
Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid

 ,
,  , ,
, ,
Abstract
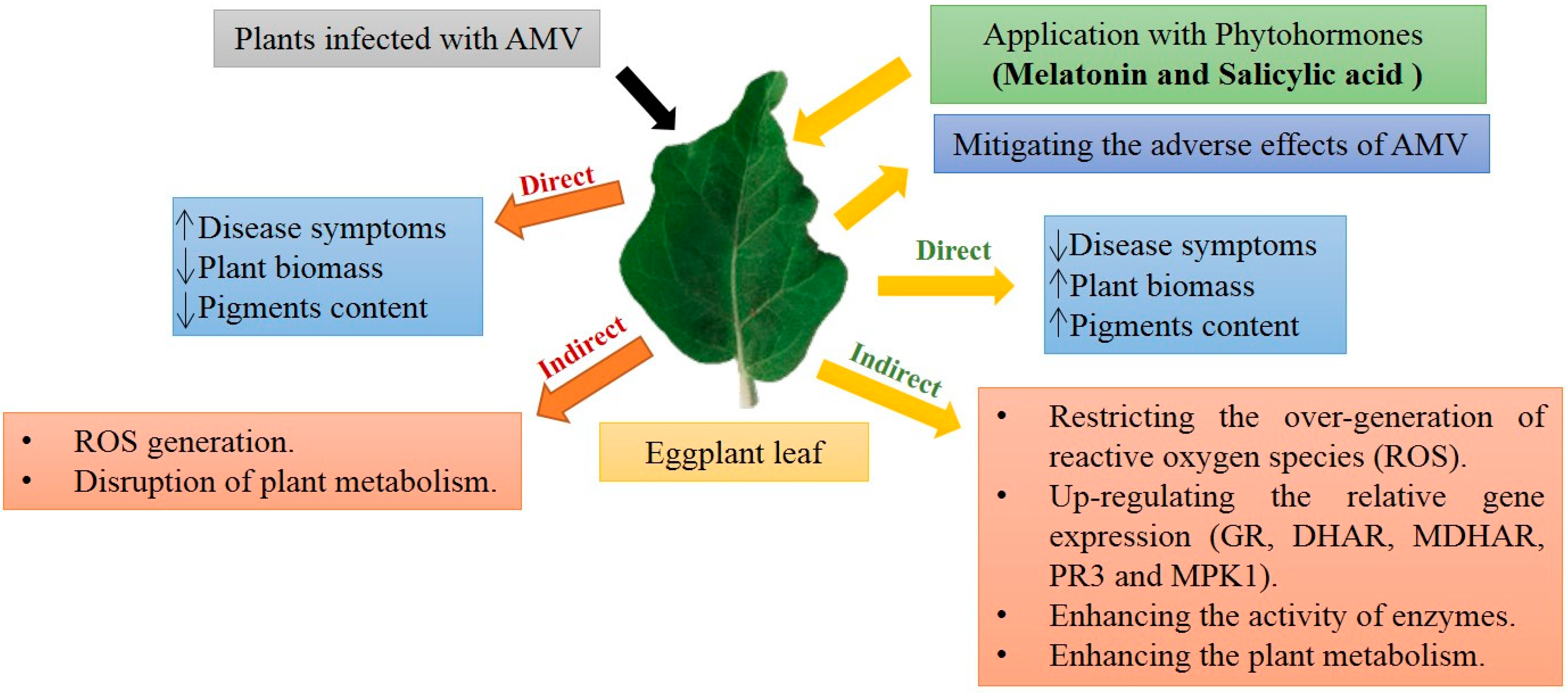
:1. Introduction
2. Results
2.1. Symptomatology, Examination and Biological Purification of AMV-Eggplant Egyptian Isolate
2.2. Molecular Characterization of AMV-Eggplant Egyptian Isolate
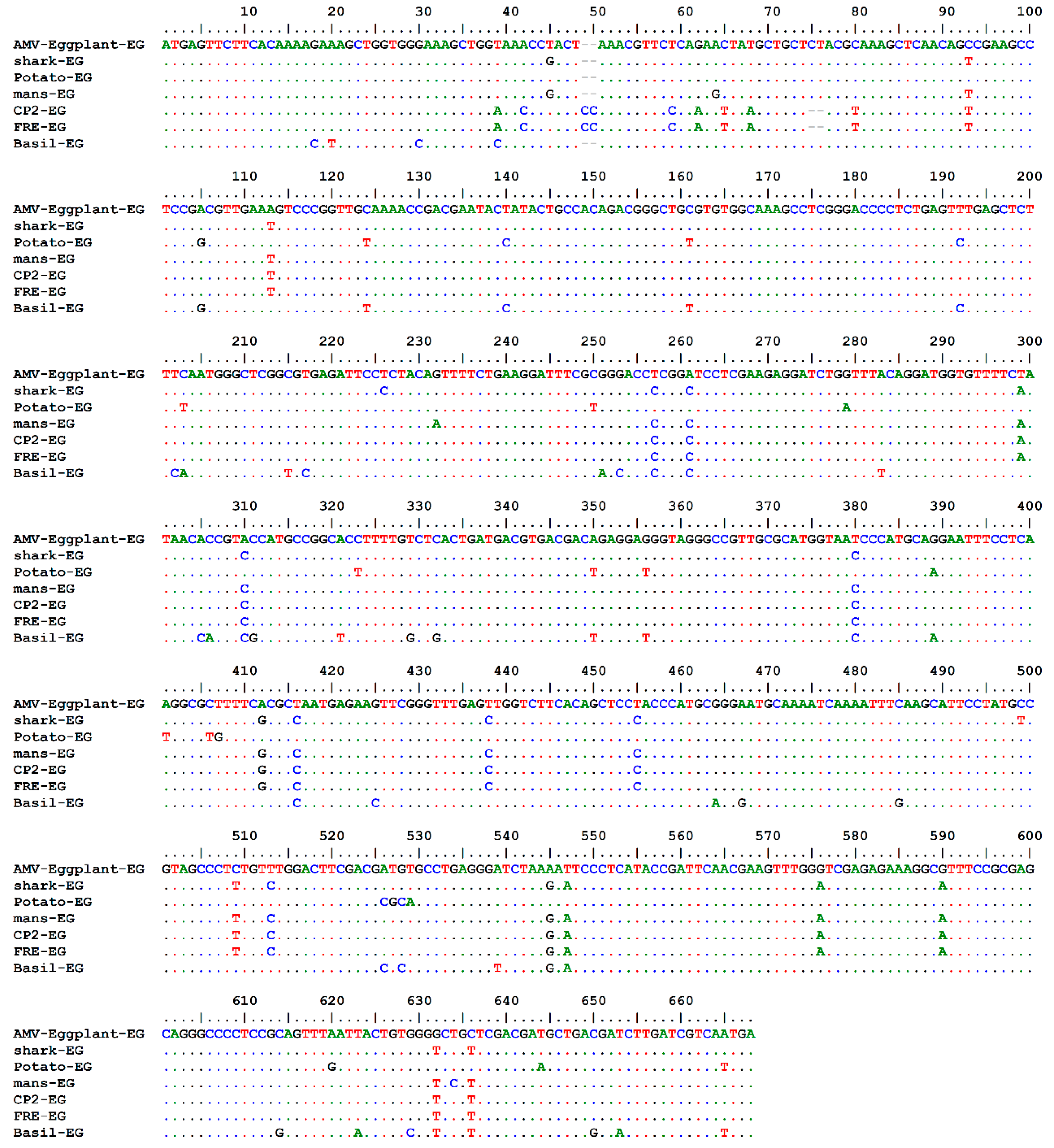
2.2.1. AMV-Eggplant-EG Coat Protein Gene Amplification and Sequencing
