
Medicago ABI3 Splicing Isoforms Regulate the Expression of Different Gene Clusters to Orchestrate Seed Maturation


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
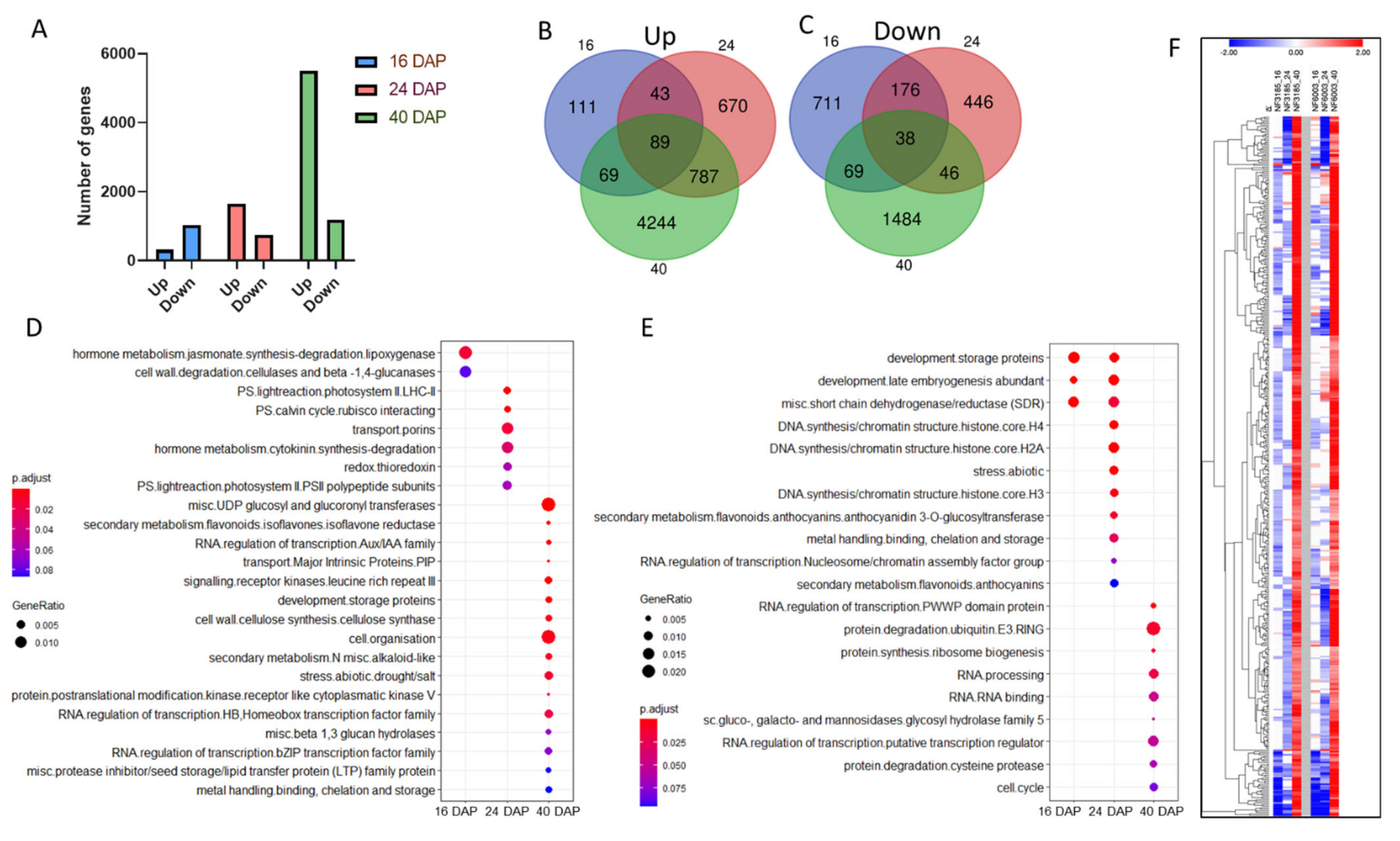
2.1. ABI3 Mutant Transcriptomic Analysis
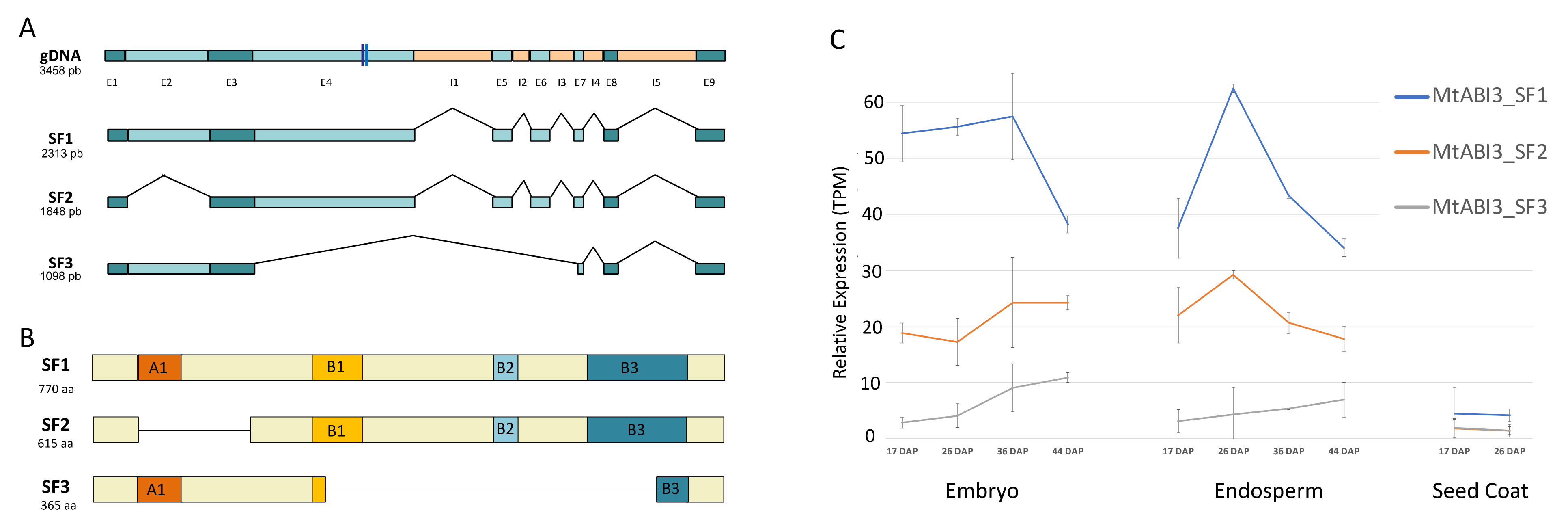
2.2. Identification of Three ABI3 Splicing Forms in Medicago Seed Development
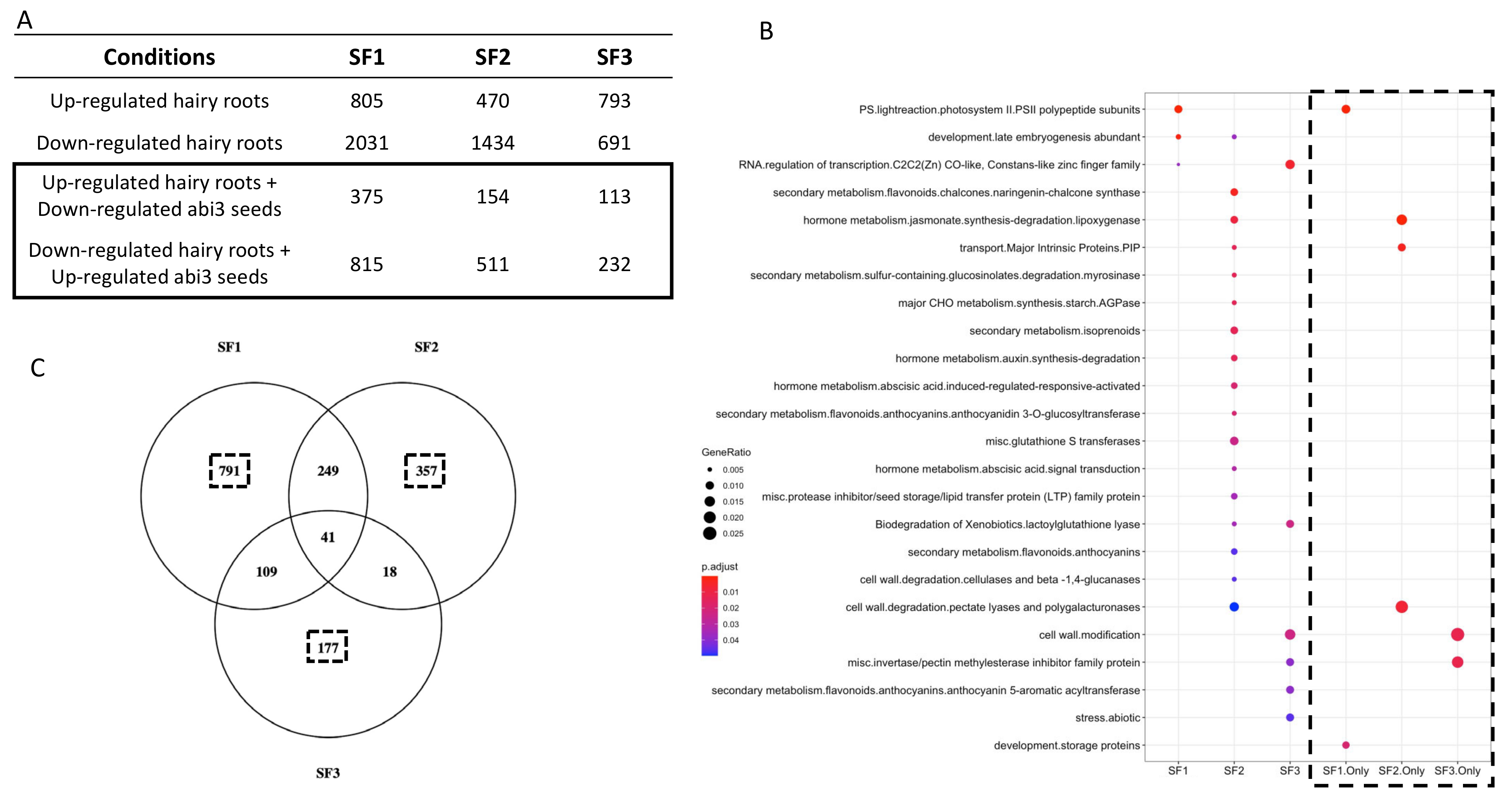
2.3. Ectopic Expression of the Three Different ABI3 Splicing Forms and Potential Associated Molecular Functions
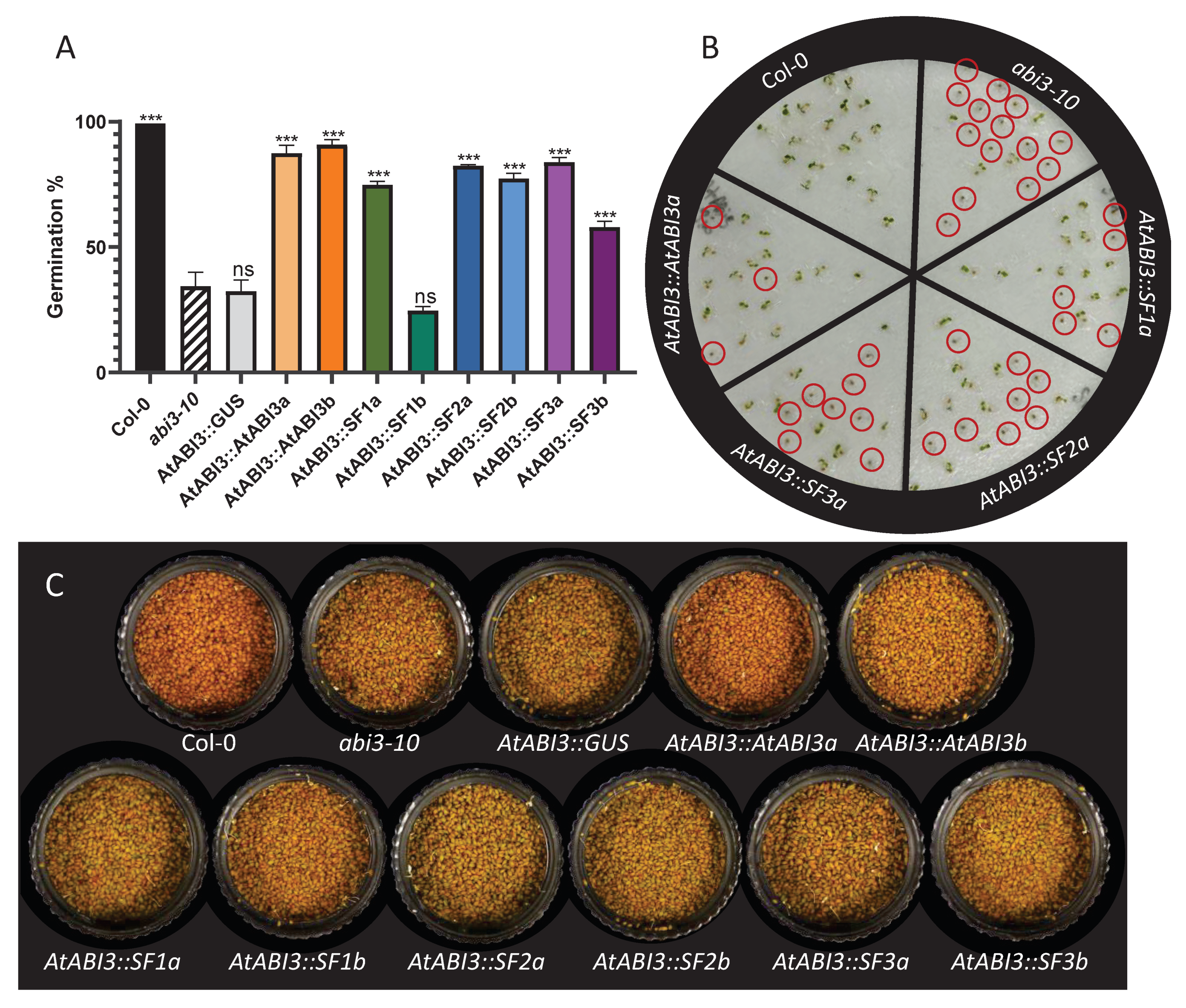
2.4. Complementation of Arabidopsis ABI3 Mutant Lines Using the Three Medicago Transcript Isoforms
3. Discussion
3.1. Alternative Splicing of ABI3 during Medicago Truncatula Seed Development
3.2. Splicing Isoforms of Medicago ABI3 Regulate Different Set of Genes but Restore Seed Viability Phenotype of Mutant Lines
4. Materials and Methods
4.1. Plant Material
4.2. RNA Extraction and Hybridization Arrays
4.3. Cloning and Ectopic Expression in Hairy Root
4.4. Arabidopsis ABI3 Complementation with Different Medicago Splicing Forms
4.5. Expression Profiles of MtABI3 Splicing Isoforms Using Existing RNA-Seq Data
4.6. Gene Annotation and Over-Representation Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Popelka, J.C.; Terryn, N.; Higgins, T.J.V. Gene Technology for Grain Legumes: Can It Contribute to the Food Challenge in Developing Countries? Plant Sci. 2004, 167, 195–206. [Google Scholar] [CrossRef]
- Varshney, R.K.; Dubey, A. Novel Genomic Tools and Modern Genetic and Breeding Approaches for Crop Improvement. J. Plant Biochem. Biotechnol. 2009, 18, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Doyle, J.J.; Luckow, M.A. The Rest of the Iceberg. Legume Diversity and Evolution in a Phylogenetic Context. Plant Physiol. 2003, 131, 900–910. [Google Scholar] [CrossRef] [Green Version]
- Gepts, P.; Beavis, W.D.; Brummer, E.C.; Shoemaker, R.C.; Stalker, H.T.; Weeden, N.E.; Young, N.D. Legumes as a Model Plant Family. Genomics for Food and Feed Report of the Cross-Legume Advances through Genomics Conference. Plant Physiol. 2005, 137, 1228–1235. [Google Scholar] [CrossRef] [Green Version]
- Pecrix, Y.; Staton, S.E.; Sallet, E.; Lelandais-Brière, C.; Moreau, S.; Carrère, S.; Blein, T.; Jardinaud, M.; Latrasse, D.; Zouine, M.; et al. Whole-Genome Landscape of Medicago Truncatula Symbiotic Genes. Nat. Plants 2018, 4, 1017–1025. [Google Scholar] [CrossRef]
