Antioxidant, Anti-tyrosinase, Anti-α-amylase, and Cytotoxic Potentials of the Invasive Weed Andropogon virginicus

 ,
,  ,
,  , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Yield and Total Phenolic (TPC) and Flavonoid (TFC) Contents of Extracts from A. virginicus
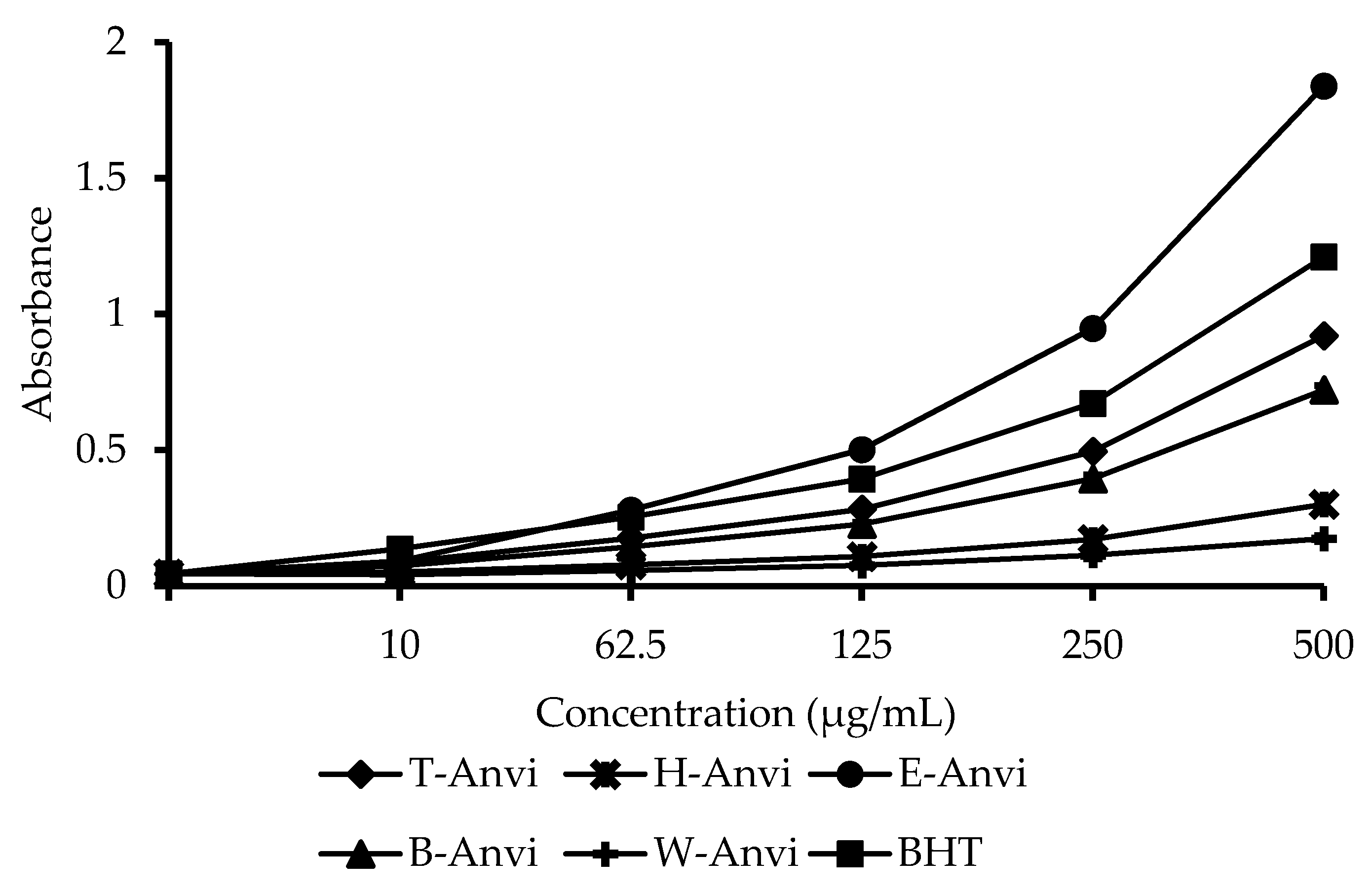
2.2. Antioxidant Activity of Extracts from A. virginicus
2.3. Tyrosinase and α-Amylase Inhibitory Activities of A. virginicus Extracts
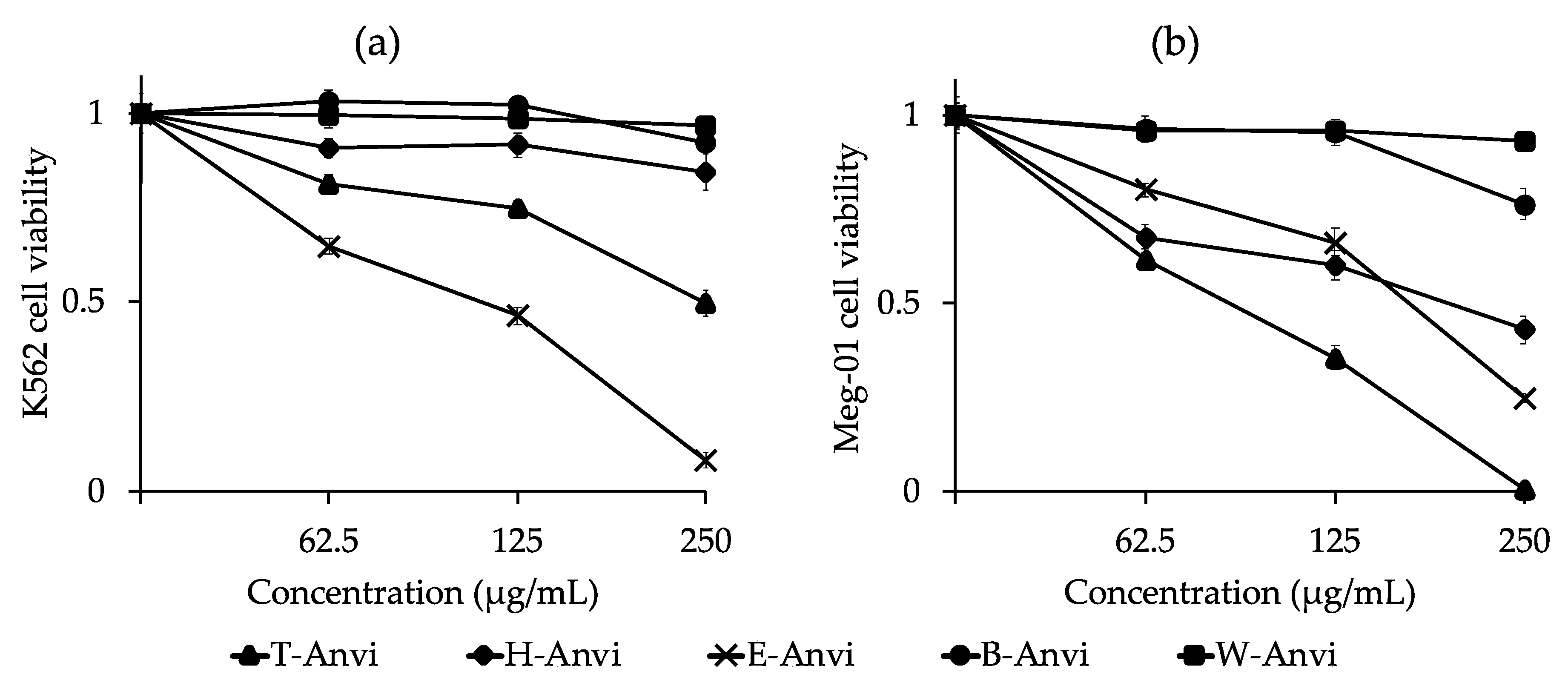
2.4. Cytotoxic Activity of Extracts from A. virginicus against K562 and Meg-01 Cell Lines
2.5. Correlations between Total Phenolic (TPC) and Flavonoid (TFC) Contents and Biological Properties of A. virginicus Extracts
2.6. GC-MS Results
2.7. HPLC-ESI-MS/MS Results
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Chemicals and Cell Lines
4.1.2. Plant Materials
4.2. Sample Preparation and Extraction
4.3. Total Phenolic (TPC) and Flavonoid (TFC) Contents
4.4. Antioxidant Activity
4.4.1. ABTS Cation Decolorization Assay
4.4.2. DPPH Radical Scavenging Assay
4.4.3. β-Carotene Bleaching Assay
4.4.4. Reducing Power Assay
4.5. Enzymatic Inhibitory Assays
4.5.1. Tyrosinase Inhibition
4.5.2. α-Amylase Inhibition
4.6. Cytotoxic Activity against K562 and Meg-01 Cell Lines
4.7. Identification of Phytochemical Component
4.7.1. GC-MS
- (1)
- Initial temperature: 50 °C (without holding),
- (2)
- Rushing temperature: 10 °C/min for 30 min and 20 min in maintenance,
- (3)
- Injector and detector temperatures: 300 °C and 320 °C, respectively.
