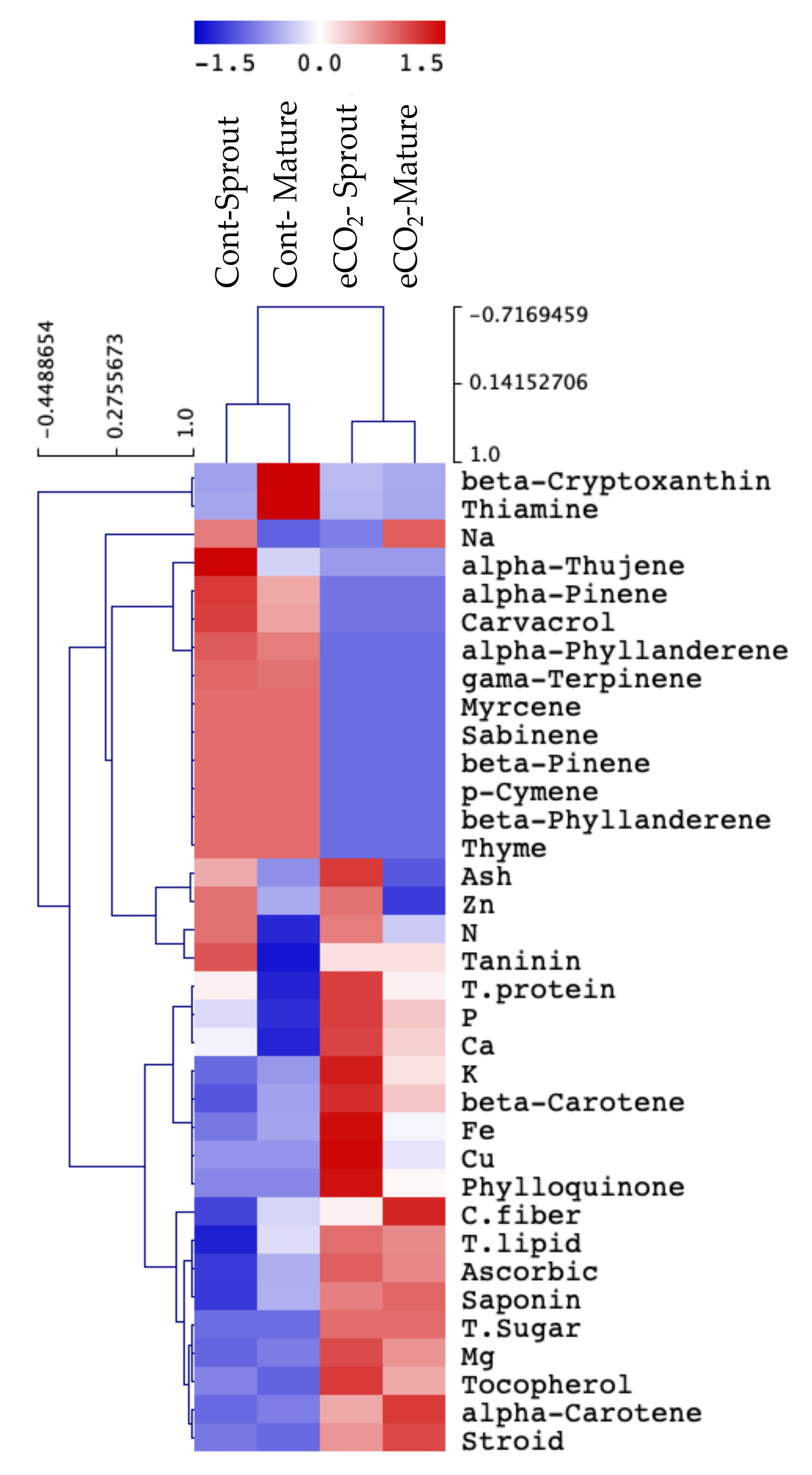
2.2. Impact of eCO2 on Minerals and Primary Metabolites of Caraway Sprouts and Mature Plants
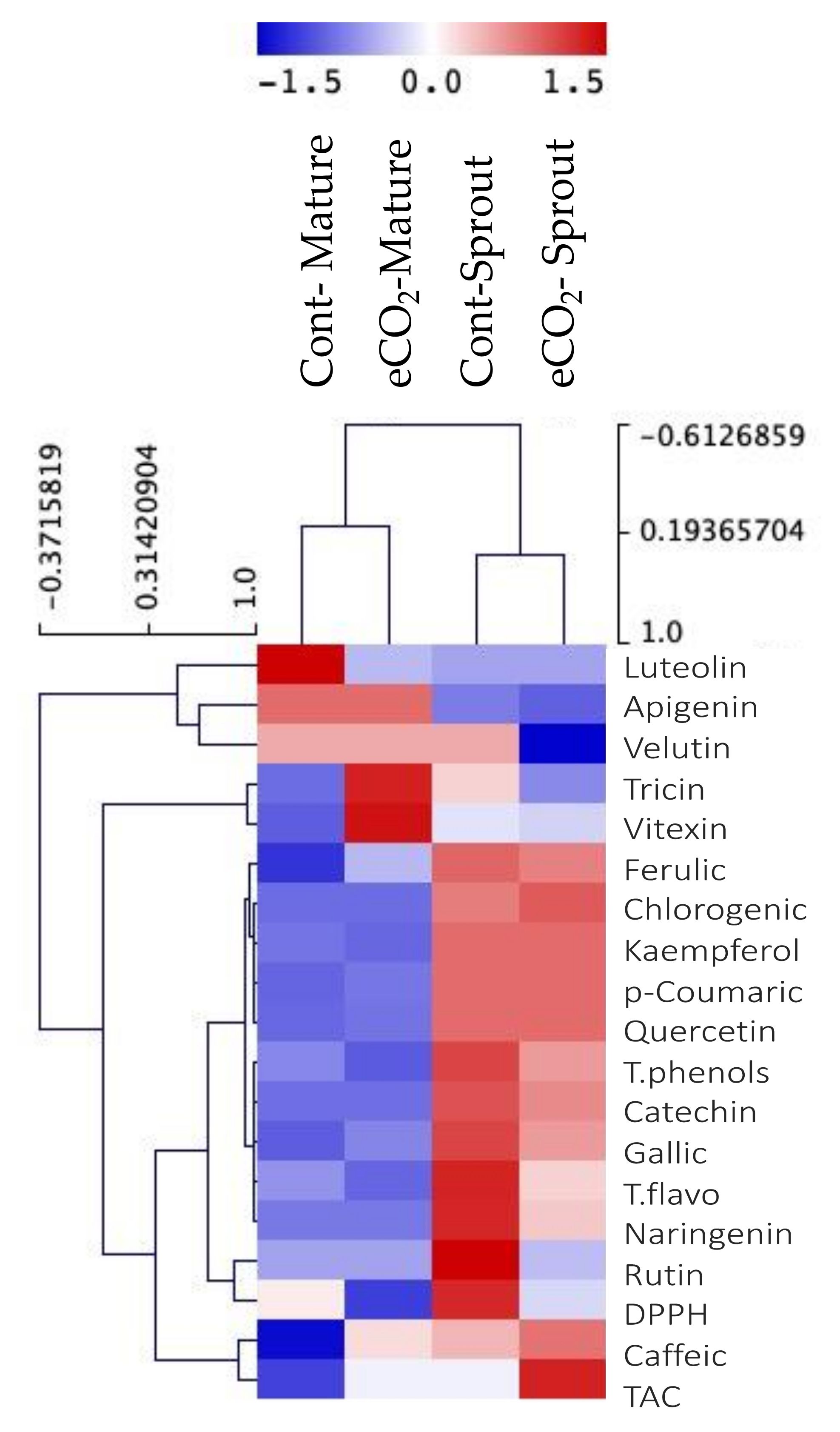
2.3. Secondary Metabolites of Caraway Sprouts and Mature Plants as Influenced by eCO2
2.4. Effect of eCO2 on Antioxidant Activities of Caraway Sprouts and Mature Plants
2.5. Antimicrobial Activity of Caraway Sprouts and Mature Plants under eCO2 Treatment
3. Discussion
3.1. eCO2 Enhanced Biomass Production of Caraway Sprouts and Mature Plants
3.2. eCO2 Similarly Affected the Primary Chemical Composition of Caraway Sprouts and Mature Tissue
3.3. eCO2 Differentially Affected the Mineral and Secondary Chemical Compostion of Caraway Sprouts and Mature Tissue
3.4. eCO2 Treatments Induced Higher Biological Activities in Sprouts than in Mature Tissues of Caraway Plants
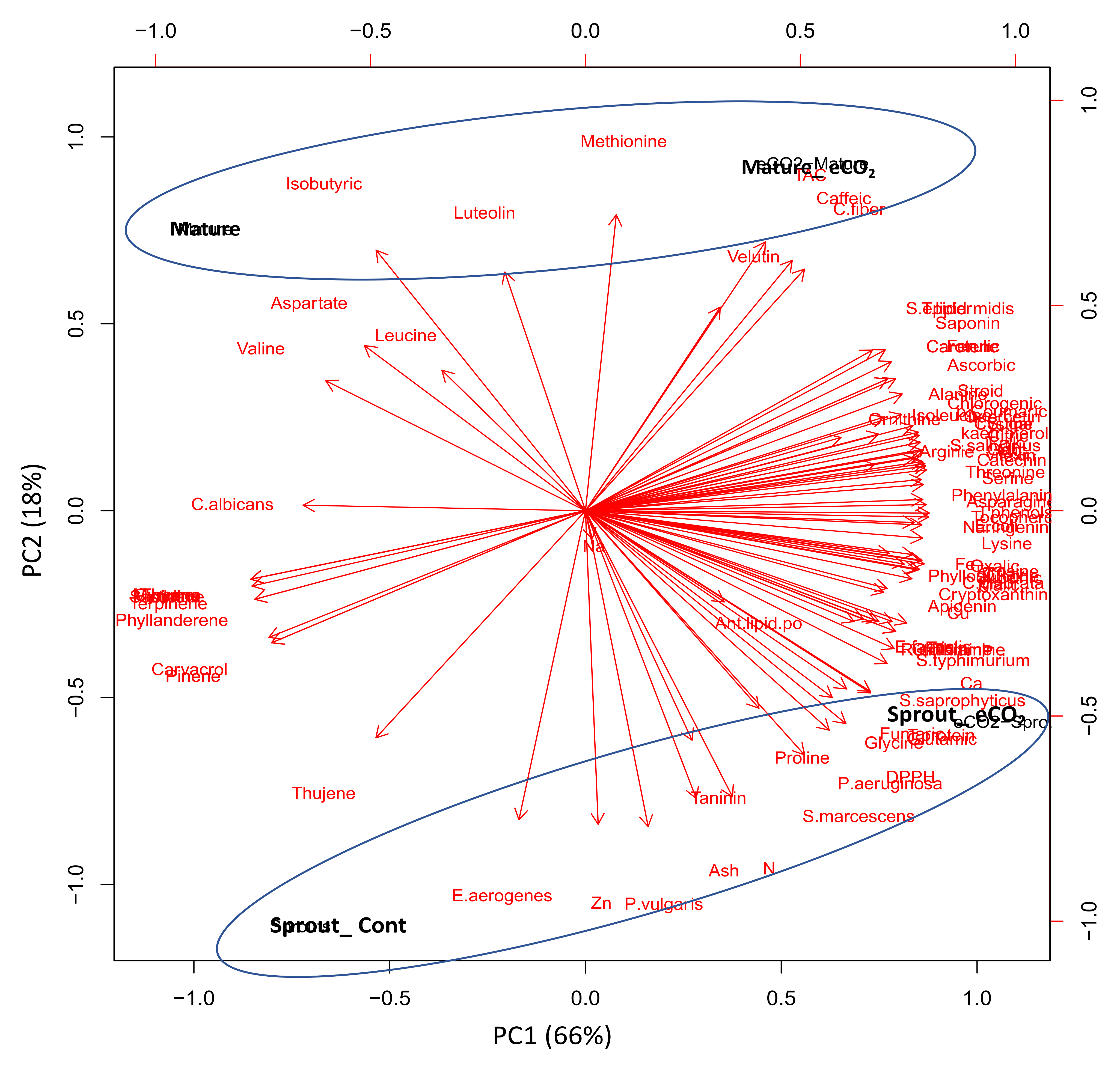
3.5. Principal Component Analysis (PCA) Confirmed the Developmental Stage-Specific Response of Caraway Plants to eCO2
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Determination of Photosynthetic Rate
4.3. Pigment Analysis
4.4. Preparation of Caraway Extracts
4.5. Analysis of Mineral Contents
4.6. Measurement of Phenolic Acids, Flavonoids and Vitamins
4.7. Measurement of Antioxidant Capacity
4.7.1. Ferric Reducing Antioxidant Power (FRAP) Method
4.7.2. DPPH• Assay
4.7.3. Lipid Peroxidation Assay
4.8. Determination of Total Carbohydrates, Protein, Lipids and Fibers
4.9. Analysis of Essential Oils, Organic Acids, and Amino Acids
4.9.1. Analysis of Essential Oils
4.9.2. Organic Acids Analysis
4.9.3. Amino Acid Analysis
4.10. Antibacterial Activity of Caraway Extracts
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Galieni, A.; Falcinelli, B.; Stagnari, F.; Datti, A.; Benincasa, P. Sprouts and Microgreens: Trends, Opportunities, and Horizons for Novel Research. Agronomy 2020, 10, 1424. [Google Scholar] [CrossRef]
- Silva, L.R.; Pereira, M.J.; Azevedo, J.; Gonçalves, R.F.; Valentão, P.; de Pinho, P.G.; Andrade, P.B. Glycine max (L.) Merr., Vigna radiata L. and Medicago sativa L. sprouts: A natural source of bioactive compounds. Food Res. Int. 2013, 50, 167–175. [Google Scholar] [CrossRef]
- Marton, M.; Mandoki, Z.S.; Csapo-Kiss, Z.S.; Csapo, J. The role of sprouts in human nutrition. A review. Acta Univ. Sapientiae 2010, 3, 81–117. [Google Scholar]
- Kapoor, L.D. Handbook of Ayurvedic Medicinal Plants; CRC Press Inc.: Boca Raton, FL, USA, 1990. [Google Scholar]
- Tyler, V.E.; Brady, L.R.; Robbers, J.E. Pharmacognosy, 9th ed.; Lea Fabiger: Philadelphia, PA, USA, 1988. [Google Scholar]
- Dubey, N.K.; Mishra, A.K. Evaluation of some essential oils against dermatophytes. Indian Drugs 1990, 27, 529–531. [Google Scholar]
- Boskabady, M.; Shaikhi, J. Inhibitory effect of Carum copticum on histamine (H1) receptors of isolated guinea-pig tracheal chains. J. Ethnopharmacol. 2000, 69, 217–227. [Google Scholar] [CrossRef]
