
The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review



1
Centre for Tissue Engineering and Regenerative Medicine, Faculty of Medicine, Universiti Kebangsaan Malaysia, Cheras, Kuala Lumpur 56000, Malaysia
2
School of Health Sciences, Universiti Sains Malaysia, Kubang Kerian 16150, Malaysia
*
Author to whom correspondence should be addressed.
Pharmaceutics 2021, 13(7), 981; https://doi.org/10.3390/pharmaceutics13070981
Submission received: 12 April 2021
/
Revised: 23 June 2021
/
Accepted: 25 June 2021
/
Published: 29 June 2021
(This article belongs to the Section Biopharmaceutics)
Abstract
:A diverse range of normal flora populates the human skin and numbers are relatively different between individuals and parts of the skin. Humans and normal flora have formed a symbiotic relationship over a period of time. With numerous disease processes, the interaction between the host and normal flora can be interrupted. Unlike normal wound healing, which is complex and crucial to sustaining the skin’s physical barrier, chronic wounds, especially in diabetes, are wounds that fail to heal in a timely manner. The conditions become favorable for microbes to colonize and establish infections within the skin. These include secretions of various kinds of molecules, substances or even trigger the immune system to attack other cells required for wound healing. Additionally, the healing process can be slowed down by prolonging the inflammatory phase and delaying the wound repair process, which causes further destruction to the tissue. Antibiotics and wound dressings become the targeted therapy to treat chronic wounds. Though healing rates are improved, prolonged usage of these treatments could become ineffective or microbes may become resistant to the treatments. Considering all these factors, more studies are needed to comprehensively elucidate the role of human skin normal flora at the cellular and molecular level in a chronic injury. This article will review wound healing physiology and discuss the role of normal flora in the skin and chronic wounds.
1. Introduction
Skin wound healing is a highly complex and dynamic mechanism involving various regulatory cells and molecules integrating to complete the wound re-epithelialization cascade [1]. Once the cutaneous layer is broken, the embedded cellular and molecular substances within the skin layers will synchronize at the designated phases to initiate the healing mechanism. Even though cutaneous wound healing is a systematic process, the phases are overlapping; therefore, it is known as one of the most complicated biological processes in the human body [2]. A chronic wound can be described as a stalled wound or wound that cannot heal in the expected time frame (of less than 3 months) [3]. It is characterized by an abnormal phenotype of epidermis and dermis cells that originates from the wound bed. A chronic wound is caused by a local factor (infection), systemic factor (diabetes), or both. The local factor affects the wound healing recovery of a particular wounded area of the skin, while systemic factors refer to the progression of the wound healing altered by the medical condition or history [4]. Generally, the chronic wound is a healthcare and socioeconomic burden. Approximately 2% of the population in developed countries has the potential to develop chronic wounds, especially leg ulcers, once in a lifetime [5]. Chronic wounds affected around 5.7 million people in the USA alone, at the cost of $20 billion for treatment and management yearly. The incidence is predicted to rise significantly in the elderly due to diabetes [6]. Besides skin lesion and diabetes, there are various underlying diseases such as sickle cell anemia, calciphylaxis, systemic lupus erythematosus (SLE), skin disease, or impaired physiological states that include paralysis, malnourishment (lack of nutrients), aging, and poor mobility that can affect the sequence of healing events, resulting in non-healing or chronic wounds [7].
In diabetes, for instance, several complications can lead to chronic wounds that are microvascular or macrovascular. However, diabetic foot ulcer (DFU) is considered more alarming than any other complications as it has become the primary cause of morbidity and increased hospital care for diabetic patients [8]. In addition, poor vascular flow and life-threatening infections are the major causative factors in diabetic chronic wounds impairing the wound healing process [9]. According to Armstrong and his team (2017), the cost of treatment for diabetic foot care has exceeded the cost for common cancers, as the diabetic wound is responsible for more admissions than any other diabetic complication. The researchers further stated that diabetic patients who developed foot ulcerations are at two times higher risk of death within 5 years upon diagnosis than patients who are not diagnosed with foot ulcers [10]. Based on the 2015 prevalence data, the International Diabetes Federation (IDF) reported that approximately 463 million adults are living with diabetes in 2019 [11], and diabetic foot with lower extremity complications affects about 40 to 60 million people globally (International Diabetes Federation-Complications 2020). In addition, the World Health Organization (WHO) has estimated that diabetes will be the seventh foremost death cause in 2030 [12]. Recently, it has been reported that every 30 s, one leg is being amputated due to DFU in some part of the world [13]. For the past 10 years in Malaysia, there has been an increase in diabetic cases, mainly affecting individuals of 30 years or older [14] in which 15% of the diabetic patients developed DFU [15]. Meanwhile, a statistic showed that 17% of patients admitted to the General Hospital of Kuala Lumpur are mainly due to DFU [16].
2. Anatomy and Physiology of Skin
The skin is the largest organ of the body and is divided into three layers; epidermis, dermis, and hypodermis, also commonly known as subcutaneous tissue [17] (Figure 1). The function of the epidermis is to protect the skin from microbial infections, chemical hazards, and mechanical as well as thermal hazards [18]. The epidermis also helps to maintain body temperature and prevent water loss through homeostasis [19].
The epidermis layer is composed of five stratum layers, keratinocytes, melanocytes, Langerhans cells, and Merkel cells. Beneath the epidermal layer lies the dermis or dermal layer that is rich with blood supply, extracellular matrix (ECM), nutrients, mast cells, nerve endings, lymphatic, fibroblasts, and epidermal appendages. The dermis provides structural integrity for the skin and modulates all cellular activities, including thermoregulation and cell restoration. The hypodermis acts as an insulator by trapping heat, providing shock absorption, and structural support to the skin [20,21].
3. Skin Wound Healing
Normal skin wounds take around one to two months to heal [22]. It is a natural, biological, and sophisticated process that occurs after an injury in tissue, which involves blood cells, connective tissue, parenchymal cells, ECM, and soluble mediators such as cytokines and growth factors interacting with each other during the wound healing mechanism. Classically, there are four major phases in wound healing; hemostasis, inflammation, tissue proliferation, and tissue maturation or remodeling [23,24]. The underlying connective tissue is exposed upon injury to the skin, and the collagens attract platelets to the injury site. This triggers platelet aggregation to deliver clotting factors such as prothrombin and fibrinogen, which initiates platelet clotting, coagulation, and a complement cascade through extrinsic and intrinsic pathways. During the coagulation phase, the platelets release chemicals from their granules into the plasma, including cytokines, growth factors, and pro-inflammatory mediators such as ADP, serotonin, Von Willebrand factor, and prostaglandin. These further assist the platelets in adhering to the injury site, forming a platelet plug, activating clotting chemicals, and maintaining vasoconstriction. Thrombin is a coagulation factor II that converts fibrinogen into fibrin, an insoluble protein, as a final product at the end of the complement cascade, thus arresting the blood flow at the wounded site. After clot formation, the coagulation process is switched off to prevent thrombosis. Fibrinolysis removes the fibrin and maintains vascular patency in balance with blood coagulation and fibrin formation [25,26,27,28].
In the inflammation phase, aggregated platelets secrete chemoattractant to activate the inflamed cells. These aggregated platelets release growth factors and pro-inflammatory chemokines to recruit neutrophils and macrophages to phagocyte the debris and fight against infection. Meanwhile, the endothelial cells from the wounded site are being stimulated to produce growth factors such as epidermal growth factor (EGF), transforming growth factor-beta (TGF-β), and platelet-derived growth factor (PDGF) to synthesize fibroblasts. After neutrophils have cleared out debris from the injury site, they undergo cell death or return back into circulation. Macrophages predominate the inflammation phase by clearing the apoptotic neutrophils, unwanted necrotic tissue, dead cells, and toxic production from the site. Macrophages transform into M1 (pro-inflammatory) and M2 (anti-inflammatory); M1 macrophages modulate cytokines such as interleukin (IL)-1β, TNFα, IL-6, IL-12, and matrix metalloproteinase (MMPs) while M2 macrophages produce arginase, TGF-β, CCL18, PGE2, and IL-10. These macrophages have their distinct lineages respective to their specific functions. For example, M1 macrophages are responsible for wound clearance of microorganisms and inflammation effects on the wounded bed, while M2 macrophages stimulate an anti-inflammatory effect, modulation, and encourage wound adhesion. On top of that, M2 macrophages also help in reducing inflammation and promoting angiogenesis and tissue regeneration in the healing process [29,30]. This is followed by the proliferation phase in which fibroblasts, keratinocytes, and endothelial cells migrate, proliferate, and re-epithelialize through the denuded wound to form new blood vessels [31,32].
During this time, new ECM are produced and some of its constituents such as collagen, elastin, proteoglycans, and hyaluronic acid aid in the construction of the granulation layer to restore platelet clot formation and contracting the wound size [33,34]. In a normal wound, keratinocytes relocate to the wounded area as a cell sheet over the granulation tissue and differentiate to re-epithelize the skin via integrin-mediated binding interactions with ECM molecules. However, these cells become dysfunctional and fail to regenerate the damaged tissue in the chronic wound [35]. While it undergoes a remodeling or maturation phase, the wound typically swells, reddens, and becomes itchy. The newly formed granulation tissue contains collagen fibers, becomes more vascular, and decreases the number of cells or changes in cell size. Additionally, the epidermal layer is also constructed, whereby this layer is subjacent to the basal lamina through the external and internal layers of the wound. At the same time, all unused proteins are degraded [36]. The collagen is remodeled from collagen type III to collagen type I, regulated by MMPs, while myofibroblasts produce collagen fibers for wound contraction [37]. In addition, myofibroblast highly expresses cytoskeleton α-actin, a type of smooth muscle protein that is essential for the contraction of the blood vessel [38]. At the final stage of wound healing, the myofibroblasts will be destroyed while cross-linked collagen fibers are rearranged and regulated by growth factors to give the newly formed scar a better tensile strength [39]. Table 1 shows the difference between acute and chronic wounds.
4. Pathophysiology of Chronic Wounds and Diabetic Foot Ulcers
Healthy skin is essential for wound recovery when a minor injury is inflicted on the skin layer. This could be only possible with the association of molecules and cells residing within the skin. In chronic wounds, the cells are unhealthy or in abnormal conditions that cause prolong wound healing or showed no improvement. Generally, fibroblasts play a critical role in wound healing by synthesizing ECM (fibronectins, hyaluronan, and proteoglycans) and collagen [44]. Unlike acute wounds, fibroblasts in chronic wounds become senescent, decrease in proliferation, and migration to the wounded area is inhibited thus, hampering collagen production and deposition [45], as shown in Figure 2. Consequently, fibroblasts are unable to remodel the ECM, causing elevations of enzymes such as MMPs, collagenase, serine protease, and elastase. MMP is a type of zinc endopeptidase that can deteriorate the ECM components. There are 23 types of MMP classifications, however, only collagenase (MMP-1, MMP-8) and gelatinase (MMP-2 and MMP-9) actively participate in the wound healing stages. Under normal conditions, the secretions ratio between MMPs and tissue inhibitors of metalloproteinases (TIMPs) are equal. However, the imbalance expressions of MMPs and TIMPs can cause degradation and inhibition of growth factors and ECM components thus, delaying the wound healing process [46,47,48]. Keratinocytes are the building blocks for the epidermis, which trigger the release of cytokines when in contact with an injury or wound [49]. They also involve in the initiation, maintenance, and completion of the wound healing process to restore the epidermal barrier [50]. Keratinocytes at the wound edge become stalled in proliferation or hyperproliferative and show poor migratory activity to re-epithelialize, suggesting no improvement in wound recovery. However, the malfunction of keratinocytes during wound restoration in the molecular aspects is not clearly defined [51]. For example, the role of transforming growth factors (TGF-β) in wound healing is still elusive from the molecular aspects due to the paradoxical roles during the inflammatory phase. TGF-β either supports persistent inflammation by activating several pro-inflammatory cytokines (IL-1β, Tumour Necrosis Factor Alpha (TNF-α), and IL-6) or inhibits the activation of T cells to become effector T cells and encourage wound regeneration. In chronic wounds, some studies reported that high levels of TGF-β1 and β3 were observed in chronic ulcers, especially in the hyperkeratotic epidermis, while some studies observed low secretion levels of TGF-β in chronic wounds, which could be a possible reason for failure in wound closure. Overall, more evidence and in-depth understanding are needed to rule out the exact role of TGF-β in chronic wounds [51]. Cytokines such as TNF-α exhibit a negative effect on the healing process [52]. The increased production of TNF-α signifies prolonged inflammation, and once the dermis fibroblasts are destroyed, collagen and ECM production will stop [53].