4.7.2. HPLC-ESI-MS/MS
4.8. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Campbell, C. Andropogon. In Flora of North America North of Mexico, Volume 25: Magnoliophyta: Commelinidae (in Part): Poaceae, Part 2; Flora of North America Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2003; Volume 25, p. 814. [Google Scholar]
- European and Mediterranean Plant Protection Organization. Andropogon virginicus L. Bull. OEPP/EPPO Bull. 2019, 49, 61–66. [Google Scholar] [CrossRef] [Green Version]
- U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). Available online: https://www.fs.fed.us/database/feis/plants/graminoid/andvir/all.html (accessed on 20 October 2020).
- CABI—Invasive Species Compendium. Available online: http://www.cabi.org/isc/datasheet/5286 (accessed on 14 October 2020).
- US Fish and Wildlife Service, Conserving the Nature of America. Available online: https://ecos.fws.gov/speciesProfile/profile/speciesProfile.action?spcode=Q38V (accessed on 20 October 2020).
- Weeds of Australia—Biosecurity Queensland Edition Fact Sheet, Andropogon virginicus. Available online: https://keyserver.lucidcentral.org/weeds/data/media/Html/andropogon_virginicus.pdf (accessed on 20 October 2020).
- Balandier, P.; Collet, C.; Miller, J.H.; Reynolds, P.E.; Zedaker, S.M. Designing forest vegetation management strategies based on the mechanisms and dynamics of crop tree competition by neighbouring vegetation. Forestry 2006, 79, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Collins, T. Acute and chronic inflammation. In Pathologic Basis of Disease; Cotran, R.S., Kumar, V., Collins, T., Eds.; WB Saunders: Philadelphia, PA, USA, 1999; pp. 50–88. [Google Scholar]
- Anderson, M.T.; Staal, F.J.T.; Gitler, C.; Herzenberg, L.A.; Herzenberg, L.A. Separation of oxidant initiated and redox-regulated steps in the NF-κB signal transduction pathway. Proc. Natl. Acad. Sci. USA 1994, 91, 11527–11531. [Google Scholar] [CrossRef] [Green Version]
- Flohe, L.; Brigelius-Flohe, R.; Saliou, C.; Traber, M.G.; Packer, L. Redox regulation of NF-κB activation. Free Radic. Biol. Med. 1997, 22, 1115–1126. [Google Scholar] [CrossRef]
- Biswas, S.K.; de Faria, J.B.L. Which comes first: Renal inflammation or oxidative stress in spontaneously hypertensive rats? Free Radic. Res. 2007, 41, 216–224. [Google Scholar] [CrossRef]
- Ambade, A.; Mandrekar, P. Oxidative stress and inflammation: Essential partners in alcoholic liver disease. Int. J. Hepatol. 2012, 2012, 853175. [Google Scholar] [CrossRef]
- Cachofeiro, V.; Goicochea, M.; de Vinuesa, S.D.; Oubina, P.; Lahera, V.; Luno, J. Oxidative stress and inflammation, a link between chronic kidney disease and cardiovascular disease. Kidney Int. Suppl. 2008, 111, S4–S9. [Google Scholar] [CrossRef] [Green Version]
- Tucker, P.S.; Scanlan, A.T.; Dalbo, V.J. Chronic kidney disease influences multiple systems: Describing the relationship between oxidative stress, inflammation, kidney damage, and concomitant disease. Oxid. Med. Cell. Longev. 2015, 2015, 806358. [Google Scholar] [CrossRef]
- Sies, H. Introductory remarks. In Oxidative Stress; Sies, H., Ed.; Academic Press: Orlando, FL, USA, 1985; pp. 1–7. [Google Scholar]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 3rd ed.; Oxford University Press: London, UK, 1999. [Google Scholar]
- Biswas, S.K.; de Faria, J.B.L. Hypertension induces oxidative stress but not macrophage infiltration in the kidney in the early stage of experimental diabetes mellitus. Am. J. Nephrol. 2006, 26, 415–422. [Google Scholar] [CrossRef]