- Vasudevan, K.; Vembar, S.; Veeraraghavan, K.; Haranath, P.S. Influence of intragastric perfusion of aqueous spice extracts on acid secretion in anesthetized albino rats. Indian J. Gastroenterol. 2000, 19, 53–56. [Google Scholar]
- Pradeep, K.U.; Geervani, P.; Eggum, B.O. Common Indian spices: Nutrient composition, consumption and contribution to dietary value. Plant Foods Hum. Nutr. 1993, 44, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Ruszkowska, J. Main Chemical Constituents of Carum. Caraway-The Genus Carum; Németh, É., Ed.; Harwood Academic Publishers: Amsterdam, The Netherlands, 1998; pp. 38–60. [Google Scholar]
- Sedláková, J.; Kocourková, B.; Kubáň, V. Determination of essential oils content and composition in caraway (Carum carvi L.). Czech J. Food Sci. 2013, 19, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Johri, R. Cuminum cyminum and Carum carvi: An update. Pharmacogn. Rev. 2011, 5, 63–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedges, L.J.; Lister, C.E. The nutritional attributes of legumes. Crop. Food Res. Confid. Rep. 2006, 1745, 50. [Google Scholar]
- Almuhayawi, M.S.; Al Jaouni, S.K.; Almuhayawi, S.M.; Selim, S.; Abdel-Mawgoud, M. Elevated CO2 improves the nutritive value, antibacterial, anti-inflammatory, antioxidant and hypocholestecolemic activities of lemongrass sprouts. Food Chem. 2021, 357, 129730. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Feng, H.; Kushad, M.M.; Fan, X. Effects of Ultrasound, Irradiation, and Acidic Electrolyzed Water on Germination of Alfalfa and Broccoli Seeds and Escherichia coli O157: H7. J. Food Sci. 2006, 71, M168–M173. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Hassan, A.H.A.; Abdel-Mawgoud, M.; Khamis, G.; Selim, S.; Al Jaouni, S.K.; AbdElgawad, H. Laser light as a promising approach to improve the nutritional value, antioxidant capacity and anti-inflammatory activity of flavonoid-rich buckwheat sprouts. Food Chem. 2020, 345, 128788. [Google Scholar] [CrossRef]
- Saleh, A.M.; Selim, S.; Al Jaouni, S.; AbdElgawad, H. CO2 enrichment can enhance the nutritional and health benefits of parsley (Petroselinum crispum L.) and dill (Anethum graveolens L.). Food Chem. 2018, 269, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.M.; Abdel-Mawgoud, M.; Hassan, A.R.; Habeeb, T.H.; Yehia, R.S.; AbdElgawad, H. Global metabolic changes in-duced by arbuscular mycorrhizal fungi in oregano plants grown under ambient and elevated levels of atmospheric CO2. Plant Physiol. Biochem. 2020, 151, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Wang, J.; Shen, S.; Wang, Y.; Zhu, J.; Wang, Y.; Yang, L. The impact of elevated CO2 and temperature on grain quality of rice grown under open-air field conditions. J. Sci. Food Agric. 2016, 96, 3658–3667. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Li, H.; Zong, Y.; Li, F.Y.; Han, Y.; Hao, X. Photosynthesis and metabolite responses of Isatis indigotica Fortune to ele-vated [CO2]. Crop J. 2017, 5, 345–353. [Google Scholar] [CrossRef]
