
OsKEAP1 Interacts with OsABI5 and Its Downregulation Increases the Transcription of OsABI5 and the ABA Response Genes in Germinating Rice Seeds


Abstract
:1. Introduction
2. Results
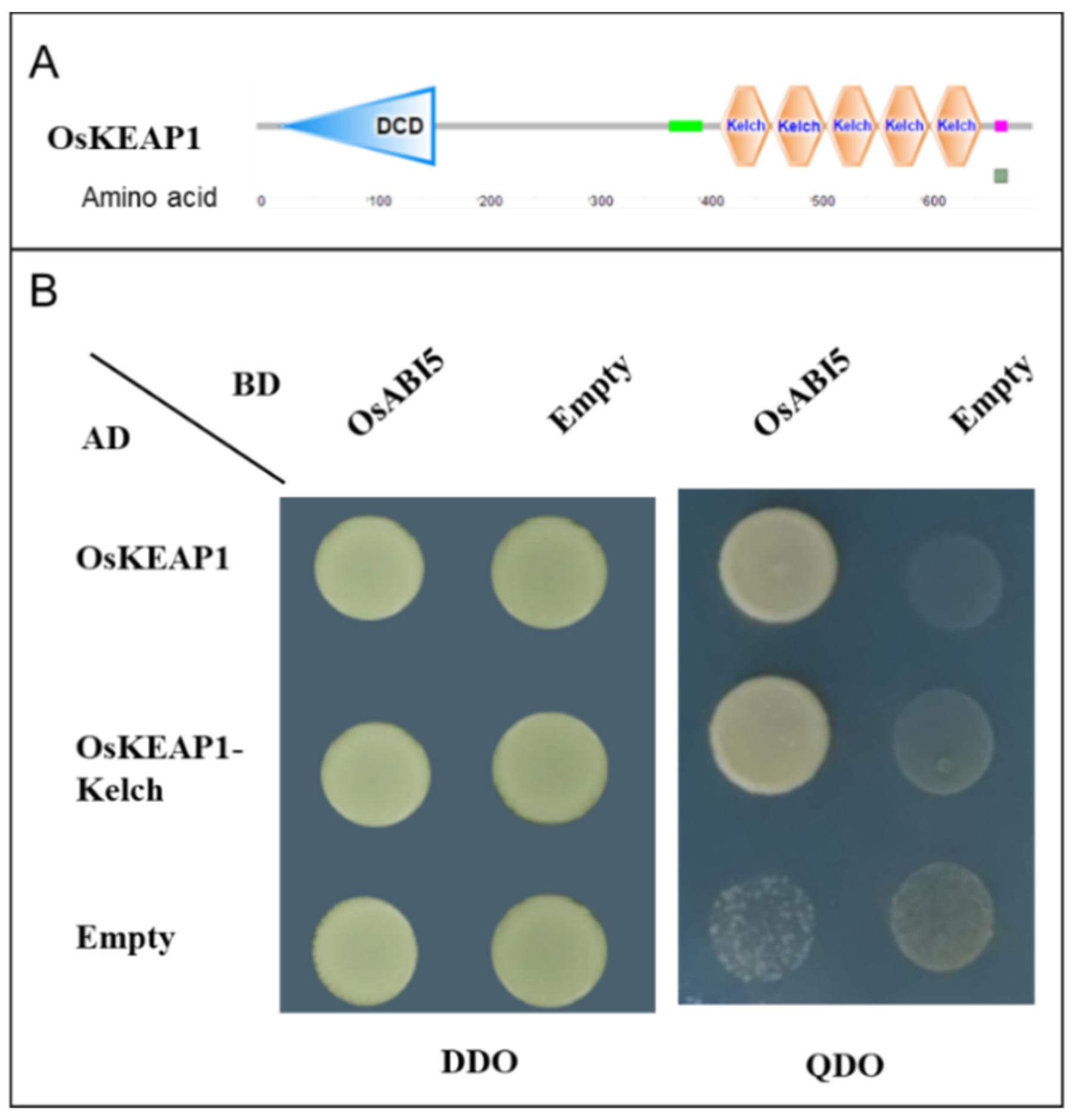
2.1. OsKEAP1 Interacts with OsABI5 in the Nucleus
2.2. Downregulation of OsKEAP1 Enhanced the Transcription of OsABI5
2.3. Downregulation of OsKEAP1 Had Little Effect on the Expression of Genes Regulating OsABI5
2.4. Downregulation of OsKEAP1 Altered the Transcription of ABA Response Genes
3. Discussion
3.1. The Likely OsKEAP1–OsABI5 Working Mechanism
3.2. The Effect of OsKEAP1 Downregulation on OsABI5 Expression and Its Consequences
3.3. Further Studies
4. Materials and Methods
4.1. Search of NRF2 Orthologs
4.2. Plant Materials
4.3. Yeast Two-Hybrids Assay
4.4. Subcellular Localization and BiFC Assay
4.5. Real-Time Quantitative PCR
4.6. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, Y.; Li, W.J.; Su, Z.Y.; Kong, A.N.T. The complexity of the Nrf2 pathway: Beyond the antioxidant response. J. Nutr. Biochem. 2015, 26, 1401–1413. [Google Scholar] [CrossRef]
- Raghunath, A.; Sundarraj, K.; Nagarajan, R.; Arfuso, F.; Bian, J.S.; Kumar, A.P.; Sethi, G.; Perumal, E. Antioxidant response elements: Discovery, classes, regulation and potential applications. Redox Biol. 2018, 17, 297–314. [Google Scholar] [CrossRef] [PubMed]
- Paunkov, A.; Chartoumpekis, D.V.; Ziros, P.G.; Sykiotis, G.P. A Bibliometric review of the Keap1/Nrf2 pathway and its related antioxidant compounds. Antioxidants 2019, 8, 353. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Mo, Z. Keap1-Nrf2 signaling pathway in angiogenesis and vascular diseases. J. Tissue Eng. Regen. Med. 2020, 14, 869–883. [Google Scholar] [CrossRef]
- Leiser, S.F.; Miller, R.A. Nrf2 Signaling, a mechanism for cellular stress resistance in long-lived mice. Mol. Cell. Biol. 2010, 30, 871–884. [Google Scholar] [CrossRef] [Green Version]
- Uruno, A.; Motohashi, H. The Keap1-Nrf2 system as an in vivo sensor for electrophiles. Nitric Oxide-Biol. Chem. 2011, 25, 153–160. [Google Scholar] [CrossRef]
- Jain, A.; Lamark, T.; Sjøttem, E.; Larsen, K.B.; Awuh, J.A.; Øvervatn, A.; McMahon, M.; Hayes, J.D.; Johansen, T. p62/SQSTM1 is a target gene for transcription factor NRF2 and creates a positive feedback loop by inducing antioxidant response element-driven gene transcription. J. Biol. Chem. 2010, 285, 22576–22591. [Google Scholar] [CrossRef] [Green Version]
