
Environmental and Management Effects on Demographic Processes in the U.S. Threatened Platanthera leucophaea (Nutt.) Lindl. (Orchidaceae)


 ,
,
Abstract
:1. Introduction
- (a)
- How do long-term trends in sizes of flowering plant populations correspond to habitat and climatic variation?
- (b)
- How are flowering and fecundity affected by habitat and fire, and how do they interact with climatic variation?
- (c)
- How is plant survival affected by habitat and fire, and how do these affects interact with climatic variation?
- (d)
- How do crossing rates (selfing and outcrossing within and among populations) affect germination?
- (e)
- What are the effects of these factors on population demography?
1.1. Study Species
1.2. Study Area
2. Results
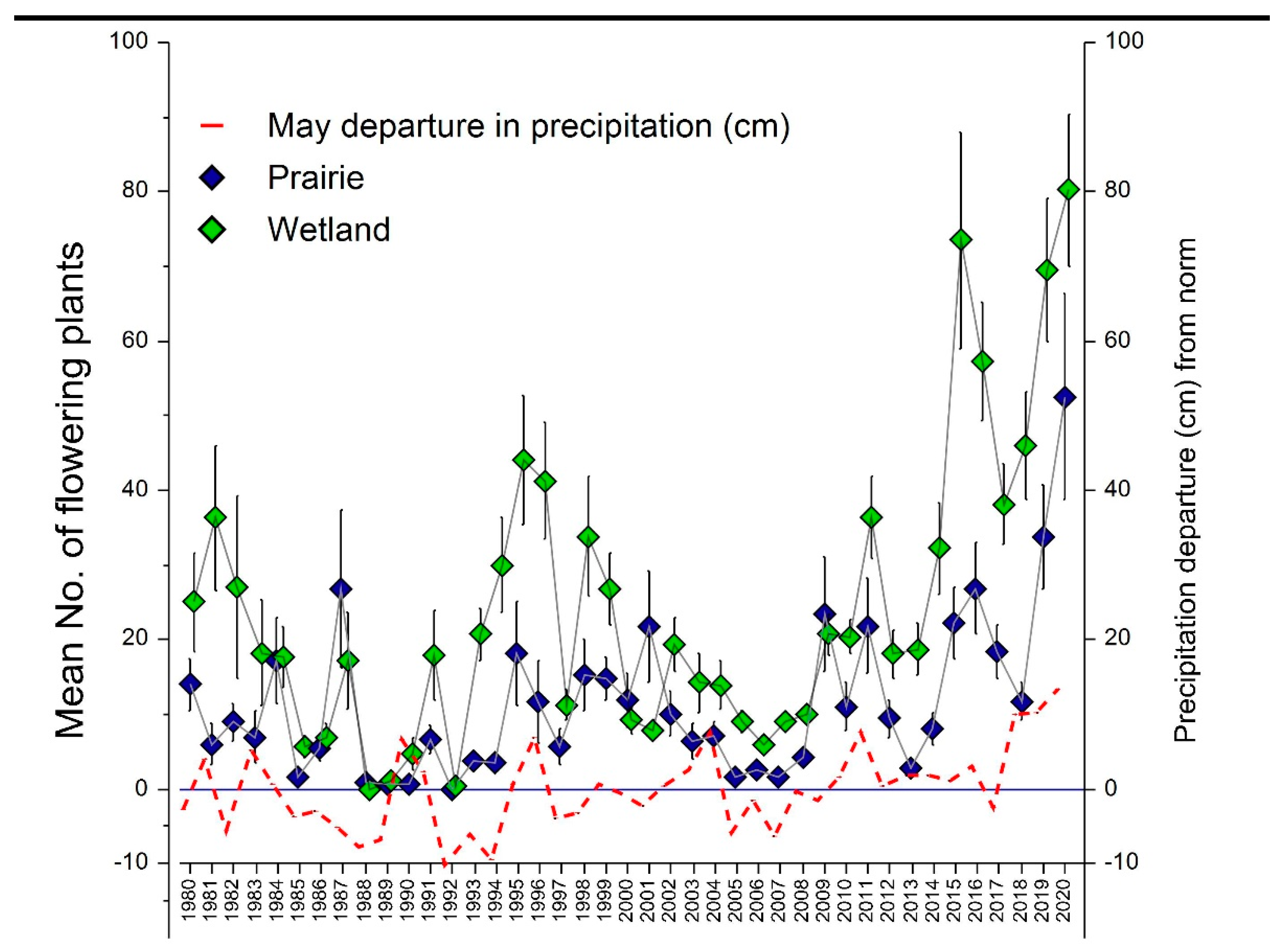
2.1. Long-Term Trends in Prairie and Wetland Populations
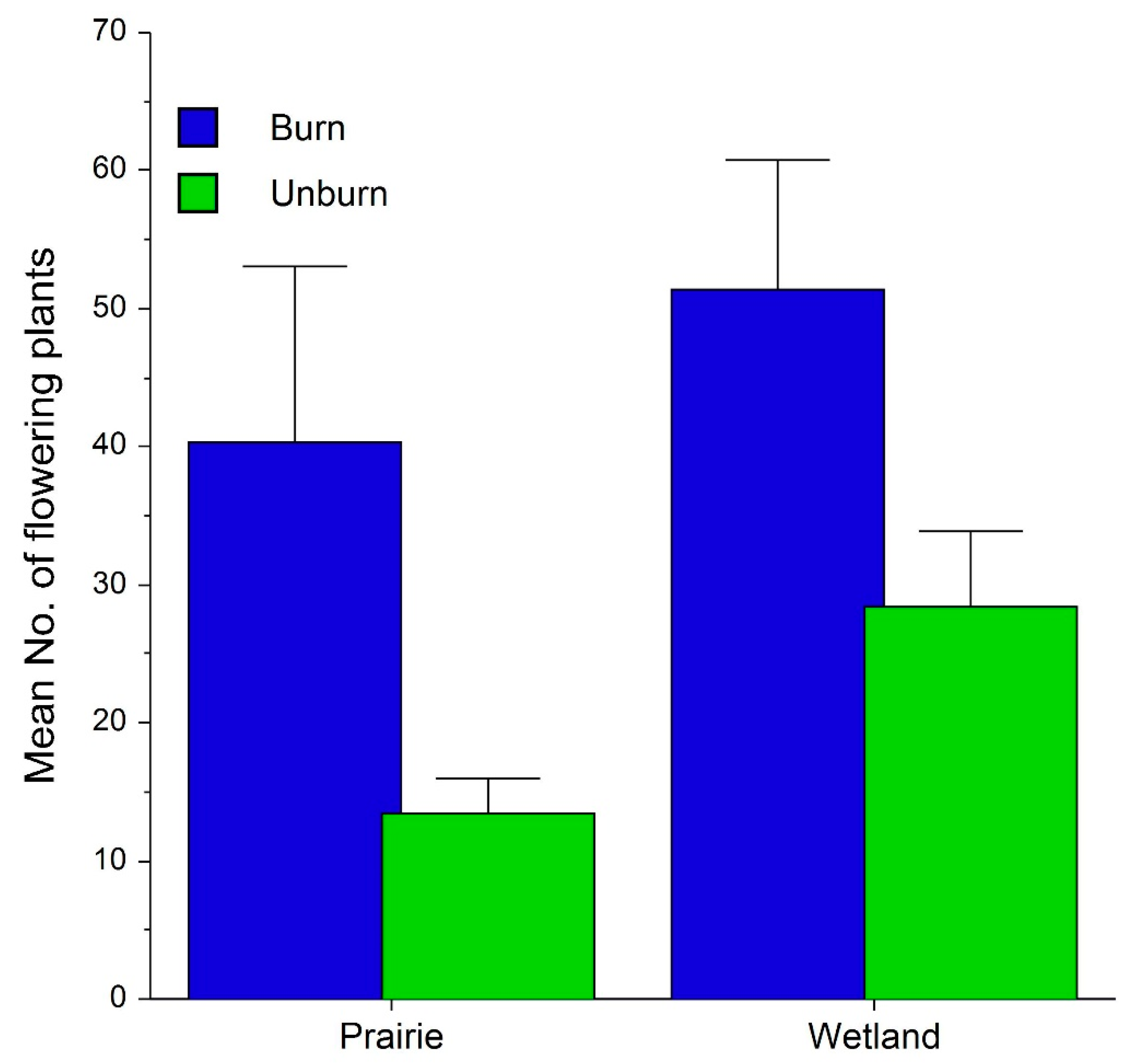
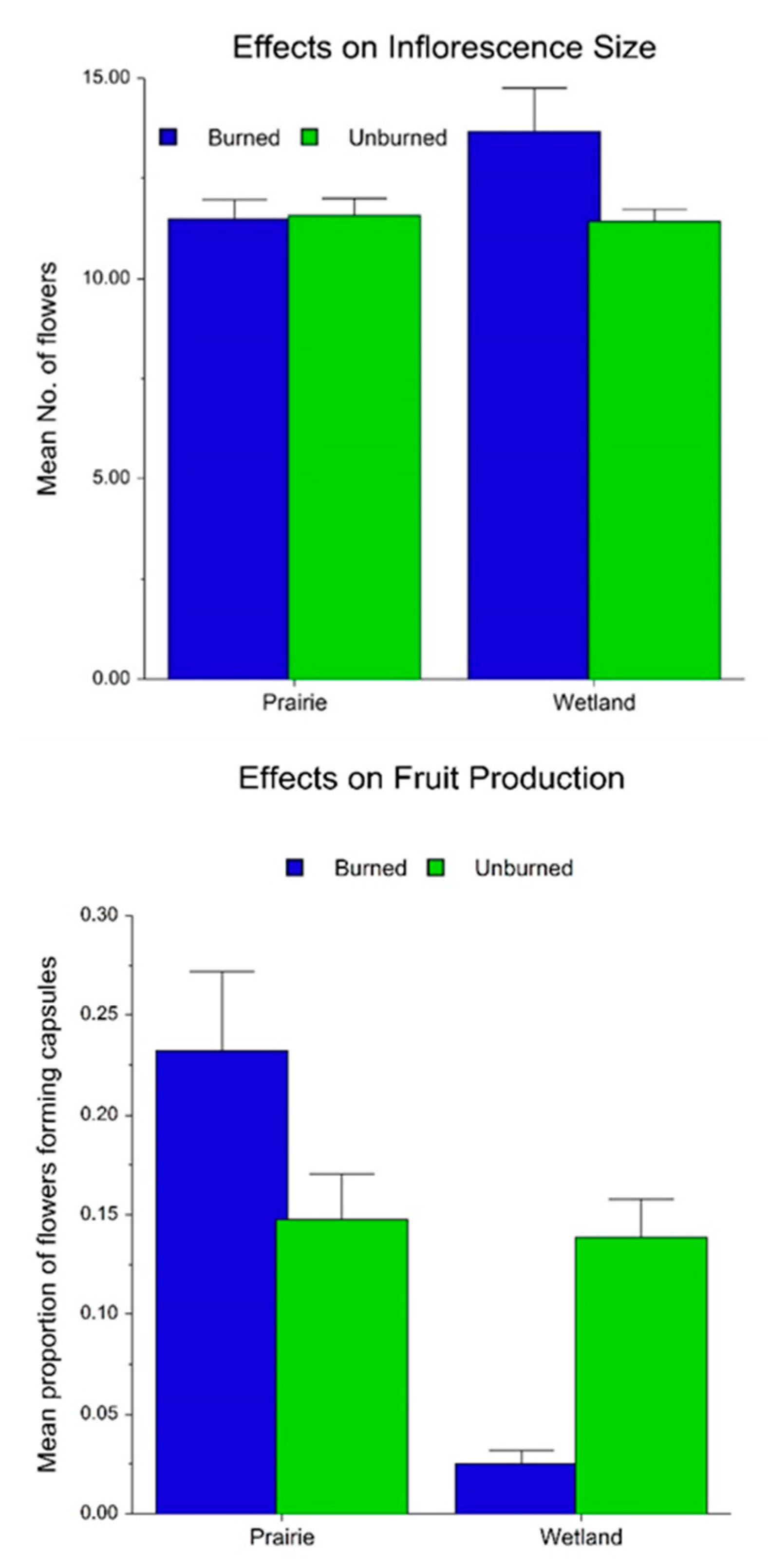
2.2. Fire, Habitat and Climatic Effects on Flowering and Fecundity
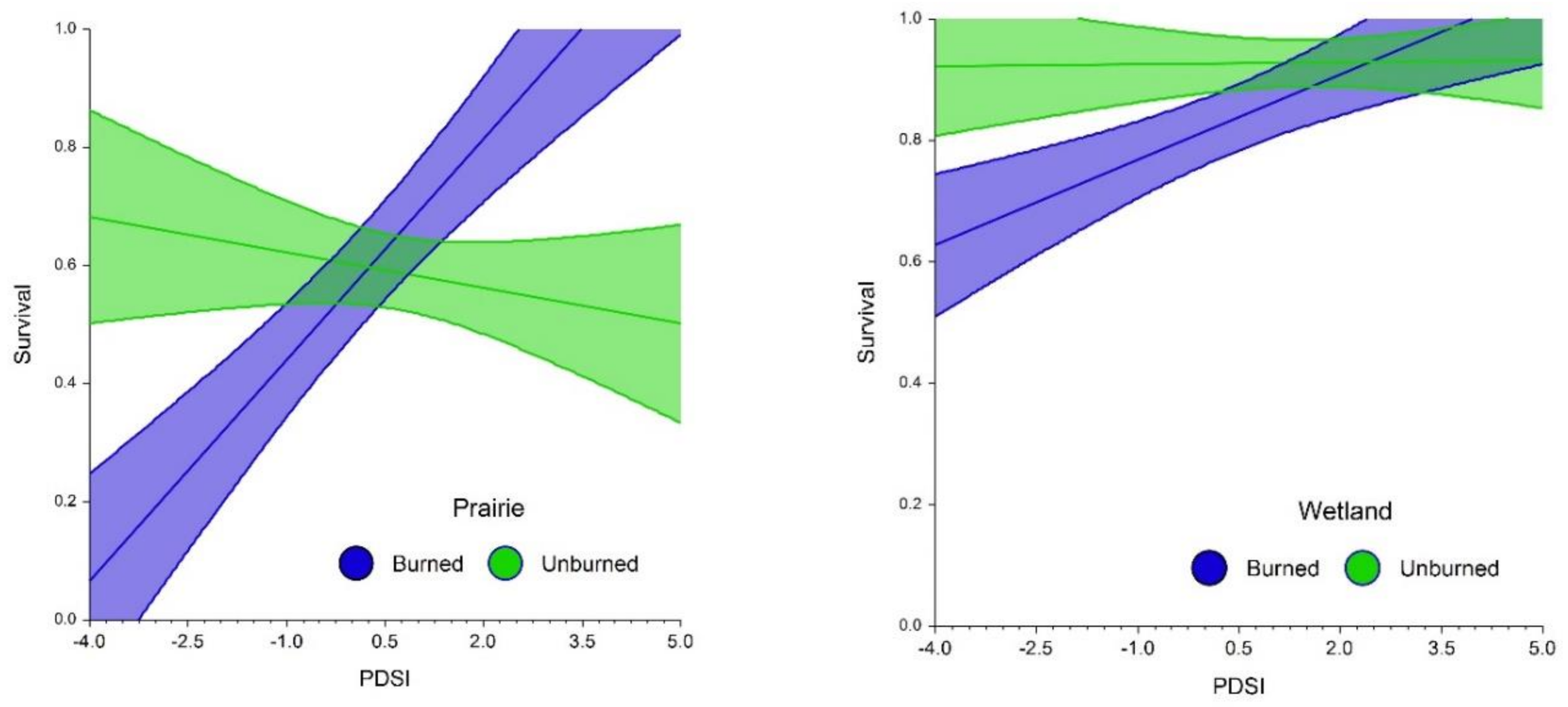
2.3. Fire, Habitat and Climatic Effects on Survival
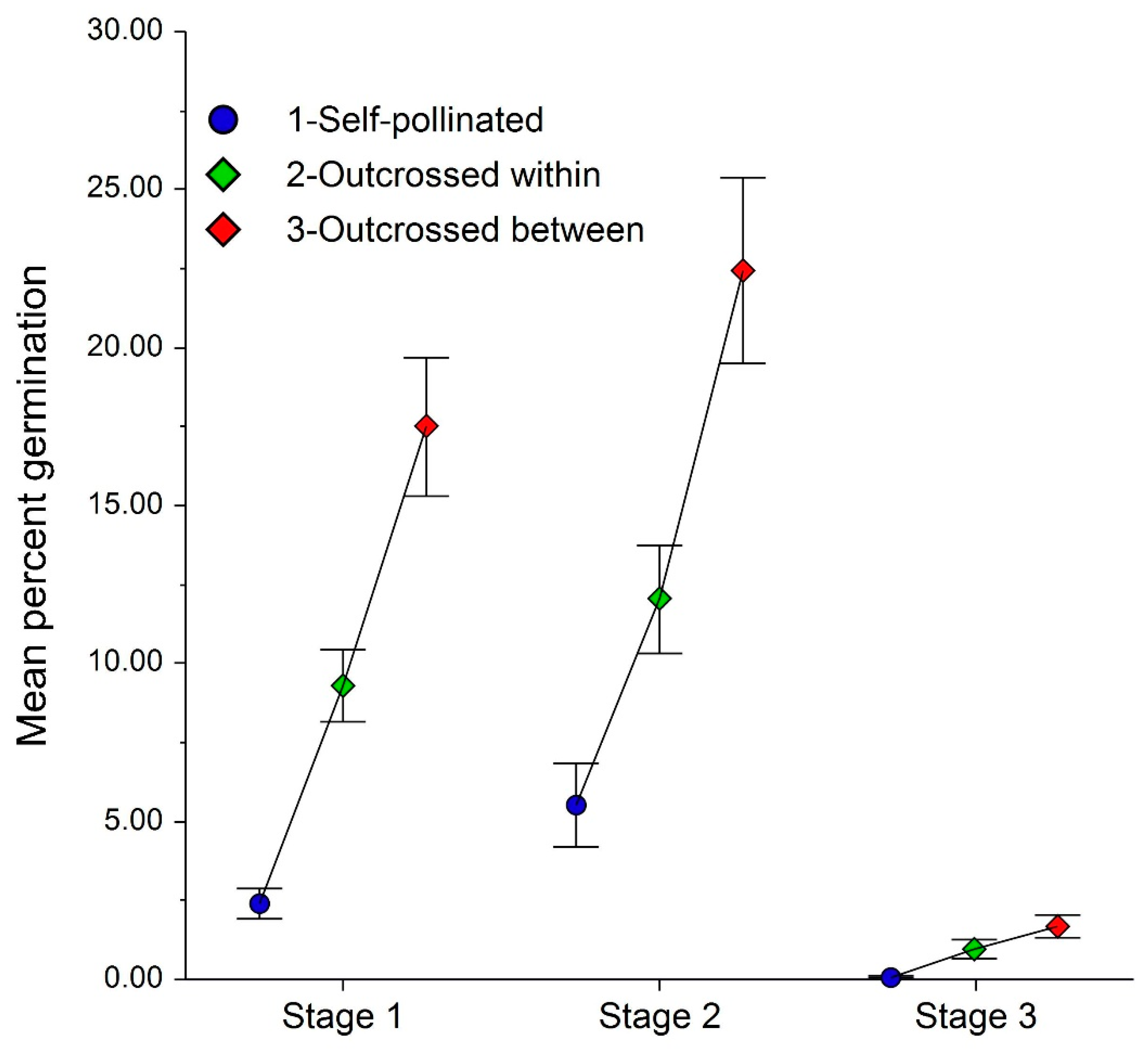
2.4. Crossing Effects on Germination
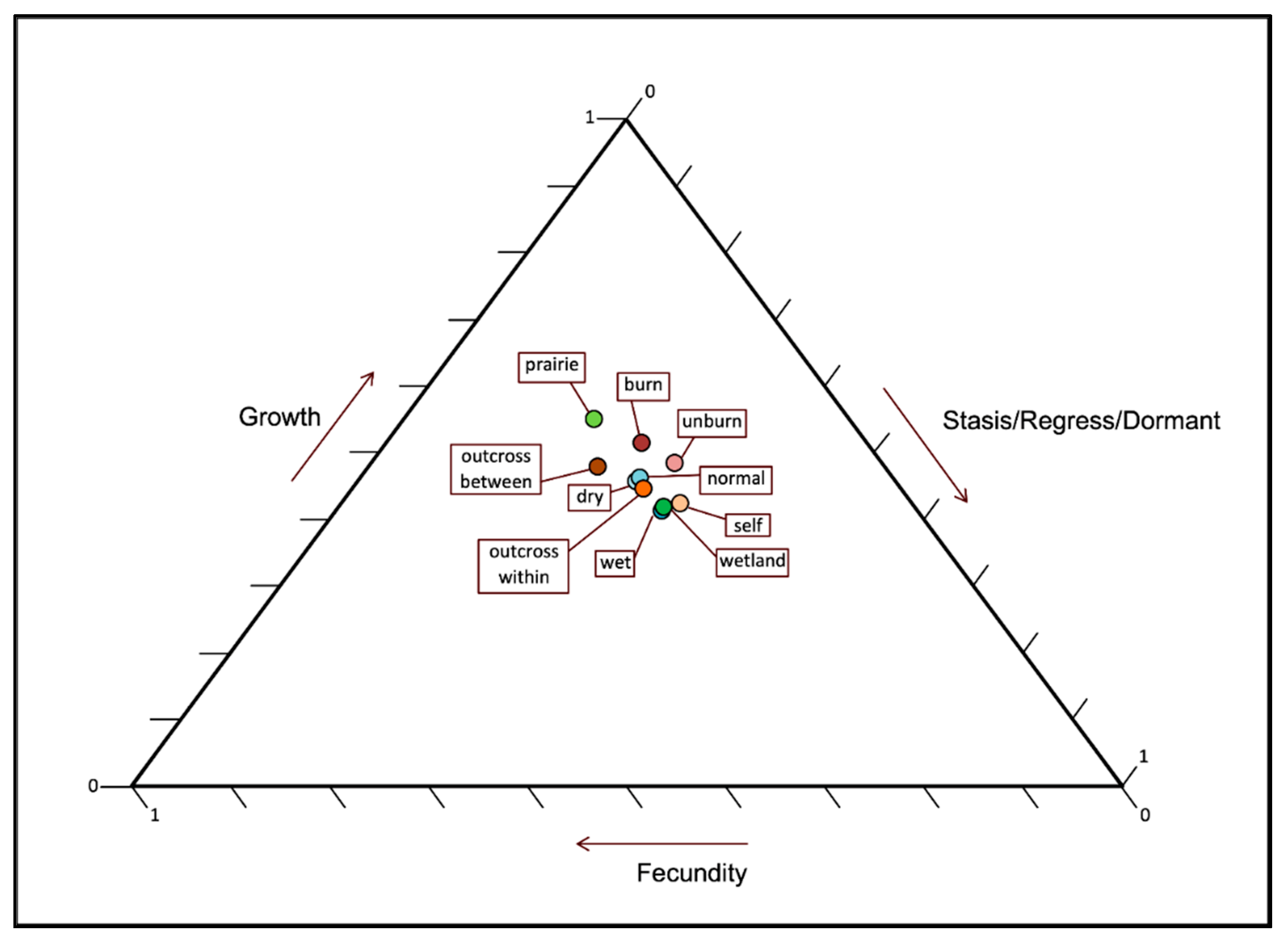
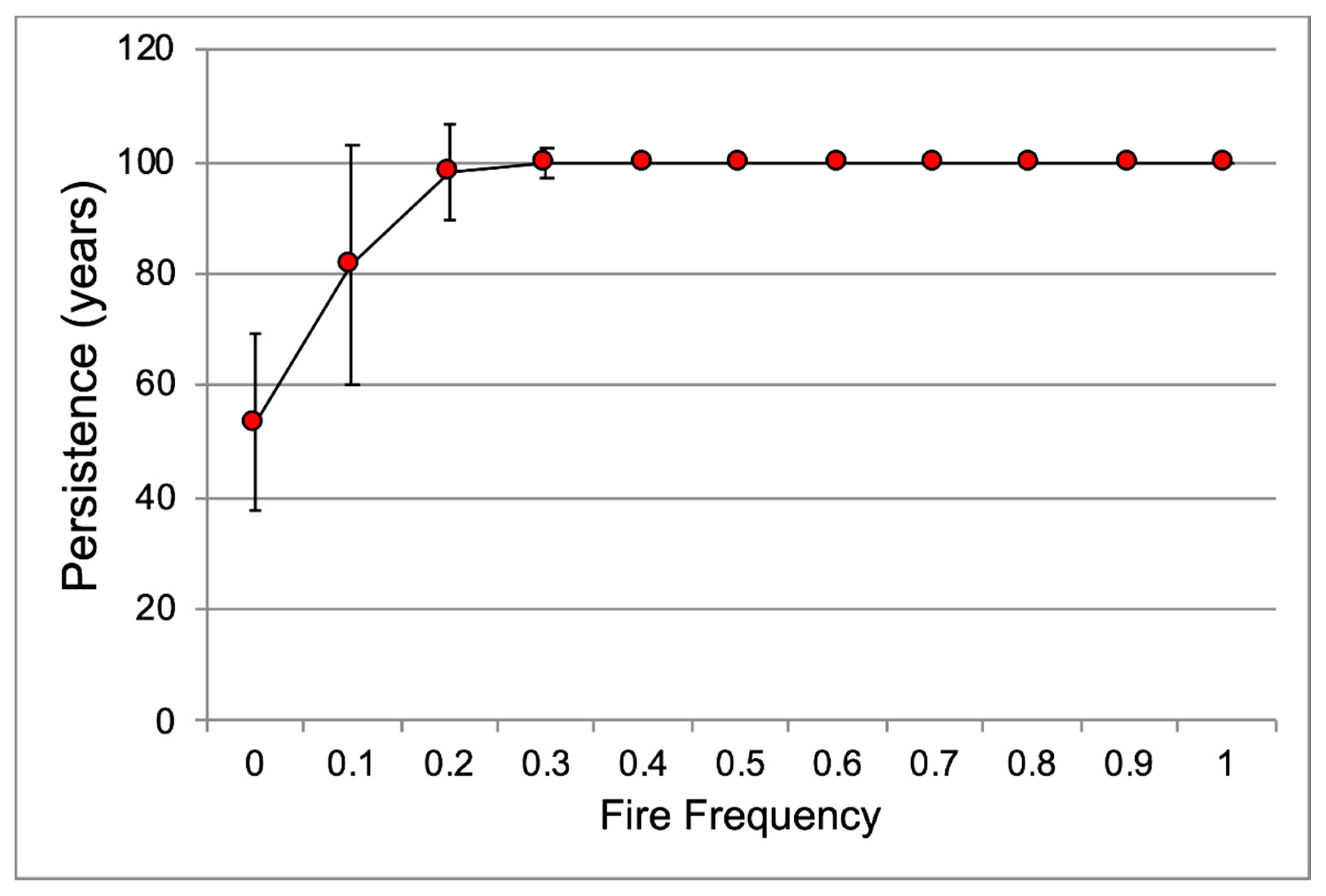
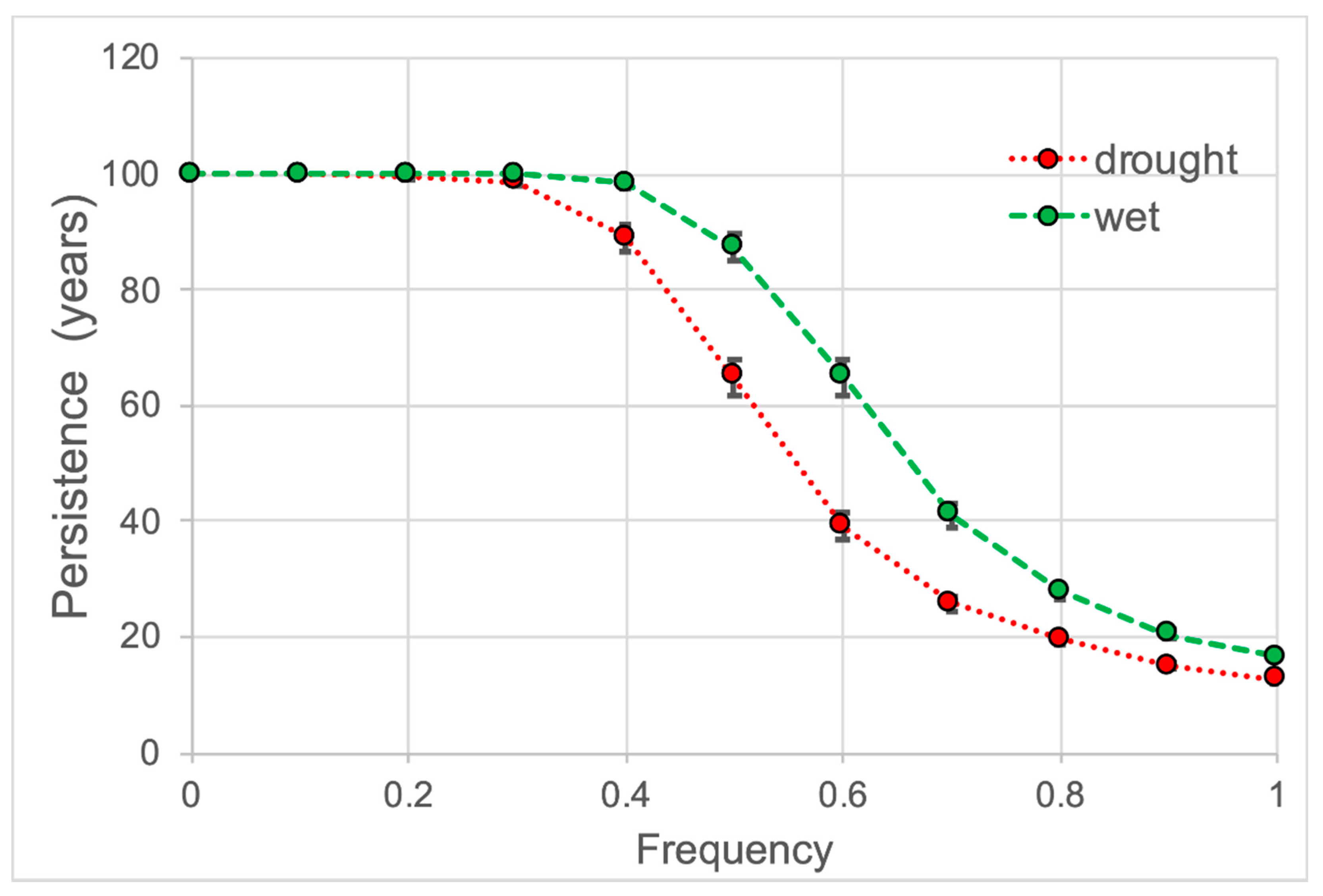
2.5. Effects on Population Demography
3. Discussion
3.1. Long-Term Trends in Prairie and Wetland Populations
3.2. Effects on Flowering, Fecundity and Survival
3.3. Crossing Effects on Seed Germination
