
Analysis of PRX Gene Family and Its Function on Cell Lignification in Pears (Pyrus bretschneideri)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of Class III Peroxidase Genes (PRXs) in Pear
2.2. Phylogenetic Analysis, Gene Structure and Conserved Motif Analysis
2.3. Chromosomal Location and Synteny Analysis
2.4. Calculating Ka and Ks of the PbPRX Gene Family
2.5. Gene Ontology Enrichment Analysis
2.6. Genome-Wide Expression Analysis of PRX Gene Family
2.7. RNA Extraction and First-Strand cDNA Synthesis
2.8. Quantitative Real-Time PCR Analysis
2.9. Determination of Stone Cells and Lignin Content
3. Results
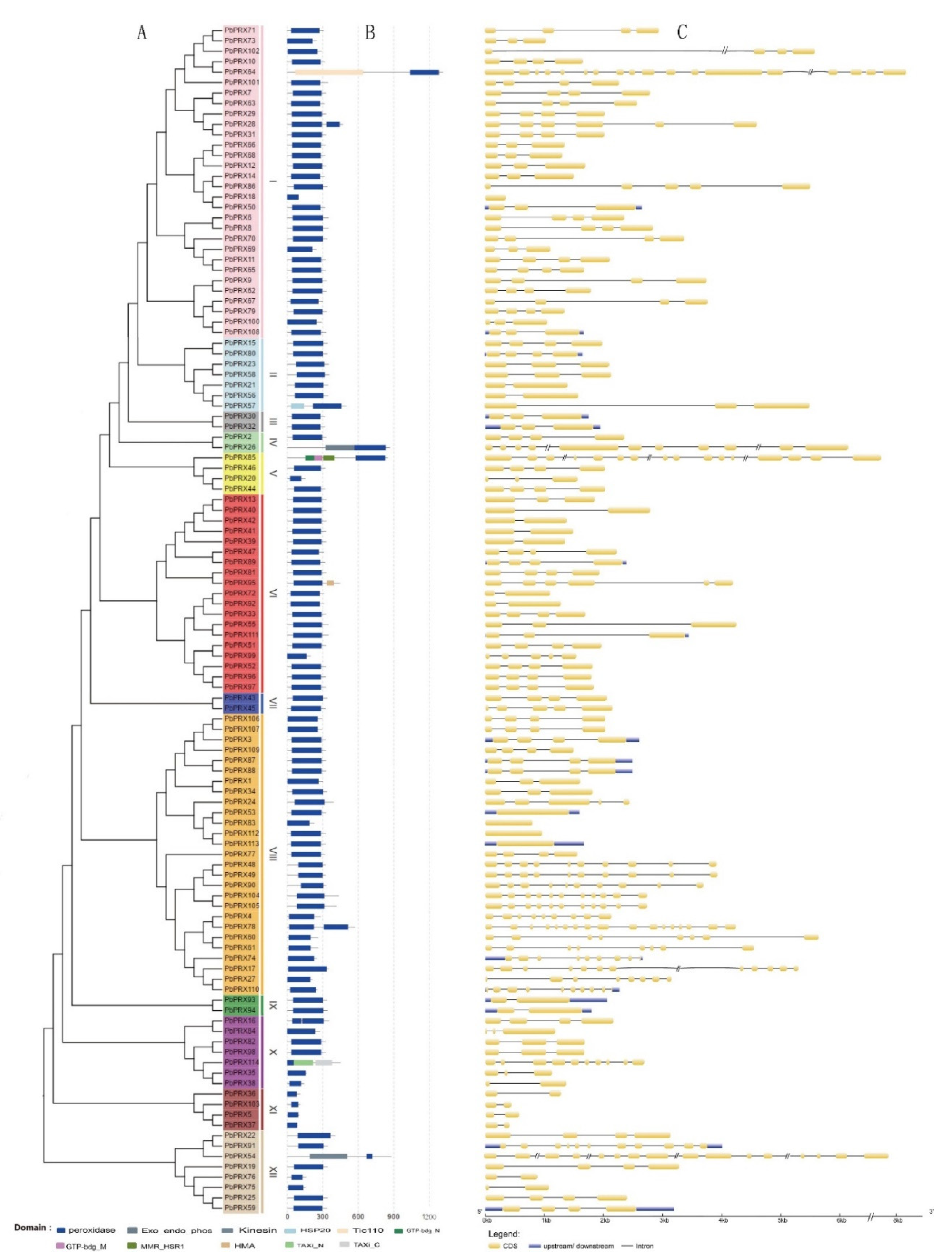
3.1. Identification and Construction of Phylogenetic Tree of Class III Peroxidase Gene Family (PRXs) in Pear
3.2. Analysis of Conserved Motifs and Domain
3.3. Chromosomal Locations, Gene Duplication, and Collinearity Analyzes
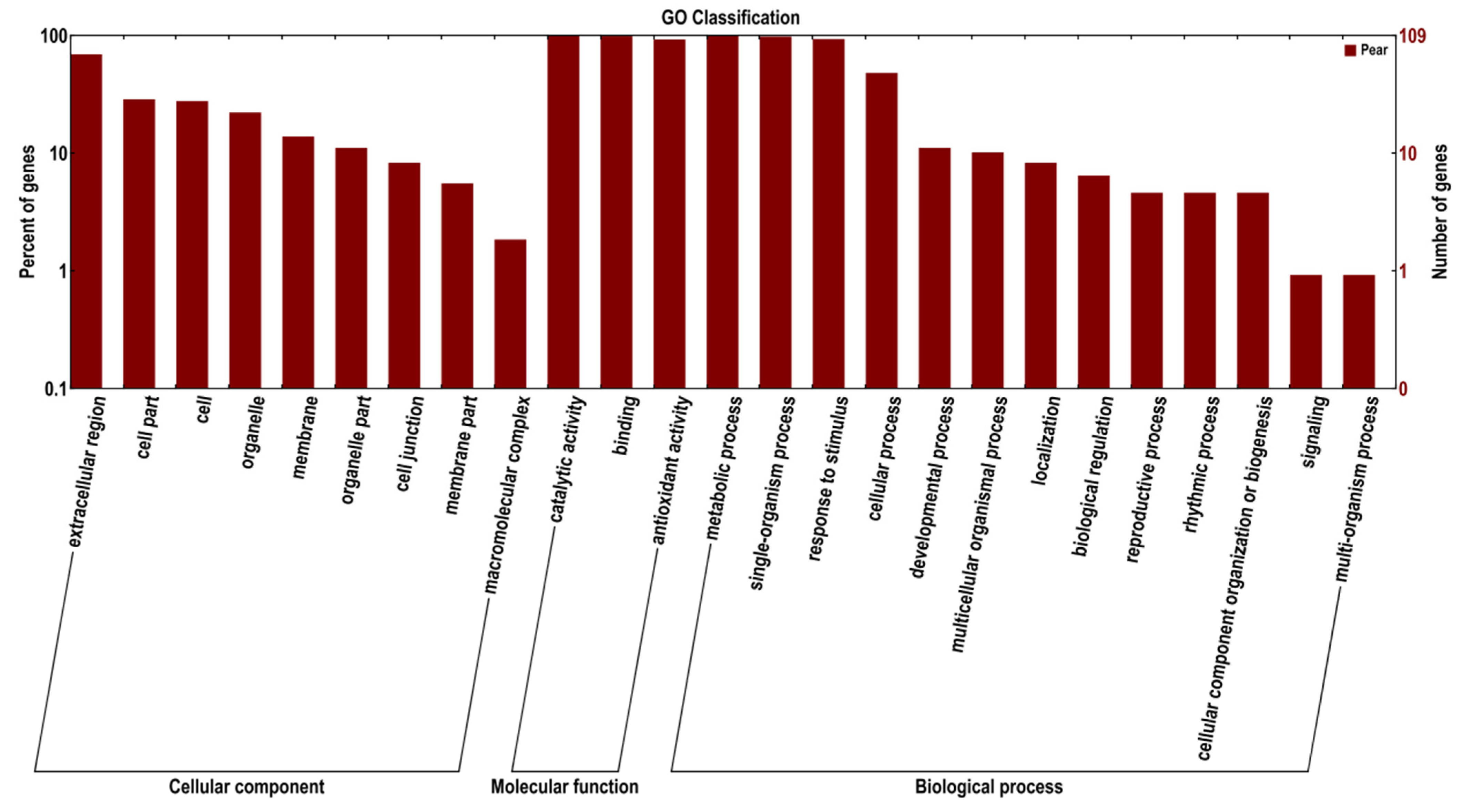
3.4. Functional Annotation with Gene Ontology
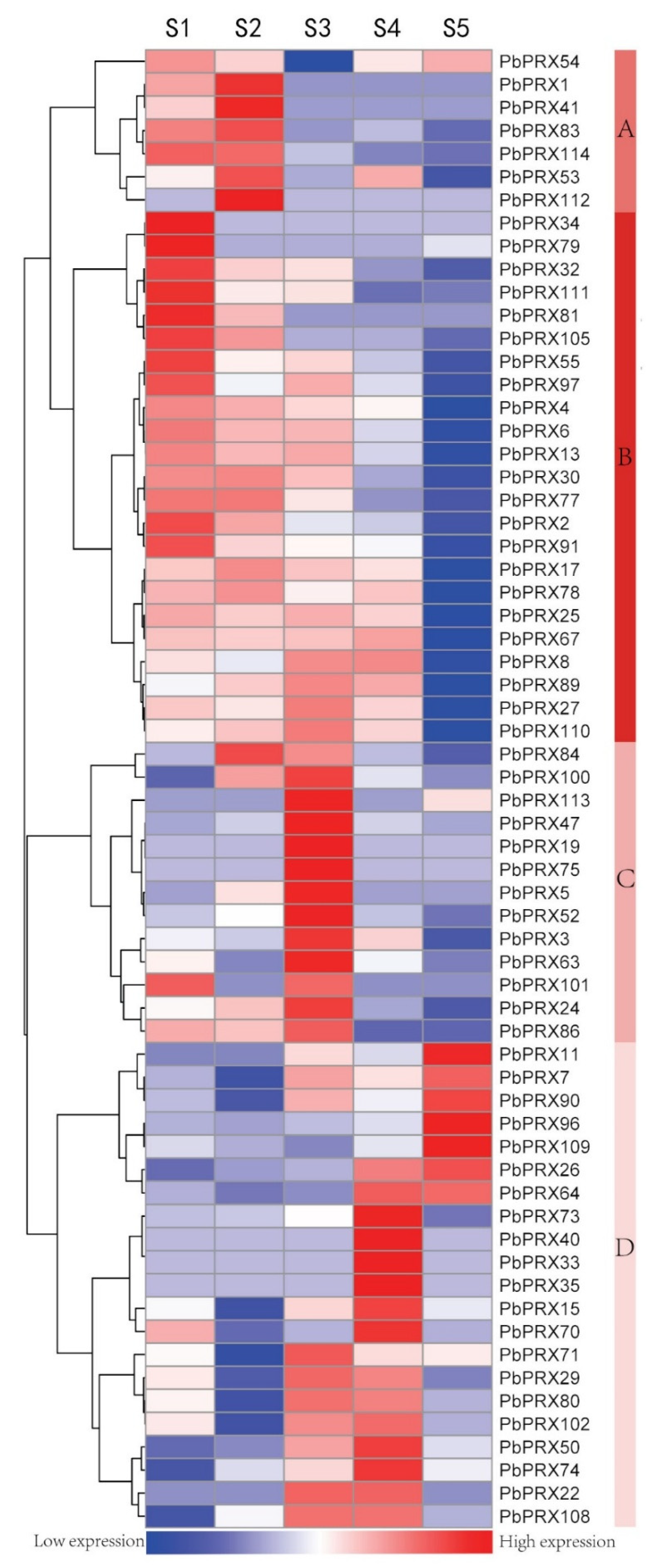
3.5. Expression of the PRX Gene Family in Pears
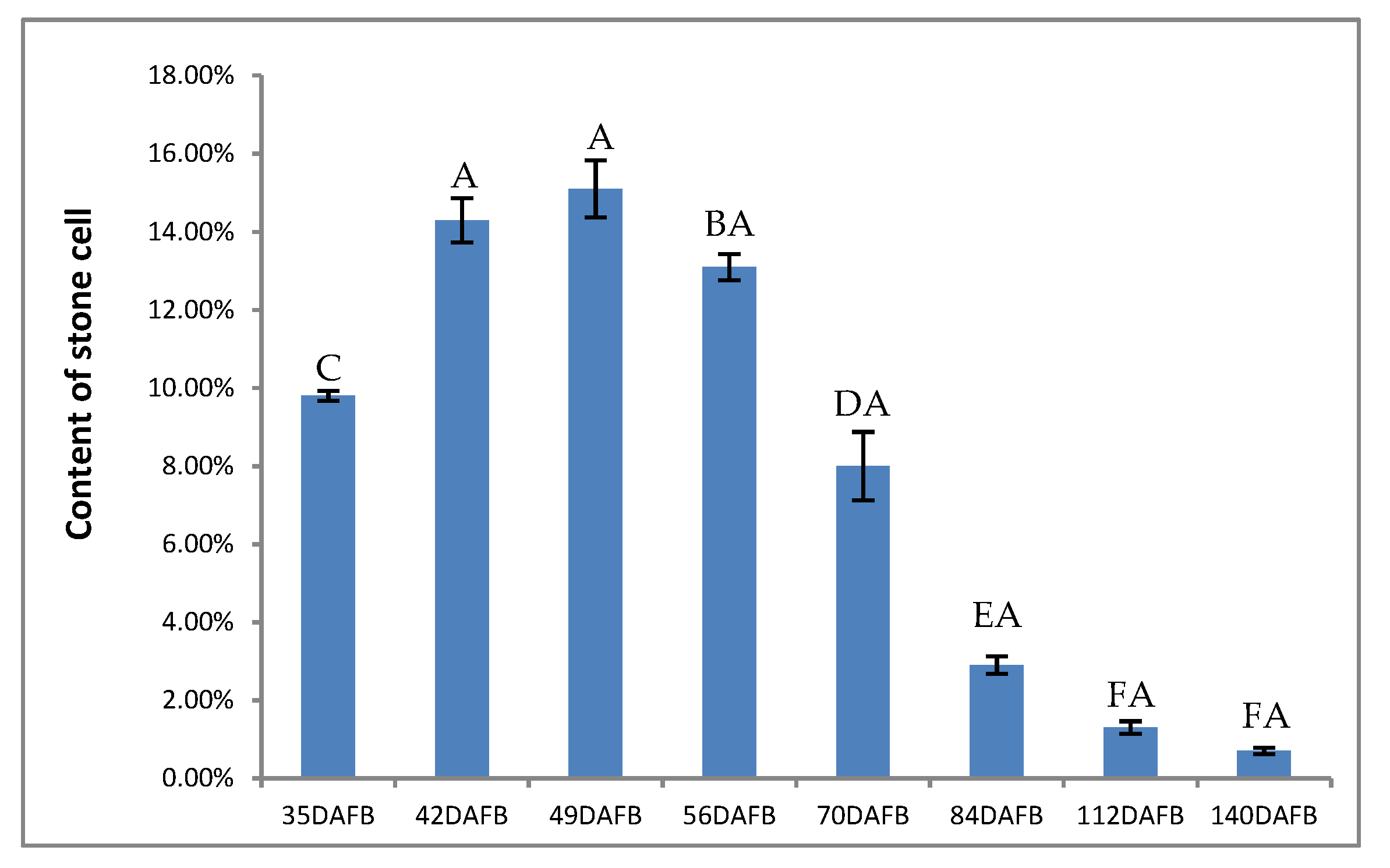
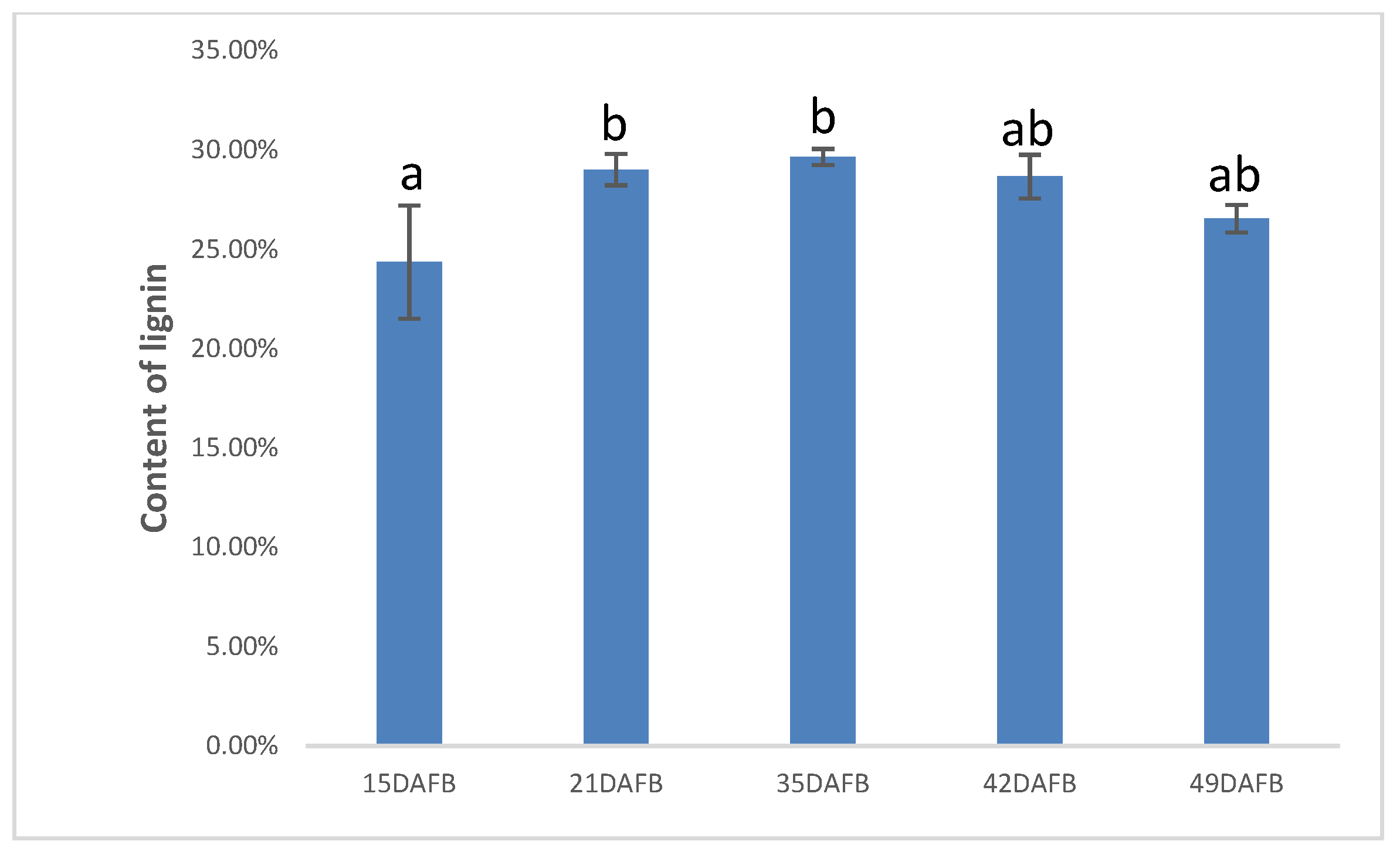
3.6. Stone Cells and Lignin Content of Pear Fruit during Pear Fruit Development
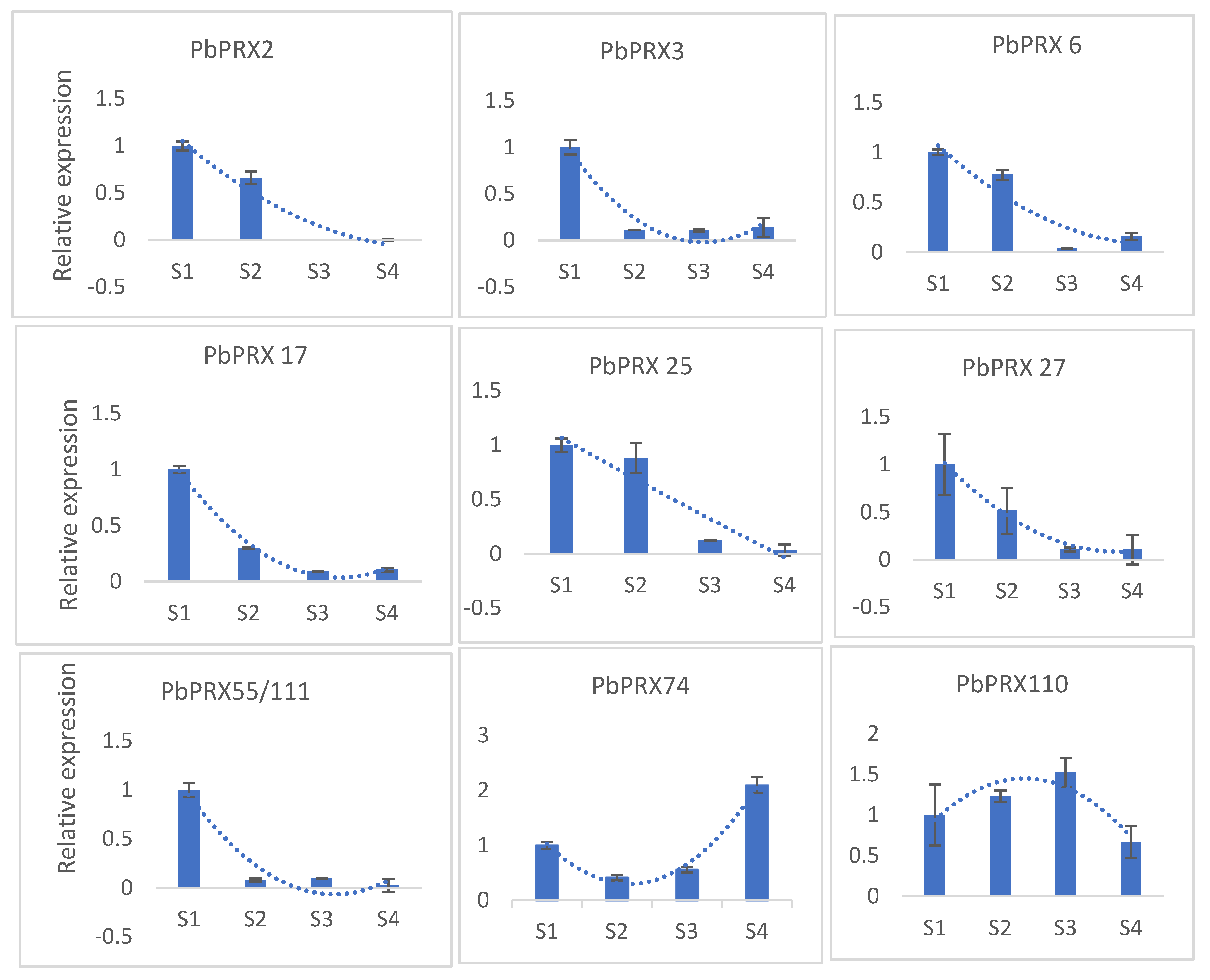
3.7. Verification of Gene Expression by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cosio, C.; Dunand, C. Specific functions of individual class III peroxidase genes. J. Exp. Bot. 2009, 60, 391–408. [Google Scholar] [CrossRef]
- Bindschedler, L.V.; Dewdney, J.; Blee, K.A.; Stone, J.M.; Asai, T.; Plotnikov, J.; Denoux, C.; Hayes, T.; Gerrish, C.; Davies, D.R.; et al. Peroxidase-dependent apoplastic oxidative burst in Arabidopsis required for pathogen resistance. Plant J. 2006, 47, 851–863. [Google Scholar] [CrossRef] [Green Version]
- Daudi, A.Z.; Cheng, J.A.; Brien, O.; Mammarella, N.; Khan, S.; Ausubel, F.M.; Bolwell, G.P. The apoplastic oxidative burst peroxidase in Arabidopsis is a major component of pattern-triggered immunity. Plant Cell 2012, 24, 275–287. [Google Scholar] [CrossRef] [Green Version]
- Herrero, J.; Fernández-Pérez, F.; Yebra, T.; Novo-Uzal, E.; Pomar, F.; Pedreo, M.; Cuello, J.; Guéra, A.; Esteban-Carrasco, A.; Zapata, J.M. Bioinformatic and functional characterization of the basic peroxidase 72 from Arabidopsis thaliana involved in lignin biosynthesis. Planta 2013, 237, 1599–1612. [Google Scholar] [CrossRef] [PubMed]
- Welinder, K.G. Plant peroxidases: Structure–function relationships. In Plant Peroxidases; University of Geneva: Geneva, The Switzerland, 1992; pp. 1–24. [Google Scholar]
- Welinder, K.G. Superfamily of plant, fungal and bacterial peroxidases. Curr. Opin. Struct. Biol. 1992, 2, 388–393. [Google Scholar] [CrossRef]
- Passardi, F.; Longet, D.; Penel, C.; Dunand, C. The class III peroxidase multigenic family in rice and its evolution in land plants. Phytochemistry 2004, 65, 1879–1893. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Liu, Y.; Qian, T.; Qi, L.; Wang, X.; Zeng, Q. Subcellular relocalization and positive selection play key roles in the retention of duplicate genes of populus class III peroxidase family. Plant Cell 2014, 26, 2404–2419. [Google Scholar] [CrossRef] [Green Version]
