
Phytocannabinoids Biosynthesis in Angiosperms, Fungi, and Liverworts and Their Versatile Role


1
Department of Botany, Plant Physiology Section, Faculty of Life Sciences, Aligarh Muslim University, Aligarh 202002, India
2
Department of Biology and Plant Ecology, Faculty of Biology, University of Bialystok, Ciolkowskiego 1J, 15-245 Bialystok, Poland
*
Author to whom correspondence should be addressed.
Plants 2021, 10(7), 1307; https://doi.org/10.3390/plants10071307
Submission received: 6 June 2021
/
Revised: 22 June 2021
/
Accepted: 25 June 2021
/
Published: 28 June 2021
(This article belongs to the Special Issue 10th Anniversary of Plants—Recent Advances and Perspectives)
Abstract
:Phytocannabinoids are a structurally diverse class of bioactive naturally occurring compounds found in angiosperms, fungi, and liverworts and produced in several plant organs such as the flower and glandular trichrome of Cannabis sativa, the scales in Rhododendron, and oil bodies of liverworts such as Radula species; they show a diverse role in humans and plants. Moreover, phytocannabinoids are prenylated polyketides, i.e., terpenophenolics, which are derived from isoprenoid and fatty acid precursors. Additionally, targeted productions of active phytocannabinoids have beneficial properties via the genes involved and their expression in a heterologous host. Bioactive compounds show a remarkable non-hallucinogenic biological property that is determined by the variable nature of the side chain and prenyl group defined by the enzymes involved in their biosynthesis. Phytocannabinoids possess therapeutic, antibacterial, and antimicrobial properties; thus, they are used in treating several human diseases. This review gives the latest knowledge on their role in the amelioration of abiotic (heat, cold, and radiation) stress in plants. It also aims to provide synthetic and biotechnological approaches based on combinatorial biochemical and protein engineering to synthesize phytocannabinoids with enhanced properties.
1. Introduction
Phytocannabinoids are meroterpenoids bearing a resorcinol core with an isoprenyl, alkyl, or aralkyl para-positioned side chain, or alkyl group usually containing an odd number of carbon atoms—cannabinoids that have an even number of carbon atoms in the side chain are rare. Phytocannabinoids can be obtained from angiosperms (flowering plants), fungi, and liverworts (Figure 1). The first phytocannabinoid was isolated from the Cannabis sativa family Cannabaceae, but it has a long controversial history of its use and abuse [1,2]. From C. sativa more than 113 phytocannabinoids were isolated and classified into several groups such as cannabidiols (CBDs), cannabigerols (CBGs), cannabicyclols (CBLs), cannabidiols (CBNDs), cannabinols (CBNs), cannabitriols (CBTs), cannabichromenes (CBCs), (−)-Δ9-trans-tetrahydrocannabinol (Δ9-THC) and miscellaneous cannabinoids [1,3,4,5]. Compounds obtained from C. sativa predominately generate alkyl-type phytocannabinoids with a monoterpene isoprenyl and the pentyl side chain [4,6]. In C. sativa, CBD, CBG, CBC, cannabichromevarine (CBCV), and Δ9-THC are the most abundant cannabinoids in their respective acidic form. The acidic form of the cannabinoid (C22, “pre-cannabinoids”) is the final step of the cannabinoid biosynthetic pathway. Oxidation, decarboxylation, and cyclization lead to the development of modified phytocannabinoid via spontaneous breakdown or conversion product. The conversion mainly occurs due to the poor oxidative stability of phytocannabinoids, especially with the alkyl group. C. sativa produces the most common phytocannabinoids. In addition to this, the brains of mammals have receptors that respond to the C. sativa cannabinoid, so they were termed as cannabinoid receptor types 1 and 2 (CB1R and CB2R) and thus participated in the endocannabinoid system [1,3,4,7,8].
The Endocannabinoid system in humans and animals revealed that it participates in the regulation of biological functions such as memory, brain system, mood and addiction along with cellular and metabolic processes such as glycolysis, lipolysis, and the energy balance system [7,9]. Other angiosperms such as Helichrysum umbraculigerum (Asteraceae) native to South Africa, Amorpha fruticosa (Fabaceae), and Glycyrrhiza foetida (Fabaceae) contains a bioactive compound bearing a cannabinoid backbone (Figure 2); they are characterized as prenylated bibenzyl derivatives because the aralkyl side chain occurs [1,10].
Many Rhododendron species (family Ericaceae) such as Rh. dauricum native to Northeastern Asia, Rh. adamsii found in Eastern Siberia and Mongolia, Rh. rubiginosum var. rubiginosum native to Southwest China, and Rh. anthopogonoides grown in Southern China, all generate active monoterpenoids that have a cannabinoid backbone. Phytocannabinoids are CBC types with an orcinol or methyl group side chain (Figure 3). Rh. dauricum particularly produces cannabinoids bearing sesquiterpene moiety such as daurichromenic acid (DCA), grifolic acid (GFA), confluentin, and rhododaurichromenic acid [11,12,13]. Rh. adamsii produces cannabigerorcinic acid, DCA, cannabigerorcinic acid methylase, chromane, and chromene monoterpenoids; Rh. rubiginosum produces cannabinoid rubiginosins A–G [14,15]. Rh. anthopogonoides contains chromane/chromene derivatives such as cannabiorcicyclolic acid, cannabiorcichromenic acid, anthopogochromenic acid, and anthopogocyclolic acid (Figure 3) [16].
Liverworts, such as Radula marginata, R. perrottetii, and R. laxirameae native to New Zealand, produce active cannabinoids with bibenzyl backbones such as lunularic acid and its dimeric form—vittatin (Figure 4) [17,18,19,20]. Some fungi, e.g., Albatrellus (Albatrellaceae, mycorrhizal fungi) species, also produce GFA along with its derivative confluentin, grifolin, and neogrifolin (Figure 4). Additionally, Cylindrocarpon olidum generates cannabiorcichromenic acid and halogenated cannabinoid, i.e., 8-chlorocannabiorcichromenic acid (Figure 4) [1,21].
This review focuses on the biosynthesis of different active phytocannabinoids in several cellular compartments of C. sativa, Rhododendron, and Radula species. In this topic framework, the most crucial criterion is the synthetic and biotechnological techniques for the production of phytocannabinoids. The current review highlights the multi-faceted role of different active phytocannabinoids in humans and plants. Interestingly, this review briefly highlights the antimicrobial, antibacterial, and antibiotic properties of phytocannabinoid based on recent papers. Additionally, the role of phytocannabinoids in ameliorating pathogenic attack, and environmental stresses, e.g., cold, heat, and UV radiation, is also briefly assessed.
2. Phytocannabinoid Biosynthesis Sites
In C. sativa, phytocannabinoids are stored in glandular trichomes, located all over the aerial part of the plant, so root surface and root tissues do not keep phytocannabinoid. Female flowers possess a high density of phytocannabinoid [22,23]. Glandular trichomes have balloon shaped secretory vessicle which store cannabinoid. High temperature or herbivory leads to trichome rupture, which releases the sticky contents on the plant parts with viscous and non-crystallizing properties [24,25]. Higher temperatures increase cannabinoid production. Furthermore, cannabinoid production is raised in the cannabis flower after UV-B exposure. Nevertheless, phytocannabinoids act as a sun shield that absorbs lethal UV radiation [26]. Rhododendron genus lepidote consists of small leaves surrounded by glandular scales on both the abaxial and adaxial surfaces. These scales have lipophilic globules that contain major bioactive compounds such as cannabinoids, terpenes; the apoplastic space of the glandular scale also contains cannabinoids such as DCA in the Rhododendron [27]. Liverworts have oil bodies that are membrane-bound cellular structures that contain cannabinoids, aromatic oil, and terpenoid (cis configuration), mostly sesquiterpenoids and diterpenoids. Oil bodies are odiferous bitter, pungent compounds, which make them biologically active. Furthermore, these possess several ecological advantages such as tolerance from temperature, light, or radiation [28,29].
3. Biosynthesis of Phytocannabinoids
This section focuses on detailed events undergone in the production of several phytocannabinoids inside C. sativa, Rhododendron, and liverworts. Moreover, biosynthesis of phytocannabinoids via biotechnological approaches in a heterologous host and synthetic methods are discussed.
3.1. Cannabis sativa
Phytocannabinoids are prenylated polyketides, i.e., terpenophenolic compounds, which are derived from isoprenoid and fatty acid precursors. Phytocannabinoid biosynthesis occurs in different cellular compartments: gland cells cytosol, the plastids, and the extracellular storage cavity. In the cytosol, oxidative cleavage of fatty acid such as palmitic acid yields hexanoic acid; it further synthesizes olivetolic acid (OA). The next step is prenylation of phenolic moiety (the polyketide derivatives, 5-pentenyl resorcinolic acid, and OA) with the terpenoid geranyl pyrophosphate (GPP). This step originates from the methylerythritol-4-phosphate (MEP) pathway in plastids. Cyclization (oxidative) and storage of the final products take place outside the gland cells. Transport proteins and vesicle trafficking participate in mobilizing intermediates across the morphologically highly specialized interface between the gland cells and storage cavity [30,31,32].
In C. sativa, phytocannabinoids biosynthesis is divided into three important places: cytosol (for polyketide pathway), plastids (MEP pathway for prenylation), and apoplastic spaces (oxidocyclization and storage) (Figure 5). Inside cytosol, biosynthesis of phytocannabinoids participates in the integration of major steps in polyketide and isoprenoid metabolism. Fatty acids (C18) are sequentially desaturated, peroxygenated, and cleaved into the hexanoic acid (C6) and C12 product via enzyme desaturase, lipoxygenase (LOX), and hydroperoxide lyases, respectively. Hexanoic acid is converted into thioester hexanoyl-CoA; this reaction is catalyzed by acyl-activated enzyme 1 (AAE1). Later, hexanoyl-CoA and malonyl-CoA (C2 donor) together via the action of olivetol synthase (OLS) and olivetolic acid cyclase (OAC) synthesizes OA. Moreover, it was reported that OAC is the dimeric (α+β) barrel protein, it is the first plant enzyme that catalyzes (C2-C7) intramolecular aldol condensation along with carboxylate retention; OAC contains distinctive active-site bearing the pentyl-binding hydrophobic pocket and polyketide binding site, whereas it is devoid of aromatase and thioesterase activities [31,33,34].
Inside plastids, the MEP pathway synthesizes GPP. It prenylates OA, which forms the intermediate branch-point and first cannabinoid which is cannabigerolic acid (CBGA), and this reaction is catalyzed by cannabigerolic acid synthase (CBGAS). CBGA is an essential cannabinoid because it acts as the precursor of several cannabinoids with an alkylic pentyl side chain. In contrast, CBGAS is a transmembrane aromatic prenyltransferase (PT) that transfers plastid signals. Then CBGA is converted into Δ9-THCA and cannabidiolic acid (CBDA) with the help of two enzymes which are CBDAS and Δ9-tetrahydrocannabinolic acid synthase (Δ9-THCAS). This conversion continues by reducing oxygen (O2) into hydrogen peroxide (H2O2) via oxidative cyclization reactions. Additionally, CBDAS and Δ9-THCAS are necessary flavoprotein enzymes that are dependent on O2 (electron acceptor) [1,30,31].
Another important enzyme, cannabichromenic acid synthase (CBCAS), dependent from FAD and O2, takes part in the synthesis of cannabichromenic (CBCA). Additionally, enzymes Δ9-THCAS and CBCAS have high sequence similarity at about a 96% nucleotide level. Both remain active inside resin space, which shows that CBCAS participates as an O2 dependent flavoprotein that converts CBGA to cannabichromenic acid (CBCA) with H2O2 as the side product via an oxidocyclization reaction. Active cannabinoid Δ9-THCA, CBDA, and CBCA with a pentyl side chain are synthesized in the apoplastic cannabis space. Furthermore, these active phytocannabinoids undergo decarboxylation and spontaneous rearrangement reactions on exposure to heat, radiation, or during storage. Some phytocannabinoid having unknown C1-C4 alkyl side chain are synthesized from acetyl-CoA, propanoyl-CoA, or pentanoyl-CoA [1,5,31,35].
3.2. Rhododendron
DCA and its derivative are produced and stored inside specialized glandular scales in Rh. dauricum. DCA utilizes carbon atoms from acetyl-CoA and farnesyl-CoA, with two significant intermediates, i.e., orsellinic acid (OSA) and GFA. DCA biosynthesis in Rhododendron is split between the cytosol, plastid, and apoplastic spaces (Figure 6) [11,36].
The biosynthesis of DCA starts in the cytosol with polyketide formation, type III polyketide synthase (PKS) helps in acetyl-CoA chain extension. Then another enzyme, orcinol synthase (ORS), catalyzes orcinol, OSA, triacetic acid, tetracetic acid, lactone and phloroacetophenone, where malonyl-CoA (three units) act as a carbon donor. Furthermore, tetraketide cyclase catalyzes OSA from ORS [36,37]. OA is transported to the plastid via a transporter, which is still unknown. Inside the plastids, the MEP pathway derives farnesyl-CoA. The inhibition of the MEP pathway via clomazone decreases OSA and DCA synthesis. In contrast, inhibition of the mevalonate-dependent pathway via mevastatin led to an increment in OSA and DCA biosynthesis. Moreover, aromatic farnesyltransferase Rh. dauricum prenyltransferase (PT) helps in regiospecific farnesylation; this enzyme moderates sequence identity with UbiA aromatic PTs that lie within chloroplasts. Geranyl-CoA and geranylgeranyl-CoA serve as the alternative prenyl donors used by PT, but their activity rate is 13%, and 2.5% of the activity acquired by farnesyl-CoA. GFA is synthesized as the intermediate within the plastids. Then, within apoplastic spaces, an oxidocyclization reaction takes place via DCA synthase (DCAS) forming CBC scaffold; reaction moves forward by H2O2 release. Like Δ9-THCAS and CBDAS, DCAS is active enzymatically outside apoplastic spaces and dependent on O2. In Rh. dauricum, DCA decarboxylated forms produce confluentin; spontaneous decarboxylation occurs via heat, irradiation, and during storage, similar to the decarboxylation acidic to neutral phytocannabinoids in C. sativa trichomes [5,11,37].