- Lotan, T.; Ohto, M.A.; Matsudaira Yee, K.; West, M.A.L.; Lo, R.; Kwong, R.W.; Yamagishi, K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Arabidopsis LEAFY COTYLEDON1 Is Sufficient to Induce Embryo Development in Vegetative Cells. Cell 1998, 93, 1195–1205. [Google Scholar] [CrossRef] [Green Version]
- Sano, N.; Rajjou, L.; North, H.M.; Debeaujon, I.; Marion-Poll, A.; Seo, M. Staying Alive: Molecular Aspects of Seed Longevity. Plant Cell Physiol. 2016, 57, 660–674. [Google Scholar] [CrossRef] [Green Version]
- Leprince, O.; Pellizzaro, A.; Berriri, S.; Buitink, J. Late Seed Maturation: Drying without Dying. J. Exp. Bot. 2016, 363. [Google Scholar] [CrossRef] [Green Version]
- Santos-Mendoza, M.; Dubreucq, B.; Baud, S.; Parcy, F.; Caboche, M.; Lepiniec, L. Deciphering Gene Regulatory Networks That Control Seed Development and Maturation in Arabidopsis. Plant J. 2008, 54, 608–620. [Google Scholar] [CrossRef]
- Linkies, A.; Graeber, K.; Knight, C.; Leubner-Metzger, G. The Evolution of Seeds. New Phytol. 2010, 186, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Kwong, R.W.; Bui, A.Q.; Lee, H.; Kwong, L.W.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. LEAFY COTYLEDON1-LIKE Defines a Class of Regulators Essential for Embryo Development. Plant Cell 2003, 15, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Grimault, A.; Gendrot, G.; Chaignon, S.; Gilard, F.; Tcherkez, G.; Thévenin, J.; Dubreucq, B.; Depège-Fargeix, N.; Rogowsky, P.M. Role of B3 Domain Transcription Factors of the AFL Family in Maize Kernel Filling. Plant Sci. 2015, 236, 116–125. [Google Scholar] [CrossRef]
- Boulard, C.; Fatihi, A.; Lepiniec, L.; Dubreucq, B. BBA-Gene Regulatory Mechanisms Regulation and Evolution of the Interaction of the Seed B3 Transcription Factors with NF-Y Subunits. Biochim. Biophys. Acta Gene Regul. Mech. 2017, 1860, 1069–1078. [Google Scholar] [CrossRef]
- Giraudat, J.; Hauge, B.M.; Valon, C.; Smalle, J.; Parcy, F.; Goodman, H.M. Isolation of the Arabidopsis ABI3 Gene by Positional Cloning. Plant Cell 1992, 4, 1251–1261. [Google Scholar] [CrossRef]
- Suzuki, M.; Kao, C.Y.; McCarty, D.R. The Conserved B3 Domain of VIVIPAROUS1 Has a Cooperative DNA Binding Activity. Plant Cell 1997, 9, 799–807. [Google Scholar] [CrossRef] [Green Version]
