
Artepillin C Reduces Allergic Airway Inflammation by Induction of Monocytic Myeloid-Derived Suppressor Cells

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Artepillin C (ArtC) Isolation
2.2. Animals
2.3. OVA-Induced Asthma Model
2.4. Treatment
2.5. Bronchoalveolar Lavage
2.6. Lung Homogenate
2.7. Histopathology
2.8. MDSC Differentiation
2.9. Treg Cell Differentiation
2.10. Flow Cytometry
2.11. M-MDSC Adoptive Transfer
2.12. Cytokines
2.13. Statistical Analysis
3. Results
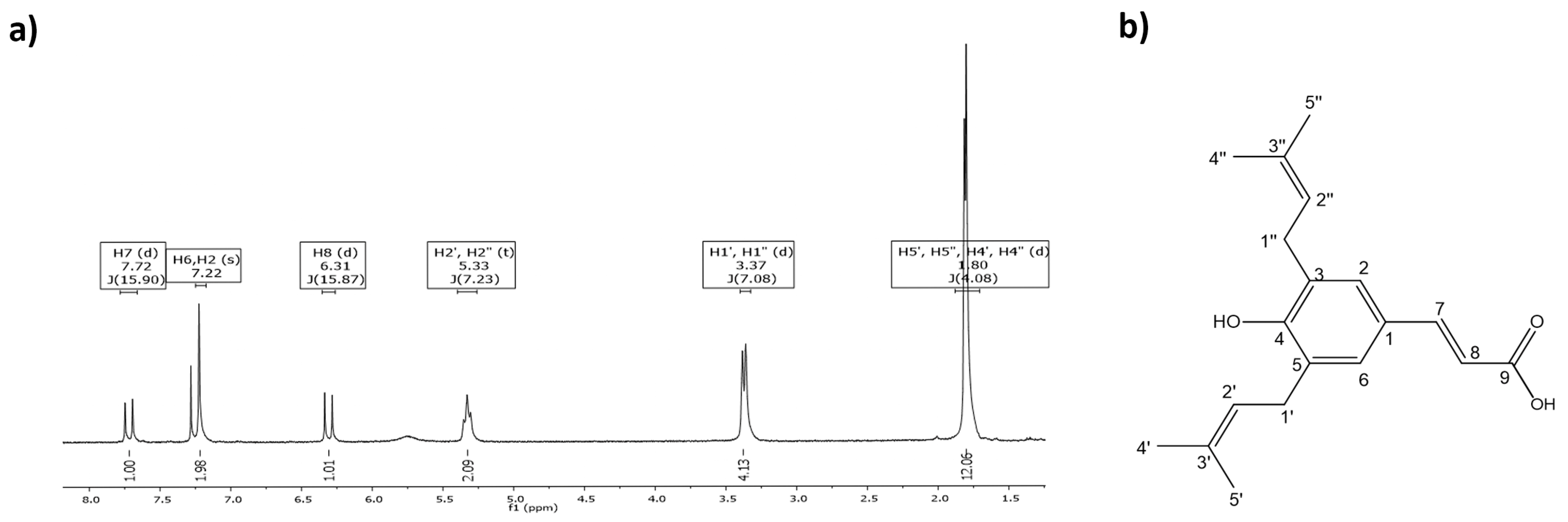
3.1. ArtC Isolation
3.2. ArtC Attenuates Allergic Airway Inflammation
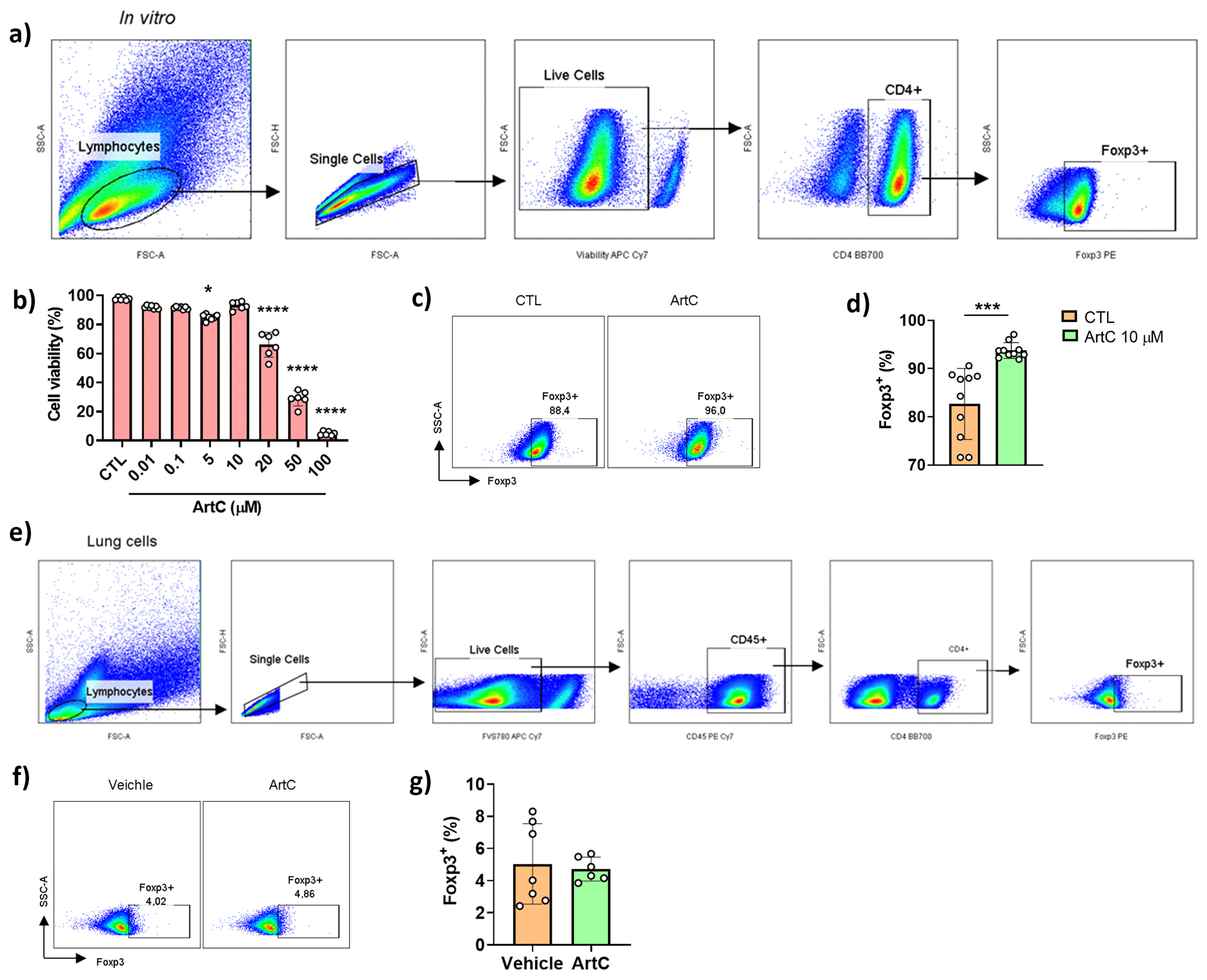
3.3. ArtC Induces Treg Cells Differentiation In Vitro, but Not In Vivo
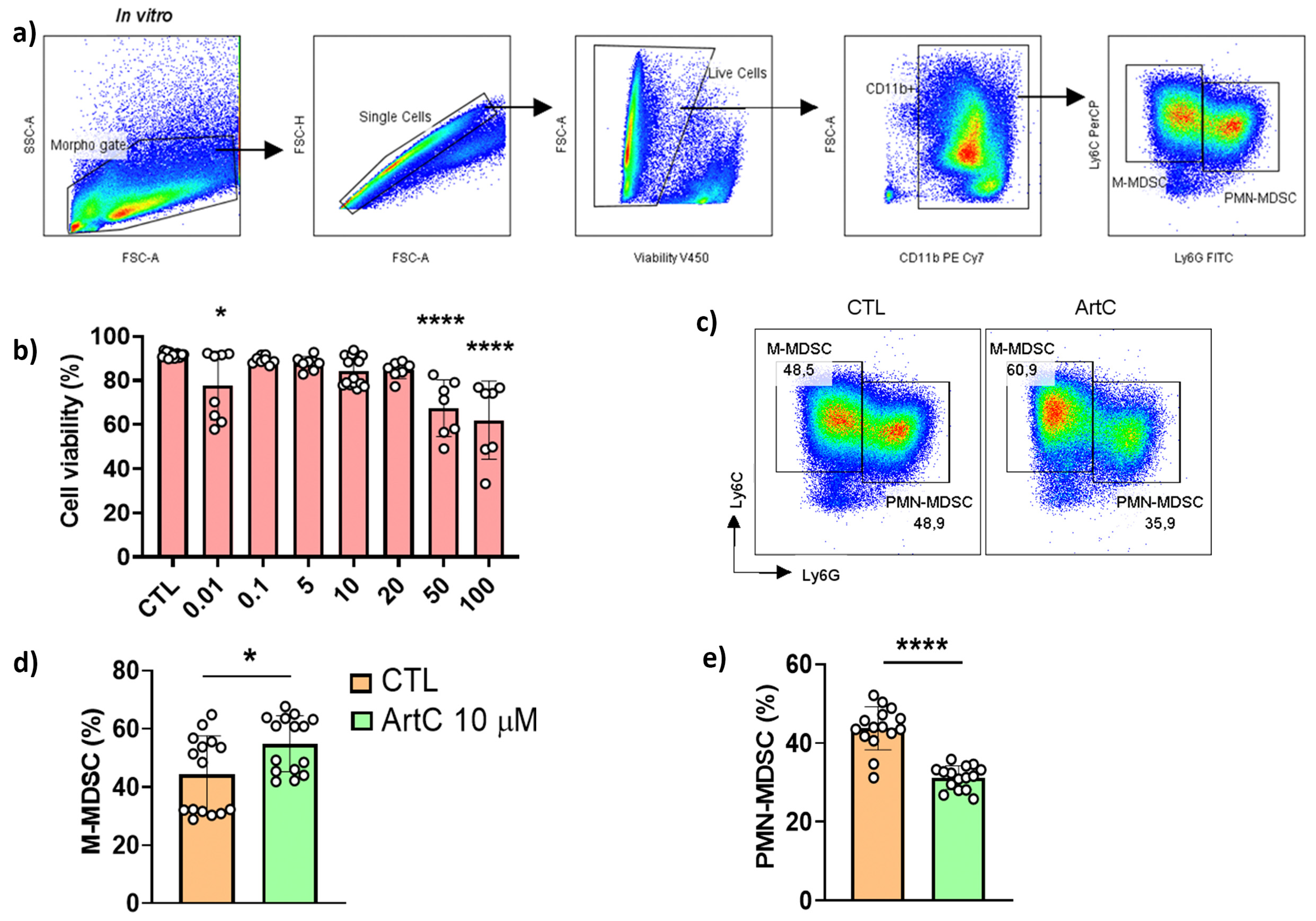
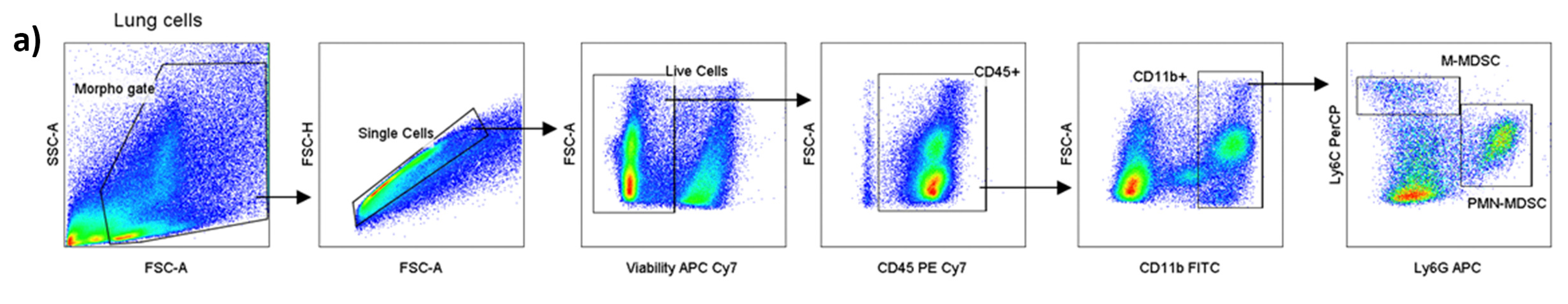
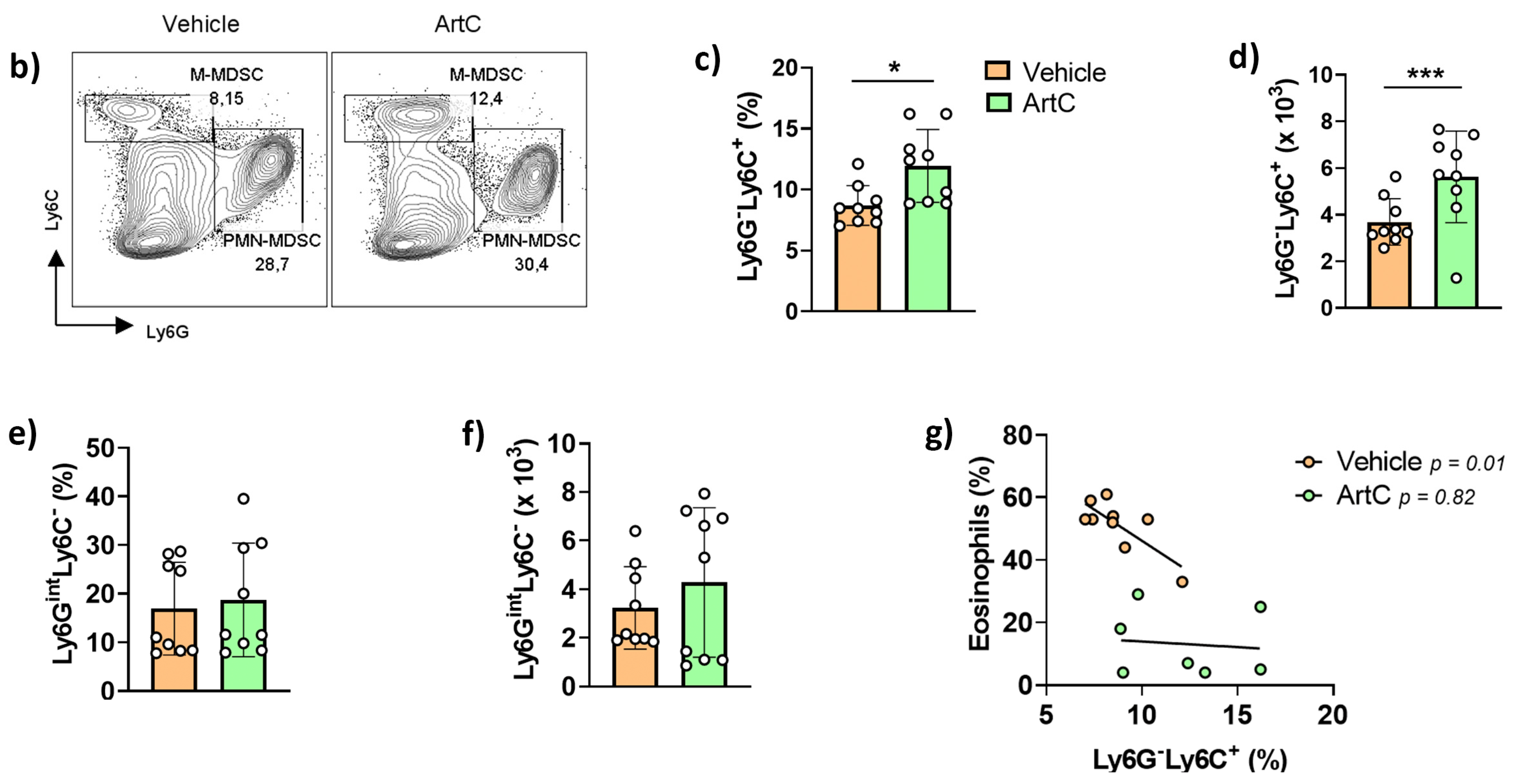
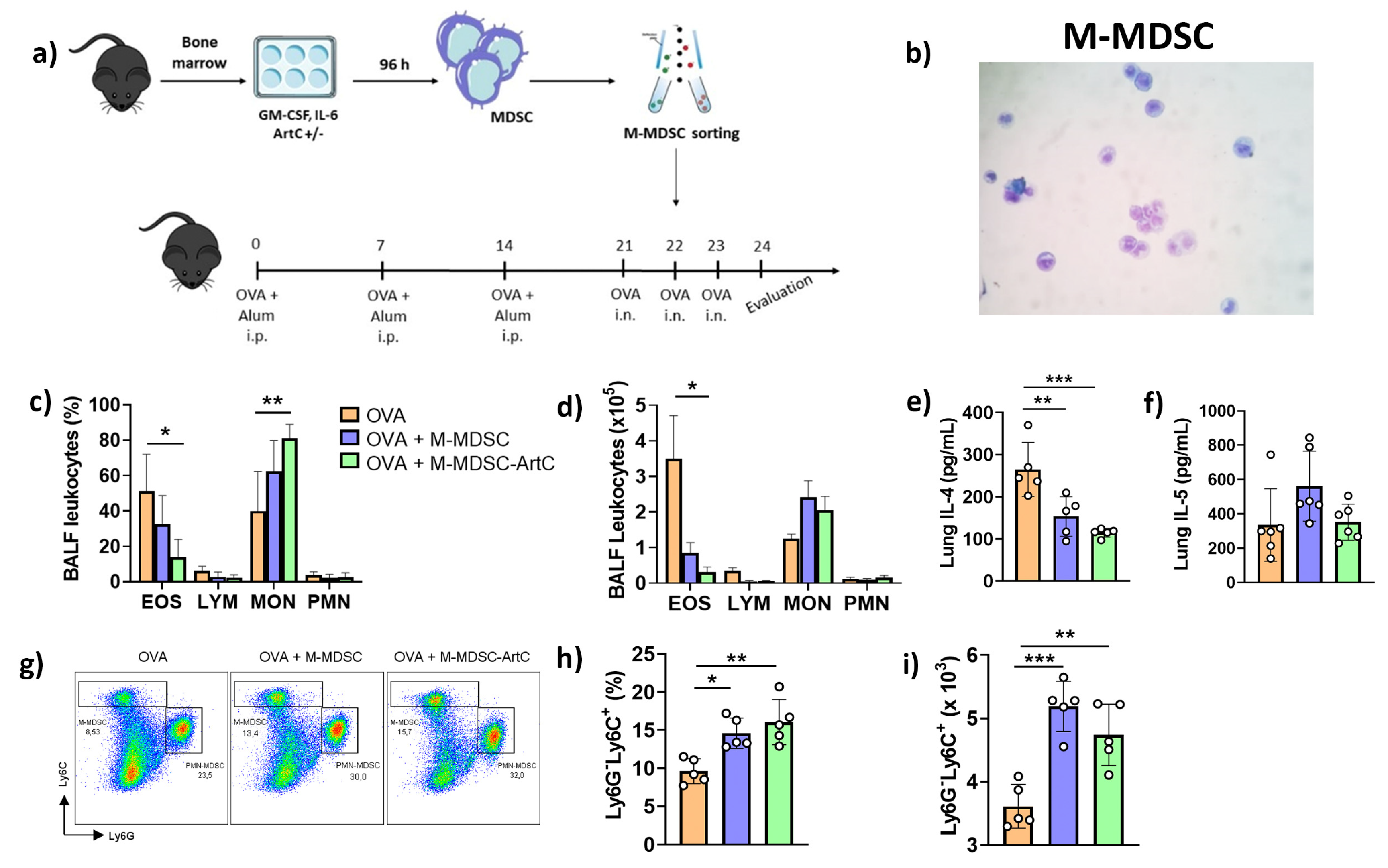
3.4. ArtC Augments M-MDSC Frequency In Vitro and In Vivo
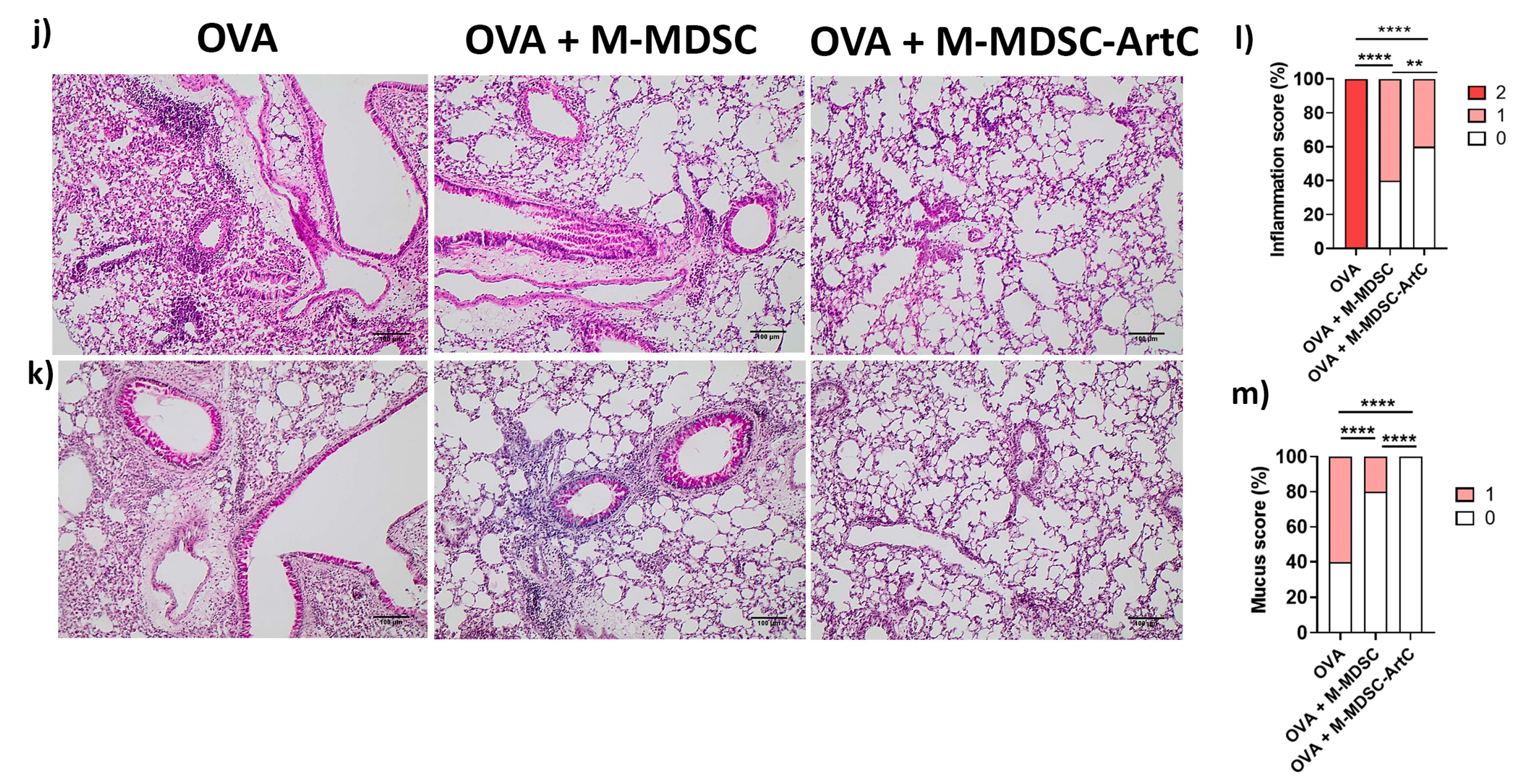
3.5. M-MDSC Suppress the Allergic Airway Inflammation
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Bazo, A.P.; Rodrigues, M.A.M.; Sforcin, J.M.; de Camargo, J.L.V.; Ribeiro, L.R.; Salvadori, D.M.F. Protective action of propolis on the rat colon carcinogenesis. Teratog. Carcinog. Mutagen. 2002, 22, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Cornara, L.; Biagi, M.; Xiao, J.; Burlando, B. Therapeutic properties of bioactive compounds from different honeybee products. Front. Pharmacol. 2017, 8, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Orsi, R.O.; Sforcin, J.M.; Funari, S.R.C.; Bankova, V. Effects of Brazilian and Bulgarian propolis on bactericidal activity of macrophages against Salmonella Typhimurium. Int. Immunopharmacol. 