Apoplastic spaces serve as storage for many metabolites, essential oils, DCA, and confluentin. Moreover, GFA and DCA act as phytotoxic compounds in Rh. dauricum cell culture, as they induce cell death. Similarly, H2O2 formed as a side product in DCA biosynthesis also increases cell death by enhancing apoptosis-related reactions. However, to overcome cell death, autotoxicity, and cell damage, DCA storage occurs in the apoplast, and H2O2 is released to participate in the plant-defense system and provide plant immunity. In Rh. dauricum, transport proteins and vesicle trafficking mechanisms are still not well understood and remain a valuable and exciting approach for further future investigations [1,2,37].
3.3. Liverworts
It was reported that Radula marignata possesses enzymes for GPP biosynthesis and helps in the biosynthesis of bibenzyl cannabinoid. Moreover, the production of bibenzyl CBGA analog (i.e., lunularic acid, perrottetinenic acid, and perrottetinene) needs precursor stilbene acid or dihydrostilbene acid, which is very rare, and compounds of this type were found in Hydrangea macrophylla var. thunbergii and liverworts such as Marchantia polymorpha and Convolvulus hystrix (Figure 7) [1,17,38].
Stilbene acid is synthesized from type III PKS via coumaroyl-CoA or dihydrocoumaroyl-CoA, CoA-activated precursors. Starter molecules are extended by malonyl-CoA with decarboxylation, followed by a condensation reaction, which produces polyketide intermediate, which synthesizes different core structures. Hydrangic acid is the starter molecule that acts as coumaroyl-CoA. It is extended utilizing malonyl-CoA (three units) as a C2 donor to synthesize tetraketide intermediate. These reactions are catalyzed via stilbene synthase (STS)-type PKS enzymes. Ketoreductase (KR) leads to polyketide reduction followed by STS-like C2 to C7 intramolecular aldol condensation, here retention of the carboxylic group produces hydrangic acid. KR is involved in the loss of the C5-hydroxyl group on the aromatic ring structure of hydrangic acid, contrary to the stilbene acid structure. In R. marginata, the precursor of stilbene acid or dihydrostilbene is derived from the coumaroyl CoA or cinnamoyl-CoA; type III PKS enzyme helps in chain elongation, later putative tetraketide cyclase or (dihydrostilbene acid cyclase, DHAC) helps in cyclization [1,18,38,39].
The lunularic acid precursor, prelunularic acid is produced by the type III PKS, named bibenzyl synthase (BBS). Moreover, BBS catalyzes the reaction where dihydrocoumaroyl-CoA serves as the starter molecule for the extension utilizing malonyl-CoA (3-units), which serves as the carbon donor. Later, cyclization occurs on reduced polyketide, with a lack of C5-hydroxyl group on the aromatic ring structure. Furthermore, type III PKS plays a crucial role in the bibenzyl cannabinoid and its analog synthesis by also catalyzing the carboxylate retaining reaction mechanism. KR is involved in the cyclization and assists proper ring formation. Thus, after cyclization, lunularic acid is synthesized [40,41,42].
Perrottetinenic acid (PA) is synthesized in R. marginate [43]. The transcriptomic approach of liverworts bears the mRNA encoding for type III PKS (responsible for chain elongation), which were later recognized as STS. Furthermore, it also exhibits a 60% homology of the amino acid sequence to stilbene-carboxylate synthase in Marchantia polymorpha. Other enzymes, such as double-bond reductase (DBR)—aromatic PT and oxidocyclase (perrottetinenic acid synthase, PAS), are also responsible for the production of PA. DBR catalyzes compounds that are precursors of phenylpropanoid, and generates dihydrocinnamoyl-CoA. The production of bibenzyl phytocannabinoid is dispersed across the liverworts cells in the same way as in cannabis and Rhododendron. Therefore, DHAC and DBR reside in the cytosol, PT is localized in the plastids and PAS inside the oil body. Signal peptides act as the crucial indicators for the encoding genes selection [18,39,41,44]. Another bibenzyl cis-THC, (−)-cis-perrottetinene (cis-PET) was isolated from the liverwort R. perrottetii [45]. The cis configuration in the cyclohexene ring in cis-PET is comparable with Δ9-trans-THC. PET resembles Δ9-THC in its 3D shape, and can bind to many of the same cannabinoid receptors (CBRs) as Δ9-THC. Interestingly, PET also reduces the level of prostaglandins in the brain—a compound with inflammatory properties that increase in response to Δ9-THC and may be responsible for adverse effects [17].
3.4. Application of Biotechnological Approaches to Phytocannabinoids Production in Heterologous Hosts
Phytocannabinoids can be generated in different heterologous hosts such as fungi, bacteria, and plants with the help of biotechnological techniques. Several investigations reported that in vitro culture of phytocannabinoids biosynthesis in C. sativa, with the help of explants and micropropagation, is a widely used biotechnological approach for phytocannabinoid production. Moreover, apart from C. sativa, Nicotiana benthamiana also emerged as the favorable heterologous host for the production of phytocannabinoids. It exhibits the production of several proteins and bioactive compounds and has glandular trichomes that help overcome cell death, autotoxicity, and cell damage caused due to intermediates produced during phytocannabinoids biosynthesis. In recent research, it was found that a major biotechnological tool for phytocannabinoids production and inducing genetic modification is established via the micropropagation technique. Additionally, cell suspension culture, hairy root, and adventitious root culture also produce a small quantity of cannabinoids. In Saccharomyces cerevisiae, galactose produces phytocannabinoids such as CBG, CBDA, Δ9-THCA, and minor phytocannabinoids such as cannabidivarinic acid (CBDVA) and Δ9-tetrahydrocannabidivarinic acid (Δ9-THCVA) [46,47,48,49,50,51].
3.5. Production of Phytocannabinoids through Synthetic Approaches
Phytocannabinoids are terpenophenolic compounds that are produced by polyketide and the MEP pathway. In the heterologous biosynthesis approach, two important pathways that are optimizing and engineering provide abundant precursors for cannabinoid production. Additionally, these pathways are linked via aromatic PT, ubiquitous in plants, animals, bacteria, and fungi [52,53]. Thus, this helps in the production of aromatic metabolites such as coumarin, flavonoid, and phenylpropanoid at different reaction spectra [54]. As discussed, biosynthesis of phytocannabinoid is different in plants, fungi, and liverworts; thus, the recent techniques which emerge out to be beneficial is by using the aromatic PT-based approach to generate novel phytocannabinoids in the heterologous hosts formed on combinational utilization of module over several species [52,53,54].
Similar to Humulus lupulus, inside trichomes of C. sativa, chalcone isomerases such as proteins (CHILs) are expressed. Additionally, CHILs are polyketide binding proteins, and their co-expression in heterologous cannabinoid production mechanism plays a pivotal role in augmenting biosynthesis. Apart from combinatorial biochemistry techniques, enzyme-based approaches also play a remarkable role in phytocannabinoid production. Furthermore, some non-natural precursors such as pentanoic acid, hexanoic acid, heptanoic acid are incorporated inside the generated cannabinoid. Derivatization of phytocannabinoid diversifies their functionality. Glycosylation of phytocannabinoid reduces cell damage and autotoxicity and increases cell stability and life; in a therapeutic context, glycosylated phytocannabinoid enhances absorption, distribution, metabolism and, excretion (ADME) features. Halogenation of cannabinoids (like DCA) by co-expression of halogenase AscD derived from Fusarium exhibits several antifungal, antibacterial, antitumor, antiparasitic properties. Thus, the synthetic biology technique serves as a new insight into the production and designing of phytocannabinoids [55,56,57,58].
4. Phytocannabinoid Storage and Maintenance of Cell Homeostasis
The last step of phytocannabinoid biosynthesis and their storage both occur in different cellular compartments like in C. sativa storage organs are resin present inside glandular trichomes, in Rh. dauricum they storage takes place inside apoplast of glandular scale while in R. marginata oil bodies are the storage organs [1]. In C. sativa, nuclear magnetic resonance (NMR) based metabolomics of trichomes revealed storage of phytocannabinoids along with terpenes, sugars, organic acid, amino acid, and choline [6]. Sugars, organic acids, and choline, which present in appropriate stoichiometric ratios, give rise to natural deep eutectic solvents (NADES) [59]. NMR-based techniques revealed that constituents for NADES display proton-associated intermolecular interactions, forming aggregates to form larger structures in the liquid phase [60]. NADES is a distinguished solvent bearing this property when the water content is less than 40%; this unique property of NADES incorporates them in the third membrane-less solvent phase inside biological systems [61,62]. Δ9-THCA and CBDA support NADES mediated solubilization in trichomes and oil bodies as they are virtually insoluble in water. In C. sativa and Rh. dauricum, oxidocyclase releases H2O2. Furthermore, it participates in a plant-defense system and provides plant immunity. NADES stabilizes the activity of oxidocyclases, whereas it also serves as a biological solvent where the biosynthetic catalytic enzyme remains active and functional. NADES plays a crucial role in a plant cell to maintain cell homeostasis. Thus, it plays a chief role in stabilizing phytocannabinoid biosynthesis enzymes, their storage and further induces cellular homeostasis. Recently, cytochrome P450 (CYP) enzymes, CYP79A1, CYP71E1, and P450 oxidoreductase involved in the biosynthesis of the cyanogenic glucoside dhurrin, which was stabilized NADES. Biosynthesis and appropriate storage of cannabinoids in heterologous hosts need to be engineered to reduce autotoxicity and apoptosis [63,64].
5. Biotechnology and In Vitro Propagation of Cannabis
Cannabis is an essential plant as it yields phytocannabinoid, which has multiple applications. Cannabis cultivation is regulated in several countries, so alternative biotechnological and in vitro tissue culture approaches require attention. Meanwhile, these approaches are beneficial to preserve cultivars or clones of plants with specific metabolites such as phytocannabinoid. Micropropagation generates genetically stable plants; thus, this method is beneficial for the clonal multiplication of cannabis. Nevertheless, micropropagation of the cannabis plant is performed via adventitious buds and axillary buds located on the nodal segment. Another method that is synthetic seed technology (encapsulation of axillary bud or nodal segment in calcium alginate seeds) has also gained importance in plant clonal propagation. It displays homogenous growth and development of plants and genetic stability even after long storage [65,66].
Cannabis is not a well-known recalcitrant plant for transformation. The plant’s regeneration capacity is low and is completely dependent on plant tissue, age, explants, and a combination of plant growth regulators (PGRs), i.e., indole-3-acetic acid (IAA), indole-3-butyric acids, 1-naphthaleneacetic acid (NAA), 2,4-dichlorophenoxyacetic acid, and kinetin. Moreover, successful transformation is performed via Agrobacterium tumefaciens; cells that do not regenerate the shoots are undifferentiated and can be transformed via phosphomannose isomerize and calorimetric assays, which showed efficacious expression of the transgene [67]. Cannabis transformation using shoot tip explants regenerates a high amount of cannabinoid after infection with Agrobacterium tumefaciens [68,69]. For the successful regeneration of cannabis, thidiazuron (behavior as cytokinin-like compound) with kinetin (represent cytokinins) has not only the effect on the shoots but also shows a high yielding capacity of phytocannabinoids. Dicamba (3,6-dichloro-2-methoxybenzoic acid) herbicide is also known for inducing the regeneration of shoots from calli. Moreover, a most potent and efficient transformation of cannabis was carried out using 1–2 cm of hypocotyl explants and by supplementation of zeatin and 6-benzylaminopurine [65].
Another important tissue culture technique used for the production of bioactive cannabinoids showing pharmaceutical effects is hairy root culture. In this system, both Agrobacterium tumefaciens and Agrobacterium rhizogenes are used for transformation to yield cannabinoids by establishing a hairy root system. Meanwhile, the most responsive tissue for infection in this system is hypocotyl, which produces large quantities of hypocotyls or engineered plants for the production of bioactive compounds that are industrially valuable; for example, Δ9-THCA is produced by expressing Δ9-THCAS. The hairy root technique is obtained by the increased growth rate, independent of a hormone, and has the same metabolic potential as the original plant organ. In the hairy root system, cannabis callus was cultured on a full-strength B5 medium feed with an NAA or IAA displayed increase in the accumulation of phytocannabinoids. Rhizogenesis induction in cannabis undifferentiated cells is crucial, as it can be performed on calli overexpressing some important transcription factors (TFs) and genes which are involved in cannabinoid synthesis. Additionally, cannabis hairy root culture is an efficient method implemented with adsorbents to reduce toxicity problems [65,67].
Cannabis cell suspension culture transformation with the genes involved in the cannabinoid synthesis pathway offers a great opportunity to enhance bioactive phytocannabinoid production possessing pharmacological potential. Additionally, increased production via cannabis cell suspension culture can be obtained through TF. TF have cascade mechanism (that is when the master regulator of cannabinoid synthesis is identified; there it can be expressed inducibly or constitutively inside cell suspension culture). Thus, TF constitutes potent and efficient tool involved in plant metabolic engineering [65].
TF, belonging to the MYB family, was involved in cannabinoid production and tolerant oxidative stress. TF expression in an inducible manner confers toxicity tolerance caused due to the accumulation of phytocannabinoids during the growth of transformed cells. Genetic engineering and plant cell suspension culture were elicited to induce the production of specific cannabinoids. Additionally, in cannabis suspension cells, induction with biotic and abiotic elicitors did not cause an increment in phytocannabinoid synthesis. Non-essential elements, such as silicon, play a key role in mitigating biotic and abiotic stress. Additionally, silicon has a stimulatory effect on the production of cannabis secondary metabolites like cannabinoids. Cyclodextrins are cyclic oligosaccharides bearing five or more than five α-D-glucopyranose residues; these form an inclusion complex structure with lipophilic compounds like cannabinoids. In plant cell suspension culture, cyclodextrins are used for the production of non-polar compounds such as stilbenes and other cannabinoids. Moreover, cyclodextrins improve cannabinoid and other metabolite solubilities in an aqueous environment. Cyclodextrins possess a similar structure to alkyl-derived oligosaccharide released from the plant cell wall during a fungal infection; thus they act as elicitors for the production of secondary metabolites. More investigations are needed on the effect of cyclodextrins on the synthesis of non-polar cannabinoids in cannabis suspension cultures [65,70].
6. Phytocannabinoids and Their Derivative Role and Bioactivity in Humans and Animals
Phytocannabinoids are used as medicines since antiquity. They possess antimicrobial, antibacterial, and antifungal activity and serve as remarkable antibiotics (Table 1). Additionally, cannabinoids show little antifungal power because fungi can metabolize cannabinoids except a few, such as Phomopsis ganjae. An increasing number of developing countries are relaxing their legislation around phytocannabinoids and cannabis-derived products; legalized cannabis-derived product industries are increasing worldwide. They are likely to constitute a projected US$ 57 billion market by the year 2027 [71].