- Fan, W.; Tang, Z.; Chen, D.; Moughon, D.; Ding, X.; Chen, S.; Zhu, M.; Zhong, Q. Keap1 facilitates p62-mediated ubiquitin aggregate clearance via autophagy. Autophagy 2010, 6, 614–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, A.; Wang, X.-J.; Zhao, F.; Villeneuve, N.F.; Wu, T.; Jiang, T.; Sun, Z.; White, E.; Zhang, D.D. A noncanonical mechanism of Nrf2 activation by autophagy deficiency: Direct interaction between Keap1 and p62. Mol. Cell. Biol. 2010, 30, 3275–3285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komatsu, M.; Kurokawa, H.; Waguri, S.; Taguchi, K.; Kobayashi, A.; Ichimura, Y.; Sou, Y.S.; Ueno, I.; Sakamoto, A.; Tong, K.I.; et al. The selective autophagy substrate p62 activates the stress responsive transcription factor Nrf2 through inactivation of Keap1. Nat. Cell Biol. 2010, 12, 213–223. [Google Scholar] [CrossRef]
- Gacesa, R.; Dunlap, W.C.; Barlow, D.J.; Laskowski, R.A.; Long, P.F. Rising levels of atmospheric oxygen and evolution of Nrf2. Sci. Rep. 2016, 6, 27740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gacesa, R.; Dunlap, W.C.; Long, P.F. Bioinformatics analyses provide insight into distant homology of the Keap1–Nrf2 pathway. Free Radic. Biol. Med. 2015, 88, 373–380. [Google Scholar] [CrossRef]
- Mhamdi, A.; van Breusegem, F. Reactive oxygen species in pant development. Development 2018, 145, dev164376. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.H.; Jiang, M.; Li, R.Q.; Basnet, R.; Huang, J.Z.; Song, S.Y.; Shu, Q.Y. Identification, characterization, and mutational analysis of a probable Keap1 ortholog in rice (Oryza sativa L.). Plants 2020, 9, 1450. [Google Scholar] [CrossRef]
- Skubacz, A.; Daszkowska-Golec, A.; Szarejko, I. The role and regulation of ABI5 (ABA-Insensitive 5) in plant development, abiotic stress responses and phytohormone crosstalk. Front. Plant Sci. 2016, 7, 1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nonogaki, H. ABA responses during seed development and germination. Adv. Bot. Res. 2019, 92, 171–217. [Google Scholar] [CrossRef]
- Stone, S.L.; Williams, L.A.; Farmer, L.M.; Vierstra, R.D.; Callis, J. KEEP ON GOING, a RING E3 ligase essential for Arabidopsis growth and development, is involved in abscisic acid signaling. Plant Cell 2006, 18, 3415–3428. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Stone, S.L. Abscisic acid increases Arabidopsis ABI5 transcription factor levels by promoting KEG E3 ligase self-ubiquitination and proteasomal degradation. Plant Cell 2010, 22, 2630–2641. [Google Scholar] [CrossRef] [Green Version]
- Albertos, P.; Romero-Puertas, M.C.; Tatematsu, K.; Mateos, I.; Sánchez-Vicente, I.; Nambara, E.; Lorenzo, O. S-nitrosylation triggers ABI5 degradation to promote seed germination and seedling growth. Nat. Commun. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zou, M.J.; Guan, Y.C.; Ren, H.B.; Zhang, F.; Chen, F. Characterization of alternative splicing products of bZIP transcription factors OsABI5. Biochem. Biophysic. Res. Commun. 2007, 360, 307–313. [Google Scholar] [CrossRef]