2005, 5, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Sforcin, J.M. Propolis and the immune system: A review. J. Ethnopharmacol. 2007, 113, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Machado, J.L.; Assunção, A.K.M.; Da Silva, M.C.P.; Dos Reis, A.S.; Costa, G.C.; Arruda, D.D.S.; Rocha, B.A.; Vaz, M.M.; Paes, A.M.; Guerra, R.N.M.; et al. Brazilian green propolis: Anti-inflammatory property by an immunomodulatory activity. Evid.-Based Complement. Altern. Med. 2012, 2012, 157652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Hariri, M.T.; Eldin, T.G.; Hashim, T.; Chathoth, S.; Alswied, A. Propolis Modulates Inflammatory Mediators and Improves Histopathology in Male Rats with L-arginine-induced Acute Pancreatitis. Sultan Qaboos Univ. Med. J. 2019, 19, 103. [Google Scholar] [CrossRef] [PubMed]
- Costa, P.; Somensi, L.B.; da Silva, R.d.C.M.V.d.A.; Mariano, L.N.B.; Boeing, T.; Longo, B.; Perfoll, E.; de Souza, P.; Gushiken, L.F.S.; Pellizzon, C.H.; et al. Role of the antioxidant properties in the gastroprotective and gastric healing activity promoted by Brazilian green propolis and the healing efficacy of Artepillin C. Inflammopharmacology 2020, 28, 1009–1025. [Google Scholar] [CrossRef] [PubMed]
- Sy, L.B.; Wu, Y.L.; Chiang, B.L.; Wang, Y.H.; Wu, W.M. Propolis extracts exhibit an immunoregulatory activity in an OVA-sensitized airway inflammatory animal model. Int. Immunopharmacol. 2006, 6, 1053–1060. [Google Scholar] [CrossRef] [PubMed]