- Högy, P.; Wieser, H.; Köhler, P.; Schwadorf, K.; Breuer, J.; Franzaring, J.; Muntifering, R.; Fangmeier, A. Effects of elevated CO2 on grain yield and quality of wheat: Results from a 3-year free-air CO2 enrichment experiment. Plant Biol. 2009, 11, 60–69. [Google Scholar] [CrossRef]
- Azam, A.; Khan, I.; Mahmood, A.; Hameed, A. Yield, chemical composition and nutritional quality responses of carrot, radish and turnip to elevated atmospheric carbon dioxide. J. Sci. Food Agric. 2013, 93, 3237–3244. [Google Scholar] [CrossRef] [PubMed]
- Al Jaouni, S.; Saleh, A.M.; Wadaan, M.A.M.; Hozzein, W.N.; Selim, S.; AbdElgawad, H. Elevated CO2 induces a global met-abolic change in basil (Ocimum basilicum L.) and peppermint (Mentha piperita L.) and improves their biological activity. J. Plant Physiol. 2018, 224, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Almuhayawi, M.; AbdElgawad, H.; Al Jaouni, S.; Selim, S.; Hassan, A.H.A.; Khamis, G. Elevated CO2 improves glucosinolate metabolism and stimulates anticancer and anti-inflammatory properties of broccoli sprouts. Food Chem. 2020, 328, 127102. [Google Scholar] [CrossRef]
- Almuhayawi, M.S.; Hassan, A.H.A.; Al Jaouni, S.K.; Alkhalifah, D.H.M.; Hozzein, W.N.; Selim, S.; AbdElgawad, H.; Khamis, G. Influence of elevated CO2 on nutritive value and health-promoting prospective of three genotypes of Alfalfa sprouts (Medicago Sativa). Food Chem. 2020, 340, 128147. [Google Scholar] [CrossRef] [PubMed]
- Leakey, A.; Ainsworth, E.A.; Bernacchi, C.; Rogers, A.; Long, S.; Ort, D.R. Elevated CO2 effects on plant carbon, nitrogen, and water relations: Six important lessons from FACE. J. Exp. Bot. 2009, 60, 2859–2876. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, G.; Sun, B.; Zhang, S.; Zhang, Y.; Liao, Y.; Zhou, Y.; Xia, X.; Shi, K.; Yu, J. Stimulated Leaf Dark Respiration in Tomato in an Elevated Carbon Dioxide Atmosphere. Sci. Rep. 2013, 3, 3433. [Google Scholar] [CrossRef] [Green Version]
- Noguchi, K.; Watanabe, C.K.; Terashima, I. Effects of Elevated Atmospheric CO2 on Primary Metabolite Levels inArabidopsis thalianaCol-0 Leaves: An Examination of Metabolome Data. Plant Cell Physiol. 2015, 56, 2069–2078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hozzein, W.N.; Saleh, A.M.; Habeeb, T.H.; Wadaan, M.A.M.; AbdElgawad, H. CO2 treatment improves the hypocholester-olemic and antioxidant properties of fenugreek seeds. Food Chem. 2020, 308, 125661. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and Signaling Aspects Underpinning the Regulation of Plant Carbon Nitrogen Interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef]
- Kanunnikova, N.P. Role of brain glutamic acid metabolism changes in neurodegenerative pathologies. J. Biol. 2012, 2, M1–M10. [Google Scholar]
- Zieliński, H.; Frias, J.; Piskuła, M.K.; Kozłowska, H.; Vidal-Valverde, C. Vitamin B 1 and B 2, dietary fiber and minerals content of Cruciferae sprouts. Eur. Food Res. Technol. 2005, 221, 78–83. [Google Scholar] [CrossRef]