- Zou, M.; Guan, Y.; Ren, H.; Zhang, F.; Chen, F. A bZIP transcription factor, OsABI5, is involved in rice fertility and stress tolerance. Plant Mol. Biol. 2008, 66, 675–683. [Google Scholar] [CrossRef]
- Du, L.; Xu, F.; Fang, J.; Gao, S.; Tang, J.; Fang, S.; Wang, H.; Tong, H.; Zhang, F.; Chu, J.; et al. Endosperm sugar accumulation caused by mutation of PHS8/ISA1 leads to pre-harvest sprouting in rice. Plant J. Cell Mol. Biol. 2018, 95, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Kim, D.; Han, S.-H.; Kim, S.-H.; Piao, W.; Yanagisawa, S.; An, G.; Paek, N.-C. Multilayered regulation of membrane-bound ONAC054 is essential for abscisic acid-induced leaf senescence in rice. Plant Cell 2020, 32, 630–649. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Deng, W.; Wang, X.; Yang, C.; Li, Z. Maize (Zea mays L.) homologue of ABA-insensitive (ABI) 5 gene plays a negative regulatory role in abiotic stresses response. Plant Growth Regul. 2012, 68, 383–393. [Google Scholar] [CrossRef]
- Zhou, X.; Yuan, F.; Wang, M.; Guo, A.; Zhang, Y.; Xie, C.G. Molecular characterization of an ABA insensitive 5 orthologue in Brassica oleracea. Biochem. Biophys. Res. Commun. 2013, 430, 1140–1146. [Google Scholar] [CrossRef]
- Collin, A.; Daszkowska-Golec, A.; Kurowska, M.; Szarejko, I. Barley ABI5 (Abscisic Acid INSENSITIVE 5) is involved in abscisic acid-dependent drought response. Front. Plant Sci. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Zinsmeister, J.; Lalanne, D.; Terrasson, E.; Chatelain, E.; Vandecasteele, C.; Vu, B.L.; Dubois-Laurent, C.; Geoffriau, E.; Signor, C.L.; Dalmais, M.; et al. ABI5 is a regulator of seed maturation and longevity in legumes. Plant Cell 2016, 28, 2735–2754. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 system: A thiol-based sensor-effector apparatus for maintaining redox homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Watai, Y.; Kobayashi, A.; Nagase, H.; Mizukami, M.; McEvoy, J.; Singer, J.D.; Itoh, K.; Yamamoto, M. Subcellular localization and cytoplasmic complex status of endogenous Keap1. Genes Cells 2007, 12, 1163–1178. [Google Scholar] [CrossRef] [PubMed]
- Mundy, J.; Yamaguchi-Shinozaki, K.; Chua, N.H. Nuclear proteins bind conserved elements in the abscisic acid-responsive promoter of a rice rab gene. Proc. Natl. Acad. Sci. USA 1990, 87, 1406–1410. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Wang, G.; Wu, H.; Fan, X.; Liang, L.; Zhao, H.; Li, S.; Hu, Y.; Liu, H.; Ayaad, M.; et al. OsMFT2 is involved in the regulation of ABA signaling-mediated seed germination through interacting with OsbZIP23/66/72 in rice. Plant J. 2020, 103, 532–546. [Google Scholar] [CrossRef]