Diabetic skin ulcers present as a painful sore with the disintegration of skin layers including the subcutaneous tissue. The ulcerations are usually found on the lower limbs, especially on the foot, and are known as DFU [54]. In most cases, DFU causes severe destructions to the joints, bones, and soft tissues of the ankle and foot [55]. DFU can be categorized based on the depth of the wound, ranking from 1–5, starting with superficial ulcer right up to foot gangrene following the Wagner grade system [56]. In most DFU cases, chronic wound healing impairment leads to bacterial infection that can result in tissue destruction and, if untreated, could cause lower limb amputation [57]. Similar to normal wounds, ulcerated skin appears red and swollen, except that it is warm, and produces pus or excessive exudate followed by an unpleasant odor [58]. Additionally, the ulcerated foot can be seen with systemic infection, extensive cellulitis (more than 2 cm distant from the ulceration), bone necrosis, or gangrene, with reduced oxygen to the limb [59]. Apart from this, high glucose content in the blood produces substances known as advanced glycation end products (AGEs), which trigger the release of TNF-α and IL-1β that interrupt the synthesis of collagen and diminishes proliferation to re-epithelialize the wound [60]. High sugar levels also cause factors like thromboxane, Von Willebrand factor, factor VIII fibrinogen, and plasminogen activator inhibitor levels to be elevated in DFU patients. These factors not only contribute to poor platelet adhesion and thrombosis but also impede fibrinolysis [61]. The pathophysiology of DFU is multifactorial and the ulcerations can be generated not only by one but several factors, which include immunopathy, neuropathy, neuroarthropathy, vasculopathy, and mechanical stress [62]. Figure 3 summarises the pathophysiology of DFU.
5. The Role of Normal Flora as a Protective Agent towards Skin
The skin’s normal flora plays a vital role in the maturation and homeostasis of cutaneous immunity. The human skin harbors around 1000 types of bacterial normal flora. Theoretically, normal flora is referred to as the bacterial normal flora since bacteria typically populates the human skin without causing any harm to healthy individuals [63]. Most of these bacteria live in the superficial layers of the stratum corneum and the upper parts of the hair follicles [64]. Some bacteria, however, reside in the deeper areas of the hair follicles that are beyond the reach of ordinary disinfection procedures. These bacteria are the reservoir for re-colonization after the removal of the surface bacteria [65]. Other common types of bacterial normal flora that can be found on the skin are Actino-bacteria, Firmicutes, Proteobacteria, and Bacteroidetes. However, the most abundant type of bacteria that take up the huge space of the skin are Staphylococcus epidermidis [66] and Staphylococcus aureus [67] followed by Staphylococcus haemolyticus, Staphylococcus hominis [68], and Micrococci species [69].
A varying range of microbial flora inhabits the skin immediately after a person is born. Most of them are either mutualistic or show commensalism, and once they have established settlement, the skin microbial communities remain constant and stable on the skin over time [70]. The interactions between these microbe—host can be interrupted if some factors cause alterations to the skin structure, whether internally or externally [71]. These factors could include gender, age, medications (antibiotics), disease, different geographical regions, and lifestyle [72]. Haro et al.’s (2016) study on age and gender factors postulated that the microbes could change into different cell morphologies on the skin at the early and late years of a person’s life [73]. In addition, the difference of skin surfaces based on gender due to inconsistent secretions of hormones, skin pH, oil, and sweat could lead to favouritism for some microbes to populate the skin more than the others [74]. In relation to females, males have a greater abundance of lipophilic bacteria, especially Propionibacterium and Corynebacterium, because their skins are highly acidic, the enormous micro-colony size of bacteria, high production of sweat, and inadequate hygiene [72]. However, Shami et al. (2019) argued that gender did not influence the number or diversity of microbes on the cutaneous layer [69]. In addition, antibiotics are antimicrobial substances to inhibit proliferation or destroy microorganisms. In some skin conditions, these antibiotics profoundly obstruct the growth of normal flora and provide entry to opportunistic pathogens for disease exacerbation [75]. For example, patients with rosacea indicated that Cutibacterium acne protected the skin against harmful microbes by converting the oil into fatty acid, which could acidify the skin surface and inhibit colonization of pathogens [76,77]. However, in a different case with acne, antibiotic minocycline suppresses Cutibacterium acne thus, increasing the growth of Streptococcus species and Pseudomonas species while destroying other microbial flora that inhabit the skin [78]. A study was conducted to observe the behavior of skin normal flora among children and teenagers in rural and urban areas. The findings revealed that the type of outdoor activities and some physiological changes that occur due to different age range could diversify the dynamics of the normal flora on the skin [79]. Lifestyle, particularly hygiene and cosmetics, also influence the dynamics of the normal flora [80]. Even though soaps, cosmetics, and other body shower products help clean the skin, they can eradicate the healthy normal flora already residing on the skin, which could lead to the growth of foreign microbes [81].
The bacterial survival and the extent of colonization partially depends on (1) exposure of the skin to a particular environment, (2) innate and species-specific bactericidal activity on the skin, and (3) a high degree of specificity involved in the adherence of bacteria to the skin epithelial surfaces. Most bacteria do not attach to the skin. Staphylococci are mostly found as nasal flora, and this species outnumbered the viridan streptococci in conquering the nasal mucosa site. On the contrary, viridan streptococci can be seen dominating the oral canal but not the skin or nose [82,83]. The colonization of normal flora depends on the part of the body that is suitable for their growth, which includes oily (face), moist (armpit), or some dry environments (forearm and buttock). Staphylococcus species and Propionibacterium are commonly found in the oily part of the body, while Corynebacteria and b-Proteobacteria inhabit the moist areas of the body. The dry environments are occupied mainly by bacteria such as b-Proteobacteria, Corynebacteria, and Flavobacteriales [84,85]. The skin normal flora harmonizes with various innate factors, including complement, IL-1α, and antimicrobial peptides (AMPs) [86]. Normal flora release substances called phenol soluble modulins (PSMs) and bacteriocins, particularly from S. epidermidis and acnecin from P. acnes (or also known as Cutibacterium acne), to maintain the skin barrier from harmful microbes [87]. On top of that, S. epidermidis synthesizes secretome to reduce skin inflammation caused by S. aureus during an allergic reaction. This will be achieved by omitting several factors such as peptidoglycan, lipopeptide LP01, lipoteichoic acid (LTA) acid, and activation of IL-10 through modulation with innate and adaptive immunity [88]. S. epidermidis also induces the secretion of AMP (human beta defensin, HBd-2 and cathelicidins, and LL-37) produced by epidermal keratinocytes and skin commensal organisms. Then, it activates the toll-like receptor (TLR)-2 signaling pathway through the innate immune system. This response is important to promote the eradication of pathogenic bacteria and encourage wound healing [89,90].
S. epidermidis is able to increase the frequency of CD8 T cells along with IL-17A or IFN-γ to enhance the epidermal barrier and limit pathogen invasions [91]. Besides this, a reduced number of skin resident microbes with an increase of Staphylococcus species and Corynebacterium bovis were detected in a mice model of atopic dermatitis (AD). The findings showed that lack of skin normal flora, dysbiosis, and overgrowth of both S. aureus and C. bovis are responsible for acute atopic flares in patients diagnosed with AD [92]. Despite this, there is another mechanism utilized by skin normal flora, known as pattern recognition receptors (PRRs), which secrete nucleotide-binding oligomerization domain containing 2 (NOD2) that bind to peptidoglycans of Gram-positive and Gram-negative bacteria [93]. The mechanism is equally important because it helps the skin commensals recognize potential pathogens and initiate the innate immune system for further elimination. NOD2 with TLR2 and TLR6 are more specific to protect the skin from S. aureus, and likewise, TLR 2, 3, 7, 8, and 9 act as a defender against infections with herpesviruses, papillomaviruses, and poxviruses [94].
Besides bacteria, other types of microbes reside on the skin, such as fungi, viruses, and parasites considered normal flora of a healthy skin [95]. Examination of 14 different body locations such as foot, heel, toenail, and toe web of 10 healthy adult participants showed that Malassezia fungi accountable for most fungi colonization on healthy skin [96]. These fungi species are found in large numbers on oily skin containing lipid secreted by the sebaceous gland [97]. Although the benefits of Malassezia species to the skin host are still unclear, a study showed that this fungi species might enhance the activity of the epithelial cells by synthesizing aryl hydrocarbon receptor (Ahr) ligands that act as a shield against UV light [98]. Aspergillus and Penicillium are also commensal fungi, however, they are expressed in a lower numbers. Meanwhile, Candida and Dermatophytes can be either commensal or parasitic depending on the skin condition of the host [99]. For example, Candida albicans increases the activation of antigen-specific Th17 cells and destroys any non-commensal fungi that inhabit the local tissue by binding to their respective epitopes [100]. The bacterial–fungi interactions possess more virulence and resistance properties than with a single organism. The biofilms of both, containing Staphylococcus epidermidis and Candida albicans, exhibit resistance towards antibiotics compared to their single biofilms [101]. An in vivo study on mice was conducted to investigate the effects of the papilloma virus on carcinogenic skin cancer by introducing the strains into the immunocompetent mice. The investigation revealed that the commensal papilloma virus could trigger T cells to reduce the virulence of the virus, hence suppressing the growth of cancerous cells in the immunocompetent mice [102,103,104].
6. The Role of Microbes in Wound Healing
6.1. Acute Wound Healing
Normal skin aims to maintain its integrity by controlling the skin inhabitants from becoming harmful to the skin or preventing any penetration into the underlying tissue [105]. A slight alteration on the skin barrier may cause clinical changes to the skin, such as skin inflammation, allergic reaction, skin infections, formation of tumors in the superficial layer of the skin, and impaired skin healing [106]. Like other living organisms, microorganisms need essential requirements such as suitable temperature, pH, and nutrients for them to survive and multiply [107]. This can be achieved through the open cutaneous wound, however, the total population and the type of microorganisms depend on the severity of the wound and the immune level of the host [108]. The normal pH of healthy skin for both males and females is between 4 to 5, which is acidic to significant fractions of the human body [109]. The acidic condition is one of the ways to protect the skin from exogenous microorganisms [110]. The acidic pH of the skin is a desirable condition for S. epidermidis to preserve and attach to the skin, whereby this condition is unfavorable or could inhibit the growth of some skin pathogens such as S. aureus and P. acne. Most skin pathogens prefer an alkaline environment [111]. The pH of the wound either facilitates the growth or suppresses the colonization of pathogenic bacteria into the skin [112]. Under acute wound healing, when the wound becomes acidic, the amount of lactic acid and oxygen will reduce the pH of the skin. Acidic condition of the wound is needed for macrophages response, multiplication of fibroblasts, angiogenesis, collagen synthesis, and DNA formation to facilitate wound closure. In contrast, a wound that develops infection appears to increase in an alkalinity environment [113,114]. Temperature influences the wound healing rate. Any temperature that falls below the average body temperature will delay the wound healing process. In normal wound healing, the temperature will be elevated in the first few days before returning to the normal temperature due to the activities of the immune system and other physiological responses related to the healing process [115,116].