6.1. Cannabinoid Receptors in Humans and Their Role
CB1R (cannabinoid receptor type 1, first cloned in 1990) mRNA is highly expressed in the brain. It is also found in the heart, lung, ovary, adrenal gland, thymus, testes, prostate, tonsils, bone marrow, and uterus. CB1R is also widely distributed inside non-neuronal tissues and inside various cells, tissues and also co-exist with other CBRs [86,87]. CB1R is found inside the brain and displays high protein density and expression inside brain parts such as the cerebellar molecular layer, substantia nigra pars reticulate, globus pallidus external and internal, olfactory bulb, olfactory nucleus (anterior), hippocampus and layers II–III, Va and VI of the cerebral cortex. Additionally, in humans, the highest CB1R level is found inside the cingulated gyrus, motor cortices and frontal secondary somatosensory. CB1R is found in moderate levels inside the hypothalamus and ventral striatum and low levels in the brainstem; other respiratory control centers lack CB1R. Moreover, this is the main reason CB1R has a low effect on respiratory and cardiovascular activities [88]. Another important receptor, CB2R, has high density inside immune cells and tissues, whereas a low amount is expressed inside the brain. Inside the brain, most of the presynaptic CBRs are observed as CB1R, whereas it was displayed that presynaptic inhibitory CB2R in GABAergic terminals of the hippocampus [86,87]. Phytocannabinoid applies its strong physio-psychotropic and psychotogenic actions along with the modulation of fast synaptic transmission in the brain showing its action on receptor CB1R, present in the synaptic region. Additionally, fast synaptic activity encloses signaling via GABA and glutamate (activate ionotropic receptors). However, CB1R contributes to psychoactivity and neurophysiology along with phytocannabinoid [89]. CB2R is found inside pyramidal cells (II/III) of the medial prefrontal cortex; when it activates, it causes IP3-dependent opening of calcium-activated chloride channels and ultimately resulted in the inactivation of neuronal firing [90,91].
G-protein-coupled receptors (GPR), such as GPR18 and GPR55, serve as potent phytocannabinoid receptors. GPR18 is a “deorphanized” receptor present in cells and tissues of the thymus, spleen, small intestine, lymph nodes, leukocytes, and gametes [92,93,94,95]. Phytocannabinoids (e.g., Δ9-THC) are agonists to GPR18. It plays a potent role in signalizing toxin-sensitive G-protein, Akt, PI3K, and p42/44 mitogen-activated protein kinase [96,97]. Additionally, cannabinoid (e.g., Δ9-THC) induces β-arrestin recruitment in the cells which are transfected with GPR18 [86,95]. GPR55 is the important “deorphanized” metabotropic receptor that interacts with several active phytocannabinoids; it is present in the central nervous system (CNS), adipose tissues, adrenal gland, immune cells, small intestine, osteoblasts and osteoclast [98,99]. GPR55 displays a low sequence with both CB1R and CB2R (about 15%); GPRs interact with Gα12 or Gα13, which results in activation of several pathways such as Rho-associated protein kinase, Ras homolog gene family member A (RhoA), p38, MAPK/ERK pathways [91,100]. Some phytocannabinoids potently activate GPR5. Moreover, GPR55 is the most complex CBR. The same ligand acts as an agonist, antagonist, or neutral; it also possesses the allosteric modulator site [86,87]. Phytocannabinoids serve as weak and partial agonists on the GPR55, but via the help of some cannabinoids (CBD, cannabidivarine, CBDV), they inhibit lysophosphatidylinositol linked GPR55 activation. Thus, GPR55, along with cannabinoids, can participate in curing cancer cells, obesity, inflammation, neuropathic pain, osteoporosis, and neuromodulation. Transient receptor potential (TRP) superfamily channels have 27 polymodal sensor cation channels classified into six broad types in humans [101]. Among these six families, four groups interact with cannabinoids and other derivatives. Thus, these participate in boosting the immune system, with analgesia (pain reliever), and nociception (processing of sensory nervous system); they also modulate inflammations and pain sensations [102,103,104].
Phytocannabinoids exerts strong therapeutic potential in humans assessed by the meta-analysis of several clinical trials; it possesses a strong effect on health conditions such as nausea, vomiting, insomnia (sleeping disorder), depression, anxiety, paraplegia (paralyzes in a lower limb due to spinal cord injury), psychosis (emotional metal disorder), appetite stimulation in several syndromes [86]. Additionally, cannabinoids play a significant role in ailments of human diseases and syndromes such as Tourette syndrome (nervous system disorder such as continuous movements), AIDS, and treating intraocular pressure of the eye in glaucoma [105]. The endocannabinoid system (internal lipid retrograde neurotransmitters which bind on CBR) along with phytocannabinoid is involved in modulating pharmacological, physiological, biological, and cognitive processes [106]. Due to the limited space, in this review, we only name diseases and ailments cured by phytocannabinoids that are liver encephalopathy, hepatic disease, asthma, respiratory tract changes, bronchospasm, bone remodeling and metabolism, osteoarthritis, and osteoporosis. Phytocannabinoids may also be used to treat diseases related to CNS such as brain trauma, stroke, brain aging, neuroinflammation, and neurodegradation. Additionally, they have served as an anti-solid depressant to cure patients with suicidal tendencies; apart from all this, they are a strong pain reliever and immunomodulator. Phytocannabinoids are also given to pregnant women to avoid miscarriage and preimplantation embryo development [86,105,107].
Phytocannabinoids have several medicinal properties in curing diseases and disorders and assist as an important template for chemists to form novel and synthetic medicine [108]. Several investigations display the potential of synthetic cannabinoids in human ailments. Δ9-THC and CBD together or alone form synthetic cannabinoids such as nabiximols, Sativex®, nabilone, dronabinol, levonantradol, and other synthetic Δ9-THC-analogs. These synthetic cannabinoids are used to cure cancer pain, neuropathic pain, and spasticity caused by sclerosis [75,105].
Phytocannabinoids have diverse therapeutic potential, but they have several adverse risks, mainly due to fewer trials. Certainly, there is a necessity to support research and investigation on phytocannabinoid legally and ethically; globally, the legalization of cannabinoids encounters several controversies by society, researchers, and health practitioners. Many countries such as Canada and the US also support phytocannabinoid production for medical use. Colorado’s state is employing private support from the medical cannabinoid (marijuana) industry to endorse future research on phytocannabinoids and cannabis [108,109].
6.2. Bioactivity of Phytocannabinoids
Phytocannabinoids are of great clinical use in humans and mammals, but the plant sources do not accumulate at high levels and are also regarded as endangered species. This enhances the demand for the biosynthesis of phytocannabinoids via biotechnological approaches to understand their bioactivity and role in medication. The prerequisite and establishment for such biotechnological approaches should be proper and deep knowledge of the pathways and mechanisms involved in phytocannabinoid production in C. sativa, liverworts and Rhododendron species [1,2,78,105].
6.2.1. Neutral Cannabinoids
In humans, phytocannabinoids play a potent role in signal transduction. They show crucial interaction with the G-protein coupled cannabinoid receptors (GPCRs such as CB1R and CB2R), peroxisome proliferator-activated receptor γ (PPARγ), and TRP ion channel. CB1R is the most abundant GPCR located in CNS, whereas CB2R is found in immune cells and tissues. Thus, cannabinoids play a crucial role in signaling, immune, and CNS proper functioning [72,74]. In humans, Δ9-THC displays pleiotropic effects such as analgesic (relieve pain), relaxation, pain tolerance, and dysphoria (anxiety disorder); thus this reveals Δ9-THC displays agonistic effects by the activation of CB1R of β-arrestin 2 recruitment and signaling; also Δ9-THC is the essential psychoactive (alters mood, behavior, feeling and thoughts by affecting CNS) bioactive constituent [73,104]. Δ9-THC is found in the drug dronabinol (Marinol®), and the constituent of sesame oil plays potent antiemetic (prevents vomiting) for cancer patients receiving chemotherapy. Additionally, it also stimulates appetite in a patient with AIDS (acquired immunodeficiency syndrome); chronic administration of doses increases weight and appetite. Δ9-THC is also given to patients with insomnia and depression as it improves sleep. In humans, Δ8-THC decreases intraocular pressure (fluid pressure inside the eye) and exhibits antiglaucoma activity [76]. Phytocannabinoid CBG possesses non-psychotic activity that has a lower affinity for both CB1R and CB2R, several times lower than Δ9-THC. CBG has remarkable activity towards ligand-gated cation channels of superfamily TRP. CBG serves as an agonist of TRP type ankyrin 1 (TRPA1) and TRP type vanilloid 1 (TRPV1), whereas it acts as an antagonistic of TRP type melastatin 8 (TRPM8). Δ9-tetrahydrocannabivarinic acid (Δ9-THCV) serves as a non-psychoactive that is antagonistic to CB1R. In particular, Δ9-THCV potentially acts against obesity-linked glucose intolerance. In humans, it acts as the potential phytocannabinoid for treating metabolic disorders, hepatosteatosis syndrome, and obesity. It also helps recover fasting plasma glucose and pancreatic β cell functioning. Additionally, Δ9-THCV reduces adiponectin and apolipoprotein [75].
CBD plays a potent pharmacological role in humans by affecting CNS and some peripheral portions, as it is highly antagonistic to CB1R and CB2R. Additionally, CBD also acts as an essential allosteric modulator of receptor μ-opioid (pain reliever), thus used to relieve pain [75]. In humans, doses in the range of 10 to 700 mg CBD are not toxic and are delivered to patients in the form of Epidiolex® with epilepsy treatment-resistant, interlinked with CDKL5 deficiency disorder and many other disorders and syndromes [80]. CBD does not act as a psychoactive compound and can be delivered in patients receiving pharmacotherapy. Furthermore, CBD reduces Δ9-THC-elicited psychotic disorders and decreases the deleterious effect of Δ9-THC on memory which is hippocampus-dependent. CBD acts as the potential cannabinoid to cure obesity, convulsive disorder, and rheumatoid arthritis. Furthermore, CBD also possesses antipsychotic, antinausea, and antianxiety properties [79].
CBC acts as the non-psychotropic which does not interact with CB1R and CB2R [5,78]. Additionally, CBC inhibits endocannabinoid inactivation and further activates TRPA1, which helps recover intestinal inflammation and has several protective roles [110,111]. Additionally, CBC also possesses anti-inflammatory activity for curing lipopolysaccharide-enhanced edema. In particular, till 2021, no CBC study involves humans [112].
6.2.2. Cannabinoid Acid
In C. sativa, CBDA, CBCA, CBDA, CBGA, and Δ9-THCA belong to the cannabinoid acid group, which does not possess cannabimimetic or psychotropic bioactivity [5]. Continuous decarboxylation of Δ9-THCA leads to a low level of Δ9-THC [113]. Δ9-THCA has more affinity than Δ9-THC and binds to PPARγ, whereas it has a low affinity towards CB1R and CB2R. Additionally, Δ9-THCA possesses the neuroprotective property [77]. Phytocannabinoids of cannabis and their derivatives hamper the level of tumor necrosis factor α after stimulating lipopolysaccharide in culture supernatants of some macrophages. Additionally, phytocannabinoids, i.e., cannabinoid acid, also display their effect on phospholipase activity specific to phosphatidylcholine; thus, this effects signaling. Cannabinoid acids, e.g., CBDA, possess antihyperalgesic (i.e., reduce abnormally enhance pain) and anti-inflammatory properties [114]. In mice, cannabinoid acid (i.e., CBDA, and Δ9-THCA) treatment increased antihyperalgesic and anti-inflammatory properties with inflammations induced by carrageenan. Additionally, in mice, oral administration of cannabinoids increases the antihyperalgesic property before carrageenan. In intestinal segments of house musk shrews, CBDA enhances tissue reduction in the resting state with the help of non-neuronal-associated pathways or pathways that are independent or are not associated with CB1R or CB2R [82,115].
6.2.3. Bibenzyl Cannabinoids
Analogs of bioactive bibenzyl (aralkyl) compound such as CBG from few liverworts have a phenethyl side chain; so bibenzyl CBG displays reduced affinity to cannabinoids receptors [116]. Additionally, it has an affinity to TRPA1 and TRP 1-4, which are ionotropic receptors, and shows a strong association to TRPM8 [10]. Bibenzyl phytocannabinoids belonging to the class amofrutin were extracted from Glycyrrhiza foetida and Amorpha fruticosa. They possess anti-inflammatory properties and are a strong activator of PPARγ [84]. Bibenzyl cannabinoid perrottetinene produced by Radula species shows a structural resemblance to cannabis Δ9-THC, distinguishing it by having an aromatic side chain instead of a pentyl. In mammals, perrottetinene acts as the psychoactive compound that is agonistic to CB1R. Furthermore, it enhances behavioral tetrads of analgesia, catalepsy, hypolocomotion, and hypothermia. Interestingly, enantiomers produced in liverworts are the major hallmark of their bioactivity and metabolic responses [28].
6.2.4. Rhododendron Cannabinoids
Rhododendron produces phytocannabinoids in the form of prenylated orcinoids. They are generally classified in CBC or CBL type as they have chromane or chromene scaffold. Prenylated orcinoids and their derivatives play a potent role in building and boosting up the immune system in mammals and humans. Rhododendron cannabinoids possess anticancer property; hence can be recommended to a cancer patient. It also has antithrombotic activity. Furthermore, it also displays anti-inflammatory, antimicrobial, and antipsychotic properties and has very low toxicity in humans when administered [5,37,83,117]. Other essential phytocannabinoids of the Rhododendron species, DCA and rhododaurichromenic acids, serve as the potent bioactive compounds in curing HIV as they possess anti-HIV bioactivities. It was found that when DCA was administered in infected H9 cells, it showed a half-maximal effective concentration (EC50) of 15 nM, much lower than the azidothymidine (a drug used to cure HIV), which has EC50 of 44 nM. Moreover, DCA with potent anti-HIV bioactivity has a therapeutic index (TI) of 3701 and EC50 of 5.67 ng mL−1 [81]. Moreover, rhododaurichromanic acid possesses anti-HIV properties with TI of 92 and EC50 of 370 ng mL−1. Active phytocannabinoids from Rh. anthopogonoides such as cannabiorcichromenic acid (CBC-type), anthopogocyclolic acid, and anthopogochromenic acid, which are CBL-type possess antiallergic activity as they inhibit the release of histamine [5,16,117].