- De Farias, J.H.C.D.; Reis, A.S.; Araújo, M.A.R.; Araújo, M.J.A.M.; Assunção, A.K.M.; De Farias, J.C.; Fialho, E.M.S.; Silva, L.A.; Costa, G.C.; Guerra, R.N.M.; et al. Effects of stingless bee propolis on experimental asthma. Evid.-Based Complement. Altern. Med. 2014, 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piñeros, A.R.; de Lima, M.H.F.; Rodrigues, T.; Gembre, A.F.; Bertolini, T.B.; Fonseca, M.D.; Berretta, A.A.; Ramalho, L.N.Z.; Cunha, F.Q.; Hori, J.I.; et al. Green propolis increases myeloid suppressor cells and CD4+Foxp3+ cells and reduces Th2 inflammation in the lungs after allergen exposure. J. Ethnopharmacol. 2020, 252, 112496. [Google Scholar] [CrossRef] [PubMed]
- Global Asthma Network. The Global Asthma Report 2018; Global Asthma Network: Auckland, New Zeland, 2018; ISBN 9780473465230. [Google Scholar]
- Lambrecht, B.N.; Hammad, H. The immunology of asthma. Nat. Immunol. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.J.; Brightling, C.E. Pathogenesis of asthma: Implications for precision medicine. Clin. Sci. 2017, 131, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- De Leonibus, C.; Attanasi, M.; Roze, Z.; Martin, B.; Marcovecchio, M.L.; Di Pillo, S.; Chiarelli, F.; Mohn, A. Influence of inhaled corticosteroids on pubertal growth and final height in asthmatic children. Pediatr. Allergy Immunol. 2016, 27, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Beserra, F.P.; Gushiken, L.F.S.; Hussni, M.F.; Ribeiro, V.P.; Bonamin, F.; Jackson, C.J.; Pellizzon, C.H.; Bastos, J.K. Artepillin C as an outstanding phenolic compound of Brazilian green propolis for disease treatment: A review on pharmacological aspects. Phytother. Res. 2021, 35, 2274–2286. [Google Scholar] [CrossRef] [PubMed]