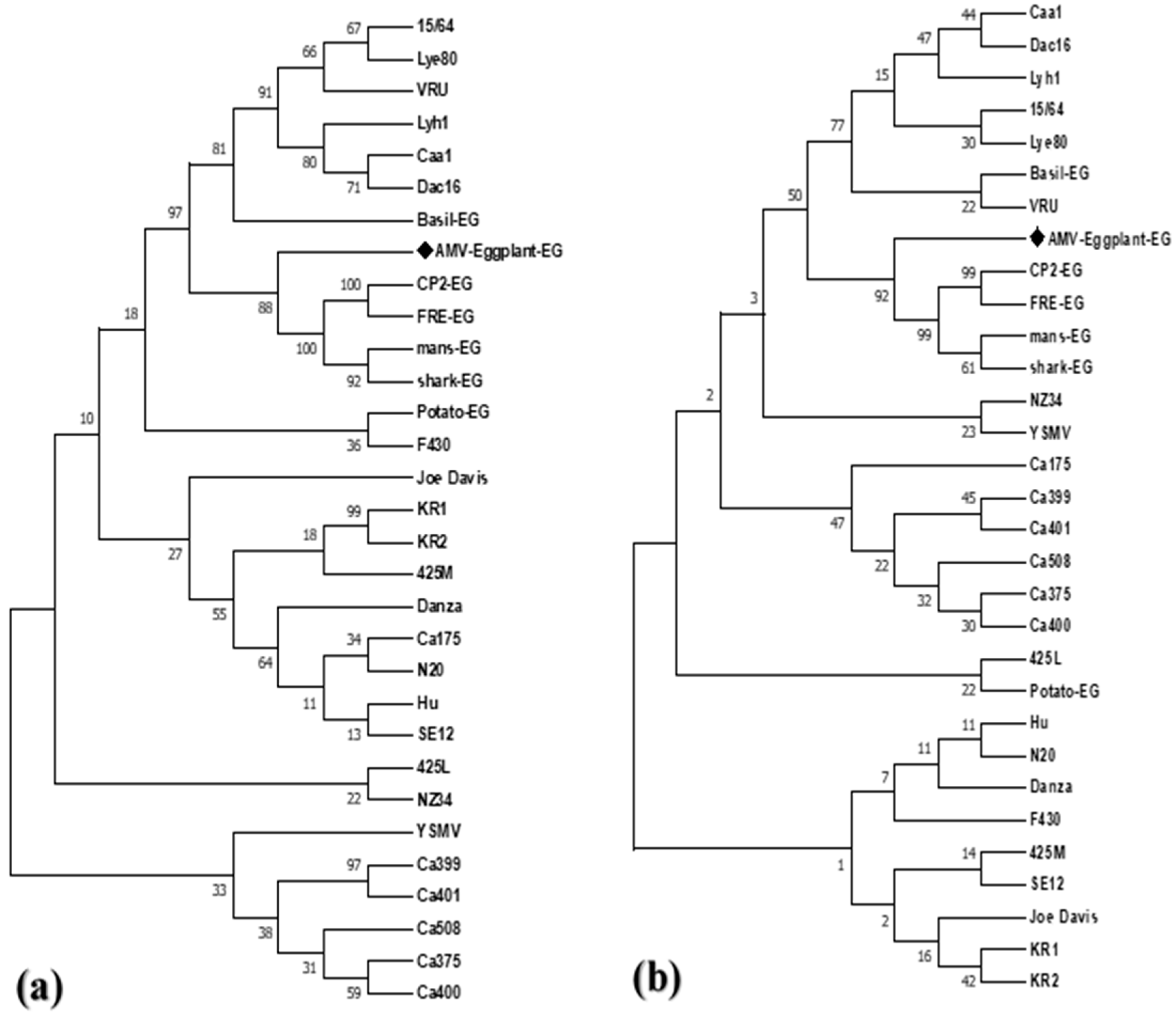
2.2.2. Viroinformatics Analysis of AMV-Eggplant-EG/CP Gene
2.3. Systemic Protection against Alfalfa mosaic virus (AMV) in Eggplant
2.4. Physiological and Biochemical Studies
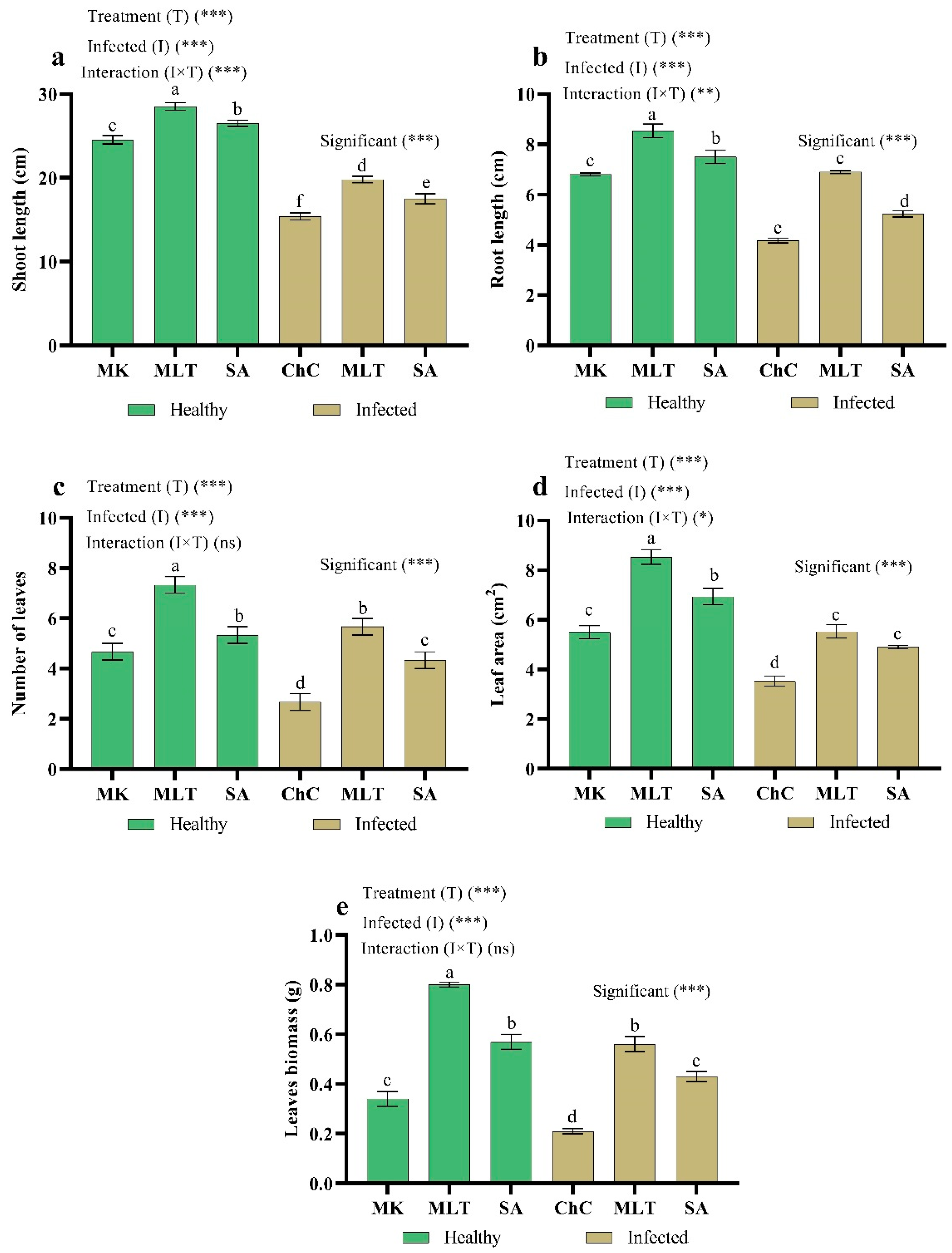
2.4.1. Changes in Plant Growth
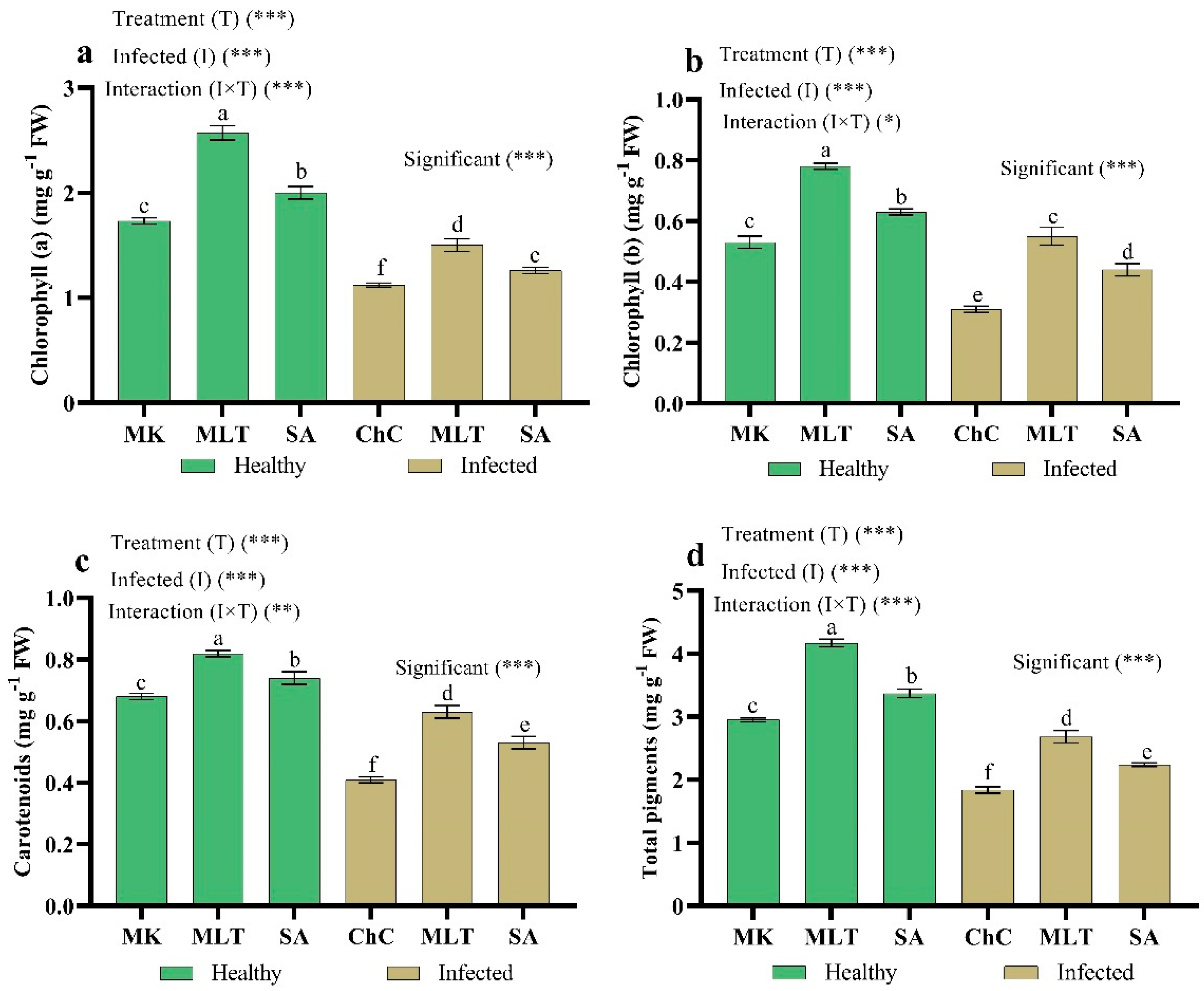
2.4.2. Changes in Photosynthetic Pigments