7. Antibacterial and Antimicrobial Property of Phytocannabinoids
Cannabis fibers are used in textile industries as they are rich in active phytocannabinoids, which provide high biomass in less time. Hemp fibers, also called bast fibers, are rich in Δ9-THC, cellulose, lignin, and other metabolites, so they are used in industries, animal bedding, and used as a substitute for glass fibers. Hemp fibers also possess high antibacterial properties because of the presence of phytocannabinoids, so they are used as antibacterial finishing agents and in surgical industries. Recently it was found that cannabis powder contains a high amount of cannabinoid content which possesses antibacterial properties against Escherichia coli. Interestingly, phytocannabinoids have high pharmacological, antibacterial, and antimicrobial activities [65,78,105,118,119].
Cannabis extract contains antibacterial property against Bacillus subtilis and Staphylococcus aureus, which are Gram-positive bacteria; also against Gram-negative bacteria E. coli, Pseudomonas aeruginosa. In contrast, it displays no activity against fungal infections caused by Aspergillus niger and Candida albicans. Phytocannabinoids, e.g., Δ9-THC, CBG, CBN, CBD, and CBC, possess antibiotic properties against methicillin-resistant Staphylococcus aureus. CBD and Δ9-THC display bactericidal activity against streptococci and staphylococci. GFA and DCA also show antimicrobial properties against Gram-positive bacteria. Moreover, the prenyl group shows structural similarities to bioactive monoterpenes [58,65,85,108,120].
8. Phytocannabinoids in Stress Tolerance
Phytocannabinoids have a diverse role in humans and also exhibit antimicrobial, antibacterial, and antibiotic activity. Apart from this, they also have some other biological beneficial properties in mitigating biotic and abiotic stress in the plant. Cannabis trichomes possess phytocannabinoid in large quantities. High temperatures or herbivory cause trichome rupture and the release of the phytocannabinoid content, which protects the plant from desiccation and high-temperature stress. Therefore, they enhance plant tolerance to heat stress [2,79]. It was also reported that phytocannabinoid production was enhanced in cannabis flowers after UV-B-induced stress. Thus, phytocannabinoids serve as a sun shield against destructive UV-B radiation [26]. Liverworts oil bodies possess phytocannabinoid, so they are not damaged by fungi and bacteria, insect larvae and adults, slugs, snails, and small mammals. Significantly, phytocannabinoids inside oil bodies provide tolerance against several abiotic stresses such as cold temperature, heat, excessive light, and UV radiation [26,29,58].
Additionally, phytocannabinoids provide resilience to desiccation. Liverworts are unable to produce abscisic acid (ABA); besides this, they have phytocannabinoid, i.e., lunularic acid that shows the same activity as ABA, which is a plant hormone. There is a lack of knowledge about the role of phytocannabinoids in biotic and abiotic stress mitigation. Thus, more in-depth studies are still required to understand the mechanism involved in stress tolerance via phytocannabinoids. However, studies must understand their role in other plants’ stress amelioration [1].
9. Phytocannabinoid Foe
Why is cannabis cultivation illegal in many countries and what are the disadvantages associated with phytocannabinoid consumption? Cannabis is the third most popular substance consumed after alcohol and tobacco. The USA and New Zealand are at the top rates (42%) of consuming it; however, it is the most illicit drug used globally. Its consumption is escalating among teenagers, and they are consuming it in the form of hashish and marijuana [86,121,122]. It was reported that almost everyone who tried cannabis (marijuana or hashish) becomes a habitual abuser and dependent on cannabis; thus, this enhanced the cannabis use disorder [123,124]. Cannabis elicits a plethora of biological, metabolic, and physical responses. Nevertheless, everything depends on its utilization and what dose it is consumed in (what, when, and how). Most commonly used cannabis marijuana is prepared by drying leaves and flowering buds of the cannabis plant. Moreover, hashish which is eventually stronger than marijuana, is the concentrated resin from the female cannabis plant and is directly consumed via chewing and smoking. Kief is a trichomes mass obtained from flowers and leaves, which is used to make hashish. Additionally, hashish contains a mixture of several phytocannabinoids and terpenes [125]. Smoking or dabbing marijuana is the most common way in the consumption of cannabinoids. Phytocannabinoid delays the onset effect by half to two hour circulating level; circulating level is greater when they are smoked or intravenously consumed. In particular, the delay time in the onset of psychobiological effect is due to the biosynthesis of bioactive phytocannabinoid metabolite or induction in endocannabinoid availability or to reach receptors to show its effect. Cannabis consumption produces two kinds of effects: short-term (acute) and long-term (chronic) [86]. In humans, acute consumption of cannabis effect is classified into two groups: (i) physiological effect, which displays therapeutic potential, and (ii) recreation effects which show the effect on brain identical to psychotic state (psychotomimetic) [126,127]. Acute physiological effects include relaxation, hyperlocomotion, resting of heart, and increase in thirst, appetite, and food palatability. Acute psychotomimetic effects encompass enhanced sociability, euphoria (sense of great happiness), relaxation, hallucinations, delusions, conceptual disorganization, paranoia, social withdrawal, blurred vision, and decreased attention [128,129]. Furthermore, it also leads to anxiety, panic attacks, and dysphoria; all of these effects are similar to schizophrenia. Long-term (chronic) consumption of cannabis is related to addiction risk which depends on age, gender, lifestyle, frequency of intake, dosage, and genetic makeup [122]. Long-term cannabis consumption increases various alterations inside the human body (mainly heart, brain, and nervous system) and develops the serious risk of several disorders such as schizophrenia, neuropsychiatric syndrome, cerebellar infarction, vasoconstriction, and several fatal cardiovascular problems. Additionally, cannabis consumption has a detrimental effect on metabolic and biological processes; but these disorders are low if consumption of alcohol is low; apart from this, cannabis consumption confers useful effects on the risk of cardiometabolic factors such as insulin sensitivity, glycemia, lipid amount. Nevertheless, it remains to be demonstrated whether long-term cannabis consumption in humans functions as an activator or deactivator of CBRs [79,105,108,109,122,126].
10. Conclusions and Future Prospects
Phytocannabinoids are bioactive naturally occurring terpenoids that were earlier thought to be exclusive to C. sativa but have now also been produced in Rhododendron species, some legumes, the liverwort genus Radula, and some fungi. Bioactive phytocannabinoids show remarkable non-hallucinogenic biological properties determined by the variable nature of the side chain and prenyl group defined by the enzymes involved in their biosynthesis. The present review focused on genes and enzymes involved in biosynthesis across several plant species such as cannabis, Rhododendron and liverworts were discussed here. Moreover, these species are used in the combinatorial fashion to construct a required new bioactive phytocannabinoid structure established on integrating peculiar prenyl moieties, side chains, and unique cyclized core structures. Meanwhile, phytocannabinoids biosynthesis involves a large collection of enzymes that can potently develop bioactive phytocannabinoids via genetic engineering. This review gives a better understanding of the diverse role of phytocannabinoids in humans, plants, microbiology, and biotechnology (Figure 8). In particular, active phytocannabinoids play a crucial role in treating several diseases in humans. They possess antibacterial and antimicrobial properties in several industries. Recent studies also appraise phytocannabinoids role in cold, heat, and radiation stress tolerance. Phytocannabinoids also protect the plant from pathogens and herbivory. Moreover, the review also focuses on the foe of improper use of phytocannabinoids and why it is banned in many countries.
The hypothesis is that phytocannabinoids have versatile use and are beneficial for humans and plants if appropriately used. Further investigations are needed on genes and enzymes involved in the biosynthetic pathway in different plant species. Comparative study between type III PKS from cannabis and liverwort might explain their specificity to particular molecular structure and insight of genetic and protein engineering. Semi-synthetic techniques and chemical modification integration can be used for the biosynthesis of different types of cannabinoids—it also modulates their bioactivity and bioavailability. The availability of naturally occurring cannabinoids provides an insight into scrutinizing their role in plant stress tolerance, including individual and combinatorial exogenous effects. Lastly, quantitative chemical profiling can also give a deep knowledge of the occurrence and importance of NADES and their other possible role in plants and their role in reducing autotoxicity.
Author Contributions
Conceptualization, S.H.; methodology, S.H.; writing—original draft preparation, Y.A. and P.S.; writing—review and editing, S.H. and A.B.; visualization, A.B.; supervision, S.H. and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Yamshi Arif acknowledges the receipt of Inspire fellowship (IF 180778) by the Department of Science & Technology, Government of India, New Delhi.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| AAE1 | acyl-activated enzyme 1 |
| ABA | abscisic acid |
| ADME | absorption, distribution, metabolism, excretion |
| BBS | bibenzyl synthase |
| C4H | cinnamate 4-hydroxylase |
| 4CL | 4-coumarate:CoA ligase |
| CBR | cannabinoid receptor |
| CB1R | cannabinoid receptor type 1 |
| CB2R | cannabinoid receptor type 2 |
| CBC | cannabichromene |
| CBCA | cannabichromenic acid |
| CBCAS | cannabichromenic acid synthase |
| CBCV | cannabichromevarine |
| CBCVA | cannabichromevarinic acid |
| CBD | cannabidiol |
| CBDA | cannabidiolic acid |
| CBDAS | cannabidiolic acid synthase |
| CBDV | cannabidivarine |
| CBDVA | cannabidivarinic acid |
| CBE | cannabielsoin |
| CBG | cannabigerol |
| CBGA | cannabigerolic acid |
| CBGAS | cannabigerolic acid synthase |
| CBL | cannabicyclol |
| CBN | cannabinol |
| CBND | cannabidiol |
| CBT | cannabitriol |
| CHIL | chalcone isomerases-like protein |
| CNS | central nervous system |
| CYP | cytochrome P450 |
| DBR | double-bond reductase |
| DCA | daurichromenic acid |
| DCAS | daurichromenic acid synthase |
| DHAC | dihydrostilbene acid cyclase |
| Dicamba | 3,6-dichloro-2-methoxybenzoic acid |
| EC50 | effective concentration |
| GFA | grifolic acid |
| GPP | geranyl pyrophosphate |
| GPR | G-protein-coupled receptor |
| GPCR | G-protein-coupled cannabinoid receptor |
| IAA | indole-3-acetic acid |
| KR | ketoreductase |
| LOX | lipoxygenase |
| MEP | methylerythritol-4-phosphate |
| NAA | 1-naphthaleneacetic acid |
| NADES | natural deep eutectic solvent |
| OA | olivetolic acid |
| OAC | olivetolic acid cyclase |
| OLS | olivetol synthase |
| ORS | orcinol synthase |
| OSA | orsellinic acid |
| PA | perrottetinenic acid |
| PAL | phenylalanine ammonia-lyase |
| PAS | perrottetinenic acid synthase |
| PET | perrottetinene |
| PGR | plant growth regulator |
| PKS | polyketide synthase |
| PPARγ | peroxisome proliferator-activated receptor γ |
| PT | prenyltransferase |
| STS | stilbene synthase |
| TAL | tyrosine ammonia-lyase |
| TF | transcription factor |
| TI | therapeutic index |
| TRP | transient receptor potential |
| TRPA1 | TRP type ankyrin 1 |
| TRPM8 | TRP type melastanin 8 |
| TRPV1 | TRP type vanilloid 1 |
| Δ8-THC | Δ8-tetrahydrocannabinol |
| Δ9-THC | Δ9-tetrahydrocannabinol |
| Δ9-THCA | Δ9-tetrahydrocannabinolic acid |
| Δ9-THCAS | Δ9-tetrahydrocannabinolic acid synthase |
| Δ9-THCV | Δ9-tetrahydrocannabivarinic acid |
| Δ9-THCVA | Δ9-tetrahydrocannabidivarinic acid |
References
- Gülck, T.; Møller, B.L. Phytocannabinoids: Origins and biosynthesis. Trends Plant Sci. 2020, 25, 985–1004. [Google Scholar] [CrossRef] [PubMed]
- Hanuš, L.O.; Meyer, S.M.; Muñoz, E.; Taglialatela-Scafati, O.; Appendino, G. Phytocannabinoids: A unified critical inventory. Nat. Prod. Rep. 2016, 33, 1357–1392. [Google Scholar] [CrossRef] [Green Version]
- ElSohly, M.A.; Slade, D. Chemical constituents of marijuana: The complex mixture of natural cannabinoids. Life Sci. 2005, 78, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Berman, P.; Futoran, K.; Lewitus, G.M.; Mukha, D.; Benami, M.; Shlomi, T.; Meiri, D. A new ESI-LC/MS approach for comprehensive metabolic profiling of phytocannabinoids in Cannabis. Sci. Rep. 2018, 8, 14280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pollastro, F.; Caprioglio, D.; Del Prete, D.; Rogati, F.; Minassi, A.; Taglialatela-Scafati, O.; Munoz, E.; Appendino, G. Cannabichromene. Nat. Prod. Commun. 2018, 13, 1189–1194. [Google Scholar] [CrossRef] [Green Version]
- Happyana, N.; Kayser, O. Monitoring metabolite profiles of Cannabis sativa L. trichomes during flowering period using 1H NMR-based metabolomics and real-time PCR. Planta Med. 2016, 82, 1217–1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chanda, D.; Neumann, D.; Glatz, J.F.C. The endocannabinoid system: Overview of an emerging multi-faceted therapeutic target. Prostaglandins Leukot. Essent. Fatty Acids 2019, 140, 51–56. [Google Scholar] [CrossRef]
- Flores-Sanchez, I.J.; Verpoorte, R. Secondary metabolism in Cannabis. Phytochem. Rev. 2008, 7, 615–639. [Google Scholar] [CrossRef]
- Dei Cas, M.; Casagni, E.; Casiraghi, A.; Minghetti, P.; Fornasari, D.M.M.; Ferri, F.; Arnoldi, S.; Gambaro, V.; Roda, G. Phytocannabinoids profile in medicinal cannabis oils: The impact of plant varieties and preparation methods. Front. Pharmacol. 2020, 11, 570616. [Google Scholar] [CrossRef]
- Pollastro, F.; De Petrocellis, L.; Schiano-Moriello, A.; Chianese, G.; Heyman, H.; Appendino, G.; Taglialatela-Scafati, O. Amorfrutin-type phytocannabinoids from Helichrysum umbraculigerum. Fitoterapia 2017, 123, 13–17. [Google Scholar] [CrossRef] [Green Version]