- McCarty, D.R.; Hattori, T.; Carson, C.B.; Vasil, V.; Lazar, M.; Vasil, I.K. The Viviparous-1 Developmental Gene of Maize Encodes a Novel Transcriptional Activator. Cell 1991, 66, 895–905. [Google Scholar] [CrossRef]
- Nakamura, S.; Lynch, T.J.; Finkelstein, R.R. Physical Interactions between ABA Response Loci of Arabidopsis. Plant J. 2001, 26, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.; Nantel, A.; Rock, C.D.; Quatrano, R.S. A Conserved Domain of the Viviparous-1 Gene Product Enhances the DNA Binding Activity of the BZIP Protein EmBP-1 and Other Transcription Factors. J. Biol. Chem. 1996, 271, 3366–3374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bies-Etheve, N.; Da, A.; Conceicao, S.; Giraudat, J.; Koornneef, M.; Léon-Kloosterziel, K.; Valon, C.; Delseny, M. Importance of the B2 Domain of the Arabidopsis ABI3 Protein for Em and 2S Albumin Gene Regulation. Plant Mol. Biol. 1999, 40, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Ezcurra, I.; Wycliffe, P.; Nehlin, L.; Ellerström, M.; Rask, L. Transactivation of the Brassica Napus Napin Promoter by ABI3 Requires Interaction of the Conserved B2 and B3 Domains of ABI3 with Different Cis-Elements: B2 Mediates Activation through an ABRE, Whereas B3 Interacts with an RY/G-Box. Plant J. 2000, 24, 57–66. [Google Scholar] [CrossRef]
- Nag, R.; Maity, M.K.; DasGupta, M. Dual DNA Binding Property of ABA Insensitive 3 like Factors Targeted to Promoters Responsive to ABA and Auxin. Plant Mol. Biol. 2005, 59, 821–838. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.Y.; Weselake, R.J. Genome-Wide Identification and Analysis of the B3 Superfamily of Transcription Factors in Brassicaceae and Major Crop Plants. Theor. Appl. Genet. 2013, 126, 1305–1319. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Molina, L.; Mongrand, S.; McLachlin, D.T.; Chait, B.T.; Chua, N.H. ABI5 Acts Downstream of ABI3 to Execute an ABA-Dependent Growth Arrest during Germination. Plant J. 2002, 32, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Zhao, X.Y.; Shao, X.X.; Wang, F.; Zhou, C.; Liu, Y.G.; Zhang, Y.; Zhang, X.S. Abscisic Acid Regulates Early Seed Development in Arabidopsis by ABI5-Mediated Transcription of SHORT HYPOCOTYL UNDER BLUE1. Plant Cell 2014, 26, 1053–1068. [Google Scholar] [CrossRef] [Green Version]