- Tognolli, M.; Penel, C.; Greppin, H.; Simon, P. Analysis and expression of the class III peroxidase large gene family in Arabidopsis thaliana. Gene 2002, 288, 129–138. [Google Scholar] [CrossRef]
- Tao, S.T. Characterization of Sclereid Structure and Composition and Cloning of Sclereid Related Enzyme Genes in Pear (Pyrus) Fruit. Ph.D. Thesis, Nanjing Agricultural University, Nanjing, China, 2009. [Google Scholar]
- Tao, S.T.; Khanizadeh, S.; Zhang, H.; Zhang, S.L. Anatomy, ultrastructure and lignin distribution of stone cells in two Pyrus species. Plant Sci. 2009, 176, 413–419. [Google Scholar] [CrossRef]
- Van de Peer, Y.; Meyer, A. Large-scale gene and ancient genome duplications. Evol. Genome 2005, 9, 329–368. [Google Scholar]
- Fawcett, J.A.; Maereand, S.; Van de Peer, Y. Plants with double genomes might have had a better chance to survive the Cretaceous-Tertiary extinction event. Proc. Natl. Acad. Sci. USA 2009, 106, 5737–5742. [Google Scholar] [CrossRef] [Green Version]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A. The genome of the domesticated apple (Malus × domestica Borkh). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Taylor, J.S.; Raes, J. Duplication and divergence: The evolution of new genes and old ideas. Annu. Rev. Genet. 2004, 38, 615–643. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Wang, J.; Lin, W.; Song, G.; Li, H.; Zhou, J.; Ni, P.; Dong, W. The Genomes of Oryza sativa: A History of Duplications. PLoS Biol. 2005, 3, 266–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharoni, A.M.; Nuruzzaman, M.; Satoh, K.; Shimizu, T.; Kondoh, H.; Sasaya, T.; Choi, I.; Omura, T.; Kikuchi, S. Gene Structures, Classification and Expression Models of the AP2/EREBP Transcription Factor Family in Rice. Plant Cell Physiol. 2011, 2, 344–360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.X.; Cai, M.J.; Zhang, X.B.; Li, Y.R.; Zhang, J.H.; Zhao, H.L.; Kong, F.; Zheng, Y.L.; Qiu, F.Z. Genome-Wide Identification, Evolution and Expression Analysis of mTERFGene Family in Maize. PLoS ONE 2014, 9, 1–18. [Google Scholar]
- Wu, J.; Wang, Z.W.; Shi, Z.B.; Zhang, S.; Ming, R.; Zhu, S.L.; Khan, M.A.; Tao, S.T.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Bateman, A.; Clements, J.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Heger, A.; Hetherington, K.; Holm, L.; Mistry, J.; et al. Pfam: The protein families database. Nucleic Acids Res. 2014, 42, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Doerks, T.; Bork, P. SMART 7: Recent updates to the protein domain annotation resource. Nucleic Acids Res. 2012, 40, 302–305. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Jin, J.; Guo, A.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 8, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Bailey, T.L.; Elkan, C. Unsupervised Learning of Multiple Motifs in Biopolymers Using Expectation Maximization. Mach. Learn. 1995, 21, 51–80. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.H.; Tang, H.; Wang, X.; Paterson, A.H. PGDD: A database of gene andgenome duplication in plants. Nucleic Acids Res. 2013, 41, 1152–1158. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee Th Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of genesynteny and collinearity. Nucleic Acids Res. 2012, 40, 49. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A Toolkit Incorporating Gamma-Series Methods and Sliding Window Strategies. Genom. Proteom. Bioinform. 2010, 1, 77–80. [Google Scholar] [CrossRef] [Green Version]
- Zdobnov, E.M.; Apweiler, R. InterProScan—An integration platform for the signature-recognition methods in InterPro. Bioinformatics 2001, 17, 847–848. [Google Scholar] [CrossRef] [Green Version]
- Harris, M.; Clark, J.; Ireland, A.; Lomax, J.; Ashburner, M.; Foulger, R.; Eilbeck, K.; Lewis, S.; Marshall, B.; Mungall, C.; et al. The gene ontology (GO) database and informatics resource. Annu. Rev. Genet. 2004, 32, 258. [Google Scholar]
- Ye, J.; Fang, L.; Zheng, H.; Zhang, Y.; Chen, J.; Zhang, Z.; Wang, J.; Li, S.; Li, R.; Bolund, L.; et al. WEGO: A web tool for plotting GO annotations. Nucleic Acids Res. 2006, 34, 293–297. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-DDCT Method. Methods 2001, 4, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Syros, T.; Yupsanis, T.; Zafiriadis, H.; Economou, A. Activity and isoforms of peroxidases, lignin and anatomy, during adventitious rooting in cuttings of Ebenus cretica L. J. Plant Physiol. 2004, 161, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Cannon, C.P.; Braunwald, E.; McCabe, C.H.; Rader, D.J.; Rouleau, J.L.; Belder, R.; Joyal, S.V.; Hill, K.A.; Pfeffer, M.A.; Skene, A.M. Intensive versus Moderate Lipid Lowering with Statins after Acute Coronary Syndromes. N. Engl. J. Med. 2004, 350, 1495–1504. [Google Scholar] [CrossRef] [PubMed]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef] [Green Version]
- Rogozin, I.B.; Wolf, Y.I.; Sorokin, A.V.; Mirkin, B.G.; Koonin, E.V. Remarkable Interkingdom Conservation of Intron Positions and Massive, Lineage-Specific Intron Loss and Gain in Eukaryotic Evolution. Curr. Biol. 2003, 17, 1512–1517. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chr(Mbp) z | Stra-Nd y | Start | End | CDS Length(bp) | Genomic Sequence Length(Bp) | Protein Length(aa) | Molecular Weight(da) | Pi x | Exon Number |
|---|---|---|---|---|---|---|---|---|---|---|---|
| PbPRX1 | Pbr032785.1 | Chr1(10.7) | − | 8171107 | 8172424 | 906 | 1318 | 302 | 32,907.3 | 8.13 | 3 |
| PbPRX2 | Pbr040489.1 | Chr2(22.1) | + | 15570372 | 15572304 | 996 | 1933 | 332 | 36,405.6 | 9.33 | 4 |
| PbPRX3 | Pbr035186.1 | Chr2(22.1) | − | 12486774 | 12488911 | 978 | 2138 | 326 | 35,266.2 | 8.82 | 4 |
| PbPRX4 | Pbr023311.1 | Chr2(22.1) | + | 16007519 | 16009267 | 855 | 1749 | 285 | 31,612.2 | 9.04 | 9 |
| PbPRX5 | Pbr003171.1 | Chr2(22.1) | + | 22018927 | 22019388 | 312 | 462 | 104 | 11,184.8 | 7.85 | 2 |
| PbPRX6 | Pbr000691.1 | Chr3(27.4) | + | 19118330 | 19120264 | 1053 | 1935 | 351 | 37,093.4 | 4.48 | 4 |
| PbPRX7 | Pbr013075.1 | Chr3(27.4) | + | 22733308 | 22735595 | 984 | 2288 | 328 | 35,036.7 | 9.29 | 4 |