- Biswas, S.K.; Peixoto, E.B.; Souza, D.S.; Lopes de Faria, J.B. Hypertension increases pro-oxidant generation and decreases antioxidant defense in the kidney in early diabetes. Am. J. Nephrol. 2008, 28, 133–142. [Google Scholar] [CrossRef]
- Halliwell. Oxidative stress and neurodegeneration: Where are we now? J. Neurochem. 2006, 97, 1634–1658. [Google Scholar] [CrossRef]
- Onyango, I.G. Mitochondrial dysfunction and oxidative stress in Parkinson’s disease. Neurochem. Res. 2008, 33, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Hald, A.; Van Beek, J.; Lotharius, J. Inflammation in Parkinson’s disease: Causative or epiphenomenal? Subcell. Biochem. 2007, 42, 249–279. [Google Scholar] [PubMed]
- Tarling, C.A.; Woods, K.; Zhang, R.; Brastianos, H.C.; Brayer, G.D.; Andersen, R.J.; Withers, S.G. The search for novel human pancreatic α-amylase inhibitors: High-throughput screening of terrestrial and marine natural product extracts. ChemBioChem 2008, 9, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Linda, G.H. Dermatological complications of obesity. Am. J. Clin. Dermatol. 2002, 3, 497–506. [Google Scholar] [CrossRef]
- Demirseren, D.D.; Emre, S.; Akoglu, G.; Arpacı, D.; Arman, A.; Metin, A.; Cakır, B. Relationship between skin diseases and extracutaneous complications of diabetes mellitus: Clinical analysis of 750 patients. Am. J. Clin. Dermatol. 2014, 15, 65–70. [Google Scholar] [CrossRef]
- Klein, E.; Ben-Bassat, H.; Neumann, H.; Ralph, P.; Zeuthen, J.; Polliack, A.; Vanky, F. Properties of the K562 cell line, derived from a patient with chronnic myeloid leukemia. Int. J. Cancer 1976, 18, 421–431. [Google Scholar] [CrossRef]
- NIH—National Cancer Institute. Available online: https://www.cancer.gov/publications/dictionaries/cancer-terms/def/white-blood-cell?fbclid=IwAR1Jr1RfMklHWtlLj2eQ_HdJp9xY6-h8OQHhYkg2fnQWBeDLJbzscm9tLO8 (accessed on 20 October 2020).
- Koeffler, H.P.; Golde, D.W. Human myeloid leukemia cell lines: A review. Blood 1980, 56, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Ogura, M.; Saito, H.; Satoh, M.; Takeuchi, M. Production of platelet-like particles by a human megakaryoblastic leukemia cell line (Meg-01). Exp. Cell Res. 1991, 193, 223–226. [Google Scholar] [CrossRef]
- Ogura, M.; Morishima, Y.; Ohno, R.; Kato, Y.; Hirabayashi, N.; Nagura, H.; Saito, H. Establishment of a novel human megakaryoblastic leukemia cell line, MEG-01, with positive Philadelphia chromosome. Blood 1985, 66, 1384–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjelakovic, G.; Nikolova, D.; Gluud, L.L.; Simonetti, R.G.; Gluud, C. Mortality in randomized trials of antioxidant supplements for primary and secondary prevention: Systematic review and meta-analysis. JAMA 2007, 297, 842–857. [Google Scholar] [CrossRef]
- Zhang, S.M.; Cook, N.R.; Albert, C.M.; Gaziano, J.M.; Buring, J.E.; Manson, J.E. Effect of combined folic acid, vitamin B6, and vitamin B12 on cancer risk in women: A randomized trial. JAMA 2008, 300, 2012–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sesso, H.D.; Buring, J.E.; Christen, W.G.; Kurth, T.; Belanger, C.; MacFadyen, J.; Bubes, V.; Manson, J.E.; Glynn, R.J.; Gaziano, J.M. Vitamins E and C in the prevention of cardiovascular disease in men: The physicians’ health study II randomized controlled trial. JAMA 2008, 300, 2123–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.K. Does the interdependence between oxidative stress and inflammation explain the antioxidant paradox? Oxid. Med. Cell. Longev. 2016, 2016, 5698931. [Google Scholar] [CrossRef] [Green Version]