- Taura, F.; Iijima, M.; Kurosaki, F. Daurichromenic acid and grifolic acid: Phytotoxic meroterpenoids that induce cell death in cell culture of their producer Rhododendron dauricum. Plant Signal. Behav. 2018, 13, e1422463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taura, F.; Iijima, M.; Lee, J.-B.; Hashimoto, T.; Asakawa, Y.; Kurosaki, F. Daurichromenic acid-producing oxidocyclase in the young leaves of Rhododendron dauricum. Nat. Prod. Commun. 2014, 9, 1329–1332. [Google Scholar] [CrossRef] [Green Version]
- Kashiwada, Y.; Yamazaki, K.; Ikeshiro, Y.; Yamagishi, T.; Fujioka, T.; Mihashi, K.; Mizuki, K.; Cosentino, L.M.; Fowke, K.; Morris-Natschke, S.L.; et al. Isolation of rhododaurichromanic acid B and the anti-HIV principles rhododaurichromanic acid A and rhododaurichromenic acid from Rhododendron dauricum. Tetrahedron 2001, 57, 1559–1563. [Google Scholar] [CrossRef]
- Rogachev, A.; Komarova, N.; Korchagina, D.; Dolgikh, M.; Sorokina, I.; Baev, D.; Tolstikova, T.; Fomenko, V.; Salakhutdinov, N. Some prenylated phenols of Rhododendron adamsii: Isolation, modification and pharmacological tests. Chem. Sustain. Dev. 2009, 17, 185–193. [Google Scholar]
- Yang, Y.-X.; Wang, J.-X.; Wang, Q.; Li, H.-L.; Tao, M.; Luo, Q.; Liu, H. New chromane and chromene meroterpenoids from flowers of Rhododendron rubiginosum Franch. var. rubiginosum. Fitoterapia 2018, 127, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Iwata, N.; Kitanaka, S. New cannabinoid-like chromane and chromene derivatives from Rhododendron anthopogonoides. Chem. Pharm. Bull. 2011, 59, 1409–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chicca, A.; Schafroth, M.A.; Reynoso-Moreno, I.; Erni, R.; Petrucci, V.; Carreira, E.M.; Gertsch, J. Uncovering the psychoactivity of a cannabinoid from liverworts associated with a legal high. Sci. Adv. 2018, 4, eaat2166. [Google Scholar] [CrossRef] [Green Version]
- Hussain, T.; Espley, R.V.; Gertsch, J.; Whare, T.; Stehle, F.; Kayser, O. Demystifying the liverwort Radula marginata, a critical review on its taxonomy, genetics, cannabinoid phytochemistry and pharmacology. Phytochem. Rev. 2019, 18, 953–965. [Google Scholar] [CrossRef]
- Métoyer, B.; Lebouvier, N.; Hnawia, E.; Herbette, G.; Thouvenot, L.; Asakawa, Y.; Nour, M.; Raharivelomanana, P. Chemotypes and biomarkers of seven species of new Caledonian liverworts from the Bazzanioideae subfamily. Molecules 2018, 23, 1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pryce, R.J. Lunularic acid, a common endogenous growth inhibitor of liverworts. Planta 1971, 97, 354–357. [Google Scholar] [CrossRef]
- Quaghebeur, K.; Coosemans, J.; Toppet, S.; Compernolle, F. Cannabiorci- and 8-chlorocannabiorcichromenic acid as fungal antagonists from Cylindrocarpon olidum. Phytochemistry 1994, 37, 159–161. [Google Scholar] [CrossRef]
- Elhendawy, M.A.; Wanas, A.S.; Radwan, M.M.; Azzaz, N.A.; Toson, E.S.; ElSohly, M.A. Chemical and biological studies of Cannabis sativa roots. Med. Cannabis Cannabinoids 2019, 1, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Happyana, N.; Agnolet, S.; Muntendam, R.; Van Dam, A.; Schneider, B.; Kayser, O. Analysis of cannabinoids in laser-microdissected trichomes of medicinal Cannabis sativa using LCMS and cryogenic NMR. Phytochemistry 2013, 87, 51–59. [Google Scholar] [CrossRef]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis glandular trichomes alter morphology and metabolite content during flower maturation. Plant J. 2019, 101, 37–56. [Google Scholar] [CrossRef]
- Mahlberg, P.G.; Kim, E.-S. Cuticle development on glandular trichomes of Cannabis sativa (Cannabaceae). Am. J. Bot. 1991, 78, 1113–1122. [Google Scholar] [CrossRef]
- Eichhorn Bilodeau, S.; Wu, B.-S.; Rufyikiri, A.-S.; MacPherson, S.; Lefsrud, M. An update on plant photobiology and implications for cannabis production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef]
- Iijima, M.; Munakata, R.; Takahashi, H.; Kenmoku, H.; Nakagawa, R.; Kodama, T.; Asakawa, Y.; Abe, I.; Yazaki, K.; Kurosaki, F.; et al. Identification and characterization of daurichromenic acid synthase active in anti-HIV biosynthesis. Plant Physiol. 2017, 174, 2213–2230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asakawa, Y.; Ludwiczuk, A. Chemical constituents of bryophytes: Structures and biological activity. J. Nat. Prod. 2017, 81, 641–660. [Google Scholar] [CrossRef]
- He, X.; Sun, Y.; Zhu, R.-L. The oil bodies of liverworts: Unique and important organelles in land plants. Crit. Rev. Plant Sci. 2013, 32, 293–302. [Google Scholar] [CrossRef]
- Geissler, M.; Volk, J.; Stehle, F.; Kayser, O.; Warzecha, H. Subcellular localization defines modification and production of Δ9-tetrahydrocannabinolic acid synthase in transiently transformed Nicotiana benthamiana. Biotechnol. Lett. 2018, 40, 981–987. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Reiter, M.A.; d’Espaux, L.; Wong, J.; Denby, C.M.; Lechner, A.; Zhang, Y.; Grzybowski, A.T.; Harth, S.; Lin, W.; et al. Complete biosynthesis of cannabinoids and their unnatural analogues in yeast. Nature 2019, 567, 123–126. [Google Scholar] [CrossRef]
- Rodziewicz, P.; Loroch, S.; Marczak, Ł.; Sickmann, A.; Kayser, O. Cannabinoid synthases and osmoprotective metabolites accumulate in the exudates of Cannabis sativa L. glandular trichomes. Plant Sci. 2019, 284, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Gagne, S.J.; Stout, J.M.; Liu, E.; Boubakir, Z.; Clark, S.M.; Page, J.E. Identification of olivetolic acid cyclase from Cannabis sativa reveals a unique catalytic route to plant polyketides. Proc. Natl. Acad. Sci. USA 2012, 109, 12811–12816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raharjo, T.J.; Chang, W.-T.; Choi, Y.H.; Peltenburg-Looman, A.M.G.; Verpoorte, R. Olivetol as product of a polyketide synthase in Cannabis sativa L. Plant Sci. 2004, 166, 381–385. [Google Scholar] [CrossRef]
- Laverty, K.U.; Stout, J.M.; Sullivan, M.J.; Shah, H.; Gill, N.; Holbrook, L.; Deikus, G.; Sebra, R.; Hughes, T.R.; Page, J.E.; et al. A physical and genetic map of Cannabis sativa identifies extensive rearrangements at the THC/CBD acid synthase loci. Genome Res. 2019, 29, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeki, H.; Hara, R.; Takahashi, H.; Iijima, M.; Munakata, R.; Kenmoku, H.; Fuku, K.; Sekihara, A.; Yasuno, Y.; Shinada, T.; et al. An aromatic farnesyltransferase functions in biosynthesis of the anti-HIV meroterpenoid daurichromenic acid. Plant Physiol. 2018, 178, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Taura, F.; Iijima, M.; Yamanaka, E.; Takahashi, H.; Kenmoku, H.; Saeki, H.; Morimoto, S.; Asakawa, Y.; Kurosaki, F.; Morita, H. A novel class of plant type III polyketide synthase involved in orsellinic acid biosynthesis from Rhododendron dauricum. Front. Plant Sci. 2016, 7, 1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsuyama, Y.; Kita, T.; Funa, N.; Horinouchi, S. Curcuminoid biosynthesis by two type III polyketide synthases in the herb Curcuma longa. J. Biol. Chem. 2009, 284, 11160–11170. [Google Scholar] [CrossRef] [Green Version]
- Hussain, T.; Plunkett, B.; Ejaz, M.; Espley, R.V.; Kayser, O. Identification of putative precursor genes for the biosynthesis of cannabinoid-like compound in Radula marginata. Front. Plant Sci. 2018, 9, 537. [Google Scholar] [CrossRef] [Green Version]
- Akiyama, T.; Shibuya, M.; Liu, H.-M.; Ebizuka, Y. p-Coumaroyltriacetic acid synthase, a new homologue of chalcone synthase, from Hydrangea macrophylla var. thunbergii. Eur. J. Biochem. 1999, 263, 834–839. [Google Scholar] [CrossRef]
- Austin, M.B.; Noel, J.P. The chalcone synthase superfamily of type III polyketide synthases. Nat. Prod. Rep. 2002, 20, 79–110. [Google Scholar] [CrossRef]
- Imoto, S.A.; Ohta, Y. Intracellular localization of lunularic acid and prelunularic acid in suspension cultured cells of Marchantia polymorpha. Plant Physiol. 1985, 79, 751–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toyota, M.; Shimamura, T.; Ishii, H.; Renner, M.; Braggins, J.; Asakawa, Y. New bibenzyl cannabinoid from the New Zealand liverwort Radula marginata. Chem. Pharm. Bull. 2002, 50, 1390–1392. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Matsui, T.; Kodama, T.; Mori, T.; Zhou, X.; Taura, F.; Noguchi, H.; Abe, I.; Morita, H. Structural basis for olivetolic acid formation by a polyketide cyclase from Cannabis sativa. FEBS J. 2016, 283, 1088–1106. [Google Scholar] [CrossRef] [Green Version]
- Toyota, M.; Kinugawa, T.; Asakawa, Y. Bibenzyl cannabinoid and bisbibenzyl derivative from the liverwort Radula perrottetii. Phytochemistry 1994, 37, 859–862. [Google Scholar] [CrossRef]
- Fuentes, P.; Armarego-Marriott, T.; Bock, R. Plastid transformation and its application in metabolic engineering. Curr. Opin. Biotechnol. 2018, 49, 10–15. [Google Scholar] [CrossRef]
- Kodym, A.; Leeb, C.J. Back to the roots: Protocol for the photoautotrophic micropropagation of medicinal Cannabis. Plant Cell Tissue Organ Cult. 2019, 138, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Lange, K.; Schmid, A.; Julsing, M.K. Δ9-Tetrahydrocannabinolic acid synthase: The application of a plant secondary metabolite enzyme in biocatalytic chemical synthesis. J. Biotechnol. 2016, 233, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Lata, H.; Chandra, S.; Khan, I.A.; ElSohly, M.A. Micropropagation of Cannabis sativa L. -An update. In Cannabis sativa L. Botany and Biotechnology; Chandra, S., Lata, H., ElSohly, M.A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 285–297. [Google Scholar] [CrossRef]
- Wróbel, T.; Dreger, M.; Wielgus, K.; Słomski, R. The application of plant in vitro cultures in cannabinoid production. Biotechnol. Lett. 2017, 40, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Andersen-Ranberg, J.; Kongstad, K.T.; Nielsen, M.T.; Jensen, N.B.; Pateraki, I.; Bach, S.S.; Hamberger, B.; Zerbe, P.; Staerk, D.; Bohlmann, J.; et al. Expanding the landscape of diterpene structural diversity through stereochemically controlled combinatorial biosynthesis. Angew. Chem. Int. Ed. 2016, 55, 2142–2146. [Google Scholar] [CrossRef]
- Chen, R.; Gao, B.; Liu, X.; Ruan, F.; Zhang, Y.; Lou, J.; Feng, K.; Wunsch, C.; Li, S.-M.; Dai, J.; et al. Molecular insights into the enzyme promiscuity of an aromatic prenyltransferase. Nat. Chem. Biol. 2017, 13, 226–234. [Google Scholar] [CrossRef]
- Kannangara, R.; Siukstaite, L.; Borch-Jensen, J.; Madsen, B.; Kongstad, K.T.; Staerk, D.; Bennedsen, M.; Okkels, F.T.; Rasmussen, S.A.; Larsen, T.O.; et al. Characterization of a membrane-bound C-glucosyltransferase responsible for carminic acid biosynthesis in Dactylopius coccus Costa. Nat. Commun. 2017, 8, 1987. [Google Scholar] [CrossRef] [Green Version]
- Yazaki, K.; Sasaki, K.; Tsurumaru, Y. Prenylation of aromatic compounds, a key diversification of plant secondary metabolites. Phytochemistry 2009, 70, 1739–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ban, Z.; Qin, H.; Mitchell, A.J.; Liu, B.; Zhang, F.; Weng, J.-K.; Dixon, R.A.; Wang, G. Noncatalytic chalcone isomerase-fold proteins in Humulus lupulus are auxiliary components in prenylated flavonoid biosynthesis. Proc. Natl. Acad. Sci. USA 2018, 115, E5223–E5232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gribble, G.W. Biological activity of recently discovered halogenated marine natural products. Mar. Drugs 2015, 13, 4044–4136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, E.H.; Osmani, S.A.; Kristensen, C.; Møller, B.L.; Hansen, J. Substrate specificities of family 1 UGTs gained by domain swapping. Phytochemistry 2009, 70, 473–482. [Google Scholar] [CrossRef]
- Khan, B.A.; Warner, P.; Wang, H. Antibacterial properties of hemp and other natural fibre plants: A review. BioResources 2014, 9, 3642–3659. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.H.; van Spronsen, J.; Dai, Y.; Verberne, M.; Hollmann, F.; Arends, I.W.C.E.; Witkamp, G.-J.; Verpoorte, R. Are natural deep eutectic solvents the missing link in understanding cellular metabolism and physiology? Plant Physiol. 2011, 156, 1701–1705. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Witkamp, G.-J.; Verpoorte, R.; Choi, Y.H. Tailoring properties of natural deep eutectic solvents with water to facilitate their applications. Food Chem. 2015, 187, 14–19. [Google Scholar] [CrossRef]
- Courchaine, E.M.; Lu, A.; Neugebauer, K.M. Droplet organelles? EMBO J. 2016, 35, 1603–1612. [Google Scholar] [CrossRef] [Green Version]
- Hammond, O.S.; Bowron, D.T.; Edler, K.J. The effect of water upon deep eutectic solvent nanostructure: An unusual transition from ionic mixture to aqueous solution. Angew. Chem. Int. Ed. 2017, 56, 9782–9785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knudsen, C.; Bavishi, K.; Viborg, K.M.; Drew, D.P.; Simonsen, H.T.; Motawia, M.S.; Møller, B.L.; Laursen, T. Stabilization of dhurrin biosynthetic enzymes from Sorghum bicolor using a natural deep eutectic solvent. Phytochemistry 2020, 170, 112214. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, C.; Gallage, N.J.; Hansen, C.C.; Møller, B.L.; Laursen, T. Dynamic metabolic solutions to the sessile life style of plants. Nat. Prod. Rep. 2018, 35, 1140–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The plant of the thousand and one molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lata, H.; Chandra, S.; Techen, N.; Khan, I.A.; ElSohly, M.A. Molecular analysis of genetic fidelity in Cannabis sativa L. plants grown from synthetic (encapsulated) seeds following in vitro storage. Biotechnol. Lett. 2011, 33, 2503–2508. [Google Scholar] [CrossRef] [PubMed]
- Gai, Q.-Y.; Jiao, J.; Luo, M.; Wei, Z.-F.; Zu, Y.-G.; Ma, W.; Fu, Y.-J. Establishment of hairy root cultures by Agrobacterium rhizogenes mediated transformation of Isatis tinctoria L. For the efficient production of flavonoids and evaluation of antioxidant activities. PLoS ONE 2015, 10, e0119022. [Google Scholar] [CrossRef]
- Feeney, M.; Punja, Z.K. Tissue culture and Agrobacterium-mediated transformation of hemp (Cannabis sativa L.). In Vitro Cell. Dev. Biol. Plant 2003, 39, 578–585. [Google Scholar] [CrossRef]
- Lata, H.; Chandra, S.; Khan, I.; ElSohly, M. High frequency plant regeneration from leaf derived callus of high Δ9-tetrahydrocannabinol yielding Cannabis sativa L. Planta Med. 2010, 76, 1629–1633. [Google Scholar] [CrossRef]
- Hazekamp, A.; Verpoorte, R. Structure elucidation of the tetrahydrocannabinol complex with randomly methylated β-cyclodextrin. Eur. J. Pharm. Sci. 2006, 29, 340–347. [Google Scholar] [CrossRef]
- Pellechia, T. Legal Cannabis Industry Poised for Big Growth, in North America and around the World. Forbes 2018. Available online: https://www.forbes.com/sites/thomaspellechia/2018/03/01/double-digit-billions-puts-north-america-in-the-worldwide-cannabis-market-lead/?sh=356dc9565109 (accessed on 15 April 2021).