- Watanabe, C.K.; Sato, S.; Yanagisawa, S.; Uesono, Y.; Terashima, I.; Noguchi, K. Effects of elevated CO2 on levels of primary metabolites and transcripts of genes encoding respiratory enzymes and their diurnal patterns in Arabidopsis thaliana: Possible relationships with respiratory rates. Plant Cell Physiol. 2014, 55, 341–357. [Google Scholar] [CrossRef] [Green Version]
- Asif, M.; Yilmaz, O.; Ozturk, L. Elevated carbon dioxide ameliorates the effect of Zn deficiency and terminal drought on wheat grain yield but compromises nutritional quality. Plant Soil 2017, 411, 57–67. [Google Scholar] [CrossRef]
- Teng, N.; Wang, J.; Chen, T.; Wu, X.; Wang, Y.; Lin, J. Elevated CO2 induces physiological, biochemical and structural changes in leaves of Arabidopsis thaliana. New Phytol. 2006, 172, 92–103. [Google Scholar] [CrossRef]
- Roberntz, P.; Linder, S. Effects of long-term CO2 enrichment and nutrient availability in Norway spruce. II. Foliar chemistry. Trees 1999, 14, 17–27. [Google Scholar] [CrossRef]
- LUOMALA, E.; Laitinen, K.; Sutinen, S.; Kellomäki, S.; Vapaavuori, E. Stomatal density, anatomy and nutrient concentra-tions of Scots pine needles are affected by elevated CO2 and temperature. Plant Cell Environ. 2005, 28, 733–749. [Google Scholar] [CrossRef]
- Ishikawa, T.; Takayanagi, T.; Kitajima, J. Water-Soluble Constituents of Cumin: Monoterpenoid Glucosides. Chem. Pharm. Bull. 2002, 50, 1471–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kunzemann, J.; Herrmann, K. Isolation and identification of flavon (ol)-O-glycosides in caraway (Carum carvi L.), fennel (Foeniculum vulgare Mill.), anise (Pimpinella anisum L.), and coriander (Coriandrum sativum L.), and of flavon-C-glycosides in anise. I. Phenolics of spices (author’s transl). Zeitschrift fur Lebensmittel-untersuchung und-Forschung. 1977, 164, 194–200. [Google Scholar]
- Matsumura, T.; Ishikawa, T.; Kitajima, J. Water-Soluble Constituents of Caraway: Carvone Derivatives and Their Glucosides. Chem. Pharm. Bull. 2002, 50, 66–72. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, T.; Ishikawa, T.; Kitajima, J. Water-soluble constituents of caraway: Aromatic compound, aromatic compound glucoside and glucides. Phytochemistry 2002, 61, 455–459. [Google Scholar] [CrossRef]
- Najda, A.; Dyduch, J.; Brzozowski, N. Flavonoid Content and Antioxidant Activity of Caraway Roots (Carum Carvi L.). Veg. Crop. Res. Bull. 2008, 68, 127–133. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Matsunaga, H.; Saita, T.; Mori, M.; Katano, M.; Okabe, H. Antiproliferative Constituents in Umbelliferae Plants II. Screening for Polyacetylenes in Some Umbelliferae Plants, and Isolation of Panaxynol and Falcarindiol from the Root of Heracleum moellendorffii. Biol. Pharm. Bull. 1998, 21, 257–261. [Google Scholar] [CrossRef] [Green Version]
- Idso, S.B.; Kimball, B.A.; Shaw, P.E.; Widmer, W.; Vanderslice, J.T.; Higgs, D.J.; Montanari, A.; Clark, W.D. The effect of ele-vated atmospheric CO2 on the vitamin C concentration of (sour) orange juice. Agric. Ecosyst. Environ. 2002, 90, 1–7. [Google Scholar] [CrossRef]