- Thavamoney, N.; Sivanadian, L.; Tee, L.H.; Khoo, H.E.; Prasad, K.N.; Kong, K.W. Extraction and recovery of phytochemical components and antioxidative properties in fruit parts of Dacryodes rostrata influenced by different solvents. J. Food Sci. Technol. 2018, 55, 2523–2532. [Google Scholar] [CrossRef]
- Tagousop, C.N.; Tamokou, J.-D.-D.; Ekom, S.E.; Ngnokam, D.; Voutquenne-Nazabadioko, L. Antimicrobial activities of flavonoid glycosides from Graptophyllum grandulosum and their mechanism of antibacterial action. BMC Complement. Altern. Med. 2018, 18, 252. [Google Scholar] [CrossRef]
- Kefayati, Z.; Motamed, S.M.; Shojaii, A.; Noori, M.; Ghods, R. Antioxidant activity and phenolic and flavonoid contents of the extract and subfractions of Euphorbia splendida Mobayen. Pharmacogn. Res. 2017, 9, 362–365. [Google Scholar] [CrossRef]
- Al-Saeedi, A.H.; Al-Ghafri, M.T.H.; Hossain, M.A. Comparative evaluation of total phenols, flavonoids content and antioxidant potential of leaf and fruit extracts of Omani Ziziphus jujuba L. Pac. Sci. Rev. A Nat. Sci. Eng. 2016, 18, 78–83. [Google Scholar] [CrossRef] [Green Version]
- Soobrattee, M.A.; Neergheen, V.S.; Luximon-Ramma, A.; Aruoma, O.I.; Bahorun, T. Phenolics as potential antioxidant therapeutic agents: Mechanism and actions. Mutat. Res. Fundam. Mol. Mech. Mutagen. 2005, 579, 200–213. [Google Scholar] [CrossRef]
- Kelly, E.H.; Anthony, R.T.; Dennis, J.B. Flavonoid antioxidants: Chemistry, metabolism and structure-activity relationships. J. Nutr. Biochem. 2002, 13, 572–584. [Google Scholar] [CrossRef]
- Pandey, A.K.; Mishra, A.K.; Mishra, A. Antifungal and antioxidative potential of oil and extracts derived from leaves of Indian spice plant Cinnamomum tamala. Cell. Mol. Biol. 2012, 58, 142–147. [Google Scholar]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Şöhretoğlua, D.; Sarib, S.; Barutc, B.; Özelc, A. Tyrosinase inhibition by some flavonoids: Inhibitory activity, mechanism by T in vitro and in silico studies. Bioorg. Chem. 2018, 81, 168–174. [Google Scholar] [CrossRef]
- Quan, N.V.; Xuan, T.D.; Tran, H.D.; Thuy, N.T.D.; Trang, L.T.; Huong, C.T.; Yusuf, A.; Tuyen, P.T. Antioxidant, α-amylase and α-glucosidase inhibitory activities and potential constituents of Canarium tramdenum bark. Molecules 2019, 24, 605. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Hsu, C.; Huang, L. Inhibitory potential of fatty acids on key enzymes related to type 2 diabetes. BioFactors 2013, 39, 415–421. [Google Scholar] [CrossRef]
- Su, C.H.; Lai, M.N.; Ng, L.T. Inhibitory effects of medicinal mushrooms on α-amylase and α-glucosidase enzymes related to hyperglycemia. Food Funct. 2013, 4, 644–649. [Google Scholar] [CrossRef]
- Anh, L.H.; Xuan, T.D.; Thuy, N.T.D.; Quan, N.V.; Trang, L.T. Antioxidant and α-amylase inhibitory activities and phytocompounds of Clausena indica Fruits. Medicines 2020, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Kumar, V.; Prakash, O. Enzymes inhibition and antidiabetic effect of isolated constituents from Dillenia indica. Biomed Res. Int. 2013, 2013, 382063. [Google Scholar] [CrossRef]
- Chahar, M.K.; Sharma, N.; Dobhal, M.P.; Joshi, Y.C. Flavonoids: A versatile source of anticancer drugs. Pharmacogn. Rev. 2011, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veeramuthu, D.; Raja, W.R.T.; Al-Dhabi, N.A.; Savarimuthu, I. Flavonoids: Anticancer properties. In Flavonoids—From Biosynthesis to Human Health; Justino, G.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Sakthivel, R.; Sheeja, D.; Kasi, M.; Devi, P. Phytol shows anti-angiogenic activity and induces apoptosis in A549 cells bydepolarizing the mitochondrial membrane potential. Biomed. Pharmacother. 2018, 105, 742–752. [Google Scholar] [CrossRef] [PubMed]