- Booth, J.K.; Bohlmann, J. Terpenes in Cannabis sativa—From plant genome to humans. Plant Sci. 2019, 284, 67–72. [Google Scholar] [CrossRef]
- Laprairie, R.B.; Bagher, A.M.; Kelly, M.E.M.; Dupré, D.J.; Denovan-Wright, E.M. Type 1 cannabinoid receptor ligands display functional selectivity in a cell culture model of striatal medium spiny projection neurons. J. Biol. Chem. 2014, 289, 24845–24862. [Google Scholar] [CrossRef] [Green Version]
- Kendall, D.A.; Yudowski, G.A. Cannabinoid receptors in the central nervous system: Their signaling and roles in disease. Front. Cell. Neurosci. 2017, 10, 294. [Google Scholar] [CrossRef] [Green Version]
- Pryce, G.; Riddall, D.R.; Selwood, D.L.; Giovannoni, G.; Baker, D. Neuroprotection in experimental autoimmune encephalomyelitis and progressive multiple sclerosis by cannabis-based cannabinoids. J. Neuroimmune Pharmacol. 2014, 10, 281–292. [Google Scholar] [CrossRef]
- Hippalgaonkar, K.; Gul, W.; ElSohly, M.A.; Repka, M.A.; Majumdar, S. Enhanced solubility, stability, and transcorneal permeability of δ-8-tetrahydrocannabinol in the presence of cyclodextrins. AAPS PharmSciTech 2011, 12, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Nadal, X.; Del Río, C.; Casano, S.; Palomares, B.; Ferreiro-Vera, C.; Navarrete, C.; Sánchez-Carnerero, C.; Cantarero, I.; Bellido, M.L.; Meyer, S.; et al. Tetrahydrocannabinolic acid is a potent PPARγ agonist with neuroprotective activity. Br. J. Pharmacol. 2017, 174, 4263–4276. [Google Scholar] [CrossRef] [Green Version]
- Turner, S.E.; Williams, C.M.; Iversen, L.; Whalley, B.J. Molecular pharmacology of phytocannabinoids. In Phytocannabinoids: Unraveling the Complex Chemistry and Pharmacology of Cannabis sativa; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J.I., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 61–101. [Google Scholar] [CrossRef]
- Huestis, M.A.; Solimini, R.; Pichini, S.; Pacifici, R.; Carlier, J.; Busardò, F.P. Cannabidiol adverse effects and yoxicity. Curr. Neuropharmacol. 2019, 17, 974–989. [Google Scholar] [CrossRef]
- Devinsky, O.; Verducci, C.; Thiele, E.A.; Laux, L.C.; Patel, A.D.; Filloux, F.; Szaflarski, J.P.; Wilfong, A.; Clark, G.D.; Park, Y.D.; et al. Open-label use of highly purified CBD (Epidiolex®) in patients with CDKL5 deficiency disorder and Aicardi, Dup15q, and Doose syndromes. Epilepsy Behav. 2018, 86, 131–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rok Lee, Y.; Wang, X. A short synthetic route to biologically active (±)-daurichromenic acid as highly potent anti-HIV agent. Org. Biomol. Chem. 2005, 3, 3955. [Google Scholar] [CrossRef] [PubMed]
- Verhoeckx, K.C.M.; Korthout, H.A.A.J.; van Meeteren-Kreikamp, A.P.; Ehlert, K.A.; Wang, M.; van der Greef, J.; Rodenburg, R.J.T.; Witkamp, R.F. Unheated Cannabis sativa extracts and its major compound THC-acid have potential immuno-modulating properties not mediated by CB1 and CB2 receptor coupled pathways. Int. Immunopharmacol. 2006, 6, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Costa, M.; Dias, T.A.; Brito, A.; Proença, F. Biological importance of structurally diversified chromenes. Eur. J. Med. Chem. 2016, 123, 487–507. [Google Scholar] [CrossRef]
- Weidner, C.; Wowro, S.J.; Freiwald, A.; Kawamoto, K.; Witzke, A.; Kliem, M.; Siems, K.; Müller-Kuhrt, L.; Schroeder, F.C.; Sauer, S. Amorfrutin B is an efficient natural peroxisome proliferator-activated receptor gamma (PPARγ) agonist with potent glucose-lowering properties. Diabetologia 2013, 56, 1802–1812. [Google Scholar] [CrossRef]
- Ali, E.M.M.; Almagboul, A.Z.I.; Khogali, S.M.E.; Gergeir, U.M.A. Antimicrobial activity of Cannabis sativa L. Chin. Med. 2012, 3, 61–64. [Google Scholar] [CrossRef] [Green Version]
- Solymosi, K.; Kofalvi, A. Cannabis: A treasure trove or Pandora’s Box? Mini Rev. Med. Chem. 2017, 17, 1223–1291. [Google Scholar] [CrossRef] [PubMed]
- Morales, P.; Hurst, D.P.; Reggio, P.H. Molecular targets of the phytocannabinoids: A complex picture. In Phytocannabinoids: Unraveling the Complex Chemistry and Pharmacology of Cannabis sativa; Kinghorn, A.D., Falk, H., Gibbons, S., Kobayashi, J.i., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 103–131. [Google Scholar] [CrossRef] [Green Version]
- Lenkey, N.; Kirizs, T.; Holderith, N.; Máté, Z.; Szabó, G.; Vizi, E.S.; Hájos, N.; Nusser, Z. Tonic endocannabinoid-mediated modulation of GABA release is independent of the CB1 content of axon terminals. Nat. Commun. 2015, 6, 6557. [Google Scholar] [CrossRef] [PubMed]
- Herkenham, M.; Lynn, A.B.; Johnson, M.R.; Melvin, L.S.; de Costa, B.R.; Rice, K.C. Characterization and localization of cannabinoid receptors in rat brain: A quantitative in vitro autoradiographic study. J. Neurosci. 1991, 11, 563–583. [Google Scholar] [CrossRef] [PubMed]
- Fezza, F.; Bari, M.; Florio, R.; Talamonti, E.; Feole, M.; Maccarrone, M. Endocannabinoids, related compounds and their metabolic routes. Molecules 2014, 19, 17078–17106. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, P.; Baggelaar, M.; Erdelyi, K.; Cao, Z.; Cinar, R.; Fezza, F.; Ignatowska-Janlowska, B.; Wilkerson, J.; van Gils, N.; Hansen, T.; et al. The novel, orally available and peripherally restricted selective cannabinoid CB2 receptor agonist LEI-101 prevents cisplatin-induced nephrotoxicity. Br. J. Pharmacol. 2016, 173, 446–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, S.P.H. So what do we call GPR18 now? Br. J. Pharmacol. 2012, 165, 2411–2413. [Google Scholar] [CrossRef]
- Caldwell, M.D.; Hu, S.S.-J.; Viswanathan, S.; Bradshaw, H.; Kelly, M.E.M.; Straiker, A. A GPR18-based signalling system regulates IOP in murine eye. Br. J. Pharmacol. 2013, 169, 834–843. [Google Scholar] [CrossRef] [Green Version]
- McHugh, D.; Page, J.; Dunn, E.; Bradshaw, H.B. Δ9-Tetrahydrocannabinol and N-arachidonyl glycine are full agonists at GPR18 receptors and induce migration in human endometrial HEC-1B cells. Br. J. Pharmacol. 2012, 165, 2414–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Console-Bram, L.; Brailoiu, E.; Brailoiu, G.C.; Sharir, H.; Abood, M.E. Activation of GPR18 by cannabinoid compounds: A tale of biased agonism. Br. J. Pharmacol. 2014, 171, 3908–3917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, V.B.; Puhl, H.L., 3rd; Ikeda, S.R. N-Arachidonyl glycine does not activate G protein-coupled receptor 18 signaling via canonical pathways. Mol. Pharmacol. 2013, 83, 267–282. [Google Scholar] [CrossRef] [Green Version]
- Offertáler, L.; Mo, F.-M.; Bátkai, S.; Liu, J.; Begg, M.; Razdan, R.K.; Martin, B.R.; Bukoski, R.D.; Kunos, G. Selective ligands and cellular effectors of a G protein-coupled endothelial cannabinoid receptor. Mol. Pharmacol. 2003, 63, 699–705. [Google Scholar] [CrossRef] [Green Version]