| PbPRX8 | Pbr000689.1 | Chr3(27.4) | + | 19105701 | 19108029 | 1053 | 2329 | 351 | 37,452.8 | 4.29 | 4 |
| PbPRX9 | Pbr000686.1 | Chr3(27.4) | + | 19061527 | 19064600 | 999 | 3074 | 333 | 35,773.7 | 8.36 | 4 |
| PbPRX10 | Pbr013078.1 | Chr3(27.4) | + | 22708298 | 22709659 | 960 | 1362 | 320 | 34,387.9 | 6.23 | 4 |
| PbPRX11 | Pbr000687.1 | Chr3(27.4) | + | 19071289 | 19073019 | 975 | 1731 | 325 | 34,720.2 | 4.48 | 4 |
| PbPRX12 | Pbr013077.1 | Chr3(27.4) | − | 22711452 | 22712846 | 993 | 1395 | 331 | 35,887.7 | 6.59 | 3 |
| PbPRX13 | Pbr003832.1 | Chr3(27.4) | − | 26454998 | 26456517 | 999 | 1520 | 333 | 35,669.6 | 6.88 | 3 |
| PbPRX14 | Pbr022808.1 | Chr3(27.4) | + | 1626762 | 1627999 | 945 | 1238 | 315 | 34,167.4 | 5.61 | 3 |
| PbPRX15 | Pbr013214.1 | Chr3(27.4) | − | 21696976 | 21698605 | 1026 | 1630 | 342 | 37,353.7 | 4.81 | 4 |
| PbPRX16 | Pbr021747.1 | Chr3(27.4) | − | 12196399 | 12198178 | 1065 | 1780 | 355 | undefined | undefined | 4 |
| PbPRX17 | Pbr033934.1 | Chr3(27.4) | − | 24516702 | 24522508 | 1059 | 5807 | 353 | 38,476.8 | 5.62 | 12 |
| PbPRX18 | Pbr022809.1 | Chr3(27.4) | + | 1639130 | 1639417 | 288 | 288 | 96 | 10,491.1 | 7.78 | 1 |
| PbPRX19 | Pbr032800.1 | Chr4(13.4) | − | 11190141 | 11192827 | 1026 | 2687 | 342 | 38,505.1 | 8.93 | 4 |
| PbPRX20 | Pbr006566.1 | Chr4(13.4) | + | 816311 | 817595 | 462 | 1285 | 154 | 16,924.5 | 9.47 | 3 |
| PbPRX21 | Pbr002542.1 | Chr5(28.4) | + | 22011532 | 22012678 | 1041 | 1147 | 347 | 38,598.6 | 9.05 | 2 |
| PbPRX22 | Pbr041097.1 | Chr5(28.4) | + | 20588048 | 20590614 | 1218 | 2567 | 406 | 44,469.3 | 6.03 | 4 |
| PbPRX23 | Pbr002505.1 | Chr5(28.4) | − | 21659712 | 21661441 | 1059 | 1730 | 353 | 39,178.2 | 5.91 | 3 |
| PbPRX24 | Pbr000438.1 | Chr5(28.4) | + | 25395691 | 25397689 | 1176 | 1999 | 392 | 42,972.1 | 9.22 | 5 |
| PbPRX25 | Pbr000146.1 | Chr5(28.4) | − | 27528334 | 27530297 | 1026 | 1964 | 342 | 38,740.2 | 8.30 | 4 |
| PbPRX26 | Pbr013845.1 | Chr6(23.1) | + | 19145761 | 19152732 | 2607 | 6972 | 869 | 98,120.6 | 6.33 | 11 |
| PbPRX27 | Pbr014180.2 | Chr6(23.1) | + | 8740620 | 8743196 | 648 | 2577 | 216 | 23,389.3 | 5.11 | 8 |
| PbPRX28 | Pbr002948.1 | Chr7(15.3) | − | 12585545 | 12589315 | 1416 | 3771 | 472 | 51,545.7 | 8.99 | 6 |
| PbPRX29 | Pbr010975.1 | Chr7(15.3) | + | 11895741 | 11897397 | 990 | 1657 | 330 | 35,665.4 | 9.65 | 4 |
| PbPRX30 | Pbr002950.1 | Chr7(15.3) | + | 12603529 | 12604962 | 957 | 1434 | 319 | 34,580.5 | 9.31 | 3 |
| PbPRX31 | Pbr002956.1 | Chr7(15.3) | + | 12644802 | 12646457 | 990 | 1656 | 330 | 35,812.7 | 9.73 | 4 |
| PbPRX32 | Pbr010973.1 | Chr7(15.3) | − | 11880070 | 11881667 | 957 | 1598 | 319 | 34,607.6 | 9.43 | 3 |
| PbPRX33 | Pbr040033.1 | Chr7(15.3) | − | 14720194 | 14721588 | 987 | 1395 | 329 | 35,549.3 | 9.02 | 4 |
| PbPRX34 | Pbr013905.1 | Chr7(15.3) | − | 12981008 | 12982502 | 1011 | 1495 | 337 | 37,239.5 | 8.72 | 3 |
| PbPRX35 | Pbr010977.1 | Chr7(15.3) | + | 11900730 | 11901663 | 486 | 934 | 162 | 18,183.7 | 9.11 | 3 |
| PbPRX36 | Pbr002947.1 | Chr7(15.3) | − | 12582679 | 12583729 | 330 | 1051 | 110 | 12,014.7 | 5.34 | 2 |