Recently, a study showed that exposure to UV radiation elicited an immune response through the synthesis of AMPs; however, concurrently, it caused immune suppression of specific cytokines and other reactions such as skin cancer, photoallergic, and phototoxic. Therefore, normal flora plays an important role in protecting the skin caused by ultra-radiation, thus preventing immune suppression [117]. Besides, skin normal flora can also elicit some other diseases such as impetigo, cellulitis, and systemic diseases, including endocarditis and sepsis [118]. When bacteria propagate inside the wound, they produce small chemicals used as signals to communicate with each other, as well as affecting the host’s immune cells and blood vessels. Upon detection, immune cells respond to these signals to kill the bacteria and limit the spread of the infections. One example is when epidermal keratinocytes contact pathogenic microbes, they will release a range of cytokines and chemokines to initiate a defensive response. This, in turn, will activate the production of human beta-defensin, cathelicidin, RNase-family antimicrobial peptides, and reactive oxygen species to protect the skin layer from further invasion by these pathogenic microbes [118]. A study conducted by Yang et al. (2017) postulated that re-epithelialization of epithelial cells is important for a wound to recover due to the capability of the cells to proliferate, differentiate, and protect from microbe infections. This can be achieved through IL-27, which is known to have a functional role in regulating wound re-epithelialization. The study also hypothesized that IL-27 secreted by dendritic cells and macrophages exhibited antiviral response through JAK/STAT3 signaling pathway. In addition, the expression of IL-27 is elevated in an in vitro model of mice skin after wound closure [119].
Recent studies also found that S. epidermidis plays an active role in skin immunity by activating cytotoxic CD8 T cells (CTLs) that belong to major histocompatibility complex Ib (MHC1b), and conferred N-formyl methione peptides on antigen-presenting molecules with the help of dendritic cells to CTLs for further actions. The studies also included that RNA gene sequencing on CD8 T lymphocytes was stimulated by S. epidermidis. The analysis results showed that immune regulation and genes associated with tissue repair were up-regulated. Through this pathway, both the bacteria and T cells prevent any foreign particle invasion and help hasten skin wound healing [106,120]. Moreover, a similar finding has been proven in skin biopsy samples whereby induced wound injury exposed to S. epidermidis drastically improved re-epithelialization and tissue granulation [106]. When an infection occurs, macrophages at the wounded site get in contact with pathogens and secrete cytokines such as TNF-α and IL-1β to regulate the endothelial cells to synthesis leucocytes’ adhesion molecules and chemokines. These will regulate the activity of leucocytes and help them to relocate to the infected area. Basically, leucocytes only move to the injury site after macrophages detect pathogen-associated molecular patterns (PAMPs) of the microorganisms through toll-like receptors and activation of the immune system [121]. An adopter molecule in the TLR signaling pathway known as MyD88 also accelerates the wound healing process since the MyD88 pathway assists in the production of granulation tissue, wound closure, and angiogenesis [122], as shown in Figure 4.
Bay et al. (2018) conducted a clinical study to observe bacterial infections that occurred in the wound at the superficial layer of the epidermis. The group concluded that bacteria are always looking for any opportunity to reproduce and multiply, especially when there is a region that contains all the necessary conditions for them to survive. Such a situation happens when there is an entry into the skin and favorable conditions for the bacteria to colonize and multiply excessively. Examples are keratin residues from the broken layer of stratum corneum and fibrin accumulation at the borderline of the wound bed. This finding assumes that bacterial aggregation happens because the immune cells cannot prevent bacterial entry or immune cells are not allowed to enter the particular region colonized by bacteria (Figure 5). They also concluded that bacterial accumulation is present in all types of wounds except in chronic cases, which is more notable [123]. During acute wound healing, fibroblasts synthesized keratinocytes growth factor I to produce more keratinocytes and move to the wounded site. A recent study used a porcine model to evaluate the effects of the uninfected and infected wound with single species, followed by the wound co-infected with both P. aeruginosa and methicillin-resistant S. aureus (MRSA). The findings demonstrated that epithelialization could not occur due to low regulation of keratinocyte growth factor 1, as shown by decreased synthesis of fibroblasts in the co-infected wound. This could explain that mixed-species directly influence fibroblasts’ production, thus impairing the wound healing process [124]. An investigation conducted on pro-inflammatory interleukin IL-17A by using knockout mice (KO) demonstrated that wounds gradually healed and control activity of neutrophils against skin pathogens was observed in normal wounds compared to the wild mice with recombinant IL-17A. However, Vγ4 T-derived IL-17A is the main compound that converts IL-17 to inflammatory cytokine, impeding the function of dendritic epidermal T cells (DETCs) from producing keratinocytes and halts the migration of damaged tissue. This indicates that IL-17A could be another cytokine responsible for poor wound healing if secreted excessively [125]. In most acute wound cases, Gram-positive bacteria outnumbered Gram-negative bacteria. This can be seen in acute wounds whereby Staphylococcus species populations are greater than Pseudomonas species alongside skin undergoing wound healing [126].
6.2. Chronic Wound Healing
There are few classifications of microorganisms that are accountable for infection in a chronic wound. Such microorganisms are Gram-positive bacteria, including Staphylococcus aureus and Staphylococcus epidermidis. The Gram-negative bacteria are Escherichia coli, Proteus mirabilis, Pseudomonas aeruginosa, Enterobacter species, and Morganella species. Compared to the rest of the microbes, S. aureus is mostly resistant to antibiotics and contains high virulence, contributing to the pathogenicity of the host [127]. Furthermore, the infectious bacteria present in the wound bed tend to degrade the ECM and growth factors. The bacteria that already conquered the wound sheet generally produce biofilms acting as a barrier to allow them to grow, multiply, and protect from immune cells or become resistant to the antibiotic. The biofilm architecture comprises a fraction of bacteria implanted in the extracellular polysaccharide matrix or extracellular polymeric substances (EPS). Additionally, this biofilm is toxic to the other skin cells, explaining the delay in wound healing [46,128,129]. The EPS is water-based, containing a matrix with some protein substances that help channel nutrients, movement, and communication between the bacterial communities in a biofilm. In brief, EPS is the main component of most bacterial biofilms that support colonization or recolonization, by adhering to the wounded surface area [125,130]. Most bacteria exhibit some type of chemotaxis for movement, colonization and cause disease in a host. Chemotaxis can be defined as receptor-mediated, directed cell movement in a soluble chemical attractant concentration gradient known as a chemoattractant. Skin pathogens, especially S. aureus and S. pneumoniae, are incapable of being chemoattractant use various adhesion methods to attach themselves onto the surface of host tissue or cells and utilize the available nutrients while dispersing the virulence contents into the host. The adhesions include pili, fimbriae, and lipoproteins [131].
Bacterial biofilm formation is one of the indicators for its presence in a chronic wound. More than 50% of the biofilms are detectable via microscopic observation compared to only 6% in an acute wound. The bacteria species and the relative number varied from one wound to another [132,133]. Some anaerobic bacteria can survive and multiply deeper in a biofilm even though the oxygen level is depleted. Therefore, it is vital to investigate the type of bacterial strains and their relationship within the wound since it is insufficient to kill the biofilms cells by looking into the bacterial colonies alone [133]. In another study, Iwase and colleagues (2010) showed that S. epidermidis can block biofilm formation by S. aureus [134]. Similarly, S. epidermidis can form biofilm in chronic wound infection through implanted devices and host tissue, which can lead to septicemia [135]. The biofilm of S. epidermidis contains a protein known as polysaccharide intercellular adhesion (PIA), encoded by the ica operon gene, which helps in the formation of biofilm and adhesion by the Staphylococcus species [136]. PIA or polymeric-N-acetyl-glucosamine (PNAG) is a complement activator produced by C3 and C5. On most occasions, S. epidermidis can produce PNAG-dependent biofilms as a shield to protect itself from IgG and C3 factor, inhibit phagocytosis and destruction by neutrophils and prevent the antibody from reaching the surface of the biofilm. Through this, neutrophils’ reactions could be diverted and become ineffective in destroying Staphylococcus species [137]. Unlike methicillin-resistant S. aureus commonly found in other anaerobic bacterial colonies or mainly joined with Gram-positive bacteria, Pseudomonas aeruginosa independently (planktonic state) can produce biofilm resembling archetypal mushroom with enclosed ECM [138].
Several pathways are involved in the production of Pseudomonas aeruginosa biofilm. One of the pathways is cyclic diguanosine-5′-monophosphate (c-di-GMP), which will enhance bacterial cell attachment, cell clumping, and EPS release to the surrounding wound that causes severe infections [139]. Complement cascade activation in the skin wound has shown some conversions in wound healing [140]. It is considered that complement activation helps protect against infection and inhibits wound healing if the activation is dysregulated, especially CD59. Dysregulation of CD59 increases cytokine release, cell multiplication, and inflammation that can cause damage to the tissue [141]. During Staphylococcus species invasion, the complement cascade is stimulated by three separate routes that activate the C5a and C5b factors. C5a captivates neutrophils for bacterial engulfment, whereas C5b produces a toxic vent to target the bacterial membrane and destroy gram-negative bacteria [142]. On the contrary, Gram-positive bacteria are impenetrable by membrane attack complex due to their thick peptidoglycan membrane [143]. PTX3 is a substance produced by antibody-mediated immunity that works with plasminogen and fibrin in the coagulation phase. In a chronic wound, PTX3 acts as a mediator between antibody-mediated immunity and cellular pathways to activate phagocytosis when in contact with any infectious microbes [144]. In a chronic wound, due to some catabolic reactions, the oxygen level will reduce the skin′s pH, hence altering the wound healing phases such as collagen production, blood vessel formation, and immune system [113]. Chronic wounds are characterized by high numbers of Langerhans cells, neutrophils, pro-inflammatory macrophages [42], pro-inflammatory cytokines, reactive oxygen species [145], and protease [146]. Similar to normal wound healing, these immune cells received signals from innate cellular immunity and propagated the signal to the infected wound bed for further actions. However, the immune cells are unable to perform their regular duty as they become defective to function. The dysregulation of neutrophils and macrophages becomes less effective in inflammatory effect and phagocyte the bacteria, contributing to the delay in wound healing [27,42]. Neutrophils utilize neutrophil extracellular traps (NETs) to kill infectious microbes or biofilms by releasing chromatins and granular protein contents. It is also known as NETosis. NETs accomplished their tasks using two strategies. The first is to destroy or stop the proliferation of pathogens, and the second is to disable the migration of the pathogens. Despite its role against microbe infection, accumulated evidence has shown that microbes can surpass NETs antibacterial activity and become resistant to it by degrading NETs by enzyme nucleases and inhibiting NETs synthesis [29]. In addition, S. aureus is able to induce extracellular trap formation by releasing leucocidin from the biofilm to escape the antimicrobial activity of NETs. This encourages the multiplication of bacterial colonies to disperse to a new place for new biofilm formation; thus, helps the bacteria to sustain and survive in the chronic wound for a longer time [147]. As the focus has been mainly on the diverse bacterial pathogens in chronic wounds, the fungi’s role in the wound is considered significant. The imbalance number of commensals and pathogenic fungi residing at the skin barrier also contributes to the delay of wound healing. Candida species and Trichosporon species could hinder wound healing either via its spores or biofilms disposition to exacerbate the inflammation and consume oxygen contents resulting in necrosis [138,148]. A study has illustrated that Candida albicans could defect the role of macrophage and cell apoptosis in cutaneous wound healing [149]. IL-27 produced from CD301b+ immune cells could exhibit antiviral properties suggesting its vital role in skin regeneration [119,150]. It is thought that IL-27 could be able to quell the infection caused by the Zika virus through the activation of STAT 1 signal transducer and activators of transcription 1 (dependent) to STAT 2 (independent) [151]. A different virus has different ways to seize the host for replication. For example, in a cutaneous wound, Herpes simplex virus (HSV) replicates in the epidermis by forming glycoprotein, fused, and attaches to the keratinocytes before disseminating its viral particles to initiate infection. Many studies have investigated the skin normal flora or microbiome related to protection and thermoregulation, however, studies on the role of fungi and virus as cutaneous residents are still scarce [152].