3.4. Effects of These Factors on Population Demography
3.5. The Impact of Burning on Biotic Agents—Moth Pollinators and Mycorrhizal Fungi
4. Materials and Methods
4.1. How Do Long-Term Trends in Sizes of Flowering Plant Populations Correspond to Habitat and Climatic Variation?
4.2. How Are Flowering and Fecundity Affected by Habitat and Fire and How Do They Interact with Climatic Variation?
4.3. How Is Plant Survival Affected by Habitat and Fire, and How Do These Affects Interact with Climatic Variation?
4.4. How Do Crossing Rates (Selfing, Within and Among Populations) Affect Seed Germination?
4.5. What Are the Effects of These Factors on Population Demography?
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fay, M.F.; Pallier, T.; Dixon, K.W. Orchid conservation: Making the links. Ann. Bot. 2015, 116, 377–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fay, M.F. Orchid conservation: How we meet the challenges in the twenty-first century? Bot. Stud. 2018, 59, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, C.J.; Scott, P.J. Environmental parameters influencing the distribution of Platanthera blephariglottis and Platanthera clavellata (Orchidaceae) in peatlands on the Avalon Peninsula, Newfoundland. Can. J. Bot. 1997, 75, 975–980. [Google Scholar] [CrossRef]
- Brzosko, E. The dynamics of island populations of Platanthera bifolia in the Biebrza National Park (NE Poland). Ann. Bot. Fenn. 2003, 40, 243–252. Available online: https://jstor.org/stable/23726841 (accessed on 19 March 2021).
- Mursal, N.; Mehdiyeva, N.P.; Ibrahimova, A.G. Population status and ecology of Platanthera chlorantha (Orchidaceae) in the greater Caucasus (Azerbaijan). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 114–124. [Google Scholar] [CrossRef]
- Romano, V.A.; Rosati, L.; Fascetti, S. Trends in population size of Ophrys argolica subs. biscutella in the Appennino Lucano-Val d’Agri-Lagonegrese National Park (Italy). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 155–164. [Google Scholar] [CrossRef]
- Reddoch, J.M.; Reddoch, J.H. Population ecology of Platanthera hookeri (Orchidaceae) in southwestern Quebec, Canada. J. Torrey Bot. Soc. 2007, 134, 369–378. Available online: https://www.jstor.org/stable/20063928 (accessed on 19 March 2021).