- Baker, D.; Pryce, G.; Davies, W.L.; Hiley, C.R. In silico patent searching reveals a new cannabinoid receptor. Trends Pharmacol. Sci. 2006, 27, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Sawzdargo, M.; Nguyen, T.; Lee, D.K.; Lynch, K.R.; Cheng, R.; Heng, H.H.Q.; George, S.R.; O’Dowd, B.F. Identification and cloning of three novel human G protein-coupled receptor genes GPR52, ΨGPR53 and GPR55: GPR55 is extensively expressed in human brain. Mol. Brain Res. 1999, 64, 193–198. [Google Scholar] [CrossRef]
- Shore, D.M.; Reggio, P.H. The therapeutic potential of orphan GPCRs, GPR35 and GPR55. Front. Pharmacol. 2015, 6, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilius, B.; Szallasi, A. Transient receptor potential channels as drug targets: From the science of basic research to the art of medicine. Pharmacol. Rev. 2014, 66, 676–814. [Google Scholar] [CrossRef]
- Köfalvi, A. Alternative interacting sites and novel receptors for cannabinoid ligands. In Cannabinoids and the Brain; Köfalvi, A., Ed.; Springer: Boston, MA, USA, 2008; pp. 131–160. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.H.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International union of basic and clinical pharmacology. LXXIX. Cannabinoid receptors and their ligands: Beyond CB1 and CB2. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storozhuk, M.V.; Zholos, A.V. TRP channels as novel targets for endogenous ligands: Focus on endocannabinoids and nociceptive signalling. Curr. Neuropharmacol. 2018, 16, 137–150. [Google Scholar] [CrossRef]
- Whiting, P.F.; Wolff, R.F.; Deshpande, S.; Di Nisio, M.; Duffy, S.; Hernandez, A.V.; Keurentjes, J.C.; Lang, S.; Misso, K.; Ryder, S.; et al. Cannabinoids for medical use. JAMA 2015, 313, 2456. [Google Scholar] [CrossRef]
- Owens, B. Drug development: The treasure chest. Nature 2015, 525, S6–S8. [Google Scholar] [CrossRef] [Green Version]
- Tzadok, M.; Uliel-Siboni, S.; Linder, I.; Kramer, U.; Epstein, O.; Menascu, S.; Nissenkorn, A.; Yosef, O.B.; Hyman, E.; Granot, D.; et al. CBD-enriched medical cannabis for intractable pediatric epilepsy. Seizure 2016, 35, 41–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, S.P.H. Therapeutic potential of cannabis-related drugs. Prog. Neuropsychopharmacol. Biol. Psychiatry 2016, 64, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, M. Medical marijuana: Showdown at the cannabis corral. Nature 2015, 525, S15–S17. [Google Scholar] [CrossRef] [PubMed]
- Izzo, A.A.; Capasso, R.; Aviello, G.; Borrelli, F.; Romano, B.; Piscitelli, F.; Gallo, L.; Capasso, F.; Orlando, P.; Di Marzo, V. Inhibitory effect of cannabichromene, a major non-psychotropic cannabinoid extracted from Cannabis sativa, on inflammation-induced hypermotility in mice. Br. J. Pharmacol. 2012, 166, 1444–1460. [Google Scholar] [CrossRef] [Green Version]
- Jadoon, K.A.; Ratcliffe, S.H.; Barrett, D.A.; Thomas, E.L.; Stott, C.; Bell, J.D.; O’Sullivan, S.E.; Tan, G.D. Efficacy and safety of cannabidiol and tetrahydrocannabivarin on glycemic and lipid parameters in patients with type 2 diabetes: A randomized, double-blind, placebo-controlled, parallel group pilot study. Diabetes Care 2016, 39, 1777–1786. [Google Scholar] [CrossRef] [Green Version]
- Valliere, M.A.; Korman, T.P.; Woodall, N.B.; Khitrov, G.A.; Taylor, R.E.; Baker, D.; Bowie, J.U. A cell-free platform for the prenylation of natural products and application to cannabinoid production. Nat. Commun. 2019, 10, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno-Sanz, G. Can you pass the acid test? Critical review and novel therapeutic perspectives of Δ9-tetrahydrocannabinolic acid A. Cannabis Cannabinoid Res. 2016, 1, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Rock, E.M.; Limebeer, C.L.; Parker, L.A. Effect of cannabidiolic acid and ∆9-tetrahydrocannabinol on carrageenan-induced hyperalgesia and edema in a rodent model of inflammatory pain. Psychopharmacology 2018, 235, 3259–3271. [Google Scholar] [CrossRef]
- McPartland, J.M.; MacDonald, C.; Young, M.; Grant, P.S.; Furkert, D.P.; Glass, M. Affinity and efficacy studies of tetrahydrocannabinolic acid A at cannabinoid receptor types one and two. Cannabis Cannabinoid Res. 2017, 2, 87–95. [Google Scholar] [CrossRef]
- Fuhr, L.; Rousseau, M.; Plauth, A.; Schroeder, F.C.; Sauer, S. Amorfrutins are natural PPARγ agonists with potent anti-inflammatory properties. J. Nat. Prod. 2015, 78, 1160–1164. [Google Scholar] [CrossRef]
- Iwata, N.; Kitanaka, S. Tetracyclic chromane derivatives from Rhododendron anthopogonoides. J. Nat. Prod. 2010, 73, 1203–1206. [Google Scholar] [CrossRef]
- Khan, B.A.; Wang, J.; Warner, P.; Wang, H. Antibacterial properties of hemp hurd powder against E. coli. J. Appl. Polym. Sci. 2014, 132, 41588. [Google Scholar] [CrossRef]
- Wahby, I.; Caba, J.M.; Ligero, F. Agrobacteriuminfection of hemp (Cannabis sativa L.): Establishment of hairy root cultures. J. Plant Interact. 2013, 8, 312–320. [Google Scholar] [CrossRef]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef]
- Degenhardt, L.; Chiu, W.-T.; Sampson, N.; Kessler, R.C.; Anthony, J.C.; Angermeyer, M.; Bruffaerts, R.; de Girolamo, G.; Gureje, O.; Huang, Y.; et al. Toward a global view of alcohol, tobacco, cannabis, and cocaine use: Findings from the WHO World Mental Health Surveys. PLoS Med. 2008, 5, e141. [Google Scholar] [CrossRef] [Green Version]
- Volkow, N.D.; Baler, R.D.; Compton, W.M.; Weiss, S.R.B. Adverse health effects of marijuana use. N. Engl. J. Med. 2014, 370, 2219–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrawal, A.; Lynskey, M.T.; Bucholz, K.K.; Kapoor, M.; Almasy, L.; Dick, D.M.; Edenberg, H.J.; Foroud, T.; Goate, A.; Hancock, D.B.; et al. DSM-5 cannabis use disorder: A phenotypic and genomic perspective. Drug Alcohol Depend. 2014, 134, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasin, D.S.; O’Brien, C.P.; Auriacombe, M.; Borges, G.; Bucholz, K.; Budney, A.; Compton, W.M.; Crowley, T.; Ling, W.; Petry, N.M.; et al. DSM-5 criteria for substance use disorders: Recommendations and rationale. Am. J. Psychiatry 2013, 170, 834–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierre, J.M.; Gandal, M.; Son, M. Cannabis-induced psychosis associated with high potency “wax dabs”. Schizophr. Res. 2016, 172, 211–212. [Google Scholar] [CrossRef] [PubMed]