The severity of DFU corresponds to the amount of Staphylococcus species present in the wound. The time taken for ulcer formation depends on the diversity and level of Proteobacteria in the wound [126]. Chronic ulceration of diabetes could be caused by more than one type of skin commensals, producing a synergetic effect that converts non-virulent bacterial species to virulent and causing damage to the host. This has been proved based on the high throughput of 16S rRNA gene sequencing [153]. Recently, endotoxin secretion from Gram-negative bacteria has been observed in diabetes patients and it is likely to induce insulin inefficiency through the elevation of pro-inflammatory cytokines in the adipocytes such as TNF-α [154]. During the pathogenesis of chronic wounds, an infection caused by S. aureus increases glucose resistance by blocking the insulin to its target site hence elevating the glucose level in the blood [155]. Besides that, biofilms’ continuous presence could delay wound healing in the DFU by releasing the inflammatory cytokines, free radicals, nitric oxide, and complement initiation through the activation of immune cells [156] as shown in Figure 5. In diabetic conditions, neutrophils become hyperactive and secrete high amounts of TNF-α, which will increase the process of NETs formation to destroy neutrophils, hence impairing wound healing [157]. In contrast, some skin bacteria, either Gram-positive or Gram-negative, can regulate the NETs formation by releasing the exotoxins to suppress the activity of skin pathogens, thereby boosting the defense system and mediating phagocytosis [29]. The chronic skins of diabetic patients have shown a higher number of mast cells and macrophages [158,159], while T cell receptors and CD4 T cell numbers are reduced [160]. The presence of these cells in chronic wounds could explain the prevention of wound recovery, promoting skin infection, and jeopardizing the wound healing system [159].
A study to examine the effects of the Circoviridae virus on two different groups of children with Type I diabetes and a control group revealed that Circoviridae species is more pervasive in the control group than in diabetic children. However, most children did not exhibit any signs and symptoms after being infected with this virus and showed no significant difference in both groups [161]. Although some studies reported that the virus could protect from developing type 1 diabetes, some studies revealed a contradictory report. It seems that virus infection in murine models induced with type I diabetes shows impairment in the function of pancreatic Langerhans β-cells and autoimmunity activation, which could eventually lead to cell destruction and apoptosis [162]. In diabetic wound healing, the increased level of TLR especially TLR2 signaling pathway proteins (MyD88, pIRAK, and TRIF) and their inflammatory cytokines (IL-1β and TNF-α) were high in induced mice compared to non-induced mice. The significant increase of these levels in diabetic mice did not show any improvement in wound healing, however, it extended the duration of the inflammation phase [122]. Germ-free mice (without commensals) showed increased wound epithelialization, wound closure, and angiogenesis with lesser scar formation than non-germ-free mice (with commensals). These mice showed persistent inflammation, cytokine release, and wound cessation, concluding that the presence of skin normal flora decreases the efficiency in wound healing [163]. On the contrary, mice that have been induced with different types of antibiotics before wound incision showed decelerated wound healing than mice without antibiotics [126].
7. Treatment for Chronic Wounds and Diabetic Foot Ulcers
The use of antibiotics in treating chronic wounds is challenging since the microorganisms, especially bacteria, tend to become resistant to the prolonged treatments with no improvement in wound healing besides causing kidney failure and oral tract infection. The antibiotic treatments can be administered to the patients either by parenteral, oral, or topical [164]. Since some antibiotics can reduce a type of pathogen while increasing the growth of other pathogens, an empirical antibiotic choice should be carefully selected based on the clinical examinations, the severity of the infection, antimicrobial sensitivity pattern, and the aetiological agent. A broad spectrum of parenteral antibiotics is administered for severe infections, while narrow-spectrum oral antibiotics are administered for mild infections [165].
Besides drugs, there are different types of wound dressings applied for wound treatments, including passive dressings (such as gauze), interactive dressings (such as foam or sponge), advanced dressings (such as alginate or hydrocolloid), bioactive dressings (such as Alloderm and Apligraf), and antimicrobial dressings (such as a topical antibiotic). The goal of a good dressing is to retain moisture for wound closure, prevent infections, reduce pain or irritation, and scar formation. Additionally, a selection of dressing also depends on the application and type of wound. For example, a gauze dressing is used in a shallow flat wound for less exudate production, while foam, sponge, and alginate are suitable for minor burn or deep wounds to absorb excessive exudate from the wound. Bioactive or tissue-engineered dressings accelerate wound healing by mimicking the natural function of ECM and mediating the physiology of healing phases through angiogenesis, cell proliferation, and new tissue formation. Antimicrobial dressings are mainly applied topically to chronic wounds that are infected with bacteria [6]. Traditional wound dressings are mainly used to maintain the dryness of the wounds and prevent infections but cannot absorb a large number of exudates. Different strains of normal flora and pathogens show different interactions and impacts on the usage of the wound dressing. Another study reported that besides improving the physicochemical properties of the dressing materials, loading additional substances into the dressing materials could also prevent microbial infections [166]. Jack and colleagues (2017) have used wood nanocellulose hydrogel suspension to observe the activity of P. aeruginosa from a wound. They found that reduced virulence factor and biofilm formation by P. aeruginosa is due to the material surface and porosity, which did not support the growth of this bacteria species. This study suggested that dressing made of wood nanocellulose could be a novel finding to prevent microbe’s growth and promote a moist environment for wound acceleration [166]. In a similar study, wood nanocellulose hydrogel crosslinked with ion (copper or calcium) showed inhibition growth of S. epidermidis and retarded P.aeruginosa biofilm formation [167]. In a different study, chitosan exhibited good antibacterial and antifungal effects towards bacteria (Gram-positive and Gram-negative) and fungi while maintaining its physical properties suggesting it is a good material for wound dressing [168]. Another study using thymol from natural monoterpenoid phenol integrated into wound dressing has demonstrated antibacterial properties and can destroy the biofilm formed by methicillin-sensitive S. aureus [169].
Total contact cast (TCC) is tagged as an alternative treatment approach for DFU. Although the healing rate significantly improved with TCC, yet countless side effects can be life-threatening, such as iatrogenic infections, ulcers, blisters, or skin abrasions [170]. Besides TCC, some other treatments for DFU are maggot debridement therapy (MDT), negative pressure wound therapy (NPWT), and dermaspace systems (DS) [171]. DFU patients who have severe wound infections or osteomyelitis need to undergo antibiotic therapy for at least a month without surgical intervention to resolve the infections as recommended by the Infectious Disease Society of America (IDSA) and the International Working Group on the Diabetic Foot (IWGDF) [58]. Wound dressings are also another choice of treatment for DFU. Some of the available dressings are hydrogel, foam, films, hydrofibers, hydrocolloids, acrylics, and calcium alginates. Cellulose/collagen-based dressing has shown some improvements in diabetic non-wound healing by accelerating the wound healing rate and improvement in wound closure [171]. In addition, the integration of antibacterial properties such as silver nanoparticles into the collagen wound dressing has the potency to reduce the risk of microbial invasions through the broken wound in DFU [172]. Generally, topical antibiotics are not the best choice to treat DFU due to the imbalance of moist production and contact dermatitis [173]. However, a recent finding proved that topical probiotics such as Kefir (cultured probiotic beverage) exhibit potent antimicrobial effects against S. aureus and E.coli. Probiotics help in reducing the possibility of skin infections and improve tissue regeneration by regulating skin microflora through various mechanisms in the skin [174]. Some other examples are tabulated in Table 2. The use of wound dressings alone does not help eliminate pathogens but, together with additional substances that exhibit some antibacterial/antimicrobial properties, could shed some light on treating chronic wounds, especially in DFU.
8. Conclusions and Future Perspectives
Collectively, the conclusions of these research papers has attributed to our knowledge of chronic wounds, the pathophysiology of wound healing, and interactions of normal flora-host depending on the multifaceted conditions of the host. Studies revealed that in healthy individuals, when cells are normal, the overall wound healing phases are well-orchestrated which last only a few days, but in chronic wounds, the majority of cells become debilitated, lost skin integrity, dysfunctional or adverse effects by immune cells to the skin and pathological changes that are difficult to be irreversible. Many underlying factors hamper the wound healing process; one is infection. Current technologies have portrayed a better characterization of normal flora and the potential of these commensals to become pathogen in any opportunistic niches in all stages of wound. The microbial adaptations to the microenvironment of the skin lead to virulence and impaired wound healing. Therefore, it is important to investigate further the role of microbes at the cellular and molecular levels and not only focus on bacteria but also other normal flora such as viruses and fungi to rectify new treatments for chronic wounds. The most challenging part of treating chronic wounds is polymicrobial infections and a high tendency to become resistant to prolonged antibiotic treatments. Loading additional antibacterial substances into the wound dressings, selecting suitable empirical antibiotics based on microbial profile and antimicrobial resistance pattern could reduce the infections and improve wound healing in chronic wounds. However, a deeper understanding of normal and pathological healing will illuminate the interaction between skin cells, normal flora, and their microenvironment. This information might help investigators develop better treatments or methods to eliminate the microbes present in chronic wounds completely.
Author Contributions
Conceptualization, T.M., A.A.N. and M.B.F.; methodology, T.M.; software, T.M.; validation, A.A.N. and M.B.F.; formal analysis, M.B.F.; investigation, resources, data curation and writing—original draft preparation, T.M.; writing—review and editing, A.A.N. and M.B.F.; visualization, T.M.; supervision, M.B.F.; project administration, M.B.F.; funding acquisition, M.B.F. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded through the grant provided by Ministry of Higher Education via Transdisciplinary Research Grant Scheme (TRGS/1/2019/UKM/02/1/3) in the form of database subscription. The funders do not have any contribution and decision to publish or preparation of the manuscript.