| PbPRX37 | Pbr002957.1 | Chr7(15.3) | + | 12650388 | 12650724 | 252 | 337 | 84 | 9,017.3 | 6.54 | 2 |
| PbPRX38 | Pbr010976.1 | Chr7(15.3) | + | 11897792 | 11898914 | 429 | 1123 | 143 | 16,177.7 | 8.44 | 2 |
| PbPRX39 | Pbr026505.1 | Chr8(17.1) | − | 4426466 | 4427581 | 996 | 1116 | 332 | 35,928.0 | 8.79 | 2 |
| PbPRX40 | Pbr026503.1 | Chr8(17.1) | + | 4440333 | 4442618 | 996 | 2286 | 332 | 36,048.0 | 8.62 | 2 |
| PbPRX41 | Pbr026502.1 | Chr8(17.1) | + | 4444100 | 4445303 | 984 | 1204 | 328 | 35,583.5 | 8.57 | 2 |
| PbPRX42 | Pbr026504.1 | Chr8(17.1) | − | 4434098 | 4435213 | 996 | 1116 | 332 | 36,043.2 | 8.89 | 2 |
| PbPRX43 | Pbr036549.1 | Chr8(17.1) | − | 16020359 | 16022020 | 1014 | 1662 | 338 | 36,117.8 | 5.44 | 4 |
| PbPRX44 | Pbr006119.1 | Chr8(17.1) | − | 15274439 | 15276078 | 966 | 1640 | 322 | 34,724.7 | 8.41 | 4 |
| PbPRX45 | Pbr036474.1 | Chr8(17.1) | + | 16467636 | 16469367 | 972 | 1732 | 324 | 34,752.3 | 5.32 | 5 |
| PbPRX46 | Pbr006117.1 | Chr8(17.1) | − | 15269333 | 15270972 | 966 | 1640 | 322 | 34,697.6 | 8.11 | 4 |
| PbPRX47 | Pbr004299.1 | Chr8(17.1) | + | 6081866 | 6083665 | 933 | 1800 | 311 | 33,396.9 | 4.82 | 4 |
| PbPRX48 | Pbr020588.1 | Chr8(17.1) | − | 9421357 | 9424517 | 972 | 3161 | 324 | 34,409.4 | 5.88 | 10 |
| PbPRX49 | Pbr020590.1 | Chr8(17.1) | + | 9511517 | 9514687 | 972 | 3171 | 324 | 34,409.4 | 5.88 | 10 |
| PbPRX50 | Pbr005400.1 | Chr9(22.4) | + | 5466505 | 5468645 | 957 | 2141 | 319 | 34,371.0 | 8.71 | 3 |
| PbPRX51 | Pbr018082.1 | Chr9(22.4) | + | 16626484 | 16628070 | 984 | 1587 | 328 | 35,481.6 | 9.08 | 4 |
| PbPRX52 | Pbr018080.1 | Chr9(22.4) | + | 16633205 | 16634674 | 981 | 1470 | 327 | 35,595.7 | 9.01 | 4 |
| PbPRX53 | Pbr026235.1 | Chr9(22.4) | + | 20559304 | 20560595 | 984 | 1292 | 328 | 35,943.9 | 8.58 | 1 |
| PbPRX54 | Pbr019188.1 | Chr9(22.4) | + | 10558578 | 10565436 | 2637 | 6859 | 879 | 97,910.3 | 5.85 | 16 |
| PbPRX55 | Pbr027164.1 | Chr10(26.2) | + | 24310314 | 24313743 | 1053 | 3430 | 351 | 38,056.8 | 5.20 | 3 |
| PbPRX56 | Pbr010270.1 | Chr10(26.2) | − | 1153171 | 1154446 | 1041 | 1276 | 347 | 38,616.7 | 9.26 | 2 |
| PbPRX57 | Pbr010258.1 | Chr10(26.2) | + | 1043826 | 1048256 | 1497 | 4431 | 499 | 56,140.4 | 8.20 | 3 |
| PbPRX58 | Pbr010213.1 | Chr10(26.2) | − | 612484 | 614210 | 1071 | 1727 | 357 | 39,612.6 | 5.44 | 4 |
| PbPRX59 | Pbr031894.1 | Chr10(26.2) | − | 5317405 | 5319983 | 1011 | 2579 | 337 | 38,342.9 | 8.30 | 4 |
| PbPRX60 | Pbr020725.1 | Chr10(26.2) | − | 17517628 | 17522171 | 789 | 4544 | 263 | 28,676.3 | 6.51 | 8 |
| PbPRX61 | Pbr020734.1 | Chr10(26.2) | + | 17377042 | 17381585 | 789 | 4544 | 263 | 28,676.3 | 6.51 | 8 |
| PbPRX62 | Pbr003308.1 | Chr11(30.3) | − | 22280347 | 22282143 | 999 | 1797 | 333 | 35,706.4 | 7.53 | 4 |
| PbPRX63 | Pbr011562.1 | Chr11(30.3) | + | 25372571 | 25375151 | 945 | 2581 | 315 | 33,541.9 | 9.35 | 4 |
| PbPRX64 | Pbr011557.1 | Chr11(30.3) | + | 25338613 | 25347124 | 3945 | 8512 | 1315 | 143,415.6 | 5.78 | 18 |
| PbPRX65 | Pbr003309.1 | Chr11(30.3) | − | 22273464 | 22275146 | 975 | 1683 | 325 | 347,76.3 | 4.57 | 4 |