- Szliszka, E.; Mertas, A.; Czuba, Z.P.; Król, W. Inhibition of inflammatory response by artepillin c in activated raw264.7 macrophages. Evid.-Based Complement. Altern. Med. 2013, 2013, 735176. [Google Scholar] [CrossRef] [PubMed]
- Paulino, N.; Abreu, S.R.L.; Uto, Y.; Koyama, D.; Nagasawa, H.; Hori, H.; Dirsch, V.M.; Vollmar, A.M.; Scremin, A.; Bretz, W.A. Anti-inflammatory effects of a bioavailable compound, Artepillin C, in Brazilian propolis. Eur. J. Pharmacol. 2008, 587, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.T.; Wang, C.Z. Regulatory T cells and asthma. J. Zhejiang Univ. Sci. B 2018, 19, 663–673. [Google Scholar] [CrossRef] [PubMed]
- Nagaraj, S.; Youn, J.-I.; Gabrilovich, D.I. Reciprocal Relationship between Myeloid-Derived Suppressor Cells and T Cells. J. Immunol. 2013, 191, 17–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consonni, F.M.; Porta, C.; Marino, A.; Pandolfo, C.; Mola, S.; Bleve, A.; Sica, A. Myeloid-derived suppressor cells: Ductile targets in disease. Front. Immunol. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Gantt, S.; Gervassi, A.; Jaspan, H.; Horton, H. The role of myeloid-derived suppressor cells in immune ontogeny. Front. Immunol. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrilovich, D.I. Myeloid-derived suppressor cells. Cancer Immunol. Res. 2017, 5, 3–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veglia, F.; Sanseviero, E.; Gabrilovich, D.I. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat. Rev. Immunol. 