- Duan, J.; Cai, W. OsLEA3-2, an abiotic stress induced gene of rice plays a key role in salt and drought tolerance. PLoS ONE 2012, 7, e45117. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Hu, T.; Zeng, H.; Zeng, H.; He, S.; He, S.; Wu, Y.; Wu, Y.; Wang, G.; Wang, G.; et al. Molecular analysis of OsLEA4 and its contributions to improve E. coli viability. Appl. Biochem. Biotechnol. 2012, 166, 222–233. [Google Scholar] [CrossRef]
- Hu, T.; Liu, Y.; Zhu, S.; Qin, J.; Li, W.; Zhou, N. Overexpression of OsLea14-A improves the tolerance of rice and increases Hg accumulation under diverse stresses. Environ. Sci. Pollut. Res. Int. 2019, 26, 10537–10551. [Google Scholar] [CrossRef] [PubMed]
- Hewage, K.A.H.; Yang, J.F.; Wang, D.; Hao, G.F.; Yang, G.F.; Zhu, J.K. Chemical manipulation of abscisic acid signaling: A new approach to abiotic and biotic stress management in agriculture. Adv. Sci. 2020, 7, 2001265. [Google Scholar] [CrossRef]
- Sun, Y.F.; Pri-Tal, O.; Michaeli, D.; Mosquna, A. Evolution of abscisic acid signaling module and its perception. Front. Plant Sci. 2020, 11, 934. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Ishii, T.; Wakabayashi, N.; Yamamoto, M. Regulatory mechanisms of cellular response to oxidative stress. Free Radical Res. 1999, 31, 9–324. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, S.B.; Diehl, J.A. PERK-dependent activation of Nrf2 contributes to redox homeostasis and cell survival following endoplasmic reticulum stress. J. Biol. Chem. 2004, 279, 20108–20117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhry, S.; Zhang, Y.; McMahon, M.; Sutherland, C.; Cuadrado, A.; Hayes, J.D. Nrf2 is controlled by two distinct beta-TrCP recognition motifs in its Neh6 domain, one of which can be modulated by GSK-3 activity. Oncogene 2013, 32, 3765–3781. [Google Scholar] [CrossRef] [Green Version]
- Lynch, T.; Erickson, B.J.; Finkelstein, R.R. Direct interactions of ABA insensitive (ABI)-clade protein phosphatase (PP) 2Cs with calcium-dependent protein kinases and ABA response element-binding bZIPs may contribute to turning off ABA response. Plant Mol. Biol. 2012, 80, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Hao, H.; Zhang, Y.; Bai, Y.; Zhu, W.; Qin, Y.; Yuan, F.F.; Zhao, F.Y.; Wang, M.Y.; Hu, J.J.; et al. SOS2- like protein kinase5, an SNF1-related protein kinase3-type protein kinase, is important for abscisic acid responses in Arabidopsis through phosphorylation of ABSCISIC ACID-INSENSITIVE5. Plant Physiol. 2015, 168, 659–676. [Google Scholar] [CrossRef] [Green Version]
- Finkelstein, R.R. Mutations at two new Arabidopsis ABA response loci are similar to the abi3 mutations. Plant J. 1994, 5, 765–771. [Google Scholar] [CrossRef]
- Finkelstein, R.; Gampala, S.S.; Lynch, T.J.; Thomas, T.L.; Rock, C.D. Redundant and distinct functions of the ABA response loci ABA-INSENSITIVE (ABI) 5 and ABRE-BINDING FACTOR (ABF) 3. Plant Mol. Biol. 2005, 59, 253–267. [Google Scholar] [CrossRef]