- Pellizzaro, A.; Neveu, M.; Lalanne, D.; Ly Vu, B.; Kanno, Y.; Seo, M.; Leprince, O.; Buitink, J. A Role for Auxin Signaling in the Acquisition of Longevity during Seed Maturation. New Phytol. 2020, 225, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.; Reeves, W.; Ariizumi, T.; Steber, C. Molecular Aspects of Seed Dormancy. Annu. Rev. Plant Biol. 2008, 59, 387–415. [Google Scholar] [CrossRef] [Green Version]
- Brady, S.M.; Sarkar, S.F.; Bonetta, D.; McCourt, P. The ABSCISIC ACID INSENSITIVE 3 (ABI3) Gene Is Modulated by Farnesylation and Is Involved in Auxin Signaling and Lateral Root Development in Arabidopsis. Plant J. 2003, 34, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Suzuki, M.; Mccarty, D.R. Regulation of the Seed to Seedling Developmental Phase Transition by the LAFL and VAL Transcription Factor Networks. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 135–145. [Google Scholar] [CrossRef]
- Alonso, R.; Oñate-Sánehez, L.; Weltmeier, F.; Ehlert, A.; Diaz, I.; Dietrich, K.; Vicente-Carbajosa, J.; Dröge-Laser, W. A Pivotal Role of the Basic Leucine Zipper Transcription Factor BZIP53 in the Regulation of Arabidopsis Seed Maturation Gene Expression Based on Heterodimerization and Protein Complex FormationW. Plant Cell 2009, 21, 1747–1761. [Google Scholar] [CrossRef] [Green Version]
- Barthole, G.; To, A.; Marchive, C.; Brunaud, V.; Soubigou-Taconnat, L.; Berger, N.; Dubreucq, B.; Lepiniec, L.; Baud, S. MYB118 Represses Endosperm Maturation in Seeds of Arabidopsis. Plant Cell 2014, 26, 3519–3537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmas, F.; Sankaranarayanan, S.; Deb, S.; Widdup, E.; Bournonville, C.; Bollier, N.; Northey, J.G.B.; McCourt, P.; Samuel, M.A. ABI3 Controls Embryo Degreening through Mendel’s i Locus. Proc. Natl. Acad. Sci. USA 2013, 110, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bassel, G.W.; Stamm, P.; Mosca, G.; De Reuille, P.B.; Gibbs, D.J.; Winter, R.; Janka, A.; Holdsworth, M.J.; Smith, R.S. Mechanical Constraints Imposed by 3D Cellular Geometry and Arrangement Modulate Growth Patterns in the Arabidopsis Embryo. Proc. Natl. Acad. Sci. USA 2014, 111, 8685–8690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baud, S.; Kelemen, Z.; Thévenin, J.; Boulard, C.; Blanchet, S.; To, A.; Payre, M.; Berger, N.; Effroy-Cuzzi, D.; Franco-Zorrilla, J.M.; et al. Deciphering the Molecular Mechanisms Underpinning the Transcriptional Control of Gene Expression by Master Transcriptional Regulators in Arabidopsis Seed. Plant Physiol. 2016, 171, 1099–1112. [Google Scholar] [CrossRef]