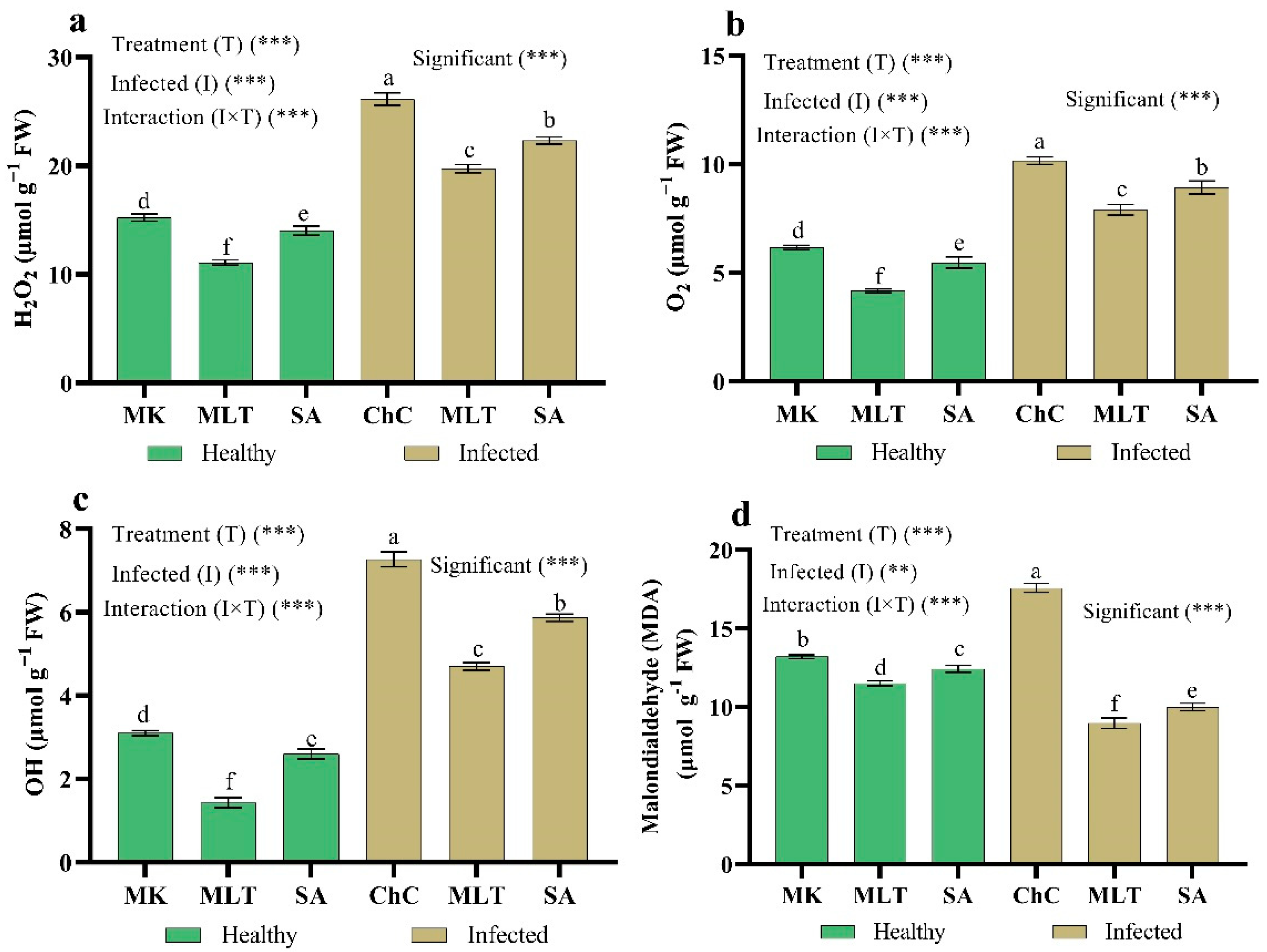
2.4.3. Changes in ROS Damage
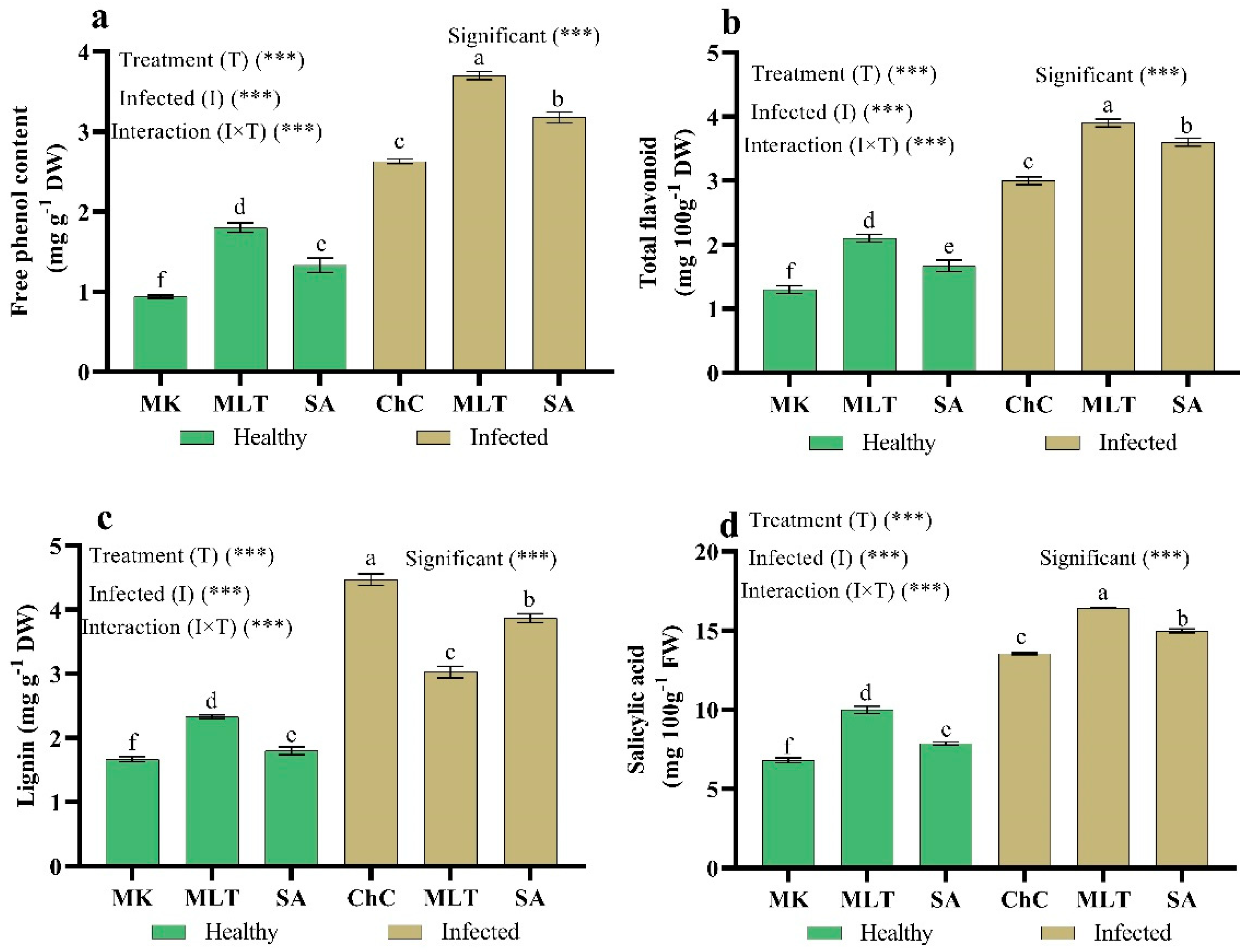
2.4.4. Changes in Secondary Metabolites and Salicylic Acid Content
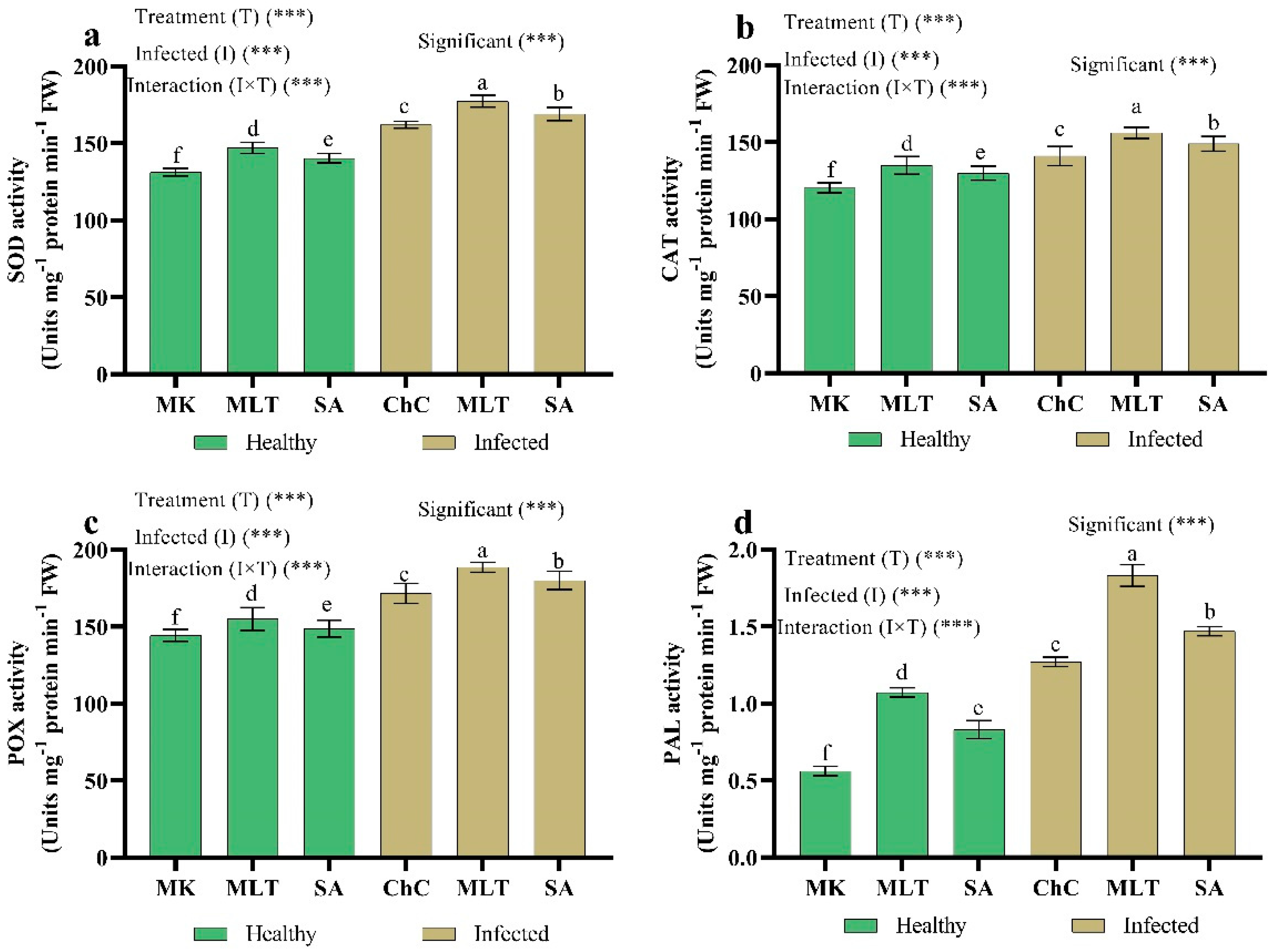
2.4.5. Changes in Antioxidant Enzymes
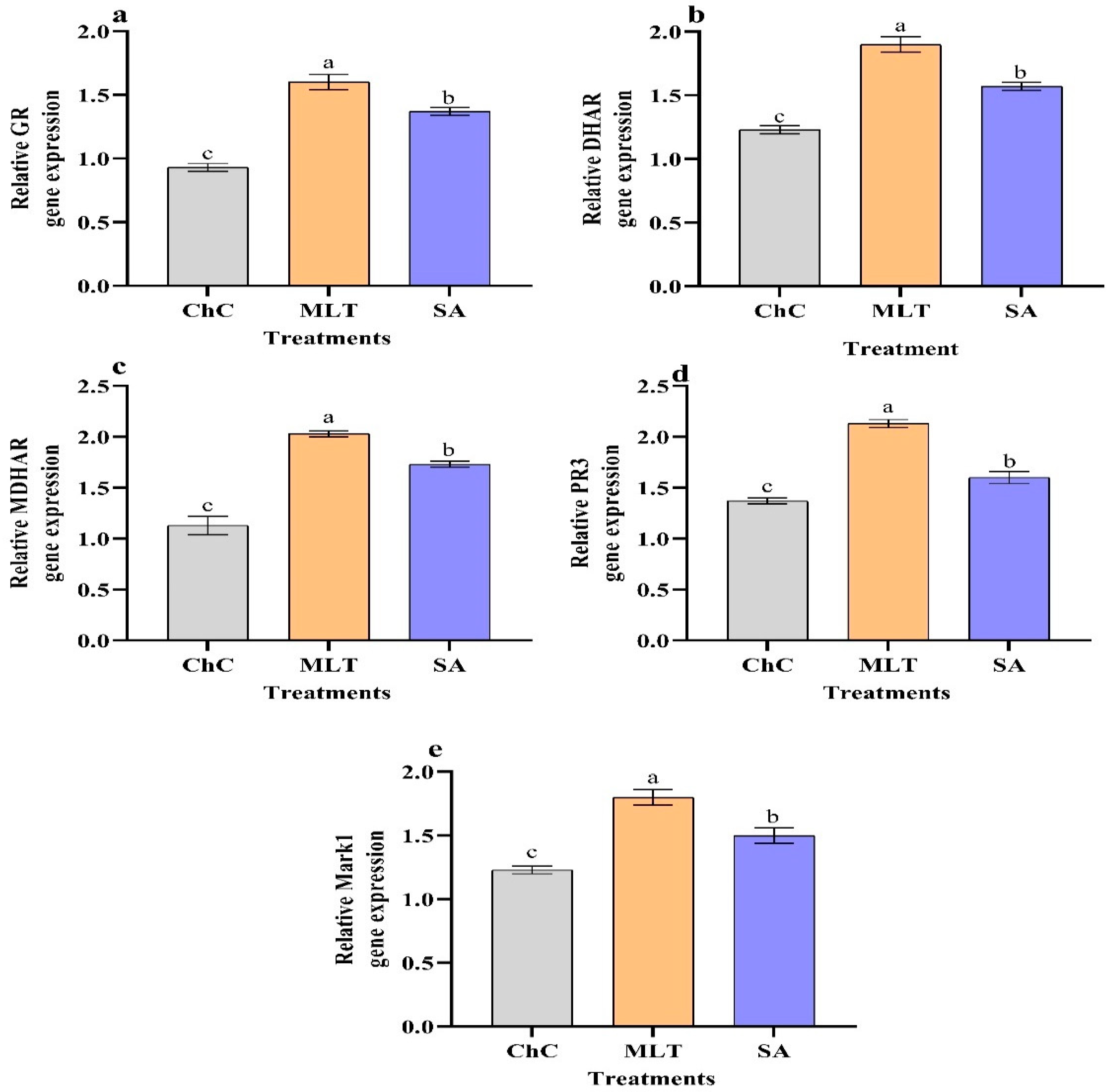
2.4.6. Changes in Gene Expression
3. Discussion
4. Materials and Methods
4.1. Sample Collection and Virus Detection
4.2. Isolation and Propagation of Virus
4.3. Total RNA Extraction and RT-PCR Amplification of AMV/CP Gene and Sequencing
4.4. Multiple Alignments and Phylogenetic Analysis
4.5. Plant Materials, Eggplant Treatments, and Alfalfa mosaic virus Inoculation
- The first group, the plants were inoculated with only phosphate buffer without virus as mock inoculation (MK).