- De Carvalho, C.C.C.R.; Da Fonseca, M.M.R. Carvone: Why and how should one bother to produce this terpene. Food Chem. 2006, 95, 413–422. [Google Scholar] [CrossRef]
- Sedláková, J.; Kocourková, B.; Lojková, L.; Kubáň, V. Determination of essential oil content in caraway (Carum carvi L.) species by means of supercritical fluid extraction. Plant Soil Environ. 2011, 49, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Oosterhaven, K.; Poolman, B.; Smid, E. S-carvone as a natural potato sprout inhibiting, fungistatic and bacteristatic compound. Ind. Crop. Prod. 1995, 4, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Chizzola, R. Composition of the Essential Oil of Wild Grown Caraway in Meadows of the Vienna Region (Austria). Nat. Prod. Commun. 2014, 9, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Darwish, M.S.; Ofir, R. Heavy metals content and essential oil yield of Juniperus phoenicea L. in different origins in Jordan. Environ. Eng. Manag. J. 2014, 13, 3009–3014. [Google Scholar] [CrossRef]
- Kolta, R.; Hornok, L. Extraction processes for essential oils. Cultiv. Process. Med. Plants 1992, 4, 93–104. [Google Scholar]
- Van der Mheen, H.J. Vergelijking en Verloop (Tijdens de Afrijping) van de Zaad en Carvonopbrengst van (Zomer) Karwij en Dille= Comparison and Development (during the Ripening Phase) of the Seed and Carvone Yield of (Summer) Caraway and Dill; PAGV: Lelystad, The Netherlands, 1994. [Google Scholar]
- Al-Essa, M.K.; Shafagoj, Y.A.; Mohammed, F.I.; Afifi, F.U. Relaxant effect of ethanol extract of Carum carvi on dispersed intestinal smooth muscle cells of the guinea pig. Pharm. Biol. 2009, 48, 76–80. [Google Scholar] [CrossRef]
- Joshi, S.G.; Joshi, S.G. Medicinal Plants; Oxford and IBH Publishing: New Delhi, India, 2000; Volume 491. [Google Scholar]
- Zheng, G.-Q.; Kenney, P.M.; Lam, L.K.T. Anethofuran, Carvone, and Limonene: Potential Cancer Chemoprotective Agents from Dill Weed Oil and Caraway Oil. Planta Med. 1992, 58, 338–341. [Google Scholar] [CrossRef]
- Lado, C.; Then, M.; Varga, I.; Szőke, É.; Szentmihályid, K. Antioxidant property of volatile oils determined by the ferric reducing ability. Zeitschrift für Naturforschung C 2004, 59, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Raphael, T.J.; Kuttan, G. Immunomodulatory Activity of Naturally Occurring Monoterpenes Carvone, Limonene, and Perillic Acid. Immunopharmacol. Immunotoxicol. 2003, 25, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, H.Z.; Ibrahim, M.H.; Karimi, E. Phenolics and Flavonoids Compounds, Phenylanine Ammonia Lyase and Antioxidant Activity Responses to Elevated CO2 in Labisia pumila (Myrisinaceae). Molecules 2012, 17, 6331–6347. [Google Scholar] [CrossRef] [Green Version]
- AbdElgawad, H.; Farfan-Vignolo, E.R.; de Vos, D.; Asard, H. Elevated CO2 mitigates drought and temperature-induced oxidative stress differently in grasses and legumes. Plant Sci. 2015, 231, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Karimi, E.; Oskoueian, E.; Oskoueian, A.; Omidvar, V.; Hendra, R.; Nazeran, H. Insight into the functional and medicinal properties of Medicago sativa (Alfalfa) leaves extract. J. Med. Plants Res. 2013, 7, 290–297. [Google Scholar]