| PbPRX66 | Pbr011560.1 | Chr11(30.3) | + | 25362475 | 25363817 | 969 | 1343 | 323 | 35,081.3 | 9.47 | 3 |
| PbPRX67 | Pbr022326.1 | Chr11(30.3) | − | 6078338 | 6082106 | 906 | 3769 | 302 | 32,772.3 | 7.54 | 4 |
| PbPRX68 | Pbr011559.1 | Chr11(30.3) | + | 25356556 | 25357860 | 963 | 1305 | 321 | 34,899.8 | 8.97 | 3 |
| PbPRX69 | Pbr003310.1 | Chr11(30.3) | − | 22266804 | 22267913 | 744 | 1110 | 248 | 26,540.9 | 4.38 | 3 |
| PbPRX70 | Pbr035815.1 | Chr12(22.8) | + | 16715336 | 16718714 | 1017 | 3379 | 339 | 35,696.5 | 5.10 | 4 |
| PbPRX71 | Pbr014607.1 | Chr12(22.8) | − | 4710343 | 4713291 | 927 | 2949 | 309 | 32,685.0 | 8.91 | 4 |
| PbPRX72 | Pbr026058.1 | Chr12(22.8) | + | 3759751 | 3760860 | 936 | 1110 | 312 | 33,748.3 | 8.09 | 2 |
| PbPRX73 | Pbr014605.1 | Chr12(22.8) | − | 4732591 | 4733625 | 750 | 1035 | 250 | 26,220.3 | 8.94 | 3 |
| PbPRX74 | Pbr008291.1 | Chr12(22.8) | − | 9391476 | 9394142 | 753 | 2667 | 251 | 27,632.2 | 5.30 | 9 |
| PbPRX75 | Pbr008320.1 | Chr12(22.8) | + | 9106502 | 9107578 | 471 | 1077 | 157 | 17,355.4 | 5.82 | 2 |
| PbPRX76 | Pbr035513.1 | Chr12(22.8) | + | 22606333 | 22607214 | 483 | 882 | 161 | 17,774.6 | 8.75 | 2 |
| PbPRX77 | Pbr034800.1 | Chr13(15.1) | + | 14344175 | 14345739 | 957 | 1565 | 319 | 34,329.0 | 9.37 | 4 |
| PbPRX78 | Pbr034821.1 | Chr13(15.1) | + | 14483716 | 14487956 | 1722 | 4241 | 574 | 63,176.3 | 6.96 | 17 |
| PbPRX79 | Pbr015016.1 | Chr13(15.1) | − | 8716264 | 8717618 | 1002 | 1355 | 334 | 35,742.2 | 4.42 | 4 |
| PbPRX80 | Pbr030045.1 | Chr13(15.1) | − | 4024846 | 4026504 | 1017 | 1659 | 339 | 36,936.2 | 4.81 | 4 |
| PbPRX81 | Pbr014793.1 | Chr13(15.1) | − | 11876947 | 11878622 | 996 | 1676 | 332 | 36,098.2 | 7.03 | 4 |
| PbPRX82 | Pbr015032.1 | Chr13(15.1) | − | 8833969 | 8835416 | 975 | 1448 | 325 | 35,052.1 | 5.49 | 3 |
| PbPRX83 | Pbr039193.1 | Chr13(15.1) | − | 3561750 | 3562436 | 687 | 687 | 229 | 25,105.6 | 8.93 | 1 |
| PbPRX84 | Pbr016853.1 | Chr14(20.3) | − | 3626045 | 3627070 | 834 | 1026 | 278 | 29,540.2 | 6.41 | 3 |
| PbPRX85 | Pbr005912.1 | Chr15(43.6) | + | 2978373 | 2985696 | 2595 | 7324 | 865 | 93,865.2 | 7.87 | 16 |
| PbPRX86 | Pbr007872.1 | Chr15(43.6) | − | 20261904 | 20266643 | 1020 | 4740 | 340 | 37,262.1 | 5.36 | 5 |
| PbPRX87 | Pbr002672.1 | Chr15(43.6) | − | 1214104 | 1216253 | 984 | 2150 | 328 | 36,549.0 | 9.52 | 4 |
| PbPRX88 | Pbr009308.1 | Chr15(43.6) | − | 3677275 | 3679424 | 984 | 2150 | 328 | 36,549.0 | 9.52 | 4 |
| PbPRX89 | Pbr010632.1 | Chr15(43.6) | + | 12931761 | 12933817 | 963 | 2057 | 321 | 34,691.7 | 7.53 | 4 |
| PbPRX90 | Pbr027845.1 | Chr15(43.6) | − | 9981997 | 9985179 | 993 | 3183 | 331 | 35,362.0 | 5.40 | 10 |
| PbPRX91 | Pbr042913.1 | Chr15(43.6) | − | 36603917 | 36607371 | 1035 | 3455 | 345 | 37,741.8 | 6.26 | 11 |
| PbPRX92 | Pbr006005.1 | Chr16(20.6) | − | 10236903 | 10238012 | 936 | 1110 | 312 | 33,748.3 | 8.09 | 2 |
| PbPRX93 | Pbr036153.1 | Chr16(20.6) | + | 62244 | 64019 | 1011 | 1776 | 337 | 36,758.4 | 8.03 | 2 |
| PbPRX94 | Pbr036152.1 | Chr16(20.6) | + | 59021 | 60571 | 1023 | 1551 | 341 | 37,562.1 | 8.28 | 2 |
| PbPRX95 | Pbr011189.1 | Chr16(20.6) | − | 6703300 | 6706908 | 1341 | 3609 | 447 | 49,287.0 | 9.10 | 5 |