2021, 21, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, D.M.; De Souza, M.C.; Arruda, C.; Pereira, R.A.S.; Bastos, J.K. The Role of Baccharis dracunculifolia and its Chemical Profile on Green Propolis Production by Apis mellifera. J. Chem. Ecol. 2020, 46, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Prado, R.Q.; Bertolini, T.B.; Piñeros, A.R.; Gembre, A.F.; Ramos, S.G.; Silva, C.L.; Borges, M.C.; Bonato, V.L.D. Attenuation of experimental asthma by mycobacterial protein combined with CpG requires a TLR9-dependent IFN-γ-CCR2 signalling circuit. Clin. Exp. Allergy 2015, 45, 1459–1471. [Google Scholar] [CrossRef] [PubMed]
- Solito, S.; Pinton, L.; De Sanctis, F.; Ugel, S.; Bronte, V.; Mandruzzato, S.; Marigo, I. Methods to Measure MDSC Immune Suppressive Activity In Vitro and In Vivo. Curr. Protoc. Immunol. 2019, 124, e61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Aidy, W.K.; Ebeid, A.A.; Sallam, A.E.R.M.; Muhammad, I.E.; Abbas, A.T.; Kamal, M.A.; Sohrab, S.S. Evaluation of propolis, honey, and royal jelly in amelioration of peripheral blood leukocytes and lung inflammation in mouse conalbumin-induced asthma model. Saudi J. Biol. Sci. 2015, 22, 780–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talmadge, J.E.; Gabrilovich, D.I. History of myeloid-derived suppressor cells. Nat. Rev. Cancer 2013, 13, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Kolahian, S.; Öz, H.H.; Zhou, B.; Griessinger, C.M.; Rieber, N.; Hartl, D. The emerging role of myeloid-derived suppressor cells in lung diseases. Eur. Respir. J. 2016, 47, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; He, Y.; Wang, X.; Liu, Y.; Shi, K.; Zheng, Z.; Su, X.; Lei, A.; He, J.; Zhou, J. Polymorphonuclear