Institutional Review Board Statement
This study was approved by the National University of Malaysia (UKM) Research ethics committee with the approval code of UKM PPI/111/8/JEP-2020-152.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
All the authors would like to express our gratitude to the Faculty of Medicine, UKM for the guidance and resources to complete this review.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Han, G.; Ceilley, R. Chronic Wound Healing: A Review of Current Management and Treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Kosaric, N.; Bonham, C.A.; Gurtner, G.C. Wound Healing: A Cellular Perspective. Physiol. Rev. 2019, 99, 665–706. [Google Scholar] [CrossRef] [PubMed]
- Anna, L.-R.; Fabrellas, N.; Saez Rubio, G.; Wilson, K. Time of chronic wound healing, as part of a prevalence and incidence study. Enferm. Glob. 2017, 16, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Mulholland, E.J.; Dunne, N.; Mccarthy, H.O. MicroRNA as Therapeutic Targets for Chronic Wound Healing. Mol. Ther. Nucleic Acids 2017, 8, 46–55. [Google Scholar] [CrossRef] [Green Version]
- Le Goff-Pronost, M.; Bénédicte, M.; Jean-Pierre, B.; Luc, T.; Hervé, B.; Dompmartin, A. Real-World Clinical Evaluation And Costs Of Telemedicine For Chronic Wound Management. Int. J. Technol. Assess. Health Care 2018, 34, 567. [Google Scholar] [CrossRef]
- Brown, M.S.; Ashley, B.; Koh, A. Wearable Technology for Chronic Wound Monitoring: Current Dressings, Advancements, and Future Prospects. Front. Bioeng. Biotechnol. 2018, 6, 47. [Google Scholar] [CrossRef] [PubMed]
- Nussbaum, S.; Carter, M.J.; Fife, C.E.; DaVanzo, J.; Haught, R.; Nusgart, M.; Cartwright, D. An Economic Evaluation of the Impact, Cost, and Medicare Policy Implications of Chronic Nonhealing Wounds. Value Health 2017, 21, 27–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandhu, D.K.S.; Singh, D.K.; Banga, D.R.K.; Sandhu, D.K.S.; Samria, D.J. Role of topical phenytoin (Diphenylhydantoin) dressing in diabetic ulcers: A comparative study with conventional dressing. Int. J. Orthop. Sci. 2018, 4, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Perez-Favila, A.; Martinez-Fierro, M.L.; Rodriguez-Lazalde, J.G.; Cid-Baez, M.A.; Zamudio-Osuna, M.D.J.; Martinez-Blanco, M.; Mollinedo-Montaño, F.E.; Rodriguez-Sanchez, I.P.; Castañeda-Miranda, R.; Garza-Veloz, I. Current therapeutic strategies in diabetic foot ulcers. Medicina 2019, 55, 714. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, D.G.; Boulton, A.J.M.; Bus, S.A. Diabetic Foot Ulcers and Their Recurrence. N. Engl. J. Med. 2017, 376, 2367–2375. [Google Scholar] [CrossRef]
- Saeedi, P.; Salpea, P.; Karuranga, S.; Petersohn, I.; Malanda, B.; Gregg, E.W.; Unwin, N.; Wild, S.H.; Williams, R. Mortality attributable to diabetes in 20–79 years old adults, 2019 estimates: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2020, 162, 108086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, S.; Srivastava, S.; Singh, M.R.; Singh, D. Mechanistic insight into diabetic wounds: Pathogenesis, molecular targets and treatment strategies to pace wound healing. Biomed. Pharmacother 2019, 112, 108615. [Google Scholar] [CrossRef] [PubMed]
- Abdissa, D.; Adugna, T.; Gerema, U.; Dereje, D. Prevalence of Diabetic Foot Ulcer and Associated Factors among Adult Diabetic Patients on Follow-Up Clinic at Jimma Medical Center, Southwest Ethiopia, 2019: An Institutional-Based Cross-Sectional Study. J. Diabetes Res. 2020, 2020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, Z.; Thu, H.E.; Shuid, A.N.; Katas, H.; Hussain, F. Recent Advances in Polymer-based Wound Dressings for the Treatment of Diabetic Foot Ulcer: An Overview of State-of-the-art. Curr. Drug Targets 2017, 19, 527–550. [Google Scholar] [CrossRef]
- Kow, R.Y.; Low, C.L.; Ruben, J.K.; Zaharul-Azri, M.Z.; Lim, B.C. Predictive factors of major lower extremity amputations in diabetic foot infections: A cross-sectional study at district hospital in Malaysia. Malays. Orthop. J. 2019, 13, 45–52. [Google Scholar] [CrossRef]
- Lim, K.P.; Nasruddi, A.B. The effect of individualised glycemic intervention on wound healing rate in diabetic foot ulcer (The EIGIFU study). J. ASEAN Fed. Endocr. Soc. 2018, 33, 22–27. [Google Scholar] [CrossRef] [Green Version]
- Kendall, A.C.; Nicolaou, A. Bioactive lipid mediators in skin inflammation and immunity. Prog. Lipid. Res. 2013, 52, 141–164. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Sellami, M.; Salem, I.; Conic, R.; Kimak, M.; Pigatto, P.D.; Damiani, G. Fasting and Its Impact on Skin Anatomy, Physiology, and Physiopathology: A Comprehensive Review of the Literature. Nutrients 2019, 11, 249. [Google Scholar] [CrossRef] [Green Version]
- Thulabandu, V.; Chen, D.; Atit, R.P. Dermal fibroblast in cutaneous development and healing. Wiley Interdiscip. Rev. Dev. Biol. 2018, 7, 1–13. [Google Scholar] [CrossRef]
- Luxenburg, C.; Zaidel-bar, R. From cell shape to cell fate via the cytoskeleton—Insights from the epidermis. Exp. Cell Res. 2019, 378, 232–237. [Google Scholar] [CrossRef]
- Nguyen, A.V.; Soulika, A.M. The Dynamics of the Skin’s Immune System. Int. J. Mol. Sci. 2019, 20, 1811. [Google Scholar] [CrossRef] [Green Version]
- Coger, V.; Million, N.; Rehbock, C.; Sures, B.; Nachev, M.; Barcikowski, S.; Wistuba, N.; Strauß, S.; Vogt, P.M. Tissue Concentrations of Zinc, Iron, Copper, and Magnesium during the Phases of Full Thickness Wound Healing in a Rodent Model. Biol. Trace Elem. Res. 2019, 191, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.; Sharp, A. Undertaking a person-centred assessment of patients with chronic wounds. Nurs. Stand. 2019, 34, 77–82. [Google Scholar] [CrossRef] [Green Version]
- Netam, A.K.; Prasad, J.; Satapathy, T. A Review on Ethnopharmacological approaches to Wound Healing and Curative Progression. J. Pharm. Pharmacodyn. 2019, 11, 37–45. [Google Scholar] [CrossRef]
- Vilar, R.; Fish, R.J.; Casini, A.; Neerman-arbez, M. Fibrinogen in human disease: Both friend and foe. Haematologica 2020, 105, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Rittié, L. Cellular mechanisms of skin repair in humans and other mammals. J. Cell Commun. Signal. 2016, 10, 103–120. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, H.N.; Hardman, M.J.; Wilkinson, H.N. Wound healing: Cellular mechanisms and pathological outcomes. Open Biol. 2020, 10. [Google Scholar] [CrossRef]
- Sabau, M.; Simona, B.; Camelia, L.B.; Gheorghe, C.; Lucia-georgeta, D.; Claudia, T.J.; Bogdan, A.B.; Claudia, M.J.; Cristian, M.D.; Delia, M.T. Legal Medicine Implications in Fibrinolytic Therapy of Acute Ischemic Stroke. BMC Med. Ethics 2019, 20, 1–9. [Google Scholar] [CrossRef]
- von Köckritz-blickwede, M.; Blodkamp, S.; Nizet, V. Interaction of Bacterial Exotoxins with Neutrophil Extracellular Traps: Impact for the Infected Host. Front. Microbiol. 2016, 7, 1–8. [Google Scholar] [CrossRef]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef] [Green Version]
- Naseri, S.; Lepry, W.C.; Nazhat, S.N. Bioactive glasses in wound healing: Hope or hype? J. Mater. Chem. B 2017, 5, 6167–6174. [Google Scholar] [CrossRef] [PubMed]
- Qing, C. The molecular biology in wound healing & non-healing wound. Chin. J. Traumatol. 2017, 20, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Huang, B.; Horng, H.; Yeh, C. Wound healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Yang, C.; Lin, Y.; Parviz, Y.; Sun, K.; Wang, W.; Ren, M.; Yan, L. Matrix metalloproteinase 9 induces keratinocyte apoptosis through FasL/Fas pathway in diabetic wound. Apoptosis 2019, 24, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.M.U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, S.; Heljasvaara, R.; Gullberg, D.; Tasanen, K.; Pihlajaniemi, T. Toward understanding scarless skin wound healing and pathological scarring. F1000Research 2019, 8, 787. [Google Scholar] [CrossRef] [Green Version]
- Thiruvoth, F.; Mohapatra, D.; Sivakumar, D.; Chittoria, R.; Nandhagopal, V. Current concepts in the physiology of adult wound healing. Plast. Aesthetic Res. 2015, 2, 250. [Google Scholar] [CrossRef] [Green Version]
- Shinde, A.V.; Humeres, C.; Frangogiannis, N.G. The role of α-smooth muscle actin in fibroblast - mediated matrix contraction and remodelling. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 298–309. [Google Scholar] [CrossRef]
- Fu, L.; Chen, X.; Cai, M.; Tao, X.; Fan, Y.; Fan, Y. Surface Engineered Metal-Organic Frameworks (MOFs) Based Novel Hybrid Systems for Effective Wound Healing: A Review of Recent Developments. Front. Bioeng. Biotechnol. 2020, 8, 1053. [Google Scholar] [CrossRef]
- Ellis, S.; Lin, E.J.; Tartar, D. Immunology of Wound Healing. Curr. Dermatol. Rep. 2018, 7, 350–358. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.; Liang, H.; Clarke, E.; Jackson, C.; Xue, M. Inflammation in Chronic Wounds. Int. J. Mol. Sci. 2016, 17, 2085. [Google Scholar] [CrossRef]
- Larouche, J.; Sheoran, S.; Maruyama, K.; MMikael, M. Immune Regulation of Skin Wound Healing: Mechanisms and Novel Therapeutic Targets. Adv. Wound Care 2018, 7, 209–231. [Google Scholar] [CrossRef]
- Dai, C.; Shih, S.; Khachemoune, A. Skin substitutes for acute and chronic wound healing: An updated review. J. Dermatol. Treat. 2018, 31, 639–648. [Google Scholar] [CrossRef]
- Worthen, C.A.; Cui, Y.; Orringer, J.S.; Johnson, T.M.; Voorhees, J.J.; Fisher, G.J. CD26 Identifies a Subpopulation of Fibroblasts that Produce the Majority of Collagen during Wound Healing in Human Skin. J. Investig. Dermatol. 2020, 140, 2515–2524. [Google Scholar] [CrossRef]
- Dickinson, L.E.; Gerecht, S. Engineered biopolymeric scaffolds for chronic wound healing. Front. Physiol. 2016, 7, 341. [Google Scholar] [CrossRef] [Green Version]
- Demidova-Rice, T.N.; Hamblin, M.R.; Herman, I.M. Acute and impaired wound healing: Pathophysiology and current methods for drug delivery, part 2: Role of growth factors in normal and pathological wound healing: Therapeutic potential and methods of delivery. Adv. Ski. Wound Care 2012, 25, 349–370. [Google Scholar] [CrossRef] [Green Version]
- Karr, J.C. Bilayered skin-substitute technology for the treatment of diabetic foot ulcers: Current insights. Chronic Wound Care Manag. Res. 2016, 4, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Hariono, M.; Yuliani, S.H.; Istyastono, E.P.; Riswanto, F.D.O.; Adhipandito, C.F. Matrix Metalloproteinase 9 (MMP9) in Wound Healing of Diabetic Foot Ulcer: Molecular Target and Structure-Based Drug Design. Wound Med. 2018, 22, 1–13. [Google Scholar] [CrossRef]