- Baud, S.; Mendoza, M.S.; To, A.; Harscoët, E.; Lepiniec, L.; Dubreucq, B. WRINKLED1 Specifies the Regulatory Action of LEAFY COTYLEDON2 towards Fatty Acid Metabolism during Seed Maturation in Arabidopsis. Plant J. 2007, 50, 825–838. [Google Scholar] [CrossRef] [PubMed]
- Kurdyukov, S.; Song, Y.; Sheahan, M.B.; Rose, R.J. Transcriptional Regulation of Early Embryo Development in the Model Legume Medicago Truncatula. Plant Cell Rep. 2014, 33, 349–362. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; He, L.; Wang, X.D.; Smith, N.; Wheeler, S.; Garg, M.L.; Rose, R.J. Regulation of Carbon Partitioning in the Seed of the Model Legume Medicago Truncatula and Medicago Orbicularis: A Comparative Approach. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delahaie, J.; Hundertmark, M.; Bove, J.; Leprince, O.; Rogniaux, H.; Buitink, J. LEA Polypeptide Profiling of Recalcitrant and Orthodox Legume Seeds Reveals ABI3-Regulated LEA Protein Abundance Linked to Desiccation Tolerance. J. Exp. Bot. 2013, 64, 4559–4573. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, T.; Song, X.; Zhang, J.; Dang, Z.; Pei, X.; Long, Y. Identification and Functional Analysis of Two Alternatively Spliced Transcripts of ABSCISIC ACID INSENSITIVE3 (ABI3) in Linseed Flax (Linum Usitatissimum, L.). PLoS ONE 2018, 13, e0191910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gagete, A.P.; Riera, M.; Franco, L.; Rodrigo, M.I. Functional Analysis of the Isoforms of an ABI3-like Factor of Pisum Sativum Generated by Alternative Splicing. J. Exp. Bot. 2009, 60, 1703–1714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Liu, J.; Zhang, Z.; Sun, X.; Zhang, N.; Fan, J.; Niu, X.; Xiao, F.; Liu, Y. Functional Characterization of Two Alternatively Spliced Transcripts of Tomato ABSCISIC ACID INSENSITIVE3 (ABI3) Gene. Plant Mol. Biol. 2013, 82, 131–145. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. ClusterProfiler: An R Package for Comparing Biological Themes among Gene Clusters. OMICS 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Mönke, G.; Seifert, M.; Keilwagen, J.; Mohr, M.; Grosse, I.; Hähnel, U.; Junker, A.; Weisshaar, B.; Conrad, U.; Bäumlein, H.; et al. Toward the Identification and Regulation of the Arabidopsis Thaliana ABI3 Regulon. Nucleic Acids Res. 2012, 40, 8240–8254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Ly Vu, B.; Leprince, O.; Verdier, J. RNA Sequencing Data for Heat Stress Response in Isolated Medicago Truncatula Seed Tissues. Data Brief 2021, 35, 106726. [Google Scholar] [CrossRef] [PubMed]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C.; Biology, C. Salmon: Fast and Bias-Aware Quantification of Transcript Expression Using Dual-Phase Inference. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szakonyi, D.; Duque, P. Alternative Splicing as a Regulator of Early Plant Development. Front. Plant Sci. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Syed, N.H.; Kalyna, M.; Marquez, Y.; Barta, A.; Brown, J.W.S. Alternative Splicing in Plants-Coming of Age. Trends Plant Sci. 2012, 17, 616–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filichkin, S.A.; Priest, H.D.; Givan, S.A.; Shen, R.; Bryant, D.W.; Fox, S.E.; Wong, W.-K.; Mockler, T.C. Genome-Wide Mapping of Alternative Splicing in Arabidopsis Thaliana. Genome Res. 2010, 20, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aghamirzaie, D.; Nabiyouni, M.; Fang, Y.; Klumas, C.; Heath, L.S.; Grene, R.; Collakova, E. Changes in RNA Splicing in Developing Soybean (Glycine Max) Embryos. Biology 2013, 2, 1311–1337. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, A.; Jiménez-Gómez, J.M.; Fornara, F.; Soppe, W.J.J.; Brambilla, V. Alternative Splicing Enhances Transcriptome Complexity in Desiccating Seeds. J. Integr. Plant Biol. 2016, 58, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Sugliani, M.; Brambilla, V.; Clerkx, E.J.M.; Koornneef, M.; Soppe, W.J.J. The Conserved Splicing Factor SUA Controls Alternative Splicing of the Developmental Regulator ABI3 in Arabidopsis. Plant Cell 2010, 22, 1936–1946. [Google Scholar] [CrossRef] [Green Version]
- Penfield, S.; Karen, E.J. A Role for an Alternative Splice Variant of PIF6 in the Control of Arabidopsis Primary Seed Dormancy. Plant Mol. Biol. 2010, 89–95. [Google Scholar] [CrossRef]