myeloid-derived suppressor cells attenuate allergic airway inflammation by negatively regulating group 2 innate lymphoid cells. Immunology 2019, 156, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, R.; Yanagisawa, M.; Takahashi, N.; Kawada, T.; Kumazawa, S.; Yamaotsu, N.; Nakagome, I.; Hirono, S.; Tsuda, T. Brazilian propolis-derived components inhibit TNF-α-mediated downregulation of adiponectin expression via different mechanisms in 3T3-L1 adipocytes. Biochim. Biophys. Acta Gen. Subj. 2011, 1810, 695–703. [Google Scholar] [CrossRef] [PubMed]
- Ray, P.; Arora, M.; Poe, S.L.; Ray, A. Lung myeloid-derived suppressor cells and regulation of inflammation. Immunol. Res. 2011, 50, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Tan, X.; Hu, X.; Zhou, M.; Yan, J.; Ding, C. Tumor Microenvironment following Gemcitabine Treatment Favors Differentiation of Immunosuppressive Ly6C high Myeloid Cells. J. Immunol. 2020, 204, 212–223. [Google Scholar] [CrossRef] [PubMed]
- Drabczyk-Pluta, M.; Werner, T.; Hoffmann, D.; Leng, Q.; Chen, L.; Dittmer, U.; Zelinskyy, G. Granulocytic myeloid-derived suppressor cells suppress virus-specific CD8+ T cell responses during acute Friend retrovirus infection. Retrovirology 2017, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaufmann, J.; Lelis, F.J.N.; Teschner, A.C.; Fromm, K.; Rieber, N.; Hartl, D.; Beer-Hammer, S. Human monocytic myeloid-derived suppressor cells impair B-cell phenotype and function in vitro. Eur. J. Immunol. 