- Wang, S.Y.; Bunce, J.A.; Maas, J.L. Elevated Carbon Dioxide Increases Contents of Antioxidant Compounds in Field-Grown Strawberries. J. Agric. Food Chem. 2003, 51, 4315–4320. [Google Scholar] [CrossRef] [PubMed]
- Szydłowska-Czerniak, A.; Tułodziecka, A.; Szłyk, E. Determination of Antioxidant Capacity of Unprocessed and Processed Food Products by Spectrophotometric Methods. Food Anal. Methods 2011, 5, 807–813. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, N.; Visioli, F.; Buratti, S.; Brighenti, F. Direct Analysis of Total Antioxidant Activity of Olive Oil and Studies on the Influence of Heating. J. Agric. Food Chem. 2001, 49, 2532–2538. [Google Scholar] [CrossRef] [PubMed]
- Achat, S.; Rakotomanomana, N.; Madani, K.; Dangles, O. Antioxidant activity of olive phenols and other dietary phenols in model gastric conditions: Scavenging of the free radical DPPH• and inhibition of the haem-induced peroxidation of linoleic acid. Food Chem. 2016, 213, 135–142. [Google Scholar] [CrossRef]
- Arabameri, M.; Nazari, R.R.; Abdolshahi, A.; Abdollahzadeh, M.; Mirzamohammadi, S.; Shariatifar, N.; Barba, F.J.; Mousavi Khaneghah, A. Oxidative stability of virgin olive oil: Evaluation and prediction with an adaptive neuro-fuzzy inference system (ANFIS). J. Sci. Food Agric. 2019, 99, 5358–5367. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Ramezan, Y.; Arab, S. A comparative study on physicochemical characteristics and antioxidant activity of su-mac (Rhus coriaria L.), cumin (Cuminum cyminum), and caraway (Carum carvil) oils. J. Food Meas. Charact. 2020, 14, 3175–3183. [Google Scholar] [CrossRef]
- Fedor, M.; Socha, K.; Urban, B.; Soroczyńska, J.; Matyskiela, M.; Borawska, M.H.; Bakunowicz-Łazarczyk, A. Serum concen-tration of zinc, copper, selenium, manganese, and Cu/Zn Ratio in children and adolescents with myopia. Biol. Trace Elem. Res. 2017, 176, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Sadowska, A.; Obidoska, G. Pharmacological uses and toxicology of caraway. In Caraway; CRC Press: Boca Raton, FL, USA, 1999; pp. 198–208. ISBN 0429219679. [Google Scholar]
- Toxopeus, H.; Bouwmeester, H. Improvement of caraway essential oil and carvone production in The Netherlands. Ind. Crop. Prod. 1992, 1, 295–301. [Google Scholar] [CrossRef]
- Hartmans, K.J.; Diepenhorst, P.; Bakker, W.; Gorris, L.G. The use of carvone in agriculture: Sprout suppression of potatoes and antifungal activity against potato tuber and other plant diseases. Ind. Crop. Prod. 1995, 4, 3–13. [Google Scholar] [CrossRef]
- Bailer, J.; Aichinger, T.; Hackl, G.; de Hueber, K.; Dachler, M. Essential oil content and composition in commercially available dill cultivars in comparison to caraway. Ind. Crop. Prod. 2001, 14, 229–239. [Google Scholar] [CrossRef]
- Frank, T.; Biert, K.; Speiser, B. Feeding deterrent effect of carvone, a compound from caraway seeds, on the slug Arion lusitanicus. Ann. Appl. Biol. 2002, 141, 93–100. [Google Scholar] [CrossRef]