| PbPRX96 | Pbr034488.2 | Chr17(25.3) | − | 22537048 | 22538599 | 978 | 1552 | 326 | 35,455.4 | 8.78 | 4 |
| PbPRX97 | Pbr034480.1 | Chr17(25.3) | − | 22583700 | 22585282 | 981 | 1583 | 327 | 35,426.4 | 9.01 | 4 |
| PbPRX98 | Pbr026772.1 | Chr17(25.3) | − | 7814591 | 7816038 | 975 | 1448 | 325 | 35,052.1 | 5.49 | 3 |
| PbPRX99 | Pbr034479.1 | Chr17(25.3) | − | 22587987 | 22589313 | 600 | 1327 | 200 | 21,632.4 | 9.41 | 5 |
| PbPRX100 | Pbr006343.1 | scaffold132.0.1(0.8) | − | 817092 | 818160 | 876 | 1069 | 292 | 32,297.8 | 5.06 | 3 |
| PbPRX101 | Pbr015968.1 | scaffold235.0(0.6) | − | 499783 | 502085 | 1035 | 2303 | 345 | 37,911.4 | 5.25 | 4 |
| PbPRX102 | Pbr015965.1 | scaffold235.0(0.6) | + | 472592 | 481134 | 876 | 8543 | 292 | 31,040.9 | 7.55 | 4 |
| PbPRX103 | Pbr015969.1 | scaffold235.0(0.6) | − | 513208 | 513655 | 330 | 448 | 110 | 11,818.5 | 5.10 | 2 |
| PbPRX104 | Pbr027136.1 | scaffold440.0(0.4) | − | 95386 | 98157 | 1308 | 2772 | 436 | 47,426.7 | 7.70 | 11 |
| PbPRX105 | Pbr027137.1 | scaffold440.0(0.4) | + | 194284 | 197055 | 1245 | 2772 | 415 | 45,080.0 | 7.71 | 11 |
| PbPRX106 | Pbr037664.1 | scaffold770.0(0.2) | − | 44241 | 46306 | 888 | 2066 | 296 | 31,950.3 | 7.53 | 4 |
| PbPRX107 | Pbr037665.1 | scaffold770.0(0.2) | + | 152615 | 154680 | 888 | 2066 | 296 | 31,950.3 | 7.53 | 4 |
| PbPRX108 | Pbr037526.1 | scaffold766.0(0.2) | − | 279 | 1961 | 990 | 1683 | 330 | 36,383.7 | 5.91 | 3 |
| PbPRX109 | Pbr041827.1 | scaffold955.0(0.1) | − | 120167 | 121688 | 981 | 1522 | 327 | 35,732.6 | 8.82 | 4 |
| PbPRX110 | Pbr000988.3 | scaffold1008.0(0.1) | + | 122239 | 124533 | 768 | 2295 | 256 | 27,802.6 | 5.82 | 9 |
| PbPRX111 | Pbr007903.1 | scaffold1455.0(0.04) | + | 21962 | 25441 | 1053 | 3480 | 351 | 38,098.9 | 5.20 | 3 |
| PbPRX112 | Pbr007909.1 | scaffold1459.0(0.04) | + | 4474 | 5451 | 978 | 978 | 326 | 36,027.2 | 8.33 | 1 |
| PbPRX113 | Pbr007908.1 | scaffold1459.0(0.04) | + | 1560 | 3254 | 978 | 1695 | 326 | 35,817.9 | 6.00 | 1 |
| PbPRX114 | Pbr008699.1 | scaffold1534.0(0.03) | − | 10892 | 13612 | 1353 | 2721 | 451 | 49,351.1 | 5.63 | 10 |
| Motif | Width | Seqs z | E-Value | Conserved Amino Acid Sequences |
|---|---|---|---|---|
| 1 | 21 | 78 | 9.8 × 10−1252 | LLRMHFHDCFVQGCDASVLLD |
| 2 | 21 | 96 | 1.0 × 10−895 | HTIGQARCTTFRARIYNETNI |
| 3 | 15 | 95 | 2.0 × 10−883 | CPGVVSCADILAIAA |
| 4 | 15 | 101 | 8.0 × 10−729 | TGGPTWKVPTGRRDG |
| 5 | 15 | 103 | 1.7 × 10−648 | FFQQFAKSMVKMGNI |
| 6 | 32 | 91 | 1.4 × 10−1208 | FDNSYFKNLIQKKGLLHSDQQLFNGGSTDSIV |
| 7 | 21 | 81 | 5.5 × 10−730 | PNNNSLRGFEVVDKIKSQVEK |
| 8 | 15 | 91 | 5.5 × 10−632 | TGSNGEIRKNCRVVN |
| 9 | 21 | 79 | 7.7 × 10−611 | GFYSRTCPTAESIVKQTVQTH |
| 10 | 15 | 100 | 1.9 × 10−581 | SHGLSQTDMVALSGA |
| 11 | 21 | 101 | 1.4 × 10−382 | ANETINLPAPTFNVSQLIQSF |
| 12 | 15 | 88 | 4.7 × 10−251 | NKTYATQLQQMCPKN |
| 13 | 57 | 8 | 2.1 × 10−180 | KTKTGGPFGTMRCPAEQAHGANNGLDIAVRLLEPIKQQFPILSYA DFYQLAGVVAVE |
| 14 | 29 | 12 | 6.5 × 10−132 | RRDLRALIYSKNCAPIMLRIAWHDAGTYD |
| 15 | 11 | 59 | 1.9 × 10−132 | DTPNFTGEKTA |
| 16 | 15 | 34 | 8.6 × 10−109 | GGDNNLSPLDVTSPT |