2020, 50, 33–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cloots, R.H.E.; Sankaranarayanan, S.; Poynter, M.E.; Terwindt, E.; van Dijk, P.; Lamers, W.H.; Eleonore Köhler, S. Arginase 1 deletion in myeloid cells affects the inflammatory response in allergic asthma, but not lung mechanics, in female mice. BMC Pulm. Med. 2017, 17, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barron, L.; Smith, A.M.; El Kasmi, K.C.; Qualls, J.E.; Huang, X.; Cheever, A.; Borthwick, L.A.; Wilson, M.S.; Murray, P.J.; Wynn, T.A. Role of Arginase 1 from Myeloid Cells in Th2-Dominated Lung Inflammation. PLoS ONE 2013, 8, e61961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Mezzadra, R.; Schumacher, T.N. Regulation and Function of the PD-L1 Checkpoint. Immunity 2018, 48, 434–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.Z.; Yu, H.P.; Yu, R.; Zhang, Y.; Deng, H.J.; Chen, X. Passive transfer of lipopolysaccharide-derived myeloid-derived suppressor cells inhibits asthma-related airway inflammation. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4171–4181. [Google Scholar] [PubMed]
- Song, C.; Yuan, Y.; Wang, X.M.; Li, D.; Zhang, G.M.; Huang, B.; Feng, Z.H. Passive transfer of tumour-derived MDSCs inhibits asthma-related airway inflammation. Scand. J. Immunol. 2014, 79, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Shi, G.; Tang, J.; Kong, D.; Bao, Y.; Xiao, B.; Zuo, C.; Wang, T.; Wang, Q.; Shen, Y.; et al. Myeloid-derived suppressor cell function is diminished in aspirin-triggered allergic airway hyperresponsiveness in mice. J. Allergy Clin. Immunol. 2014, 134, 1163–1174.e16. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Type | Antibody | Fluorochrome | Clone |