- Bi, C.; Ma, Y.; Wu, Z.; Yu, Y.T.; Liang, S.; Lu, K.; Wang, X.F. Arabidopsis ABI5 plays a role in regulating ROS homeostasis by activating CATALASE 1 transcription in seed germination. Plant Mol. Biol. 2017, 94, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Bhagat, P.K.; Verma, D.; Verma, N.; Sinha, A.K. A novel positive feedback mechanism of ABI5 phosphorylation by mitogen activated protein kinase-3 regulates ABA signaling in Arabidopsis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Rodriguez, P.L.; Lozano-Juste, J.; Albert, A. PYR/PYL/RCAR ABA receptors. Adv. Bot. Res. 2019, 92, 51–82. [Google Scholar] [CrossRef]
- Zhao, W.; Guan, C.; Feng, J.; Liang, Y.; Zhan, N.; Zuo, J.; Ren, B. The Arabidopsis CROWDED NUCLEI genes regulate seed germination by modulating degradation of ABI5 protein. J. Integr. Plant Biol. 2016, 58, 669–678. [Google Scholar] [CrossRef]
- Malhotra, D.; Portales-Casamar, E.; Singh, A.; Srivastava, S.; Arenillas, D.; Happel, C.; Shyr, C.; Wakabayashi, N.; Kensler, T.W.; Wasserman, W.W.; et al. Global mapping of binding sites for Nrf2 identifies novel targets in cell survival response through ChIP-Seq profiling and network analysis. Nucleic Acids Res. 2010, 38, 5718–5734. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Song, S.; Chen, Y.; Liu, L.; Wang, Y.; Bao, S.; Zhou, X.; Teo, Z.W.N.; Mao, C.; Gan, Y.; Yu, H. OsFTIP1-mediated regulation of florigen transport in rice is negatively regulated by the ubiquitin-like domain kinase OsUbDKγ4. Plant Cell 2017, 29, 491–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, F.; Chen, S.; Ning, Y.; Wang, G.-L. Rice (Oryza sativa) protoplast isolation and its application for transient expression analysis. Curr. Protoc. Plant Biol. 2016, 1, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Citovsky, V.; Lee, L.-Y.; Vyas, S.; Glick, E.; Chen, M.-H.; Vainstein, A.; Gafni, Y.; Gelvin, S.B.; Tzfira, T. Subcellular localization of interacting proteins by bimolecular fluorescence complementation in planta. J. Mol. Biol. 2006, 362, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Song, S.; Gan, Y.; Jiang, L.; Yu, H.; Shen, L. SHAGGY-like kinase 12 regulates flowering through mediating CONSTANS stability in Arabidopsis. Sci. Adv. 2020, 6, eaaw0413. [Google Scholar] [CrossRef] [PubMed]
- Basnet, R.; Zhang, J.; Hussain, N.; Shu, Q. Characterization and mutational analysis of a monogalactosyldiacylglycerol synthase gene OsMGD2 in rice. Front. Plant Sci. 2019, 10, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Primers | Forward Strand (5’–3’) | Reverse Strand (5’–3’) |
|---|---|---|
| Y2H analysis | ||
| KEAP1-AD | gccatggaggccagtgaattcATGGGTGCTGGAAAGAAGACTCA | cagctcgagctcgatggatccAATGGCAACGGCGCATGC |