- Alencar, M.V.O.B.; Islam, M.T.; Ali, E.S.; Santos, J.V.O.; Paz, M.F.C.J.; Sousa, J.M.C.; Dantas, S.M.M.M.; Mishra, S.K.; Cavalcante, A.A.C.M. Association of phytol with toxic and cytotoxic activities in an antitumoral perspective: A meta-analysis and systemic review. Anticancer Agents Med. Chem. 2018, 18, 1828–1837. [Google Scholar] [CrossRef]
- Pejin, B.; Kojic, V.; Bogdanovic, G. An insight into the cytotoxic activity of phytol at in vitro conditions. Nat. Prod. Res. 2014, 28, 2053–2056. [Google Scholar] [CrossRef]
- Sheeja, L.; Lakshmi, D.; Bharadwaj, S.; Parveen, K.S. Anticancer activity of phytol purified from Gracilaria edulis against human breast cancer cell line (MCF-7). Int. J. Curr. Sci. 2016, 19, 36–46. [Google Scholar]
- de Alencar, M.V.O.B.; Islam, M.T.; de Lima, R.M.T.; Paz, M.F.C.J.; Dos Reis, A.C.; da Mata, A.M.O.F.; Filho, J.W.G.O.; Cerqueira, G.S.; Ferreira, P.M.P.; Sousa, E.J.M.C.; et al. Phytol as an anticarcinogenic and antitumoral agent: An in vivo study in swiss mice with DMBA-Induced breast cancer. IUBMB Life 2019, 71, 200–212. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.W.; Cho, S.K. Phytol induces apoptosis and ROS-mediated protective autophagy in human gastric adenocarcinoma AGS cells. Anal. Biochem. 2015, 4, 4. [Google Scholar] [CrossRef]
- Endrini, S.; Rahmat, A.; Ismail, P.; Taufiq-Yap, Y.H. Cytotoxic effect of γ-sitosterol from Kejibeling (Strobilanthes crispus) and its mechanism of action towards c-myc gene expression and apoptotic pathway. Med. J. Indones. 2014, 23, 203. [Google Scholar] [CrossRef] [Green Version]
- Sundarraj, S.; Thangam, R.; Sreevani, V.; Kaveri, K.; Gunasekaran, P.; Achiraman, S.; Kannan, S. γ-Sitosterol from Acacia nilotica L. induces G2/M cell cycle arrest and apoptosis through c-Myc suppression in MCF-7 and A549 cells. J. Ethnopharmacol. 2012, 141, 803–809. [Google Scholar] [CrossRef]
- Teschke, R.; Xuan, T.D. Active nature-based ingredients for drug discovery with pivotal role of clinical efficacy: Review and prospective. J. Mod. Med. Chem. 2020, 8, 4–18. [Google Scholar] [CrossRef]
- Teschke, R.; Larrey, D.; Melchart, D.; Danan, G. Traditional Chinese medicine (TCM) and herbal hepatotoxicity: RUCAM and the role of novel diagnostic biomarkers such as microRNAs. Medicines 2016, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teschke, T.; Eickhoff, A.; Wolff, A.; Xuan, T.D. Liver injury from herbs and “dietary supplements”: Highlights of a literature review from 2015 to 2017. Curr. Pharmacol. Rep. 2018, 4, 120–131. [Google Scholar] [CrossRef]
- Teschke, T.; Xuan, T.D. Herbs including shell ginger, antioxidant profiles, aging, and longevity in Okinawa, Japan: A critical analysis of current concepts. In Aging, 2nd ed.; Preedy, V.R., Patel, V.B., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 209–222. [Google Scholar] [CrossRef]
- Teschke, R.; Xuan, T.D. Viewpoint: A contributory role of shell ginger (Alpinia zerumbet (Pers.) B.L. Burtt & R.M. Sm) for human longevity in Okinawa, Japan? Nutrients 2018, 10, 166. [Google Scholar] [CrossRef] [Green Version]
- Tropicos—Missouri Botanical Garden. Available online: https://www.tropicos.org/name/25509277 (accessed on 20 October 2020).
- National Institute for Environmental Studies. Available online: https://www.nies.go.jp/biodiversity/invasive/DB/detail/81450e.html (accessed on 20 October 2020).