|---|---|---|---|
| In vitro MDSC | CD11b | PE-Cy7 | M1/70 |
| Ly6C | PerCP Cy5.5 | AL-21 | |
| Ly6G | APC | 1A8 | |
| In vitro Treg cell | CD4 | BB700 | RM4-5 |
| Foxp3 | PE | MF23 | |
| In vivo MDSC | CD45 | PE-Cy7 | 30-F11 |
| CD11b | FITC | M1/70 | |
| Ly6C | PerCP Cy5.5 | AL-21 | |
| Ly6G | APC | 1A8 | |
| In vivo Treg cell | CD45 | PE-Cy7 | 30-F11 |
| CD4 | BB700 | RM4-5 | |
| Foxp3 | PE | MF23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martins, N.S.; de Campos Fraga-Silva, T.F.; Correa, G.F.; Boko, M.M.M.; Ramalho, L.N.Z.; Rodrigues, D.M.; Hori, J.I.; Costa, D.L.; Bastos, J.K.; Bonato, V.L.D. Artepillin C Reduces Allergic Airway Inflammation by Induction of Monocytic Myeloid-Derived Suppressor Cells. Pharmaceutics 2021, 13, 1763. https://doi.org/10.3390/pharmaceutics13111763
Martins NS, de Campos Fraga-Silva TF, Correa GF, Boko MMM, Ramalho LNZ, Rodrigues DM, Hori JI, Costa DL, Bastos JK, Bonato VLD. Artepillin C Reduces Allergic Airway Inflammation by Induction of Monocytic Myeloid-Derived Suppressor Cells. Pharmaceutics. 2021; 13(11):1763. https://doi.org/10.3390/pharmaceutics13111763
Chicago/Turabian StyleMartins, Núbia Sabrina, Thais Fernanda de Campos Fraga-Silva, Giseli Furlan Correa, Mèdéton Mahoussi Michaël Boko, Leandra Naira Zambelli Ramalho, Débora Munhoz Rodrigues, Juliana Issa Hori, Diego Luis Costa, Jairo Kenupp Bastos, and Vânia Luiza Deperon Bonato. 2021. "Artepillin C Reduces Allergic Airway Inflammation by Induction of Monocytic Myeloid-Derived Suppressor Cells" Pharmaceutics 13, no. 11: 1763. https://doi.org/10.3390/pharmaceutics13111763