| 17 | 21 | 9 | 3.0 × 10−93 | HTLGRCHKERSGFEGPWTPNP |
| 18 | 29 | 12 | 8.3 × 10−117 | EGLLKLPTDKALLDDPEFRLYVELYAKDE |
| 19 | 21 | 9 | 2.5 × 10−51 | PDNLSLAGDGFDTVIKAKAAV |
| 20 | 6 | 81 | 1.5 × 10−49 | RDSVVL |
| Paralogous Pairs | Ka | Ks | Ka/Ks | Purifying Selection | Duplicate Type |
|---|---|---|---|---|---|
| PbPRX1-PbPRX34 | 0.06607 | 0.20824 | 0.31728 | Yes | Segmental (WGD) |
| PbPRX2-PbPRX85 | 0.36046 | 2.11212 | 0.17067 | Yes | Segmental(WGD) |
| PbPRX4-PbPRX78 | 0.12760 | 0.36480 | 0.34977 | Yes | Segmental(WGD) |
| PbPRX5-PbPRX89 | 0.29346 | 0.66614 | 0.44053 | Yes | Segmental(WGD) |
| PbPRX7-PbPRX29 | 0.21605 | 1.82632 | 0.11830 | Yes | Segmental(WGD) |
| PbPRX9-PbPRX62 | 0.06251 | 0.14092 | 0.44361 | Yes | Segmental(WGD) |
| PbPRX11-PbPRX69 | 0.03789 | 0.13969 | 0.27123 | Yes | Segmental(WGD) |
| PbPRX12-PbPRX68 | 0.21650 | 1.21569 | 0.17809 | Yes | Segmental(WGD) |
| PbPRX14-PbPRX50 | 0.18371 | 0.66916 | 0.27454 | Yes | Segmental(WGD) |
| PbPRX15-PbPRX80 | 0.04732 | 0.15792 | 0.29963 | Yes | Segmental(WGD) |
| PbPRX17-PbPRX27 | 0.03944 | 0.27059 | 0.14576 | Yes | Segmental(WGD) |
| PbPRX17-PbPRX74 | 0.11130 | 1.33285 | 0.08350 | Yes | Segmental(WGD) |
| PbPRX21-PbPRX56 | 0.03789 | 0.28908 | 0.13109 | Yes | Segmental(WGD) |
| PbPRX21-PbPRX58 | 0.34305 | 2.24266 | 0.15297 | Yes | Segmental(WGD) |
| PbPRX23-PbPRX58 | 0.04549 | 0.09761 | 0.46601 | Yes | Segmental(WGD) |
| PbPRX25-PbPRX59 | 0.00946 | 0.13415 | 0.07055 | Yes | Segmental(WGD) |
| PbPRX28-PbPRX31 | 0.00600 | 0.01078 | 0.55631 | Yes | Segmental(WGD) |
| PbPRX28-PbPRX38 | 0.12609 | 0.18468 | 0.68272 | Yes | Segmental(WGD) |
| PbPRX30-PbPRX32 | 0.00414 | 0.00882 | 0.46968 | Yes | Segmental(WGD) |
| PbPRX36-PbPRX37 | 0.20381 | 0.21673 | 0.94039 | Yes | Segmental(WGD) |
| PbPRX51-PbPRX97 | 0.45986 | 4.06384 | 0.11316 | Yes | Segmental(WGD) |
| PbPRX53-PbPRX83 | 0.38368 | 0.75805 | 0.50614 | Yes | Segmental(WGD) |
| PbPRX56-PbPRX57 | 0.01007 | 0.02564 | 0.39263 | Yes | Segmental(WGD) |
| PbPRX69-PbPRX70 | 0.26718 | 3.83175 | 0.06973 | Yes | Segmental(WGD) |
| PbPRX71-PbPRX102 | 0.15772 | 0.35349 | 0.44617 | Yes | Segmental(WGD) |
| PbPRX81-PbPRX95 | 0.07174 | 0.16084 | 0.44603 | Yes | Segmental(WGD) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Z.; Rui, W.; Yuan, Y.; Song, X.; Liu, X.; Gong, X.; Bao, J.; Zhang, S.; Shahrokh, K.; Tao, S. Analysis of PRX Gene Family and Its Function on Cell Lignification in Pears (Pyrus bretschneideri). Plants 2021, 10, 1874. https://doi.org/10.3390/plants10091874
Xie Z, Rui W, Yuan Y, Song X, Liu X, Gong X, Bao J, Zhang S, Shahrokh K, Tao S. Analysis of PRX Gene Family and Its Function on Cell Lignification in Pears (Pyrus bretschneideri). Plants. 2021; 10(9):1874. https://doi.org/10.3390/plants10091874
Chicago/Turabian StyleXie, Zhihua, Weikang Rui, Yazhou Yuan, Xiaofei Song, Xing Liu, Xin Gong, Jianping Bao, Shaoling Zhang, Khanizadeh Shahrokh, and Shutian Tao. 2021. "Analysis of PRX Gene Family and Its Function on Cell Lignification in Pears (Pyrus bretschneideri)" Plants 10, no. 9: 1874. https://doi.org/10.3390/plants10091874