- Sahle, F.F.; Gebre-mariam, T.; Dobner, B.; Wohlrab, J.; HHNeubert, R. Skin Diseases Associated with the Depletion of Stratum Corneum Lipids and Stratum Corneum Lipid Substitution Therapy. Ski. Pharmacol. Physiol. 2015, 28, 42–55. [Google Scholar] [CrossRef]
- Moses, R.L.; Boyle, G.M.; Howard-jones, R.A.; Rachel, J.E.; Johns, J.P.; Gordon, V.; Reddell, P.; Steadman, R.; Moseley, R. Novel epoxy-tiglianes stimulate skin keratinocyte wound healing responses and re-epithelialization via protein kinase C activation. Biochem. Pharmacol. 2020, 178, 114048. [Google Scholar] [CrossRef]
- Liarte, S.; Bernabé-Garcia, Á.; Nicolás, F.J. Role of TGF- β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Berksoy Hayta, S.; Durmuş, K.; Elif Altuntaş, E.; Yıldız, E.; Hisarcıklıo, M.; Akyol, M. The reduction in inflammation and impairment in wound healing by using strontium chloride hexahydrate. Cutan. Ocul. Toxicol. 2017, 37, 24–48. [Google Scholar] [CrossRef]
- Rikabi AHAAl Tobin, D.J.; Suman, K.R.; Thornton, M.J. Dermal fibroblasts cultured from donors with type 2 diabetes mellitus retain an epigenetic memory associated with poor wound healing responses. Sci. Rep. 2021, 11, 1–13. [Google Scholar] [CrossRef]
- Okonkwo, U.A.; Dipietro, L.A. Diabetes and wound angiogenesis. Int. J. Mol. Sci. 2017, 18, 1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, S.C.; Chhatbar, K.C.; Kashikar, A.; Mehndiratta, A. Diabetic Foot. BMJ 2017, 359, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.; Kang, H.J.; Jeon, J.H.; Kim, M.J.; Lee, I.K. Recent advances in the pathogenesis of microvascular complications in diabetes. Arch. Pharm. Res. 2019, 42, 252–262. [Google Scholar] [CrossRef]
- Jneid, J.; Lavigne, J.P.; Scola BLa Cassir, N. The Diabetic Foot Microbiota: A Review. Hum. Microbiome J. 2017, 5–6, 1–6. [Google Scholar] [CrossRef]
- Barwell, N.D.; Devers, M.C.; Kennon, B.; Hopkinson, H.E.; McDougall, C.; Young, M.J.; Robertson, H.M.A.; Stang, D.; Dancer, S.J.; Seaton, A.; et al. Diabetic foot infection: Antibiotic therapy and good practice recommendations. Int. J. Clin. Pract. 2017, 71, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Grigoropoulou, P.; Eleftheriadou, I.; Jude, E.B.; Tentolouris, N. Diabetic Foot Infections: An Update in Diagnosis and Management. Curr. Diabetes Rep. 2017, 17. [Google Scholar] [CrossRef]
- Khan, R.; Ooi, X.Y.; Parvus, M.; Valdez, L. Advanced Glycation End Products: Formation, Role in Diabetic Complications, and Potential in Clinical Applications. Eye Foot Diabetes 2020, 3. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.M. Diabetic Foot Ulcer: Synopsis of the Epidemiology and Pathophysiology. Int. J. Diabetes Endocrinol. 2018, 3, 23–28. [Google Scholar] [CrossRef] [Green Version]
- Syafril, S. Pathophysiology diabetic foot ulcer. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2018; Volume 125, p. 12161. [Google Scholar] [CrossRef]
- Chen, P.; He, G.; Qian, J.; Zhan, Y.; Xiao, R. Potential role of the skin microbiota in Inflammatory skin diseases. J. Cosmet. Dermatol. 2021, 20, 400–409. [Google Scholar] [CrossRef]
- Maguire, M.; Maguire, G. The role of microbiota, and probiotics and prebiotics in skin health. Arch. Dermatol. Res. 2017, 309, 411–421. [Google Scholar] [CrossRef]
- Hong, B.; Winkel, A.; Ertl, P.; Stumpp, S.N.; Schwabe, K.; Stiesch, M.; Krauss, J.K. Bacterial colonisation of suture material after routine neurosurgical procedures: Relevance for wound infection. Acta. Neurochir. 2017, 160, 497–503. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chen, T.H.; Butcher, A.M.; Trzoss, L.L.; Nam, S.J.; Shirakawa, K.T.; Zhou, W.; Oh, J.; Otto, M.; Fenical, W.; et al. A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci. Adv. 2018, 4, eaao4502. [Google Scholar] [CrossRef] [Green Version]
- Jasim, H.A. Study of Type of Bacteria That Found in Ear, Mouth, Nose of Second Stage Nursing Department Student. Indian J. Forensic Med. Toxicol. 2020, 14, 2789–2793. [Google Scholar]
- Gautam, V.; Sethuraman, N.; Kaur, R.; Sachdev, S.; Marwaha, N.; Ray, P. Changing Epidemiology of Coagulase–Negative Staphylococci in Normal Flora of Skin. Indian J. Med. Microbiol. 2017, 35, 277–278. [Google Scholar] [CrossRef]
- Shami, A.; Al-mijalli, S.; Pongchaikul, P.; Al-barrag, A.; Abdurahim, S. The prevalence of the culturable human skin aerobic bacteria in Riyadh, Saudi Arabia. BMC Microbiol. 2019, 19, 189. [Google Scholar] [CrossRef]
- Chen, Y.E.; Fischbach, M.A.; Belkaid, Y. Skin microbiota—Host interactions. Nature 2018, 553, 427–436. [Google Scholar] [CrossRef]
- Stegli, A.; Jachowicz, A.; Justyna, S.; Szulc, J.; Adamiak, J.; Otlewska, A.; Pielech-Przybylska, K.; Gutarowska, B. Factors Influencing Microbiological Biodiversity of Human Foot Skin. Int. J. Environ. Res. Public Health 2019, 16, 3503. [Google Scholar] [CrossRef] [Green Version]
- Cundell, A.M. Microbial Ecology of the Human Skin. Microb. Ecol. 2016, 76, 113–120. [Google Scholar] [CrossRef]
- Haro, C.; Rangel-zú, O.A.; Alcalá-díaz, J.F.; Delgado, F.G.; Martínez, P.P.; Lista, J.D.; Navarro, G.M.Q.; Landa, B.B.; Cortés, J.A.N.; Sempere, M.T.; et al. Intestinal Microbiota Is Influenced by Gender and Body Mass Index. PLoS ONE. 2016, 11, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.; Zeng, D.N.; Chi, L.; Tan, Y.; Galzote, C.; Cardona, C.; Lax, S.; Gilbert, J.; Quan, Z.X. The influence of age and gender on skin-associated microbial communities in urban and rural human populations. PLoS ONE. 2015, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bassegoda, A.; Ivanova, K.; Ramon, E.; Tzanov, T. Strategies to prevent the occurrence of resistance against antibiotics by using advanced materials. Appl. Microbiol. Biotechnol. 2018, 5, 2075–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustamante, M.; Oomah, B.D.; Oliveira, W.P.; Burgos-díaz, C.; Rubilar, M.; Shene, C. Probiotics and prebiotics potential for the care of skin, female urogenital tract, and respiratory tract. Folia. Microbiol 2020, 65, 245–264. [Google Scholar] [CrossRef] [Green Version]
- Rainer, B.M.; Thompson, K.G.; Antonescu, C.; Florea, L.; Mongodin, E.F.; Bui, J.; Fischer, A.H.; Pasieka, H.B.; Garza, L.A.; Kang, S.C.; et al. Characterization and Analysis of the Skin Microbiota in Rosacea: A Case—Control Study. Am. J. Clin. Dermatol. 2019, 21, 139–147. [Google Scholar] [CrossRef]
- Chien, A.; Tsai, J.; Leung, S.; Mongodin, E.F.; Nelson, A.M.; Kang, S.; Garza, L.A. Association of Systemic Antibiotic Treatment of Acne With Skin Microbiota Characteristics. JAMA Dermatol. 2019, 155, 425–434. [Google Scholar] [CrossRef]
- Lehtimäki, J.; Karkman, A.; Laatikainen, T.; Laura, P.H.; Leena, V.H.; Tari, H.; Hanski, I.; Ruokolainen, L. Patterns in the skin microbiota differ in children and teenagers between rural and urban environments. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fourni, M.; Latire, T.; Souak, D.; Feuilloley, M.G.J.; Bedoux, G. Staphylococcus epidermidis and Cutibacterium acnes Two Major Sentinels of Skin Microbiota and the Influence of Cosmetics. Microorganisms 2020, 8, 1752. [Google Scholar] [CrossRef]
- Wallen-russell, C.; Wallen-russell, S. Meta-Analysis of Skin Microbiome: New Link between Skin Microbiota Diversity and Skin Health with Proposal to Use This as a Future Mechanism to Determine Whether Cosmetic Products Damage. Cosmetics 2017, 4, 14. [Google Scholar] [CrossRef] [Green Version]
- Sakr, A.; Brégeon, F.; Mège, J.; Rolain, J.; Gobert, A.P. Staphylococcus aureus Nasal Colonization: An Update on Mechanisms, Epidemiology, Risk Factors, and Subsequent Infections. Front. Microbiol. 2018, 9, 2419. [Google Scholar] [CrossRef]
- Proctor, D.M.; Relman, D.A. The Landscape Ecology and Microbiota of the Human Nose, Mouth, and Throat. Cell Host Microbe 2017, 21, 421–432. [Google Scholar] [CrossRef] [Green Version]
- Sanmiguel, A.; Grice, E.A. Interactions between host factors and the skin microbiome. Cell Mol. Life Sci. 2015, 72, 1499–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155. [Google Scholar] [CrossRef]
- Sanford, J.A.; Gallo, R.L. Functions of the Skin Microbiota in Health and Disease. Seminars in Immunology; Academic Press: Cambridge, MA, USA, 2013; Volume 25, pp. 370–377. [Google Scholar] [CrossRef] [Green Version]
- Szabo, K.; Erdei, L.; Bolla, B.S.; Tax, G.; Biro, T.; Kemeny, L. Factors shaping the composition of the cutaneous micobiota. Br. J. Dermatol. 2016, 176, 344–351. [Google Scholar] [CrossRef] [Green Version]
- Laborel-préneron, E.; Bianchi, P.; Boralevi, F.; Lehours, P. Effects of the Staphylococcus aureus and Staphylococcus epidermidis Secretomes Isolated from the Skin Microbiota of Atopic Children on CD4 + T Cell Activation. PLoS ONE 2015, 10, e0141067. [Google Scholar] [CrossRef] [Green Version]
- Claudel, J.; Auffret, N.; Leccia, M. Staphylococcus epidermidis: A Potential New Player in the Physiopathology of Acne ? Dermatology 2019, 235, 287–294. [Google Scholar] [CrossRef]
- Otto, M. Staphylococci in the human microbiome: The role of host and interbacterial interactions. Curr. Opin. Microbiol. 2020, 53, 71–77. [Google Scholar] [CrossRef]
- Ellis, S.R.; Nguyen, M.; Vaughn, A.R.; Notay, M.; Burney, W.A.; Sandhu, S.; Sivamani, R.K. The skin and gut microbiome and its role in common dermatologic conditions. Microorganisms 2019, 7, 550. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Glatz, M.; Horiuchi, K.; Kawasaki, H.; Akiyama, H.; Kaplan, D.H.; Kong, H.H.; Amagai, M.; Nagao, K. Dysbiosis and Staphyloccus aureus Colonization Drives Inflammation in Atopic Dermatitis. Immunity 2015, 42, 756–766. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.J.; Lee, H.K. The role of skin and orogenital microbiota in protective immunity and chronic immune-mediated inflammatory disease. Front. Immunol. 2018, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Coates, M.; Blanchard, S.; MacLeod, A.S. Innate antimicrobial immunity in the skin: A protective barrier against bacteria, viruses, and fungi. PLoS Pathog. 2018, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.R.; Belinda, I.G.; Mcintyre, M.K.; Dubick, M.A.; Christy, R.J.; Nicholson, S.E.; Burmeister, D.M. The Cutaneous Microbiome and Wounds: New Molecular Targets to Promote Wound Healing. Int. J. Mol. Sci. 2018, 19, 2699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Findley, K.; Oh, J.; Yang, J.; Conlan, S.; Deming, C.; Meyer, J.A.; Schoenfeld, D.; Nomicos, E.; Park, M.; Kong, H.H.; et al. Human Skin Fungal Diversity. Nature 2013, 498, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Jr, T.L.D. Host—Microbe interactions: Malassezia and human skin. Curr. Opin. Microbiol. 2017, 40, 81–87. [Google Scholar] [CrossRef]