- Nakabayashi, K.; Bartsch, M.; Ding, J.; Soppe, W.J.J. Seed Dormancy in Arabidopsis Requires Self- Binding Ability of DOG1 Protein and the Presence of Multiple Isoforms Generated by Alternative Splicing. PLoS Genet. 2015, 11, e1005737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, R.R.; Lynch, T.J. The Arabidopsis Abscisic Acid Response Gene ABI5 Encodes a Basic Leucine Zipper Transcription Factor. Plant Cell 2000, 12, 599–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bossi, F.; Cordoba, E.; Dupré, P.; Mendoza, M.S.; Román, C.S.; León, P. The Arabidopsis ABA-INSENSITIVE (ABI) 4 Factor Acts as a Central Transcription Activator of the Expression of Its Own Gene, and for the Induction of ABI5 and SBE2.2 Genes during Sugar Signaling. Plant J. 2009, 59, 359–374. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, M.; Ohme-Takagi, M. A Novel Group of Transcriptional Repressors in Arabidopsis. Plant Cell Physiol. 2009, 50, 970–975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, N.I.; Tuan, P.A.; Xu, H.; Park, S.U. Isolation and Characterization of the PgDOF Transcription Factor in Platycodon Grandiflorum. Plant Omics 2011, 4, 149–153. [Google Scholar]
- Suzuki, M.; McCarty, D.R. Functional Symmetry of the B3 Network Controlling Seed Development. Curr. Opin. Plant Biol. 2008, 11, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Sasnauskas, G.; Manakova, E.; Lapėnas, K.; Kauneckaitė, K.; Siksnys, V. DNA Recognition by Arabidopsis Transcription Factors ABI3 and NGA1. FEBS J. 2018, 285, 4041–4059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, H.; McCarty, D.R.; Suzuki, M. Distinct Roles of LAFL Network Genes in Promoting the Embryonic Seedling Fate in the Absence of VAL Repression. Plant Physiol. 2013, 163, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Terrasson, E.; Buitink, J.; Righetti, K.; Ly Vu, B.; Pelletier, S.; Zinsmeister, J.; Lalanne, D.; Leprince, O. An Emerging Picture of the Seed Desiccome: Confirmed Regulators and Newcomers Identified Using Transcriptome Comparison. Front. Plant Sci. 2013, 4, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karimi, M.; Inzé, D.; Depicker, A. GATEWAY Vectors for Agrobacterium-Mediated Plant Transformation. Trends Plant Sci. 2002, 7, 193–195. [Google Scholar] [CrossRef]