- Simic, A.; Rančic, A.; Sokovic, M.D.; Ristic, M.; Grujic-Jovanovic, S.; Vukojevic, J.; Marin, P.D. Essential Oil Composition of Cymbopogon winterianus and Carum carvi. and Their Antimicrobial Activities. Pharm. Biol. 2008, 46, 437–441. [Google Scholar] [CrossRef] [Green Version]
- Almuhayawi, M.; Mohamed, M.; Abdel-Mawgoud, M.; Selim, S.; Al Jaouni, S.; AbdElgawad, H. Bioactive Potential of Several Actinobacteria Isolated from Microbiologically Barely Explored Desert Habitat, Saudi Arabia. Biology 2021, 10, 235. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH• assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Noble, J.E.; Bailey, M.J.A. Quantitation of protein. Methods Enzymol. 2009, 463, 73–95. [Google Scholar]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Habeeb, T.H.; Abdel-Mawgoud, M.; Yehia, R.S.; Khalil, A.M.A.; Saleh, A.M.; AbdElgawad, H. Interactive Impact of Arbuscular Mycorrhizal Fungi and Elevated CO2 on Growth and Functional Food Value of Thymus vulgare. J. Fungi 2020, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Sturm, K.; Koron, D.; Stampar, F. The composition of fruit of different strawberry varieties depending on maturity stage. Food Chem. 2003, 83, 417–422. [Google Scholar] [CrossRef]
- Vázquez-Ortiz, F.A.; Caire, G.; Higuera-Ciapara, I.; Hernández, G. High Performance Liquid Chromatographic Determination of Free Amino Acids in Shrimp. J. Liq. Chromatogr. 1995, 18, 2059–2068. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic Acids | Sprouts | Mature | ||
|---|---|---|---|---|
| (mg/g FW) | Control | eCO2 | Control | eCO2 |
| Oxalic | 1.42 ± 0.14 a | 1.99 ± 0.2 b | 1.2 ± 0.10 a | 1.91 ± 0.2 b |
| Malic | 5.06 ± 0.31 a | 6.49 ± 0.5 b | 4.1 ± 0.7 a | 6.09 ± 0.2 b |
| Succinic | 2.84 ± 0.16 a | 4.95 ± 0.43 b | 2.3 ± 0.2 a | 3.95 ± 0.41 b |
| Citric | 2.45 ± 0.1 a | 3.2 ± 0.39 b | 2.5 ± 0.4 a | 3.1 ± 0.19 b |
| isobutyric | 1.06 ± 0.11 a | 1.02 ± 0.01 a | 1.16 ± 0.1 a | 1.11 ± 0.01 a |
| Fumaric | 0.42 ± 0.03 b | 0.45 ± 0.01 b | 0.32 ± 0.0 a | 0.42 ± 0.0 b |
| Amino Acids (µg/g FW) | ||||
| Lysine | 1.57 ± 0.13 a | 2.62 ± 0.12 b | 1.51 ± 0.11 a | 2.12 ± 0.2 b |
| Histidine | 1.61 ± 0.089 a | 1.13 ± 0.01 a | 1.61 ± 0.09 a | 1.13 ± 0.01 a |
| Alanine | 0.54 ± 0.04 a | 1.23 ± 0.08 b | 0.34 ± 0.041 a | 1.7 ± 0.08 b |
| Arginine | 0.98 ± 0.01 a | 1.19 ± 0.09 a | 0.78 ± 0.03 a | 1.39 ± 0.1 b |
| Isoleucine | 0.08 ± 0.00 a | 0.13 ± 0.01 a | 0.1 ± 0.00 a | 0.11 ± 0.01 a |
| Asparagine | 0.52 ± 0.04 a | 0.91 ± 0.07 b | 0.52 ± 0.05 a | 0.77 ± 0.01 a |
| Ornithine | 0.1 ± 0.02 a | 0.12 ± 0.02 a | 0.11 ± 0.03 a | 0.11 ± 0.02 a |
| Glycine | 0.6 ± 0.04 b | 0.68 ± 0.06 b | 0.2 ± 0.046 a | 0.58 ± 0.01 b |
| Phenylalanine | 0.16 ± 0.013 a | 0.32 ± 0.01 b | 0.11 ± 0.015 a | 0.31 ± 0.04 b |
| Serine | 0.18 ± 0.01 a | 0.3 ± 0.02 b | 0.19 ± 0.01 a | 0.26 ± 0.01 b |
| Proline | 0.58 ± 0.04 a | 0.57 ± 0.05 a | 0.52 ± 0.05 a | 0.57 ± 0.05 a |
| Valine | 0.24 ± 0.01 a | 0.22 ± 0.03 a | 0.34 ± 0.05 b | 0.22 ± 0.03 a |