- Limon, J.J.; Skalski, J.H.; Underhill, D.M. Review Commensal Fungi in Health and Disease. Cell Host Microbe 2017, 22, 156–165. [Google Scholar] [CrossRef]
- Jo, H.J.; Deming, C.; Kennedy, E.; Conlan, S.; Polley, E.C.; Ng, W.I.; Segre, J.A.; Kong, H.H. Diverse human skin fungal communities in children converge in adulthood. J. Investig. Dermatol. 2017, 136, 2356–2363. [Google Scholar] [CrossRef] [Green Version]
- Scheffold, A.; Bacher, P.; Landmann, S.L. T cell immunity to commensal fungi. Curr. Opin. Microbiol. 2020, 58, 116–123. [Google Scholar] [CrossRef]
- Schommer, N.N.; Gallo, R.L. Structure and function of the human skin microbiome. Trends Microbiol. 2013, 21, 660–668. [Google Scholar] [CrossRef] [Green Version]
- Strickley, J.D.; Messerschmidt, J.L.; Awad, M.E.; Li, T.; Hasegawa, T.; Ha, D.T.; Nabeta, H.W.; Bevins, P.A.; Ngo, K.H.; Asgari, M.M.; et al. Immunity to commensal papillomaviruses protects against skin cancer. Nature 2019, 575, 519–522. [Google Scholar] [CrossRef]
- Lambert, P.F.; Münger, K.; Roesl, F.; Hasche, D.; Tommasino, M. Beta human papillomaviruses and skin cancer. Nature 2020, 588, E20–E23. [Google Scholar] [CrossRef]
- Sichero, L.; Nunes, E.M. Epidemiology and biology of cutaneous human papillomavirus. Clinics 2018, 73. [Google Scholar] [CrossRef]
- Wang, W.M.; Jin, H.Z. Skin Microbiome: An Actor in the Pathogenesis of Psoriasis. Chin. Med. J. 2018, 131, 95. [Google Scholar] [CrossRef]
- Leonel, C.; Sena, I.F.G.; Silva, W.N.; Prazeres PHD, M.; Fernandes, G.R.; Agresti, P.M.; Drumond, M.M.; Mintz, A.; Azevedo, V.A.C.; Birbrair, A. Staphylococcus epidermidis role in the skin microenvironment. J. Cell Mol. Med. 2019, 23, 5949–5955. [Google Scholar] [CrossRef] [Green Version]
- Belkaid, Y.; Segre, J.A. Dialogue between skin microbiota and immunity. Science 2014, 346, 954–959. [Google Scholar] [CrossRef]
- Bowler, P.G.; Duerden, B.I.; Armstrong, D.G. Wound Microbiology and Associated Approaches to Wound Management. Clin. Microbiol Rev. 2001, 14, 244–269. [Google Scholar] [CrossRef] [Green Version]
- Choi, E.H. Gender, Age, and Ethnicity as Factors That Can Influence Skin pH. pH Ski. Issues Chall. 2018, 54, 48–53. [Google Scholar] [CrossRef]
- Lee, C.S.; Jamil, A. Skin pH and its Relationship with Transepidermal Water Loss and Disease Severity in Children with Atopic Dermatitis: A Cross-Sectional Study. J. Dermatol. Dermatol. Surg. 2020, 24, 84. [Google Scholar] [CrossRef]
- Blaak, J.; Staib, P. The Relation of pH and Skin Cleansing. Issues Chall. 2018, 54, 132–142. [Google Scholar] [CrossRef]
- Rippke, F.; Berardesca, E.; Weber, T.M. pH and Microbial Infections. Issues Chall. 2018, 54, 87–94. [Google Scholar] [CrossRef]
- Bennison, L.R.; Miller, C.N.; Summers, R.J.; Minnis, A.M.B.; Sussman, G.; McGuiness, W. The pH of wounds during healing and infection: A descriptive literature review. J. Aust. Wound Manag. Assoc. 2017, 25, 63–69. [Google Scholar] [CrossRef]
- Kruse, C.R.; Singh, M.; Targosinski, S.; Sørensen, J.A.; Eriksson, E.; Nuutila, K. The effect of pH on cell viability, cell migration, cell proliferation, wound closure and wound re-epithelialization: In vitro and in vivo study. Wound Repair Regen. 2017, 25, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Power, G.; Moore, Z.; O’Connor, T. Measurement of pH, exudate composition and temperature in wound healing: A systematic review. J. Wound Care 2017, 26, 381–397. [Google Scholar] [CrossRef]
- Nakagami, G.; Sanada, H.; Iizaka, S.; Kadono, T.; Higashino, T.; Koyanagi, H.; Haga, N. Predicting delayed pressure ulcer healing using thermography: A prospective cohort study. J. Wound Care 2010, 19, 465–472. [Google Scholar] [CrossRef]
- Moskovicz, V.; Gross, A.; Mizrahi, B. Extrinsic Factors Shaping the Skin Microbiome. Microorganisms 2020, 8, 1023. [Google Scholar] [CrossRef]
- Duckney, P.; Wong, H.K.; Serrano, J.; Yaradou, D.; Oddos, T.; Stamatas, G.N. The role of the skin barrier in modulating the effects of common skin microbial species on the inflammation, differentiation and proliferation status of epidermal keratinocytes. BMC Res. Notes 2013, 6, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Sanchez Lagunes, R.; Phillips, T.; Suwanpradid, J.; MacLeod, A. IL-27 facilitates skin wound healing through induction of epidermal proliferation and host defense. J. Investig. Dermatol. 2017, 136, S130. [Google Scholar] [CrossRef]
- Stacy, A.; Belkaid, Y. Microbial guardians of skin health. Science 2019, 363, 227–228. [Google Scholar] [CrossRef]
- Tsiouris, C.G.; Tsiouri, M.G. Human microflora, probiotics and wound healing. Wound Med. 2017, 19, 33–38. [Google Scholar] [CrossRef]
- Chen, L.; Dipietro, L.A. Toll-Like Receptor Function in Acute Wounds. Adv. Wound Care 2017, 6, 344–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, L.; Kragh, K.N.; Eickhardt, S.R.; Poulsen, S.S.; Gjerdrum, L.M.R.; Ghathian, K.; Calum, H.; Magnus, S.A.; Bjarnsholt, T. Bacterial Aggregates Establish at the Edges of Acute Epidermal Wounds. Adv. Wound Care 2018, 00, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tomic, M.; Jamie, C.; Katelyn, L.B.; Natasa, E.O.N.; Irena, S. Skin Microbiota and its Interplay with Wound Healing. Am. J. Clin. Dermatol. 2020, 21, 36–43. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Y.; Zhou, L.; Liu, M.; Liang, G.; Yan, R.; Jiang, Y.; Hao, J.; Zhang, X.; Hu, X.; et al. V γ 4 T cells inhibit the Pro-healing Functions of Dendritic epidermal T cells to Delay skin Wound closure Through IL-17A. Front. Immunol. 2018, 9, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Hsia, H.C. The impact of microbial communities on wound healing: A review. Ann. Plast Surg. 2018, 81, 113–123. [Google Scholar] [CrossRef]
- Alavi, S.M.; Khosravi, A.D.; Abdullah, S.; Dashtebozorg, A.; Montazeri, E.A. Bacteriologic Study of Diabetic Foot Ulcer. Pak. J. Med. Sci. 2007, 23, 681–684. [Google Scholar] [CrossRef] [Green Version]
- Rahim, K.; Saleha, S.; Zhu, X.; Huo, L.; Basit, A. Bacterial Contribution in Chronicity of Wounds. Microb. Ecol. 2017, 73, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [Green Version]
- Flemming, H.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef]
- Matilla, M.A.; Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42, fux052. [Google Scholar] [CrossRef] [Green Version]
- Malone, M.; Bjarnsholt, T.; Mcbain, A.J.; James, G.A.; Stoodley, P.; Leaper, D.; Tachi, M.; Schultz, G.; Swanson, T.; Wolcott, R.D. The prevalence of biofilms in chronic wounds: A systematic review and meta- analysis of published data. J. Wound Care 2017, 26, 20–25. [Google Scholar] [CrossRef] [Green Version]
- Omar, A.; Wright, J.; Schultz, G.; Burrell, R.; Nadworny, P. Microbial Biofilms and Chronic Wounds. Microorganisms 2017, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Nanoukon, C.; Argemi, X.; Sogbo, F.; Orekan, J.; Keller, D.; Affolabi, D.; Schramm, F.; Riegel, P.; Baba-moussa, L.; Prevost, G. Pathogenic features of clinically significant coagulase-negative staphylococci in hospital and community infections in Benin. Int. J. Med. Microbiol. 2016, 307, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Rathnayake, I.U.; Huygens, F. Prevalence of biofilm controlling ica genes of Staphylococcus epidermidis detected in healthy skin, blood samples from septicaemia patients and chronic wounds. Int. J. Basic Clin. Pharmacol. 2017, 6, 726–733. [Google Scholar] [CrossRef] [Green Version]
- Le, K.Y.; Park, M.D.; Otto, M. Immune Evasion Mechanisms of Staphylococcus epidermidis Biofilm Infection. Front. Microbiol. 2018, 9, 359. [Google Scholar] [CrossRef]
- Kalan, L.R.; Brennan, M.B. The role of the microbiome in nonhealing diabetic wounds. Ann. N. Y. Acad. Sci. 2019, 1435, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Wei, Q.; Zhang, Z.; Luo, J.; Kong, J.; Ding, Y.; Chen, Y.; Wang, K. Insulin treatment enhances pseudomonas aeruginosa biofilm formation by increasing intracellular cyclic di-GMP levels, leading to chronic wound infection and delayed wound healing. Am. J. Transl. Res. 2019, 11, 3261. [Google Scholar]
- Rafail, S.; Kourtzelis, I.; Foukas, P.G.; Markiewski, M.M.; Deangelis, R.A.; Guariento Mara Ricklin, D.; Grice, E.A.; Lambris, J.D. Complement Deficiency Promotes Cutaneous Wound Healing in Mice. J. Immunol. 2015, 194, 1285–1292. [Google Scholar] [CrossRef] [Green Version]
- Vlaicu, S.I.; Tatomir, A.; Boodhoo, D.; Vesa, S. The role of complement system in adipose tissue-related inflammation. Immunol. Res. 2016, 64, 653–664. [Google Scholar] [CrossRef]
- vor De, L.; Rooijakkers, S.H.M.; van Strijp, J.A.G. Staphylococci evade the innate immune response by disarming neutrophils and forming biofilms. FEBS Lett. 2020, 594, 2556–2569. [Google Scholar] [CrossRef] [Green Version]
- Berends, E.T.M.; Mohan, S.; Miellet, W.R.; Ruyken, M.; Rooijakkers, S.H.M. Contribution of the complement Membrane Attack Complex to the bactericidal activity of human serum. Mol. Immunol. 2015, 65, 328–335. [Google Scholar] [CrossRef]
- Doni, A.; D’Amico, G.; Morone, D.; Mantovani, A.; Garlanda, C. Humoral innate immunity at the crossroad between microbe and matrix recognition: The role of PTX3 in tissue damage. Semin. Cell Dev. Biol. 2016, 61, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.R.; Diegelmann, R.F. What Makes Wounds Chronic. Surg. Clin. N. Am. 2020, 100, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Lazaro, J.L.; Izzo, V.; Meaume, S.; Davies, A.H.; Lobmann, R.; Uccioli, L. Elevated levels of matrix metalloproteinases and chronic wound healing: An updated review of clinical evidence. J. Wound Care 2016, 25, 277–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, M.; Berends, E.T.M.; Chan, R.; Schwab, E.; Roy, S.; Sen, C.K. Staphylococcus aureus biofilms release leukocidins to elicit extracellular trap formation and evade neutrophil-mediated killing. Proc Natl. Acad. Sci. USA 2018, 115, 7416–7421. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M. Cooperative Evolutionary Strategy between the Bacteriome and Mycobiome. MBio 2016, 7, e01951-16. [Google Scholar] [CrossRef] [Green Version]