- Verdier, J.; Zhao, J.; Torres-Jerez, I.; Ge, S.; Liu, C.; He, X.; Mysore, K.S.; Dixon, R.A.; Udvardi, M.K. MtPAR MYB Transcription Factor Acts as an on Switch for Proanthocyanidin Biosynthesis in Medicago Truncatula. Proc. Natl. Acad. Sci. USA 2012, 109, 1766–1771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quandt, H.; Puhler, A.; Broer, I. Transgenic Root Nodules of Vicia Hirsuta: A Fast and Efficient System for the Study of Gene Expression in Indeterminate-Type Nodules. Mol. Plant Microbe Interact. 1993, 6, 699–706. [Google Scholar] [CrossRef]
- Chabaud, M.; de Carvalho-Niebel, F.; Barker, D.G. Efficient Transformation of Medicago Truncatula Cv. Jemalong Using the Hypervirulent Agrobacterium Tumefaciens Strain AGL1. Plant Cell Rep. 2003, 22, 46–51. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral Dip: A Simplified Method for Agrobacterium-Mediated Transformation of Arabidopsis Thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Zhang, B.; Lin, L.L.; Zhao, S. Evaluation and Comparison of Computational Tools for RNA-Seq Isoform Quantification. BMC Genom. 2017, 18, 1–11. [Google Scholar] [CrossRef]
- Schwacke, R.; Ponce-Soto, G.Y.; Krause, K.; Bolger, A.M.; Arsova, B.; Hallab, A.; Gruden, K.; Stitt, M.; Bolger, M.E.; Usadel, B. MapMan4: A Refined Protein Classification and Annotation Framework Applicable to Multi-Omics Data Analysis. Mol. Plant 2019, 12, 879–892. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lalanne, D.; Malabarba, J.; Ly Vu, J.; Hundertmark, M.; Delahaie, J.; Leprince, O.; Buitink, J.; Verdier, J. Medicago ABI3 Splicing Isoforms Regulate the Expression of Different Gene Clusters to Orchestrate Seed Maturation. Plants 2021, 10, 1710. https://doi.org/10.3390/plants10081710
Lalanne D, Malabarba J, Ly Vu J, Hundertmark M, Delahaie J, Leprince O, Buitink J, Verdier J. Medicago ABI3 Splicing Isoforms Regulate the Expression of Different Gene Clusters to Orchestrate Seed Maturation. Plants. 2021; 10(8):1710. https://doi.org/10.3390/plants10081710
Chicago/Turabian StyleLalanne, David, Jaiana Malabarba, Joseph Ly Vu, Michaela Hundertmark, Julien Delahaie, Olivier Leprince, Julia Buitink, and Jerome Verdier. 2021. "Medicago ABI3 Splicing Isoforms Regulate the Expression of Different Gene Clusters to Orchestrate Seed Maturation" Plants 10, no. 8: 1710. https://doi.org/10.3390/plants10081710