- Kirillova, I.A.; Kirillov, D. Impact of weather conditions on seasonal development: Population structure and reproductive success on Dactylorhiza traunsteineri (Orchidaceae) in the Komi Republic (Russia). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 77–89. [Google Scholar] [CrossRef] [Green Version]
- Wilson, G.; Page, M.; Akyuz, F.A. Precipitation and fire effects on flowering of a rare prairie orchid. Great Plains Res. 2006, 16, 37–43. [Google Scholar]
- Morrison, L.W.; Haack-Gaynor, J.L.; Young, C.C.; DeBacker, M.D. A 20-Year Record of the Western Prairie Fringed Orchid (Platanthera praeclara). Nat. Areas J. 2015, 35, 246–255. [Google Scholar] [CrossRef]
- Bowles, M.L.; Flakne, R.; Dombeck, R. Status and population fluctuations of the eastern prairie fringed orchid (Platanthera leucophaea (Nutt.) Lindl) in Illinois. Erigenia 1992, 12, 26–40. [Google Scholar]
- Pleasants, J.M. The effects of spring burns on the western prairie fringed orchid (Platanthera praeclara). In Proceedings of the 14th Annual North American Prairie Conference, Kansas State University, Manhattan, KS, USA, 12–16 July 1994; pp. 67–73. [Google Scholar]
- Alexander, B.W. An Analysis of Seed Production, Viability, Germination in Situ, and Grazing Impacts on the Western Prairie Fringed Orchid (Platanthera praeclara, Sheviak and Bowles). Ph.D. Thesis, North Dakota State University, Fargo, ND, USA, 2006. [Google Scholar]
- Perazza, G.; Decarli, M. Monitoring Cypripedium calceolus (Orchidaceae) in the Adamello-Brenta Natural Park (Italy). Nat. Conserv. Res. 2020, 5 (Suppl. 1), 178–184. [Google Scholar] [CrossRef]
- Tatarenko, I.; Dodd, I.M.; Wallace, H.; Bellamy, G.; Fleckney, A. Protecting small populations of rare species. Case study on Dactylorhiza viridis (Orchidaceae) in Fancott Woods and Meadows, Bedfordshire, UK. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 165–171. [Google Scholar] [CrossRef]
- Crone, E.E.; Menges, E.S.; Ellis, M.M.; Bell, T.; Bierzychudek, P.; Ehrlen, J.; Kaye, T.N.; Knight, T.M.; Lesica, P.; Morris, W.F.; et al. How do plant ecologists use matrix population models? Ecol. Lett. 2011, 14, 1–8. [Google Scholar] [CrossRef]
- Bowles, M.L. The tallgrass prairie orchids Platanthera leucophaea (Nutt.) Lindl. and Cypripedium candidum Muhl. ex Willd.: Some aspects of their status, biology, and ecology, and implications toward management. Nat. Areas J. 1983, 3, 14–37. [Google Scholar]
- Bleho, B.L.; Koper, N.; Borkowsky, C.L.; Hamel, C.D. Effects of weather and land management on the western prairie fringed orchid (Platanthera praeclara) at the northern limit of its range in Manitoba, Canada. Am. Midl. Nat. 2015, 174, 191–203. [Google Scholar] [CrossRef]
- Paul, J.; Budd, C.; Freeland, J.R. Conservation genetics of an endangered orchid in eastern Canada. Conserv. Genet. 2013, 14, 195–204. [Google Scholar] [CrossRef]
- Fox, K.; Anderson, M.; Andreson, R.; Foster, M.; Foster, C.; Vik, D.; Vitt, P.; Harris, M. Nectar Robbery and Thievery in the Hawk Moth (Lepidoptera: Sphingidae)-Pollinated Western Prairie Fringed Orchid Platanthera praeclara. Ann. Entomol. Soc. Am. 2015, 1–14. [Google Scholar] [CrossRef]
- Kaur, J.; Andrews, L.; Sharma, J. High specificity of a rare terrestrial orchid toward a rare fungus within the North American tallgrass prairie. Fungal Biol. 2019, 123, 895–904. [Google Scholar] [CrossRef]
- Alexander, B.W.; Kirby, D.; Biondini, M.; Dekeyser, E. Cattle grazing reduces survival and reproduction of the western prairie fringed orchid. Prairie Nat. 2010, 42, 46–49. [Google Scholar]
- Bell, T.; Bowles, M.; McEachern, K. Projecting the success of plant population restoration with viability analysis. In Population Viability in Plants; Brigham, C.A., Schwartz, M.W., Eds.; Springer: New York, NY, USA, 2003; pp. 313–348. [Google Scholar]
- Menges, E.S. Turner Review No. 16. Restoration demography and genetics of plants: When is a translocation successful? Aust. J. Bot. 2008, 56, 187–196. [Google Scholar] [CrossRef]
- Dalrymple, S.E.; Banks, E.; Stewart, G.B.; Pullin, A.S. A meta-analysis of threatened plant reintroductions from across the globe. In Plant Reintroductions from Across the Globe; Maschinski, J., Haskins, K.E., Eds.; Island Press: Washington, DC, USA, 2012; pp. 31–50. [Google Scholar]
- Neale, J.R. Genetic considerations in rare plant reintroduction: Practical applications (or how are we doing?). In Plant Reintroduction in a Changing Climate; Maschinski, J., Haskins, K.E., Eds.; Island Press: Washington, DC, USA, 2012; pp. 71–88. [Google Scholar]
- Sheviak, C.J.; Bowles, M.L. The Prairie Fringed Orchids: A Pollinator-Isolated Species Pair. Rhodora 1986, 88, 267–290. Available online: https://www.jstor.org/stable/23313939 (accessed on 19 March 2021).
- Knapp, A.K. Post-burn differences in solar radiation, leaf temperature and water stress influencing production in a lowland tallgrass prairie. Am. J. Bot. 1984, 71, 220–227. [Google Scholar] [CrossRef]
- Knapp, A.K. Effect of fire and drought on the ecophysiology of Andropogon gerardii and Panicum virgatum. Ecology 1985, 66, 1309–1320. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). Endangered and threatened wildlife and plants; Determination of threatened Status for eastern and western prairie fringed orchids. Final Rule Fed. Regist. 1989, 54, 39857–39863. [Google Scholar]
- Environment Canada. Recovery Strategy for the Eastern Prairie Fringed-Orchid (Platanthera leucophaea) in Canada. Species at Risk Act Recovery Strategy Series. Environment Canada, Ottawa. ii + 11 pp. + Appendices. 2012. Available online: https://www.canada.ca/en/environment-climate-change/services/species-risk-public-registry/recovery-strategies/eastern-prairie-fringed-orchid-2012.html (accessed on 19 March 2021).
- Treher, A.; Sharma, J.; Frances, A.; Poff, K. Platanthera leucophaea. The IUCN Red List of Threatened Species 2015, eT64176760A64176769. Available online: https://dx.doi.org/10.2305/IUCN.UK.2015-4.RLTS.T64176760A64176769.en (accessed on 19 March 2021).
- Bowles, M.L.; Zettler, L.W.; Bell, T.; Kelsey, P. Relationships between Soil Characteristics, Distribution and Restoration Potential of The Federal Threatened Eastern Prairie Fringed Orchid, Platanthera leucophaea (Nutt.) Lindl. Am. Midl. Nat. 2005, 154, 273–285. Available online: https://www.jstor.org/stable/3566690 (accessed on 19 March 2021). [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). Five Year Review: Eastern Prairie Fringed Orchid (Platanthera leucophaea); U.S. Fish and Wildlife Service (USFWS): Chicago, IL, USA, 2020; p. 20. [Google Scholar]
- Bowles, M.L.; Jacobs, K.A.; Zettler, L.W.; Wilson Delaney, T. Crossing effects on seed viability and experimental germination of the Federal threatened Platanthera leucophaea (Orchidaceae). Rhodora 2002, 104, 14–30. Available online: https://jstor.org/stable/23313546 (accessed on 11 April 2021).