- Invasive Weed; Japan Livestock Technology Association (JLTA): Tokyo, Japan, 1994; p. 31.
- Elzaawely, A.A.; Xuan, T.D.; Koyama, H.; Tawata, S. Antioxidant activity and contents of essential oil and phenolic compounds in flowers and seeds of Alpinia zerumbet (Pers.) B.L. Burtt. & R.M. Sm. Food Chem. 2007, 104, 1648–1653. [Google Scholar] [CrossRef]
- Tuyen, P.T.; Xuan, T.D.; Khang, D.T.; Ahmad, A.; Quan, N.V.; Anh, T.T.T.; Anh, L.H.; Minh, T.N. Phenolic compositions and antioxidant properties in bark, flower, inner skin, kernel and leaf extracts of Castanea crenata Sieb. et Zucc. Antioxidants 2017, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Pellegrini, N.; Serafini, M.; Colombi, B.; Del Rio, D.; Salvatore, S.; Bianchi, M.; Brighenti, F. Total antioxidant capacity of plant foods, beverages and oils consumed in Italy assessed by three different in vitro assays. J. Nutr. 2003, 133, 2812–2819. [Google Scholar] [CrossRef] [Green Version]
- Elzaawely, A.A.; Tawata, S. Antioxidant activity of phenolic rich fraction obtained from Convolvulus arvensis L. leaves grown in Egypt. Asian J. Crop. Sci. 2012, 4, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Quan, N.V.; Xuan, T.D.; Anh, L.H.; Tran, H.-D. Bio-guided isolation of prospective bioactive constituents from roots of Clausena indica (Dalzell) Oliv. Molecules 2019, 24, 4442. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Samples | Code | Extraction Yield (%) | TPC (mg GAE/g DW) | TFC (mg RE/g DW) |
|---|---|---|---|---|
| Total crude extract | T-Anvi | 9.8 | 24.80 ± 0.51 a | 37.40 ± 0.74 a |
| Hexane extract | H-Anvi | 1.6 | 1.26 ± 0.03 c | 6.30 ± 0.13 c |
| Ethyl acetate extract | E-Anvi | 3.2 | 25.34 ± 0.47 a | 25.44 ± 0.45 b |
| Butanol extract | B-Anvi | 1.2 | 3.26 ± 0.06 b | 4.91 ± 0.08 d |
| Water extract | W-Anvi | 1.1 | 0.49 ± 0.01 c | 0.25 ± 0.01 e |
| Samples | ABTS Assay IC50 (µg/mL) | DPPH Assay IC50 (µg/mL) |
|---|---|---|
| T-Anvi | 77.71 ± 1.85 d | 30.54 ± 0.40 cd |
| H-Anvi | 323.88 ± 1.22 b | 126.27 ± 4.92 b |
| E-Anvi | 43.59 ± 1.25 f | 13.96 ± 0.39 e |
| B-Anvi | 86.73 ± 2.30 c | 33.79 ± 0.24 c |
| W-Anvi | 586.31 ± 6.11 a | 386.91 ± 8.24 a |
| BHT | 63.51 ± 0.43 e | 20.81 ± 0.67 de |
| Samples | Tyrosinase Inhibition IC50 (mg/mL) | α-Amylase Inhibition IC50 (mg/mL) |
|---|---|---|
| T-Anvi | 4.57 ± 0.05 c | 3.48 ± 0.07 a |
| H-Anvi | 6.22 ± 0.08 b | 0.72 ± 0.01 c |
| E-Anvi | 2.58 ± 0.13 d | ne |
| B-Anvi | 9.40 ± 0.02 a | ne |
| W-Anvi | na | na |
| Kojic acid | 0.02 ± 0.00 e | - |
| Palmitic acid | - | 1.57 ± 0.04 b |
| TPC | TFC | ABTS | DPPH | β-Ca | RP | Tyro | α-Amy | K562 | |
|---|---|---|---|---|---|---|---|---|---|
| TFC | 0.952 * | - | - | - | - | - | - | - | - |
| ABTS | 0.833 * | 0.674 * | - | - | - | - | - | - | - |
| DPPH | 0.813 * | 0.633 * | 0.993 * | - | - | - | - | - | - |
| β-Ca | 0.423 | 0.466 | 0.589 * | 0.549 * | - | - | - | - | - |
| RP | 0.843 * | 0.667 * | 0.987 * | 0.997 * | 0.523 * | - | - | - | - |