| Kelch-AD | gccatggaggccagtgaattcGCCCGTGCATATGCGGCT | cagctcgagctcgatggatccGCTGAGTGGGCTCCCCCT |
| OsABI5-BD | gaattcATGAACATGGACGAGTTTGT | ggatccCCACATGCAGCTGCCGCTGC |
| BiFC analysis | ||
| Kelch-nEYFP | ctcaagcttcgaattcGCCCGTGCATATGCGGCT | gtcgactgcagaattcGCTGAGTGGGCTCCCCCT |
| cEYFP-OsABI5 | gaattctgcagtcgacATGGCATCGGAGATGAGCA | cgcggtaccgtcgacCCACATGCAGCTGCCGCTGC |
| Subcellular localization analysis | ||
| OsKEAP1GFP | gaattcATGGGTGCTGGAAAGAAGACTCA | ggatccCAATGGCAACGGCGCATGC |
| OsABI5GFP | gaattcATGGCATCGGAGATGAGCA | ggatccCCCACATGCAGCTGCCGCTGC |
| Name of Primers | Gene Numbers | Forward Strand (5’–3’) | Reverse Strand (5’–3’) |
|---|---|---|---|
| qrt-OsKEAP1 | Os01g0165200 | CAAGCACTGGCCAGCTTAAT | GATTAGCGCGAACAGGAGCA |
| qrt-OsABI5 | Os01g0859300 | GCTAACGACCGACAGGTAACACT | CCATCCCGTTGTACCCACC |
| qrt-OsABI1 | Os05g0572700 | ACGAGTTGGAACGAGTGGAAGC | GGCTTCAGGTAGTAGTCGCCTATG |
| qrt-OsABI2 | Os01g0513100 | CGCGATCAATAGGTGACAGA | CGCTAATGTTGTCCTTGCTC |
| qrt-OsABI3 | Os08g0101000 | CCCAACAACAAAAGCAGGAT | CCTTTGTATTGGACGAGACG |
| qrt-OsSnRK2 | Os03g0764800 | TGGCAAGACTGCTGATGTATGG | TCGAAAGGATATGCACCAACTACC |
| qrt-OsKEG | Os05g0392000 | GGCCAACATGTGCAGCTCAAAC | CTGACCACGCCATCCAAATCTAGG |
| qrt-OsDSG1 | Os09g0434200 | CCGCTTTGGAAGAATCTCTG | TTCCTGTCTTCCTCCTCTTC |
| qrt-OsMFT2 | Os01g0111600 | ACGGTGGGGATACACAGGTA | TGTGTTGAAGTTGGGCCTGT |
| qrt-Rab16A | Os11g0454300 | CAGCTCAAGCTCGTCTGA | GCTTCTCCTTGATCTTCTCCTT |
| qrt-OsbZIP23 | Os02g0766700 | AGAGCAATGTGTTCCCTCCG | ATCTTGCCGAAGCCATTGGA |
| qrt-OsLEA3-2 | Os03g0322900 | AAGATGTGATCCCCCATGAGC | TTCAGCACCACTGCACTTAGA |
| qrt-OsLEA14 | Os01g0705200 | GATGTACACGCTCGGGATGT | TTCCAGGCTTGTAGGTGCTG |
| qrt-OsLEA4 | Os01g0705200 | AGTTCATGTAGAGTTCACTTCGCT | GCATCTCCCACAACATGATACC |
| Actin | Os03g0836000 | CTTCATAGGAATGGAAGCTGCGGGTA | CGACCACCTTGATCTTCATGCTGCTA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.-H.; Jiang, M.; Li, R.-Q.; Huang, J.-Z.; Shu, Q.-Y. OsKEAP1 Interacts with OsABI5 and Its Downregulation Increases the Transcription of OsABI5 and the ABA Response Genes in Germinating Rice Seeds. Plants 2021, 10, 527. https://doi.org/10.3390/plants10030527
Liu Y-H, Jiang M, Li R-Q, Huang J-Z, Shu Q-Y. OsKEAP1 Interacts with OsABI5 and Its Downregulation Increases the Transcription of OsABI5 and the ABA Response Genes in Germinating Rice Seeds. Plants. 2021; 10(3):527. https://doi.org/10.3390/plants10030527
Chicago/Turabian StyleLiu, Yan-Hua, Meng Jiang, Rui-Qing Li, Jian-Zhong Huang, and Qing-Yao Shu. 2021. "OsKEAP1 Interacts with OsABI5 and Its Downregulation Increases the Transcription of OsABI5 and the ABA Response Genes in Germinating Rice Seeds" Plants 10, no. 3: 527. https://doi.org/10.3390/plants10030527