- Pollack, C. Restoration of The Eastern Prairie Fringed Orchid (Platanthera leucophaea): Natural Pollinators and the Abundance of Larval Host Plants. Master’s Thesis, Northeastern Illinois University, Chicago, IL, USA, 2009. [Google Scholar]
- Dressler, R.L. The Orchids: Natural History and Classification; Harvard University Press: Cambridge, MA, USA, 1981; p. 332. [Google Scholar]
- Cuthrell, D.L.; Higman, P.J.; Peskar, M.R.; Windus, J.L. The Pollinators of Ohio and Michigan Populations of Eastern Prairie Fringed Orchid (Plantanthera leucophaea); U.S. Fish and Wildlife Service: Reynoldsburg, OH, USA, 1999. [Google Scholar]
- Crosson, A.L.; Dunford, J.C.; Young, D.K. Pollination and Other Insect Interactions of the Eastern Prairie Fringed Orchid (Platanthera leucophaea (Nuttall) Lindley) in Wisconsin; A report for the U.S. Fish and Wildlife Service; University of Wisconsin: Madison, WI, USA, 1999. [Google Scholar]
- Cuthrell, D.L. Insects Associated with the Prairie Fringed Orchids. Platanthera praeclara Sheviak & Bowles and P. leucophaea (Nuttall) Lindley. Master’s Thesis, North Dakota State University, Fargo, ND, USA, 1994. [Google Scholar]
- Robertson, C. Flowers and Insects X. Bot. Gaz. 1893, 18, 47–54. Available online: https://www.jstor.org/stable/2464633 (accessed on 19 October 2016). [CrossRef]
- Wallace, L.E. The cost of inbreeding in Platanthera leucophaea (Orchidaceae). Am. J. Bot. 2003, 90, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Zettler, L.W.; Stewart, S.L.; Bowles, M.L.; Jacobs, K.A. Mycorrhizal fungi and cold-assisted symbiotic germination of the Federally threatened eastern prairie fringed orchid, Platanthera leucophaea (Nuttall) Lindley. Am. Midl. Nat. 2001, 145, 168–175. [Google Scholar] [CrossRef]
- Thixton, H.L.; Esselman, E.J.; Corey, L.L.; Zettler, L.W. Further evidence of Ceratobasidium D.P. Rogers (Basidiomycota) serving as the ubiquitous fungal associate of Platanthera leucophaea (Orchidaceae) in the North American tallgrass prairie. Bot. Stud. 2020, 61, 12. [Google Scholar] [CrossRef] [Green Version]
- Zettler, L.W.; Piskin, K.A.; Stewart, S.L.; Hartsock, J.J.; Bowles, M.L.; Bell, T. Protocorm mycobionts of the Federally threatened eastern prairie fringed orchid, Platanthera leucophaea (Nutt.) Lindley, and a technique to prompt leaf elongation in seedlings. Stud. Mycol. 2005, 53, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Zettler, L.W.; Piskin, K.A. Mycorrhizal fungi from protocorms, seedlings, and mature plants of the eastern prairie fringed orchid, Platanthera leucophaea: A comprehensive list to augment conservation. Am. Midl. Nat. 2011, 166, 29–39. [Google Scholar] [CrossRef]
- Mapes, D.R. Soil Survey of DuPage and Part of Cook Counties; Illinois U.S. Department of Agriculture, Soil Conservation Service and Illinois Agricultural Experiment Station: Urbana, IL, USA, 1979. [Google Scholar]
- Anderson, R.C. Evolution and origin of the Central Grassland of North America: Climate, fire, and mammalian grazers. J. Torrey Bot. Soc. 2006, 133, 626–647. [Google Scholar] [CrossRef] [Green Version]
- Willman, H.B. Summary of the Geology of the Chicago Area. Circular 460; Illinois State Geological Survey: Urbana, IL, USA, 1971. [Google Scholar]
- Fehrenbacher, J.B.; Alexander, D.; Jansen, I.J.; Darmody, R.G.; Pope, R.A.; Flock, M.A.; Voss, E.E.; Scott, J.W.; Andres, W.F.; Bushue, L.J. Soils of Illinois. Bulletin 778; University of Illinois at Urbana Champaign, College of Agriculture, Experiment Station and Soil Conservation Service, U.S. Department of Agriculture: Urbana-Champaign, IL, USA, 1984. [Google Scholar]
- Bowles, M.L.; Jones, M.D. Repeated burning of eastern tallgrass prairie increases richness and diversity, stabilizing late successional vegetation. Ecol. Appl. 2013, 23, 464–478. Available online: https://www.jstor.org/stable/23441009 (accessed on 23 April 2021). [CrossRef]
- Figura, T.; Tylovia, E.; Jersakava, J.; Vohnik, M.; Ponert, J. Fungal symbionts may modulate nitrate inhibitory effect on orchid seed germination. Mycorrhiza 2021, 31, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Knapp, A.K.; Seastedt, T.R. Detritus accumulation limits productivity of tallgrass prairie. BioScience 1986, 36, 662–668. [Google Scholar] [CrossRef]
- Bentivenga, S.P.; Hetrick, B.A.D. Relationship between mycorrhizal activity, burning and plant productivity in tallgrass prairie. Can. J. Bot. 1991, 69, 2597–2602. [Google Scholar] [CrossRef]
- Van der Valk, A.G. Succession in wetlands: A gleasonian approach. Ecology 1981, 62, 688–696. [Google Scholar] [CrossRef] [Green Version]
- Bowles, M.L.; McBride, J.; Bell, T.J. Long-term processes affecting restoration and viability of the Federal Threatened Mead’s Milkweed (Asclepias meadii). Ecosphere 2015, 6, 11. [Google Scholar] [CrossRef]
- Fenster, C.B.; Galloway, L.F. Inbreeding and outbreeding depression in natural populations of Chamaecristata fasciculate (Fabaceae). Conserv. Biol. 2000, 14, 1406–1412. Available online: https://www.jstor.org/stable/2641793 (accessed on 25 April 2021). [CrossRef] [Green Version]
- Menges, E.S.; Dolan, R.W. Demographic viability of populations of Silene regia in midwestern prairies; relationships with fire management, genetic variation, geographic location, population size and isolation. J. Ecol. 1998, 86, 63–78. [Google Scholar] [CrossRef]
- Kaye, T.N.; Pendergrass, K.L.; Finley, K.; Kauffman, J.B. The effect of fire on the population viability of an endangered prairie plant. Ecol. Appl. 2001, 11, 1366–1380. [Google Scholar] [CrossRef]
- Liu, H.; Menges, E.S.; Quintana-Ascencio, P.F. Population viability analysis of Chaemaecrista keyensis: Effects of fire season and frequency. Ecol. Appl. 2005, 15, 210–221. [Google Scholar] [CrossRef] [Green Version]
- Oldfather, M.F.; Ackerly, D.D. Microclimate and demography interact to shape stable population dynamics across the range of an alpine plant. New Phytol. 2019, 222, 193–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeifer, M.; Wiegand, K.; Heinrich, W.; Jetschke, G. Long-term demographic fluctuations in an orchid species driven by weather: Implications for conservation planning. J. Appl. Ecol. 2006, 43, 313–324. [Google Scholar] [CrossRef]
- Raventós, J.; González, E.; Mújica, E.; Doak, D.F. Population viability analysis of the epiphytic ghost orchid (Dendrophylax lindenii) in Cuba. Biotropica 2015, 47, 179–189. [Google Scholar] [CrossRef]
- Crain, B.J.; Tremblay, R.L.; Ferguson, J.M. Sheltered from the storm? Population viability analysis of a rare endemic under periodic catastrophic regimes. Popul. Ecol. 2019, 61, 74–92. [Google Scholar] [CrossRef]
- Opler, P.A. Management of prairie habitats for insect conservation. J. Nat. Areas Assoc. 1981, 1, 3–6. Available online: https://www.jstor.org/stable/43908524 (accessed on 23 May 2021).