| Aspartate | 0.02 ± 0.001 a | 0.02 ± 0 a | 0.025 ± 0.0 a | 0.02 ± 0.0 a |
| Cystine | 0.01 ± 0.0 a | 0.1 ± 0.01 a | 0.01 ± 0.0 a | 0.1 ± 0.0 a |
| Leucine | 0.16 ± 0 b | 0.11 ± 0 a | 0.13 ± 0.019 a | 0.21 ± 0 b |
| Methionine | 0.01 ± 0.001 a | 0.01 ± 0 a | 0.02 ± 0.002 b | 0.031 ± 0 b |
| Threonine | 0.05 ± 0.0 a | 0.08 ± 0.01 b | 0.043 ± 0.0 a | 0.08 ± 0.01 b |
| Tyrosine | 0.5 ± 0.03 a | 0.66 ± 0.05 b | 0.42 ± 0.039 a | 0.61 ± 0.05 b |
| Glutamine | 46.3 ± 3.5 b | 52.79 ± 0.6 c | 33.7 ± 4.2 a | 51.1 ± 4.2 d |
| Glutamic acid | 33.8 ± 2.5 b | 46.2 ± 5.5 c | 22.8 ± 5.8 a | 33.2 ± 5.1 b |
| Sprouts | Mature | |||
|---|---|---|---|---|
| Microbial Name | Control (mm) | eCO2 (mm) | Control (mm) | eCO2 (mm) |
| Staphylococcus saprophyticus | 15.34 ± 2.2 b | 22.48 ± 1.8 c | 11.14 ± 1.3 a | 16.08 ± 1.29 b |
| Staphylococcus epidermidis | 9.27 ± 1 a | 20.13 ± 1.6 b | 11.27 ± 1.2 a | 22.1 ± 1.6 b |
| Enterococcus faecalis | 14.57 ± 3.4 b | 17.9 ± 0.5 c | 9.52 ± 3.42 a | 16.3 ± 0.56 b |
| Streptococcus salivarius | 7.87 ± 1.15 a | 16.14 ± 2.7 b | 8.82 ± 1.12 a | 13.11 ± 0.1 b |
| Escherichia coli | 6.3 ± 0.9 b | 9.68 ± 7.8 c | 4.3 ± 0.9 a | 9.68 ± 30.2 c |
| Salmonella typhimurium | 11.17 ± 1.9 b | 19.91 ± 3.1 c | 7.17 ± 1.2 a | 12.1 ± 1.1 b |
| Pseudomonas aeruginosa | 20.02 ± 1.7 b | 27.15 ± 1.5 c | 14.02 ± 1.7 a | 17.15 ± 1.1 b |
| Proteus vulgaris | 20.8 ± 1.0 c | 18.53 ± 1.82 b | 10.8 ± 1.2 a | 13.2 ± 1.5 ab |
| Enterobacter aerogenes | 14.92 ± 0.3 c | 13.96 ± 0.8 b | 12.12 ± 0.9 ab | 9.96 ± 0.17 a |
| Serratia marcescens | 5.92 ± 0.2 b | 7.68 ± 1.1 c | 3.91 ± 0.96 a | 4.68 ± 1.6 ab |
| Aspergillus flavus | 13.93 ± 1.1 b | 14.85 ± 1.8 b | 7.91 ± 1.5 a | 8.85 ± 1.0 a |
| Candida albicans | 15.58 ± 5.07 b | 15.76 ± 1.2 a | 14.51 ± 1.01 a | 14.2 ± 0.3 a |
| Candida glabrata | 4.11 ± 0.954 a | 8.11 ± 1.4 c | 3.41 ± 0.92 a | 5.91 ± 0.04 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
AbdElgawad, H.; Okla, M.K.; Al-amri, S.S.; AL-Hashimi, A.; AL-Qahtani, W.H.; Al-Qahtani, S.M.; Abbas, Z.K.; Al-Harbi, N.A.; Abd Algafar, A.; Almuhayawi, M.S.; et al. Effect of Elevated CO2 on Biomolecules’ Accumulation in Caraway (Carum carvi L.) Plants at Different Developmental Stages. Plants 2021, 10, 2434. https://doi.org/10.3390/plants10112434
AbdElgawad H, Okla MK, Al-amri SS, AL-Hashimi A, AL-Qahtani WH, Al-Qahtani SM, Abbas ZK, Al-Harbi NA, Abd Algafar A, Almuhayawi MS, et al. Effect of Elevated CO2 on Biomolecules’ Accumulation in Caraway (Carum carvi L.) Plants at Different Developmental Stages. Plants. 2021; 10(11):2434. https://doi.org/10.3390/plants10112434
Chicago/Turabian StyleAbdElgawad, Hamada, Mohammad K. Okla, Saud S. Al-amri, Abdulrahman AL-Hashimi, Wahida H. AL-Qahtani, Salem Mesfir Al-Qahtani, Zahid Khorshid Abbas, Nadi Awad Al-Harbi, Ayman Abd Algafar, Mohammed S. Almuhayawi, and et al. 2021. "Effect of Elevated CO2 on Biomolecules’ Accumulation in Caraway (Carum carvi L.) Plants at Different Developmental Stages" Plants 10, no. 11: 2434. https://doi.org/10.3390/plants10112434