- The second group, the plants were foliar sprayed with 100 µM melatonin (MLT), combined with two drops of Tween 80 into the leaves until run-off.
- The third group, the plants were foliar sprayed with 100 µM salicylic acid (SA), combined with two drops of Tween 80 into the leaves until run-off.
- The fourth group, the plants were inoculated with Alfalfa mosaic virus as challenge control (ChC).
- The fifth group, the plants were foliar sprayed with 100 µM MLT and then inoculated after three days with AMV.
- The sixth group, the plants were foliar sprayed with 100 µM SA and then inoculated after three days with AMV.
4.6. Growth Indices
4.7. Biochemical Analysis
4.7.1. Determination of Photosynthetic Pigments
4.7.2. Determination of Oxidative Damage Marker
4.7.3. Determination of Total Phenolic and Flavonoid Compounds
4.7.4. Determination of Lignin
4.7.5. Determination of Salicylic Acid
4.7.6. Enzymatic Antioxidant Assays
4.7.7. RT-PCR Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balconi, C.; Stevanato, P.; Motto, M.; Biancardi, E. Breeding for Biotic Stress Resistance/Tolerance in Plants. In Crop Production for Agricultural Improvement; Springer: Berlin/Heidelberg, Germany, 2012; pp. 57–114. [Google Scholar]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; El-Dougdoug, N.K. Potential Effect of Plant Growth-Promoting Rhizobacteria (PGPR) on Enhancing Protection Against Viral Diseases. In Field Crops: Sustainable Management by PGPR. Sustainable Development and Biodiversity; Maheshwari, D.K., Dheeman, S., Eds.; Springer: Cham, Switzerland, 2019; Volume 23, pp. 411–445. [Google Scholar]
- Wang, S.; Xilin, H.; Ying, L. Effects of Turnip mosaic virus (TuMV) on endogenous hormones and transcriptional level of related genes in infected non-heading Chinese cabbage. J. Nanjing Agric. Univ. 2011, 34, 13–19. [Google Scholar]
- Sofy, A.R.; Dawoud, R.A.; Sofy, M.R.; Mohamed, H.I.; Hmed, A.A.; El-Dougdoug, N.K. Improving regulation of enzymatic and non-enzymatic antioxidants and stress-related gene stimulation in Cucumber mosaic cucumovirus-infected cucumber plants treated with glycine betaine, chitosan and combination. Molecules 2020, 25, 2341. [Google Scholar] [CrossRef] [PubMed]
- Sofy, A.R.; Hmed, A.A.; Alnaggar, A.E.-A.M.; Dawoud, R.A.; Elshaarawy, R.F.M.; Sofy, M.R. Mitigating effects of Bean yellow mosaic virus infection in faba bean using new carboxymethyl chitosan-titania nanobiocomposites. Int. J. Biol. Macromol. 2020, 163, 1261–1275. [Google Scholar] [CrossRef] [PubMed]
- Megahed, A.A.; El-Dougdoug, K.A.; Othman, B.A.; Lashin, S.M.; Ibrahim, M.A.; Sofy, A.R. Induction of resistance in tomato plants against Tomato mosaic tobamovirus using beneficial microbial isolates. Pak. J. Biol. Sci. 2013, 16, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Elshaarawy, R.F.M.; Mustafa, F.H.A.; Sofy, A.R.; Hmed, A.A.; Janiak, C. A new synthetic antifouling coatings integrated novel aminothiazole-functionalized ionic liquids motifs with enhanced antibacterial performance. J. Environ. Chem. Eng. 2019, 7, 102800. [Google Scholar] [CrossRef]
- Mangino, G.; Plazas, M.; Vilanova, S.; Prohens, J.; Gramazio, P. Performance of a set of eggplant (Solanum melongena) lines with introgressions from its wild relative S. incanum under open field and screenhouse conditions and detection of QTLs. Agronomy 2020, 10, 467. [Google Scholar] [CrossRef] [Green Version]
- Brenes, M.; Solana, A.; Boscaiu, M.; Fita, A.; Vicente, O.; Calatayud, A.; Prohens, J.; Plazas, M. Physiological and biochemical responses to salt stress in cultivated eggplant (Solanum melongena L.) and in S. insanum L., a close wild relative. Agronomy 2020, 10, 651. [Google Scholar] [CrossRef]
- Fu, Q.S.; Yang, R.C.; Wang, H.S.; Zhao, B.; Zhou, C.L.; Ren, S.X.; Guo, Y.D. Leaf morphological and ultrastructural performance of eggplant (Solanum melongena L.) in response to water stress. Photosynthetica 2013, 51, 109–114. [Google Scholar] [CrossRef]
- Sastry, K.S.; Mandal, B.; Hammond, J.; Scott, S.W.; Briddon, R.W. Solanum melongena (Brinjal, Eggplant, Aubergine). In Encyclopedia of Plant Viruses and Viroids; Springer: New Delhi, India, 2019; pp. 2382–2393. [Google Scholar]
- Shalaby, A.A. Characterization of an Eggplant mosaic virus strain isolated from naturally infected eggplant in Egypt. Ann. Agric. Sci. Moshtohor 1997, 35, 1315–1324. [Google Scholar]
- Ozdemir, S.; Erilmez, S.; Paylan, I.C. First report of Alfalfa mosaic virus in eggplant in Turkey. J. Plant Pathol. 2011, 93, 82. [Google Scholar]
- Al-Shahwan, I.M.; Abdalla, O.A.; Al-Saleh, M.A.; Amer, M.A. Detection of new viruses in alfalfa, weeds and cultivated plants growing adjacent to alfalfa fields in Saudi Arabia. Saudi J. Biol. Sci. 2017, 24, 1336–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaspars, E.M.; Bos, L. Alfalfa Mosaic Virus. In Descriptions of Plant Viruses; No. 229; Commonw. Mycol. Inst./Assoc. Appl. Biol.: Kew, UK, 1980. [Google Scholar]
- McLaughlin, M.R. A Greenhouse Method for Apphid Inoculation of Alfalfa Mosaic Virus in White Clover By Co-Culture of Virus, Vector, and Clover; Miscellaneous Publication-Agricultural Experiment Station, Oklahoma State University: Oklahoma, IN, USA, 1991; Volume 132, p. 318. [Google Scholar]
- Valkonen, J.P.T.; Pehu, E.; Watanabe, K. Symptom expression and seed transmission of Alfalfa mosaic virus and Potato yellowing virus (SB-22) in Solanum brevidens and S. etuberosum. Potato Res. 1992, 35, 403–410. [Google Scholar] [CrossRef]
- Jaspars, E.M.J. Interaction of Alfalfa mosaic virus nucleic acid and protein. Mol. Plant Virol. 2018, 1, 155–221. [Google Scholar]
- Bol, J.F. Alfalfa mosaic virus and ilarviruses: Involvement of coat protein in multiple steps of the replication cycle. J. Gen. Virol. 1999, 80, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Erny, C.; Schoumacher, F.; Jung, C.; Gagey, M.J.; Godefroy-Colburn, T.; Stussi-Garaud, C.; Berna, A. An N-proximal sequence of the Alfalfa mosaic virus movement protein is necessary for association with cell walls in transgenic plants. J. Gen. Virol. 1992, 73, 2115–2119. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, C.; Wu, K.; Chen, W.; Chen, Y.; Hao, X.; Wu, Y. Advances and prospects in biogenic substances against plant virus: A review. Pestic. Biochem. Physiol. 2017, 135, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Iriti, M.; Faoro, F. Abscisic acid is involved in chitosan-induced resistance to Tobacco necrosis virus (TNV). Plant Physiol. Biochem. 2008, 46, 1106–1111. [Google Scholar] [CrossRef]
- El-Dougdoug, K.A.; Sofy, A.R.; Hameed, G.A.; Dawood, R.A. Intraspecific diversity of Cucumber mosaic Cucumoviridae in Egypt. Int. J. Virol. 2014, 10, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical priming of plants against multiple abiotic stresses: Mission possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Arnao, M.; Hernández-Ruiz, J. Melatonin and reactive oxygen and nitrogen species: A model for the plant redox network. Melatonin Res. 2019, 2, 152–168. [Google Scholar] [CrossRef]