| Tyro | 0.479 | 0.525 * | 0.601 * | 0.572 * | 0.970 * | 0.555 * | - | - | - |
| α-Amy | −0.236 | −0.043 | −0.439 | −0.416 | 0.217 | −0.391 | 0.302 | - | - |
| K562 | 0.898 * | 0.736 * | 0.891 * | 0.914 * | 0.330 | 0.942 * | 0.393 | −0.307 | - |
| Meg-01 | 0.787 * | 0.913 * | 0.404 | 0.378 | 0.435 | 0.424 | 0.526 * | 0.351 | 0.554 * |
| No. | Identified Compound | RT (min) | MW | Formula | Classification | Peak Area (%) | LRI | KI | Content (mg/g DW) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Phytol | 15.80 | 296 | C20H40O | Diterpenoids | 16.42 | 1835 | 1835 | - |
| 2 | 8-Methyl-1-undecene | 16.64 | 168 | C12H24 | Alkenes | 10.77 | 1916 | 1917 | - |
| 3 | Palmitic acid | 17.02 | 256 | C16H32O2 | Fatty acids | 27.97 | 1955 | 1955 | 0.86 |
| 4 | γ-Sitosterol | 28.34 | 414 | C29H50O | Steroids | 7.38 | - | - | - |
| No. Signals | RT (min) | [M+H]− (m/z) | Chemical Classification | Tentative Identity | Molecular Formula | Exact Mass | Fragmentions (m/z) |
|---|---|---|---|---|---|---|---|
| 1 | 0.68 | 377.086 | Benzophenones | Annulatophenonoside | C18H18O9 | 378.3 | 89.024(100); 119.086(53); 143.036(45); 149.099(28) |
| 2 | 5.74 | 371.098 | Phenolic glycosides | Dihydroferulic acid 4-O-glucuronide | C16H20O10 | 372.3 | 231.158(100); 243.293(23); 225.296(19); 121.105(14) |
| 3 | 6.08 | 447.093 | Flavonoids | Kaempferol-O-galactopyranoside | C21H20O11 | 448.4 | 327.154(100); 357.141(48); 429.155(5); 369.242(4) |
| 4 | 6.35 | 431.098 | Flavonoids | Genistin | C21H20O10 | 432.4 | 311.140(100); 341.117(6); 283.142(2) |
| 5 | 6.54 | 463.088 | Flavonoids | Quercetin-3-O-β-d-glucopyranoside | C21H20O12 | 464.4 | 445.145(100); 343.148(92); 427.169(75); 373.161(48) |
| 6 | 6.67 | 447.093 | Flavonoids | Kaempferol 3-O-β-d-glucopyranoside | C21H20O11 | 448.4 | 285.167(100); 402.158(6); 428.144(5); 374.128(3) |
| 7 | 8.37 | 413.088 | Proanthocyanidins | Prodelphinidin B6 | C21H18O9 | 414.4 | 313.135(100); 297.155(73); 369.145(70); 285.154(20) |
| 8 | 9.71 | 343.082 | Flavonoids | Eupatilin | C18H16O7 | 344.3 | 311.153(100) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anh, L.H.; Quan, N.V.; Lam, V.Q.; Iuchi, Y.; Takami, A.; Teschke, R.; Xuan, T.D. Antioxidant, Anti-tyrosinase, Anti-α-amylase, and Cytotoxic Potentials of the Invasive Weed Andropogon virginicus. Plants 2021, 10, 69. https://doi.org/10.3390/plants10010069
Anh LH, Quan NV, Lam VQ, Iuchi Y, Takami A, Teschke R, Xuan TD. Antioxidant, Anti-tyrosinase, Anti-α-amylase, and Cytotoxic Potentials of the Invasive Weed Andropogon virginicus. Plants. 2021; 10(1):69. https://doi.org/10.3390/plants10010069
Chicago/Turabian StyleAnh, La Hoang, Nguyen Van Quan, Vu Quang Lam, Yu Iuchi, Akiyoshi Takami, Rolf Teschke, and Tran Dang Xuan. 2021. "Antioxidant, Anti-tyrosinase, Anti-α-amylase, and Cytotoxic Potentials of the Invasive Weed Andropogon virginicus" Plants 10, no. 1: 69. https://doi.org/10.3390/plants10010069