- Hodges, R.W. The Moths of America North of Mexico; E.W. Classey Limited and R.B.D. Publications Inc.: Middlesex, UK, 1971; p. 164. [Google Scholar]
- Panzer, R. Compatibility of prescribed burning with the conservation of insects in small, isolated prairie reserves. Conserv. Biol. 2002, 16, 1296–1307. Available online: https://www.jstor.org/stable/3095325 (accessed on 5 May 2021). [CrossRef] [Green Version]
- Panzer, R. Importance of in situ survival, recolonization, and habitat gaps in the postfire recovery of fire-sensitive prairie insect species. Nat. Areas J. 2003, 23, 14–21. [Google Scholar]
- Guerra, E.E.; Blanco, C.M.; Garrie, J. The impact of prescribed fire on moth assemblages in the Boston Mountains and Ozark Highlands, in Arkansas. J. Ark. Acad. Sci. 2019, 73, 9. Available online: https://scholarworks.uark.edu/jaas/vol73/iss1/9 (accessed on 5 May 2021).
- Young, B.E.; Auer, S.; Ormes, M.; Rapaccioulo, G.; Schweitzer, D.; Sears, N. Are pollinating hawk moths declining in the Northeastern United States? An analysis of collection records. PLoS ONE 2017, 12, e0185683. [Google Scholar] [CrossRef] [Green Version]
- Nurfadilah, S.; Swarts, N.D.; Dixon, K.W.; Lambers, H.; Merritt, D.J. Variation in nutrient-acquisition patterns by mycorrhizal fungi of rare and common orchids explains diversification in a global biodiversity hotspot. Ann. Bot. 2013, 111, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seastedt, T.R.; Ramundo, R.A. The influence of fire on below-ground processes of tallgrass prairie. In Fire in North American Tallgrass Prairies; Collins, S.L., Wallace, L.L., Eds.; University of Oklahoma Press: Norman, OK, USA, 1990. [Google Scholar]
- Jasinge, N.U.; Huynh, T.; Lawrie, A.C. Consequences of season of prescribed burning on two spring-flowering terrestrial orchids and their endophytic fungi. Aust. J. Bot. 2018, 66, 298–312. [Google Scholar] [CrossRef]
- Sharma, J.; Zettler, L.W.; Van Sambeek, J.W.; Ellerseck, M.; Starbuck, C.J. Symbiotic seed germination and mycorrhizae of federally-threatened Platanthera praeclara (Orchidaceae). Am. Midl. Nat. 2003, 149, 79–95. Available online: https://www.jstor.org/stable/3083178 (accessed on 19 March 2021). [CrossRef]
- U.S. Fish and Wildlife Service (USFWS). Five Year Review: Eastern Prairie Fringed Orchid (Platanthera leucophaea); Ft. Snelling: St. Paul, MN, USA, 2010. [Google Scholar]
- Alley, W.M. The Palmer Drought Severity Index: Limitations and assumptions. J. Clim. Appl. Meteorol. 1984, 23, 1100–1109. [Google Scholar] [CrossRef] [Green Version]
- Zettler, L.W. Terrestrial orchid conservation by symbiotic seed germination: Techniques and perspectives. Selbyana 1997, 18, 188–194. Available online: https://journals.flvc.org/seblyana/article/view/120576 (accessed on 10 May 2021).
- Hadley, G. Symbiotic germination of orchid seed. Orchid Rev. 1983, 91, 44–47. [Google Scholar]
- Zettler, L.W.; Hofer, C.J. Propagation of the little club-spur orchid (Platanthera clavellata) by symbiotic seed germination and its ecological implications. Environ. Exp. Bot. 1998, 39, 189–195. [Google Scholar] [CrossRef]
- Caswell, H. Matrix Population Models: Construction, Analysis, and Interpretation, 2nd ed.; Sinauer: Sunderland, MA, USA, 2001. [Google Scholar]
- Hood, G.M. PopTools: Software for the Analysis of Ecological Models. 2009. Available online: http://www.poptools.software.informer.com/3.2/ (accessed on 17 March 2011).
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Skalski, J.R.; Millspaugh, J.J.; Dillingham, P.; Buchanan, R.A. Calculating the variance of the finite rate of population change from a matrix model in Mathematica. Environ. Model. Softw. 2007, 22, 359–364. [Google Scholar] [CrossRef]
- Akçakaya and Root. RAMAS Metapop: Viability Analysis for Stage-Structured Metapopulations (Version 4.0); Applied Biomathematics: New York, NY, USA, 2002. [Google Scholar]
- Silvertown, J.; Franco, M.; Menges, E. Interpretation of elasticity matrices as an aid to the management of plant populations for conservation. Conserv. Biol. 1996, 10, 591–597. Available online: http://www.jstor.org/stable/2386874 (accessed on 25 March 2021). [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Growth Rate (λ) | 95% CL | Mean Persistence (years) | se | Mean Extinction Duration (years) | se | ||
|---|---|---|---|---|---|---|---|
| Pooled | 1.10529 | 0.25694 | 100 | 0 | 0 | 0 | |
| Burn effects | unburn | 1.10732 a | 0.05423 | 100 a | 0 | 0 a | 0 |
| burn | 1.63042 b | 0.03619 | 100 a | 0 | 0 a | 0 | |
| Drought Effects | drought | 0.62124 a | 0.12823 | 13.0 c | 0.30 | 87.0 c | 0.39 |
| normal | 1.10996 b | 0.11775 | 100 a | 0 | 0 a | 0 | |
| wet | 0.71422 a | 0.22808 | 15.9 b | 0.56 | 84.1 b | 0.56 | |
| Crossing effect | outcross within | 1.10529 b | 0.25694 | 100 a | 0 | 0 a | 0 |
| outcross between | 1.73496 a | 0.44092 | 100 a | 0 | 0 a | 0 | |
| self | 0.88622 c | 0.19454 | 27.1 b | 0.92 | 72.9 b | 0.92 | |
| Habitat type | wetland | 1.29147 a | 0.16234 | 75.0 a | 3.21 | 21.8 a | 3.10 |
| prairie | 0.83853 b | 0.27830 | 9.4 b | 0.44 | 90.6 b | 0.44 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bell, T.J.; Bowles, M.L.; Zettler, L.W.; Pollack, C.A.; Ibberson, J.E. Environmental and Management Effects on Demographic Processes in the U.S. Threatened Platanthera leucophaea (Nutt.) Lindl. (Orchidaceae). Plants 2021, 10, 1308. https://doi.org/10.3390/plants10071308
Bell TJ, Bowles ML, Zettler LW, Pollack CA, Ibberson JE. Environmental and Management Effects on Demographic Processes in the U.S. Threatened Platanthera leucophaea (Nutt.) Lindl. (Orchidaceae). Plants. 2021; 10(7):1308. https://doi.org/10.3390/plants10071308
Chicago/Turabian StyleBell, Timothy J., Marlin L. Bowles, Lawrence W. Zettler, Catherine A. Pollack, and James E. Ibberson. 2021. "Environmental and Management Effects on Demographic Processes in the U.S. Threatened Platanthera leucophaea (Nutt.) Lindl. (Orchidaceae)" Plants 10, no. 7: 1308. https://doi.org/10.3390/plants10071308