- Zhang, J.; Li, D.; Wei, J.; Ma, W.; Kong, X.; Rengel, Z.; Chen, Q. Melatonin alleviates aluminum-induced root growth inhibition by interfering with nitric oxide production in Arabidopsis. Environ. Exp. Bot. 2019, 161, 157–165. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trends Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. The Multi-Regulatory Properties of Melatonin in Plants. In Neurotransmitters in Plants; Roshchina, V.V., Ramakrishna, A., Eds.; Taylor & Francis-CRC: New York, NY, USA, 2018; pp. 71–101. [Google Scholar]
- Zhao, L.; Chen, L.; Gu, P.; Zhan, X.; Zhang, Y.; Hou, C.; Wu, Z.; Wu, Y.F.; Wang, Q.C. Exogenous application of melatonin improves plant resistance to virus infection. Plant Pathol. 2019, 68, 1287–1295. [Google Scholar] [CrossRef]
- Bonilla, E.; Valero, N.; Chacín-Bonilla, L.; Medina-Leendertz, S. Melatonin and viral infections. J. Pineal Res. 2004, 36, 73–79. [Google Scholar] [CrossRef]
- Falcioni, T.; Ferrio, J.P.; del Cueto, A.I.; Giné, J.; Achón, M.A.; Medina, V. Effect of salicylic acid treatment on tomato plant physiology and tolerance to Potato virus X infection. Eur. J. Plant Pathol. 2014, 138, 331–345. [Google Scholar] [CrossRef]
- Mohamed, H.I.; El-Beltagi, H.S.; Aly, A.A.; Latif, H.H. The role of systemic and non systemic fungicides on the physiological and biochemical parameters in Gossypium hirsutum plant, implications for defense responses. Fresenius Environ. Bull. 2018, 27, 8585–8593. [Google Scholar]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing adverse effects of Pb on maize plants by combined treatment with jasmonic, salicylic acids and proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- White, R.F. Acetylsalicylic acid (aspirin) induces resistance to Tobacco mosaic virus in tobacco. Virology 1979, 99, 410–412. [Google Scholar] [CrossRef]
- Li, T.; Huang, Y.; Xu, Z.; Wang, F.; Xiong, A. Salicylic acid-induced differential resistance to the Tomato yellow leaf curl virus among resistant and susceptible tomato cultivars. BMC Plant Biol. 2019, 19, 173. [Google Scholar] [CrossRef]
- Al-Saleh, M.A.; Amer, M.A. Biological and molecular variability of Alfalfa mosaic virus affecting alfalfa crop in Riyadh region. Plant Pathol. J. 2013, 29, 410–417. [Google Scholar] [CrossRef]
- Vyanjane, N.; Mali, V. Occurrence of Alfalfa mosaic virus on brinjal in India. Indian Phytopathol. 1981, 34, 164–168. [Google Scholar]
- Xu, H.; Nie, J. Identification, characterization, and molecular detection of Alfalfa mosaic virus in potato. Phytopathology 2006, 96, 1237–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chittoor, J.M.; Leach, J.E.; White, F.F. Induction of Peroxidase During Defense Against Pathogens. In Pathogenesis Related Proteins in Plants; Datta, S.K., Muthukrishnnan, S., Eds.; CRC Press: Boca Raton, FL, USA, 1999; pp. 171–193. [Google Scholar]
- Radwan, D.E.M.; Fayez, K.A.; Younis Mahmoud, S.; Lu, G. Modifications of antioxidant activity and protein composition of bean leaf due to Bean yellow mosaic virus infection and salicylic acid treatments. Acta Physiol. Plant. 2010, 32, 891–904. [Google Scholar] [CrossRef]
- Friedman, M. Overview of antibacterial, antitoxin, antiviral, and antifungal activities of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, 116–134. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, N.; Nagendra-Prasad, D.; Mohan, N.; Murugesan, K. Induction of systemic resistance in Lycopersicon esculentum cv. PKM1 (tomato) against Cucumber mosaic virus by using ozone. J. Virol. Methods 2007, 139, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Mayers, C.N.; Lee, K.; Moore, C.A.; Wong, S.; Carr, J.P. Salicylic acid-induced resistance to Cucumber mosaic virus in squash and Arabidopsis thaliana: Contrasting mechanisms of induction and antiviral action. Mol. Plant Microbe Interact. 2005, 18, 428–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shazly, M.A.; Attia, Y.A.; Kabil, F.F.; Anis, E.; Hazman, M. Inhibitory effects of salicylic acid and silver nanoparticles on Potato virus Y-infected potato plants in Egypt. Middle East J. Agric. Res. 2017, 6, 835–848. [Google Scholar]
- Radwan, D.E.M.; Lu, G.; Fayez, K.A.; Mahmoud, S.Y. Protective action of salicylic acid against Bean yellow mosaic virus infection in Vicia faba leaves. J. Plant Physiol. 2008, 165, 845–857. [Google Scholar] [CrossRef]
- Abd El-Gawad, H.G.; Bondok, A.M. Response of tomato plants to salicylic acid and chitosan under infection with Tomato mosaic virus. Am. Eur. J. Agric. Environ. Sci. 2015, 15, 1520–1529. [Google Scholar]
- Vicente, M.R.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Fariduddin, Q.; Ali, B.; Ahmad, A. Effect of salicylic acid on growth and enzyme activities of wheat seedlings. Acta Agron. Hung. 2005, 53, 433–437. [Google Scholar] [CrossRef]
- Farahat, A.S.; El-Morsi, A.A.; Soweha, H.E.; Sofy, A.R.; Refaey, E.E. Metabolic changes of cucumber plants due to two CMV Egyptian isolates. Arab Univ. J. Agric. Sci. 2018, 26, 2019–2028. [Google Scholar] [CrossRef] [Green Version]
- Rahoutei, J.; García-Luque, I.; Barón, M. Inhibition of photosynthesis by viral infection: Effect on PSII structure and function. Physiol. Plant. 2000, 110, 286–292. [Google Scholar] [CrossRef]
- Balachandran, S.; Hurry, V.M.; Kelley, S.E.; Osmond, C.B.; Robinson, S.A.; Rohozinski, J.; Seaton, G.G.R.; Sims, D.A. Concepts of plant biotic stress. Some insights into the stress physiology of virus-infected plants, from the perspective of photosynthesis. Physiol. Plant. 1997, 100, 203–213. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Z.; Lan, G.; Jiao, C.; Sun, Y. Effect of exogenous melatonin on resistance of cucumber to downy mildew. Sci. Hortic. 2019, 255, 231–241. [Google Scholar] [CrossRef]
- Ding, F.; Liu, B.; Zhang, S. Exogenous melatonin ameliorates cold-induced damage in tomato plants. Sci. Hortic. 2017, 219, 264–271. [Google Scholar] [CrossRef]
- Pacheco, A.; Cabral, C.; Fermino, E.; Aleman, C. Salicylic acid-induced changes to growth, flowering and flavonoids production in marigold plants. J. Med. Plants Res. 2013, 1, 95–100. [Google Scholar]
- Javaheri, M.; Mashayekhi, K.; Dadkhah, A.; Tavallaee, F. Effects of salicylic acid on yield and quality characters of tomato fruit (Lycopersicum esculentum Mill.). Int. J. Agric. Crop Sci. 2012, 4, 1184–1187. [Google Scholar]
- El-Dougdoug, K.A.; Dawoud, R.A.; Rezk, A.A.; Sofy, A.R. Incidence of fruit trees viroid diseases by tissue print hybridization in Egypt. Int. J. Virol. 2012, 8, 114–120. [Google Scholar] [CrossRef]
- Abd El-Rahman, S.S.; Mohamed, H.I. Application of benzothiadiazole and Trichoderma harzianum to control faba bean chocolate spot disease and their effect on some physiological and biochemical traits. Acta Physiol. Plant. 2014, 36, 343–354. [Google Scholar] [CrossRef]
- Abd El-Rahman, S.S.; Mazen, M.M.; Mohamed, H.I.; Mahmoud, N.M. Induction of defence related enzymes and phenolic compounds in lupin (Lupinus albus L.) and their effects on host resistance against Fusarium wilt. Eur. J. Plant Pathol. 2012, 134, 105–116. [Google Scholar] [CrossRef]