- Ashton, C.H. Pharmacology and effects of cannabis: A brief review. Br. J. Psychiatry 2001, 178, 101–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodondi, N.; Pletcher, M.J.; Liu, K.; Hulley, S.B.; Sidney, S. Marijuana use, diet, body mass index, and cardiovascular risk factors (from the CARDIA Study). Am. J. Cardiol. 2006, 98, 478–484. [Google Scholar] [CrossRef]
- Moosmann, B.; Roth, N.; Auwärter, V. Finding cannabinoids in hair does not prove cannabis consumption. Sci. Rep. 2015, 5, 14906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sewell, R.A.; Schnakenberg, A.; Elander, J.; Radhakrishnan, R.; Williams, A.; Skosnik, P.D.; Pittman, B.; Ranganathan, M.; D’Souza, D.C. Acute effects of THC on time perception in frequent and infrequent cannabis users. Psychopharmacology 2013, 226, 401–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Structure of phytocannabinoids in Cannabis sativa. Abbreviations: CBC, cannabichromene; CBCA, cannabichromenic acid; CBCV, cannabichromevarine; CBCVA, cannabichromevarinic acid; CBD, cannabidiol; CBDA, cannabidiolic acid; CBDV, cannabidivarine, CBE, cannabielsoin; CBG, cannabigerol; CBL, cannabicyclol; Δ8-THC, Δ8-tetrahydrocannabinol; Δ9-THC, Δ9-tetrahydrocannabinol; Δ9-THCA, Δ9-tetrahydrocannabinolic acid; Δ9-THCV, Δ9-tetrahydrocannabivarinic acid.
Figure 1.
Structure of phytocannabinoids in Cannabis sativa. Abbreviations: CBC, cannabichromene; CBCA, cannabichromenic acid; CBCV, cannabichromevarine; CBCVA, cannabichromevarinic acid; CBD, cannabidiol; CBDA, cannabidiolic acid; CBDV, cannabidivarine, CBE, cannabielsoin; CBG, cannabigerol; CBL, cannabicyclol; Δ8-THC, Δ8-tetrahydrocannabinol; Δ9-THC, Δ9-tetrahydrocannabinol; Δ9-THCA, Δ9-tetrahydrocannabinolic acid; Δ9-THCV, Δ9-tetrahydrocannabivarinic acid.

Figure 2.
Structure of phytocannabinoids in Helichrysum and Glycyrrhiza plants.

Figure 3.
Structure of phytocannabinoids in Rhododendron plants.

Figure 4.
Structure of phytocannabinoids in (a) liverworts and (b) fungi.

Figure 5.
Phytocannabinoids biosynthesis in Cannabis sativa. Abbreviations: CBCA, cannabichromenic acid; CBCAS, cannabichromenic acid synthase; CBDA, cannabidiolic acid; CBDAS, cannabidiolic acid synthase; Δ9-THCA, Δ9-tetrahydrocannabinolic acid; Δ9-THCAS, Δ9-tetrahydrocannabinolic acid synthase.
Figure 5.
Phytocannabinoids biosynthesis in Cannabis sativa. Abbreviations: CBCA, cannabichromenic acid; CBCAS, cannabichromenic acid synthase; CBDA, cannabidiolic acid; CBDAS, cannabidiolic acid synthase; Δ9-THCA, Δ9-tetrahydrocannabinolic acid; Δ9-THCAS, Δ9-tetrahydrocannabinolic acid synthase.

Figure 6.
Phytocannabinoids biosynthesis in Rhododendron. Abbreviations: DCAS, daurichromenic acid synthase; MEP, methylerythritol-4-phosphate; PT, prenyltransferase.
Figure 6.
Phytocannabinoids biosynthesis in Rhododendron. Abbreviations: DCAS, daurichromenic acid synthase; MEP, methylerythritol-4-phosphate; PT, prenyltransferase.

Figure 7.
Phytocannabinoids biosynthesis in liverworts. Abbreviations: C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; PAL, phenylalanine ammonia-lyase; TAL, tyrosine ammonia-lyase.
Figure 7.
Phytocannabinoids biosynthesis in liverworts. Abbreviations: C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate:CoA ligase; PAL, phenylalanine ammonia-lyase; TAL, tyrosine ammonia-lyase.

Figure 8.
Selective role of active phytocannabinoids in humans, mammals, plants, biotechnology, and industries. Abbreviations: CBC, cannabichromene; CBCA, cannabichromenic acid; CBD, cannabidiol; CBDA, cannabidiolic acid; CBG, cannabigerol; CBGA, cannabigerolic acid; CBN, cannabinol; CBR, cannabinoid receptor; DCA, daurichromenic acid; PA, perrottetinenic acid; Δ9-THC, Δ9-tetrahydrocannabinol.
Figure 8.
Selective role of active phytocannabinoids in humans, mammals, plants, biotechnology, and industries. Abbreviations: CBC, cannabichromene; CBCA, cannabichromenic acid; CBD, cannabidiol; CBDA, cannabidiolic acid; CBG, cannabigerol; CBGA, cannabigerolic acid; CBN, cannabinol; CBR, cannabinoid receptor; DCA, daurichromenic acid; PA, perrottetinenic acid; Δ9-THC, Δ9-tetrahydrocannabinol.


{kind=link}
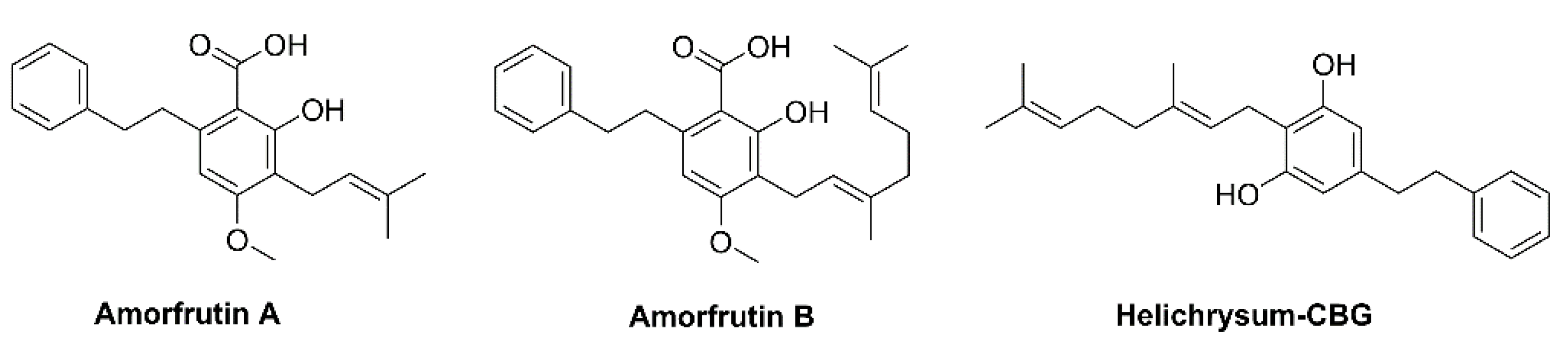{kind=link}
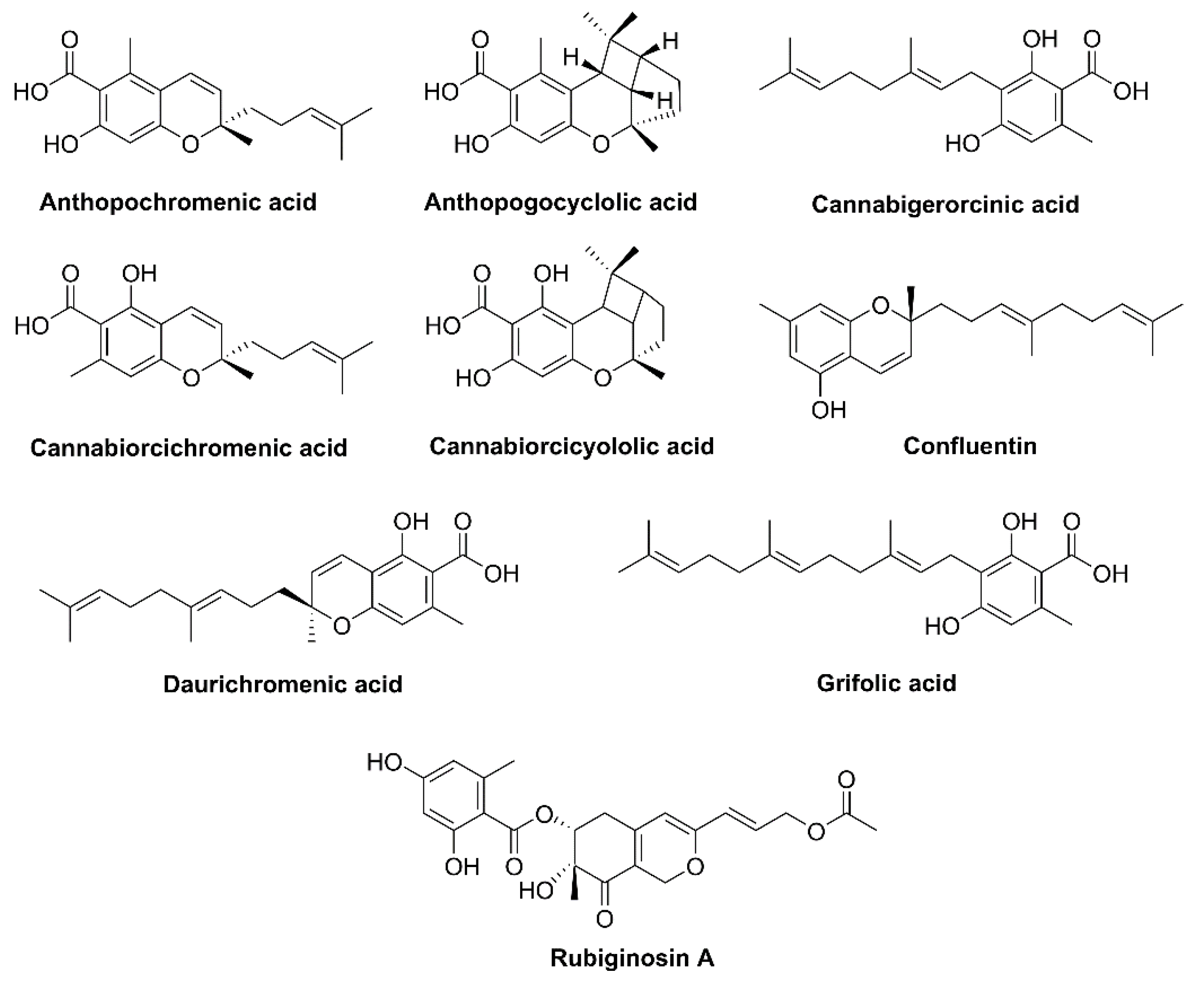{kind=link}
{kind=link}
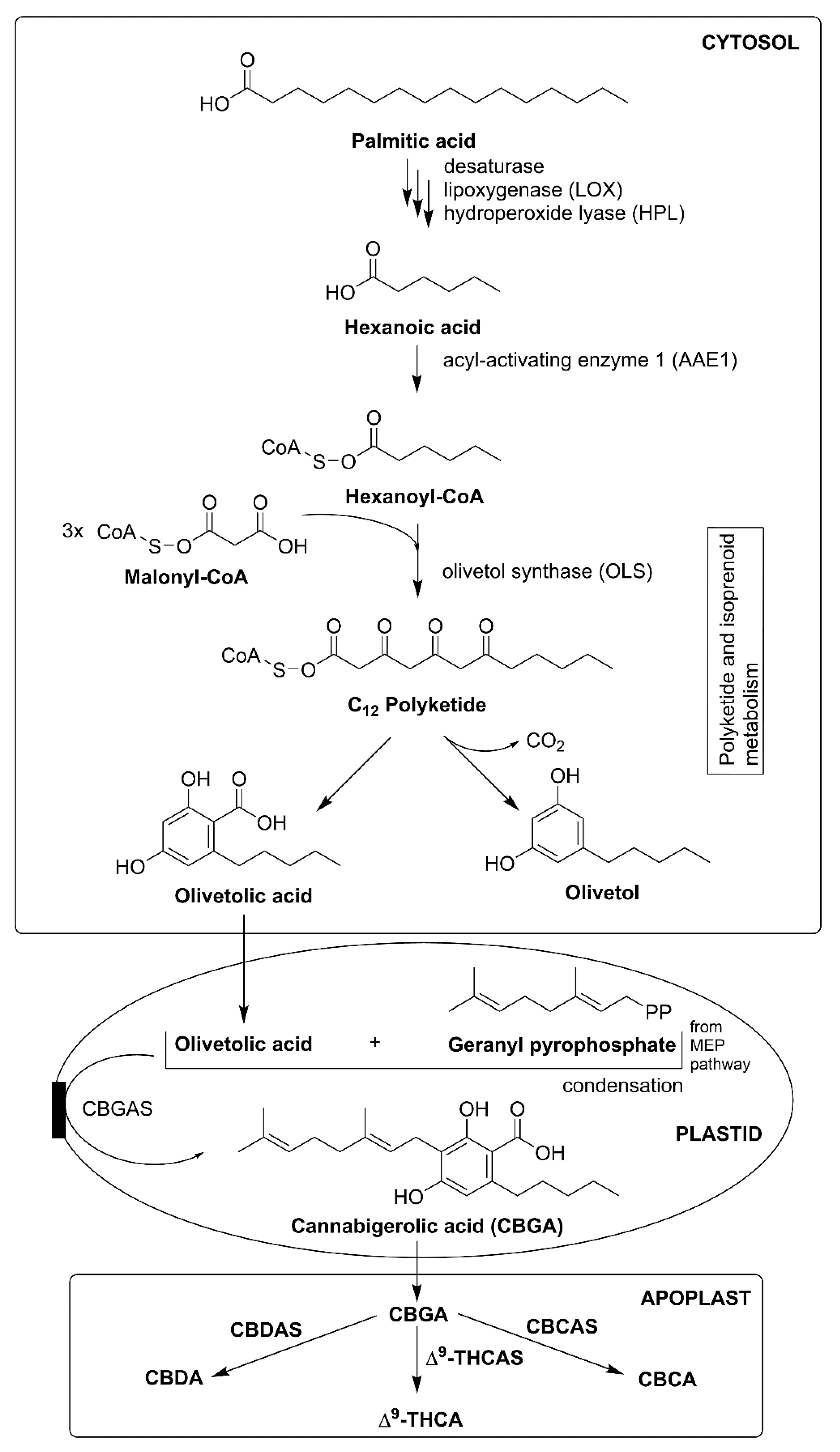{kind=link}
{kind=link}
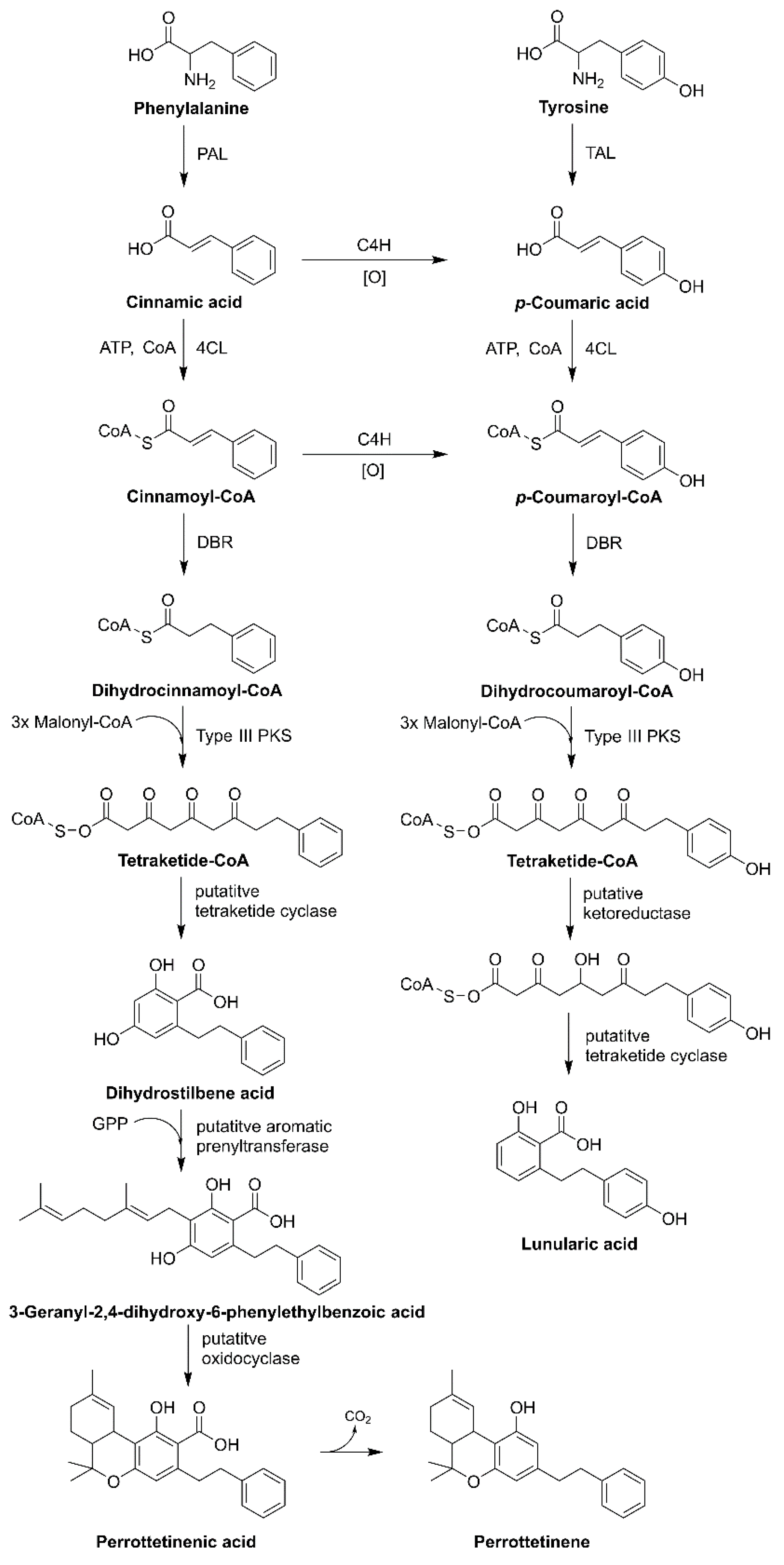{kind=link}
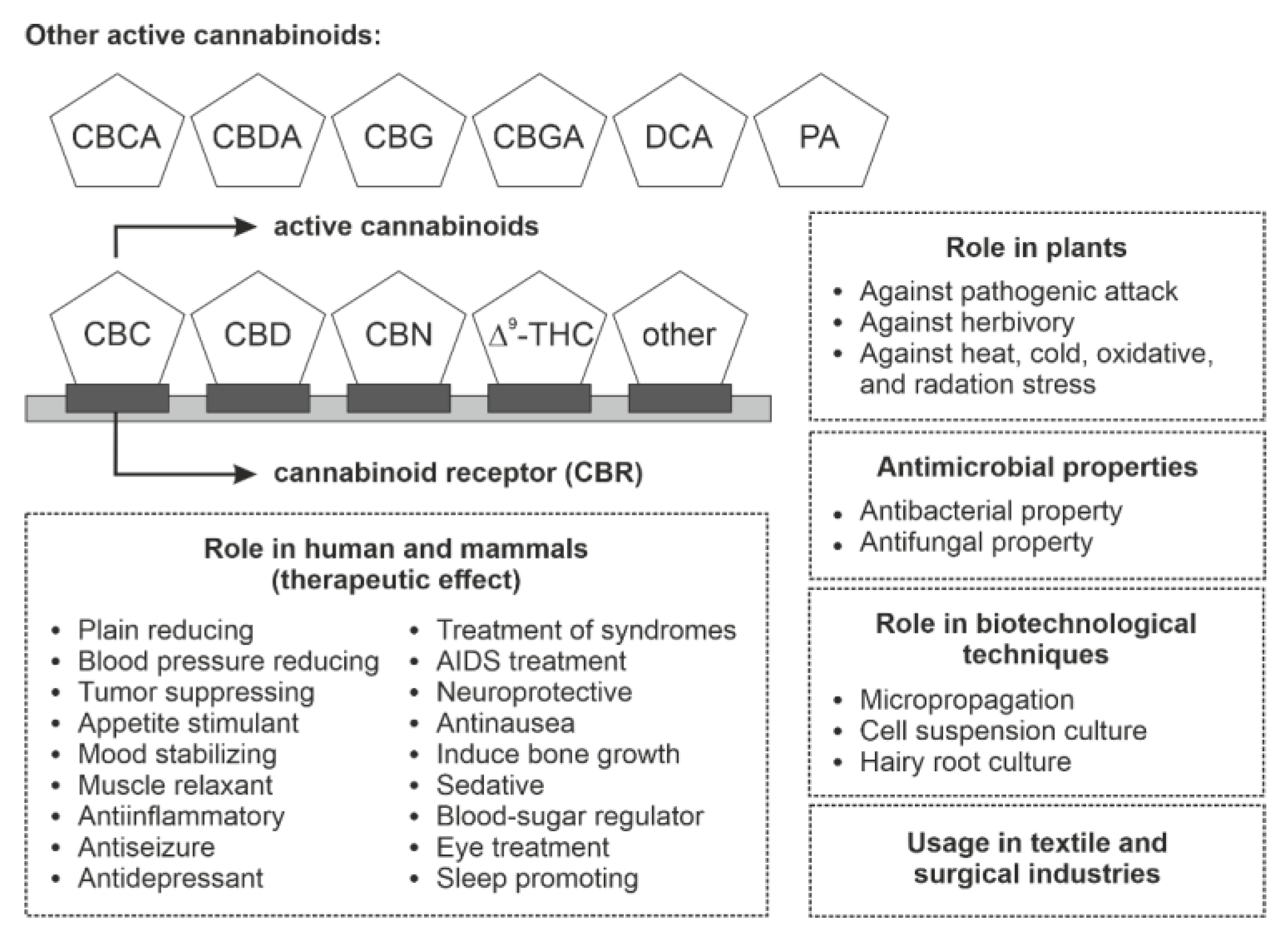{kind=link}
Table 1.
Bioactivity and effect of different phytocannabinoid in animals and humans.
| Phytocannabinoids | Bioactivity in Animals and Human | References |
|---|---|---|
| Δ9-THC | pleiotropic effects such as analgesic, muscle relaxation, and pain tolerance | [1] |
| increases weight and appetite; improves sleep and depression; alters mood, behavior, feeling, and thoughts | [72,73] | |
| cures neuropathic pain, spasticity, dysphoria, and anxiety disorders | [74,75] | |
| antiemetic activity, i.e., prevents vomiting in cancer patient during chemotherapy | [76] | |
| displays antiglaucoma activity and reduces intraocular pressure | [76] | |
| Δ9-THCA | neuroprotective and antitumor activity | [77] |
| Δ9-THCV | non-psychoactive effect; cures obesity | [75] |
| effective against metabolic disorders, pancreatic disease, and hepatosteatosis syndrome | [1] | |
| CBC | non-psychotropic and anti-inflammatory activity | [77,78] |
| CBD | cures memory loss, obesity, convulsive disorder, and rheumatoid arthritis | [79] |
| cures epilepsy; exhibits antipsychotic, antinausea, and antianxiety activity | [80] | |
| CBDA | anti-inflammatory and antihyperalgesia effect | [81,82] |
| CBG | non-psychotic activity | [75] |
| CBC, CBL, and DCA | cures HIV and cancer; possess immune boosting activity | [83] |
| CBG and amofrutin | anti-inflammatory activity | [84] |
| CBL | anti-inflammatory, anti-microbial, antipsychotic, and antiallergic activity | [5] |
| GFA and DCA | anti-microbial activity | [85] |
| PET | increases analgesia, catalepsy, hypolocomotion and hypothermia | [28] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arif, Y.; Singh, P.; Bajguz, A.; Hayat, S. Phytocannabinoids Biosynthesis in Angiosperms, Fungi, and Liverworts and Their Versatile Role. Plants 2021, 10, 1307. https://doi.org/10.3390/plants10071307
AMA Style
Arif Y, Singh P, Bajguz A, Hayat S. Phytocannabinoids Biosynthesis in Angiosperms, Fungi, and Liverworts and Their Versatile Role. Plants. 2021; 10(7):1307. https://doi.org/10.3390/plants10071307
Chicago/Turabian StyleArif, Yamshi, Priyanka Singh, Andrzej Bajguz, and Shamsul Hayat. 2021. "Phytocannabinoids Biosynthesis in Angiosperms, Fungi, and Liverworts and Their Versatile Role" Plants 10, no. 7: 1307. https://doi.org/10.3390/plants10071307
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.