- Xi, D.; Feng, H.; Lan, L.; Du, J.; Wang, J.; Zhang, Z.; Xue, L.; Xu, W.; Lin, H. Characterization of synergy between Cucumber mosaic virus and Tobacco necrosis virus in Nicotiana benthamiana. J. Phytopathol. 2007, 155, 570–573. [Google Scholar] [CrossRef]
- Deng, X.; Zhu, T.; Zou, L.; Han, X.; Zhou, X.; Xi, D.; Zhang, D.; Lin, H. Orchestration of hydrogen peroxide and nitric oxide in brassinosteroid-mediated systemic virus resistance in Nicotiana benthamiana. Plant J. 2016, 85, 478–493. [Google Scholar] [CrossRef] [Green Version]
- Kanwar, M.; Yu, J.; Zhou, J. Phytomelatonin: Recent advances and future prospects. J. Pineal Res. 2018, 65, e12526. [Google Scholar] [CrossRef] [Green Version]
- Anuradha, C.; Selvarajan, R.; Vasantha, S.; Suresha, G.S. Biochemical characterization of compatible plant virus interaction: A case study with Bunchy top virus-banana host-pathosystem. Plant Pathol. 2015, 14, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Malamy, J.; Klessig, D.F. Salicylic acid and plant disease resistance. Plant J. 1992, 2, 643–654. [Google Scholar] [CrossRef]
- Helmi, A.; Mohamed, H. Biochemical and ultrastructural changes of some tomato cultivars after infestation with Aphis gossypii Glover (Hemiptera: Aphididae) at Qalyubiyah, Egypt. Gesunde Pflanzen 2016, 68, 41–50. [Google Scholar] [CrossRef]
- Aly, A.A.; Mohamed, H.I.; Mansour, M.T.M.; Omar, M.R. Suppression of powdery mildew on flax by foliar application of essential oils. J. Phytopathol. 2013, 161, 376–381. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of ascorbic acid, glutathione and proline applied as singly or in sequence combination in improving chickpea plant through physiological change and antioxidant defense under different levels of irrigation intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.I.; Elsherbiny, E.A.; Abdelhamid, M.T. Physiological and biochemical responses of Vicia Faba plants to foliar application of zinc and iron. Gesunde Pflanzen 2016, 68, 201–212. [Google Scholar] [CrossRef]
- Akladious, S.A.; Mohamed, H.I. Physiological role of exogenous nitric oxide in improving performance, yield and some biochemical aspects of sunflower plant under zinc stress. Acta Biol. Hung. 2017, 68, 101–114. [Google Scholar] [CrossRef] [Green Version]
- El-Beltagi, H.S.; Sofy, M.R.; Aldaej, M.I.; Mohamed, H.I. Silicon alleviates copper toxicity in flax plants by up-regulating antioxidant defense and secondary metabolites and decreasing oxidative damage. Sustainability 2020, 12, 4732. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I.; Safwat, G.; Megahed, B.M.H.; Gamal, M. Evaluation of some chemical constituents, antioxidant, antibacterial and anticancer activities of Beta vulgaris L. root. Fresenius Environ. Bull. 2018, 27, 6369–6378. [Google Scholar]
- Sofy, A.R.; Mahfouze, S.A.; El-Enany, M.A.M. Isozyme markers for response of wild potato species to Potato spindle tuber viroid Egyptian isolate. World Appl. Sci. J. 2013, 27, 1010–1022. [Google Scholar]
- Gullner, C.; Tóbiás, I.; Fodor, J.; Kömives, U. Elevation of glutathione level and activation of glutathione-related enzymes affect virus infection in tobacco. Free Radic. Res. 2016, 31, 155–161. [Google Scholar] [CrossRef]
- Madhusudhan, K.N.; Srikanta, B.M.; Shylaja, M.D.; Prakash, H.S.; Shetty, H.S. Changes in antioxidant enzymes, hydrogen peroxide, salicylic acid and oxidative stress in compatible and incompatible host-tobamovirus interaction. J. Plant Interact. 2009, 4, 157–166. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhindi, K.M.; Farouk, S.; Alotaibi, M.A. Zinc and paclobutrazol mediated regulation of growth, upregulating antioxidant aptitude and plant productivity of pea plants under salinity. Plants 2020, 9, 1197. [Google Scholar] [CrossRef]
- Abu-Shahba, M.S.; Mansour, M.M.; Mohamed, H.I.; Sofy, M.R. Comparative cultivation and Bbochemical analysis of Iceberg lettuce grown in sand soil and hydroponics with or without microbubbles and macrobubbles. J. Soil Sci. Plant Nutr. 2021, 21, 389–403. [Google Scholar] [CrossRef]
- Megahed, A.A.; El-Dougdoug, K.A.; Othman, B.A.; Lashin, S.M.; Ibrahim, M.A.; Sofy, A.R. A new Egyptian satellite strain of Cucumber mosaic cucumovirus. Int. J. Virol. 2012, 8, 240–257. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, H.I.; Akladious, S.A.; El-Beltagi, H.S. Mitigation the harmful effect of salt stress on physiological, biochemical and anatomical traits by foliar spray with trehalose on wheat cultivars. Fresenius Environ. Bull. 2018, 27, 7054–7065. [Google Scholar]
- Riedle-Bauer, M. Role of reactive oxygen species and antioxidant enzymes in systemic virus infections of plants. J. Phytopathol. 2000, 148, 297–302. [Google Scholar] [CrossRef]
- Clarke, S.F.; Guy, P.L.; Burritt, D.J.; Jameson, P.E. Changes in the activities of antioxidant enzymes in response to virus infection and hormone treatment. Physiol. Plant. 2002, 114, 157–164. [Google Scholar] [CrossRef]
- Yin, L.; Wang, P.; Li, M.; Ke, X.; Li, C.; Liang, D.; Wu, S.; Ma, X.; Li, C.; Zou, Y.; et al. Exogenous melatonin improves Malus resistance to Marssonina apple blotch. J. Pineal Res. 2013, 54, 426–434. [Google Scholar] [CrossRef]
- Sharif, R.; Mujtaba, M.; Ur Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The multifunctional role of chitosan in horticultural crops; a review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chen, Y.; Tan, D.; Reiter, R.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef]
- Lee, H.; Byeon, Y.; Tan, D.; Reiter, R.J.; Back, K. Arabidopsis serotonin N-acetyltransferase knockout mutant plants exhibit decreased melatonin and salicylic acid levels resulting in susceptibility to an avirulent pathogen. J. Pineal Res. 2015, 58, 291–299. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Jia, W.; Li, B.; Li, S.; Liang, Y.; Wu, X.; Ma, M.; Wang, J.; Gao, J.; Cai, Y.; Zhang, Y. Mitogen-activated protein kinase cascade MKK7-MPK6 plays important roles in plant development and regulates shoot branching by phosphorylating PIN1 in Arabidopsis. PLoS Biol. 2016, 14, e1002550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahesh, H.M.; Murali, M.; Anup Chandra Pal, M.; Melvin, P.; Sharada, M.S. Salicylic acid seed priming instigates defense mechanism by inducing PR-Proteins in Solanum melongena L. upon infection with Verticillium dahliae Kleb. Plant Physiol. Biochem. 2017, 117, 12–23. [Google Scholar] [CrossRef]
- Hackmann, C.; Korneli, C.; Kutyniok, M.; Köster, T.; Wiedenlübbert, M.; Müller, C.; Staiger, D. Salicylic acid-dependent and -independent impact of an RNA-binding protein on plant immunity. Plant Cell Environ. 2014, 37, 696–706. [Google Scholar] [CrossRef]
- Clark, M.F.; Adams, A.N. Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection of plant viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Noordam, D. Identification of Plant Viruses. In Methods and Experiments; Center for Agricultural Publishing and Documentation: Wageningen, The Netherlands, 1973; p. 207. [Google Scholar]
- Yang, X.; Kang, L.; Tien, P.O. Resistance of tomato infected with Cucumber mosaic virus satellite RNA to Potato spindle tuber viroid. Ann. Appl. Biol. 1996, 129, 543–551. [Google Scholar] [CrossRef]
- Vernon, L.P.; Seely, G.R. The Chlorophylls; Academic Press: Cambridge, MA, USA, 1966. [Google Scholar]
- Lichtenthaler, H.K. Chlorophylls and Carotenoids: Pigments of Photosynthetic Biomembranes. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Jabs, T.; Dietrich, R.A.; Dang, J.L. Initiation of runaway cell death in an Arabidopsis mutant by extracellular superoxide. Science 1996, 273, 1853–1856. [Google Scholar] [CrossRef]
- Babbs, C.F.; Pham, J.A.; Coolbaugh, R.C. Lethal hydroxyl radical production in paraquat-treated plants. Plant Physiol. 1989, 90, 1267–1270. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Bruce, R.J.; West, C.A. Elicitation of lignin biosynthesis and isoperoxidase activity by pectic fragments in suspension cultures of castor bean. Plant Physiol. 1989, 91, 889–897. [Google Scholar] [CrossRef] [PubMed]
- Raskin, I.; Turner, I.M.; Melander, W.R. Regulation of heat-production in the inflorescences of an arum lily by endogenous salicyclic acid. Proc. Natl. Acad. Sci. USA 1989, 86, 2214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kono, Y. Generation of superoxide radical during autoxidation of hydroxylamine and an assay for superoxide dismutase. Arch. Biochem. Biophys. 1978, 186, 189–195. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1984; Volume 105, pp. 121–126. [Google Scholar]
- Thomas, R.L.; Jen, J.J.; Morr, C.V. Changes in soluble and bound peroxidase—IAA oxidase during tomato fruit development. J. Food Sci. 1982, 47, 158–161. [Google Scholar] [CrossRef]
- Assis, J.S.; Maldonado, R.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Tang, C.; Zhang, G.; Li, Y.; Wang, C.; Liu, B.; Qu, Z.; Zhao, J.; Han, Q.; Huang, L.; et al. cDNA-AFLP analysis reveals differential gene expression in compatible interaction of wheat challenged with Puccinia striiformis f. sp. tritici. BMC Genom. 2009, 10, 289. [Google Scholar] [CrossRef] [Green Version]
- Derbalah, A.S.; Elsharkawy, M.M. A new strategy to control Cucumber mosaic virus using fabricated NiO-nanostructures. J. Biotechnol. 2019, 306, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zimei, L.; Cui, J.; Jiangang, D.; Xia, X.; Liu, D.; Yu, J. Alleviation of chilling-induced oxidative damage by salicylic acid pretreatment and related gene expression in eggplant seedlings. Plant Growth Regul. 2011, 65, 101–108. [Google Scholar] [CrossRef]
- Gomez, K.A.; Gomez, A.A. Statistical Procedures for Agricultural Research; John Wiley & Sons Inc.: Singapore, 1984. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession No. | Origin | Year | Host | Strains | Identity | |
|---|---|---|---|---|---|---|
| nt (666) | aa (221) | |||||
| LN846978 | Egypt | 2015 | Tomato | shark-EG | 644/666 (96.70%) | 211/221 (95.48%) |
| HQ288892 | 2010 | S. tuberosum | Potato-EG | 644/666 (96.70%) | 210/221 (95.02%) | |
| LN846979 | 2015 | Tomato | mans-EG | 643/666 (96.55%) | 210/221 (95.02%) | |
| KY471416 | 2014 | S. tuberosum | CP2-EG | 638/668 (95.51%) | 204/221 (92.31%) | |
| KY549685 | 2014 | - | FRE-EG | 638/668 (95.51%) | 204/221 (92.31%) | |
| MH625710 | 2016 | Basil | Basil-EG | 619/666 (92.94%) | 198/221 (89.59%) | |
| HQ185569 | USA | 2006 | Soybean | Joe Davis | 643/666 (96.55%) | 210/221 (95.02%) |
| M59241 | 1991 | Lucerne | YSMV | 638/666 (95.80%) | 209/221 (94.57%) | |
| K02703 | 1983 | N.tabacum | 425M | 639/666 (95.95%) | 208/221 (94.12%) | |
| JN256026 | 2011 | Soybean | SE12 | 639/666 (95.95%) | 208/221 (94.12%) | |
| L00162 | 1977 | N. glutinosa | 425L | 635/666 (95.35%) | 205/221 (92.76%) | |
| AJ130709 | France | 1998 | Wild tomato | Lyh1 | 634/666 (95.20%) | 204/221 (92.31%) |
| AJ130708 | 1998 | Carrot | Dac16 | 634/666 (95.20%) | 204/221 (92.31%) | |
| AJ130707 | 1998 | Pepper | Caa1 | 636/666 (95.50%) | 203/221 (91.86%) | |
| AJ130703 | 1998 | Tomato | Lye80 | 629/666 (94.44%) | 201/221 (90.95%) | |
| Y09110 | Italy | 1997 | Tomato | Danza | 641/666 (96.25%) | 209/221 (94.57%) |
| AJ130706 | 1998 | Bean | F430 | 639/666 (95.95%) | 209/221 (94.57%) | |
| AF015716 | England | 1997 | Garden lupin | VRU | 634/666 (95.20%) | 205/221 (92.76%) |
| AF015717 | 1997 | Garden lupin | 15/64 | 628/666 (94.29%) | 204/221 (92.31%) | |
| DQ314753 | Canada | 2004 | Potato | Ca401 | 637/661 (96.37%) | 209/220 (95.00%) |
| DQ314751 | 2004 | Potato | Ca399 | 642/667 (96.25%) | 209/221 (94.57%) | |
| DQ314749 | 2004 | Potato | Ca375 | 640/667 (95.95%) | 209/221 (94.57%) | |
| DQ314752 | 2004 | Potato | Ca400 | 640/667 (95.80%) | 209/221 (94.57%) | |
| DQ314750 | 2004 | Potato | Ca175 | 639/667 (95.80%) | 208/221 (94.12%) | |
| DQ314754 | 2004 | Potato | Ca508 | 636/667 (95.35%) | 207/221 (93.67%) | |
| JX112759 | Australia | 2001 | Medicago sativa | Hu | 641/666 (96.25%) | 209/221 (94.57%) |
| HM807304 | 1985 | Medicago sativa | N20 | 638/666 (95.80%) | 209/221 (94.57%) | |
| AF294433 | Korea | 2000 | Solanum tuberosum | KR2 | 641/666 (96.25%) | 211/221 (95.48%) |
| AF294432 | 2000 | Solanum tuberosum | KR1 | 638/666 (95.80%) | 208/221 (94.12%) | |
| AF215664 | New Zealand | 1999 | S. tuberosum | NZ34 | 640/666 (96.10%) | 208/221 (94.12%) |
| Treatments | Virus Concentration | Percentage of Infection (%) | Disease Severity (%) |
|---|---|---|---|
| Challenge control (ChC) | 0.50 a | 100 a | 90.0 a |
| Melatonin + V | 0.18 c | 25 c | 13.33 c |
| Salicylic acid + V | 0.22 b | 40 b | 21.66 b |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| GR Glutathione reductase | TTGGTGGAACGTGTGTTCTT | TCTCATTCACTTCCCATCCA |
| DHAR Dehydroascorbate reductase | GAAGTGGAGTGTGCCTGAAA | CGTACTTCTCTTCAGCCTTGG |
| MDHAR Monodehydroascorbate reductase | TCCGAACAAACATACCTGGA | GTGTGCGTGTGTGCAGTTAG |
| PR3 Chitinase expression | AGAGAACAAGGTAGCCCAGG | TAAAAGGTCCACTCCGATGGC |
| MPK1 Mitogen-activated protein kinase 1 | CCTCCGTGGGTTGAAATAC | GTCACAACATATTCGGTCATAAAG |
| ACTIN | TGGTCGGAATGGGACAGAAG | CTCAGTCAGGAGAACAGGGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Refaey, E.E.; Mohamed, H.I.; El-Dougdoug, N.K. Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid. Plants 2021, 10, 459. https://doi.org/10.3390/plants10030459
Sofy AR, Sofy MR, Hmed AA, Dawoud RA, Refaey EE, Mohamed HI, El-Dougdoug NK. Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid. Plants. 2021; 10(3):459. https://doi.org/10.3390/plants10030459
Chicago/Turabian StyleSofy, Ahmed R., Mahmoud R. Sofy, Ahmed A. Hmed, Rehab A. Dawoud, Ehab E. Refaey, Heba I. Mohamed, and Noha K. El-Dougdoug. 2021. "Molecular Characterization of the Alfalfa mosaic virus Infecting Solanum melongena in Egypt and the Control of Its Deleterious Effects with Melatonin and Salicylic Acid" Plants 10, no. 3: 459. https://doi.org/10.3390/plants10030459