- Kirchner, S.; Lei, V. The Cutaneous Wound Innate Immunological Microenvironment. Int. J. Mol. Sci. 2020, 21, 16. [Google Scholar] [CrossRef]
- Behm, B.; Babilas, P.; Landthaler, M.; Schreml, S. Cytokines, chemokines and growth factors in wound healing. J. Eur. Acad. Dermatol. Venereol. 2011, 26, 812–820. [Google Scholar] [CrossRef]
- Kwock, J.T.; Handfield, C.; Suwanpradid, J.; Hoang, P.; Mcfadden, M.J.; Labagnara, K.F.; Floyd, L.; Shannon, J.; Uppala, R.; Sarkar, M.K.; et al. IL-27 signaling activates skin cells to induce innate antiviral proteins and protects against Zika virus infection. Sci. Adv. 2020, 6, eaay3245. [Google Scholar] [CrossRef] [Green Version]
- Lei, V.; Petty, A.J.; Atwater, A.R.; Wolfe, S.A.; Macleod, A.S. Skin Viral Infections: Host Antiviral Innate Immunity and Viral Immune Evasion. Front. Immunol. 2020, 11, 2791. [Google Scholar] [CrossRef]
- Park, J.; Oh, B.; Lee, J.P.; Choi, M.; Lee, M.; Kim, B. Influence of Microbiota on Diabetic Foot Wound in Comparison with Adjacent Normal Skin Based on the Clinical Features. Biomed. Res. Int. 2019, 2019, 10. [Google Scholar] [CrossRef]
- Piya, M.K.; McTernan, P.G.; Kumar, S. Adipokine inflammation and insulin resistance: The role of glucose, lipids and endotoxin. J. Endocrinol. 2013, 216, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Cai, Y.; Meng, C.; Ding, X.; Huang, J.; Luo, X. The role of the microbiome in diabetes mellitus. Diabetes Res. Clin. Pract. 2020, 172. [Google Scholar] [CrossRef]
- Leaper, D.; Assadian, O.; Edmiston, C.E. Approach to chronic wound infections. Br. J. Dermatol. 2015, 173, 351–358. [Google Scholar] [CrossRef]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef] [Green Version]
- Dong, J.; Chen, L.; Zhang, Y.; Jayaswal, N. Mast Cells in Diabetes and Diabetic Wound Healing. Adv. Ther. 2020, 37, 4519–4537. [Google Scholar] [CrossRef]
- Boniakowski, A.E.; Kimball, A.S.; Benjamin, N.; Kunkel, S.L.; Gallagher, K.A. Macrophage-Mediated Inflammation in Normal and Diabetic Wound Healing. J. Immunol. 2017, 199, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Moura, J.; Rodrigues, J.; Gonçalves, M.; Amaral, C.; Lima, M.; Carvalho, E. Impaired T-cell differentiation in diabetic foot ulceration. Cell Mol. Immunol. 2017, 14, 758–769. [Google Scholar] [CrossRef] [Green Version]
- Santiago-rodriguez, T.M.; Hollister, E.B. Human Virome and Disease: High-Throughput Phage-Bacteria Dysbiosis and Development of Human Gut. Viruses 2019, 11, 656. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Vatanen, T.; Droit, L.; Park, A.; Kostic, A.D.; Poon, T.W.; Vlamakis, H.; Siljander, H.; Hämäläinen, A.M.; Peet, A.; et al. Intestinal virome changes precede autoimmunity in type I diabetes-susceptible children. Proc. Natl. Acad. Sci. USA 2018, 115, E11426. [Google Scholar] [CrossRef] [Green Version]
- Canesso, M.C.C.; Vieira, A.T.; Castro, T.B.R.; Schirmer, B.G.A.; Cisalpino, D.; Martins, F.S.; Rachid, M.A.; Nicoli, J.R.; Teixeira, M.M.; Barcelos, L.S. Skin Wound Healing Is Accelerated and Scarless in the Absence of Commensal Microbiota. J. Immunol. 2014, 193, 5171–5180. [Google Scholar] [CrossRef] [Green Version]
- Tchero, H.; Kangambega, P.; Noubou, L.; Becsangele, B.; Fluieraru, S.; Teot, L. Antibiotic therapy of diabetic foot infections: A systematic review of randomized controlled trials. Wound Repair. Regen. 2018, 26, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.T.; Armstrong, D.G. Microbiology and antimicrobial therapy for diabetic foot infections. Infect. Chemother. 2018, 50, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Jack, A.A.; Nordli, H.R.; Powell, L.C.; Powell, K.A.; Kishnani, H.; Johnsen, P.O.; Pukstad, B.; Thomas, D.W.; Chinga, C.G.; Hill, K.E. The interaction of wood nanocellulose dressings and the wound pathogen P. aeruginosa. Carbohydr. Polym. 2017, 157, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Heitz, K.; Strømme, M.; Welch, K.; Ferraz, N. Ion-crosslinked wood-derived nanocellulose hydrogels with tunable antibacterial properties: Candidate materials for advanced wound care applications. Carbohydr. Polym. 2018, 181, 345–350. [Google Scholar] [CrossRef]
- Matica, M.A.; Aachmann, F.L.; Tøndervik, A.; Sletta, H.; Ostafe, V. Chitosan as a wound dressing starting material: Antimicrobial properties and mode of action. Int. J. Mol. Sci. 2019, 20, 5889. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Salinas, S.; Gámez, E.; Landa, G.; Arruebo, M.; Irusta, S.; Mendoza, G. Antimicrobial Wound Dressings against Fluorescent and Methicillin-Sensitive Intracellular Pathogenic Bacteria. ACS Appl. Mater. Interfaces 2020, 12, 51302–51313. [Google Scholar] [CrossRef]
- Messenger, G.; Masoetsa, R.; Hussain, I. A Narrative Review of the Benefits and Risks of Total Contact Casts in the Management of Diabetic Foot Ulcers. J. Am. Coll Clin. Wound Spec. 2017, 9, 19–23. [Google Scholar] [CrossRef]
- Naomi, R.; Fauzi, M.B. Cellulose/collagen dressings for diabetic foot ulcer: A review. Pharmaceutics 2020, 12, 881. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Razip Wee, M.F.M.; Tabata, Y.; Idrus, R.B.H.; Nordin, A.; Fauzi, M.B. Antibacterial-integrated collagen wound dressing for diabetes-related foot ulcers: An evidence-based review of clinical studies. Polymers 2020, 12, 2168. [Google Scholar] [CrossRef]
- Ramirez-acuña, J.M.; Cardenas-cadena, S.A.; Marquez-salas, P.A.; Garza-veloz, I.; Perez-favila, A.; Cid-baez, M.A. Diabetic Foot Ulcers: Current Advances in Antimicrobial Therapies and Emerging Treatments. Antibiotics 2019, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Knackstedt, R.; Knackstedt, T.; Gatherwright, J. The role of topical probiotics on wound healing: A review of animal and human studies. Int. Wound J. 2020, 17, 1687–1694. [Google Scholar] [CrossRef]
- Uçkay, I.; Kressmann, B.; Malacarne, S.; Toumanova, A.; Jaafar, J.; Lew, D.; Lipsky, B.A. A randomized, controlled study to investigate the efficacy and safety of a topical gentamicin-collagen sponge in combination with systemic antibiotic therapy in diabetic patients with a moderate or severe foot ulcer infection. BMC Infect. Dis. 2018, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rastogi, A.; Sukumar, S.; Hajela, A.; Mukherjee, S.; Dutta, P.; Bhadada, S.K.; Bhansali, A. The microbiology of diabetic foot infections recently treated with antibiotic therapy: A prospective study from India. J. Diabetes Complicat. 2016, 31, 407–412. [Google Scholar] [CrossRef]
- Wu, M.; Pan, H.; Leng, W.; Lei, X.; Chen, L.; Liang, Z. Distribution of Microbes and Drug Susceptibility in Patients with Diabetic Foot Infections in Southwest China. J. Diabetes Res. 2018, 2018. [Google Scholar] [CrossRef]
- Gomes, D.; Santos, R.; Soares, R.S.; Reis, S.; Carvalho, S.; Rego, P.; Peleteiro, M.C.; Tavares, L.; Oliveira, M. Pexiganan in Combination with Nisin to Control Polymicrobial Diabetic Foot Infections. Antibiotics 2020, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Elumalai, K.; Yadhav, C.S.C.; Nishad, S.S.; Srinivasan, S.; Mounika, K.; Sivannan, S.; Supriya, K.; Kumar, A. The high lymphadenopathy and subcutaneous edema are associated with development of foot ulcer in type 2 diabetes: A collagen implanted antibiotic therapy. Bull. Fac. Pharm. Cairo Univ. 2018, 56, 101–103. [Google Scholar] [CrossRef]
- Si, R.; Han, J.; Zhang, X.; Ji, F.; Yang, X.; Zhang, J. The Application of 0.75% Metronidazole Gel for the Treatment of Chronic Wounds: Two Case Reports and Literature Review; Research Square AJE LLC: Durham, NC, USA, 2020. [Google Scholar] [CrossRef]
- Ashraf, M.N.; Azhar, M.; Akhtar, N.; Afzal, M.K. Comparison of Protein Synthesis Inhibiting Antibiotics Versus Cell Wall Inhibitor Antibiotics for Treatment of Diabetic Foot. Prof. Med. J. 2020, 27, 1212–1216. [Google Scholar] [CrossRef]
Figure 1.
The anatomical structure of human skin.

Figure 2.
Diagram (A) shows the normal morphological structure of fibroblasts in an acute wound. In Diagram (B), the fibroblasts become senescent, reduce in numbers, inhibit migration to the wounded site, and alter collagen synthesis.
Figure 2.
Diagram (A) shows the normal morphological structure of fibroblasts in an acute wound. In Diagram (B), the fibroblasts become senescent, reduce in numbers, inhibit migration to the wounded site, and alter collagen synthesis.

Figure 3.
The pathophysiology of diabetic foot ulcer (DFU).

Figure 4.
(A) shows normal flora/skin microbiome (S. epidermidis) on normal wound healing. The skin is protected from gram-positive /negative bacteria by S. epidermidis. The left corner shows factors that contribute to wound healing and further preventing colonization by skin pathogens. (B) shows both S. epidermidis and T cells help in wound closure and act as protective shields against pathogen invasions.
Figure 4.
(A) shows normal flora/skin microbiome (S. epidermidis) on normal wound healing. The skin is protected from gram-positive /negative bacteria by S. epidermidis. The left corner shows factors that contribute to wound healing and further preventing colonization by skin pathogens. (B) shows both S. epidermidis and T cells help in wound closure and act as protective shields against pathogen invasions.

Figure 5.
The figures above show microbes in acute and chronic wounds. (A) In acute wounds, fibrin fragments and keratin residues are favorable conditions for pathogens invasion into the skin. Hence, bacteria aggregate to prevent immune cells from destroying the bacterial community; (B) In chronic wounds, the biofilm formation secretes inflammatory cytokines, free radicals, and nitric oxide, which are toxic to cells and cell apoptosis.
Figure 5.
The figures above show microbes in acute and chronic wounds. (A) In acute wounds, fibrin fragments and keratin residues are favorable conditions for pathogens invasion into the skin. Hence, bacteria aggregate to prevent immune cells from destroying the bacterial community; (B) In chronic wounds, the biofilm formation secretes inflammatory cytokines, free radicals, and nitric oxide, which are toxic to cells and cell apoptosis.


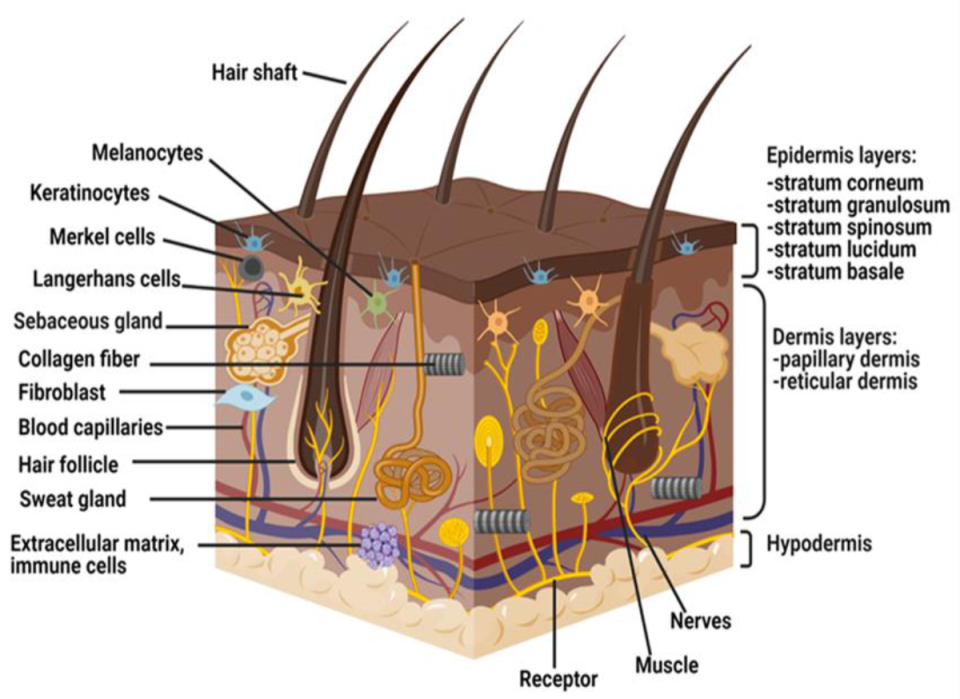{kind=link}
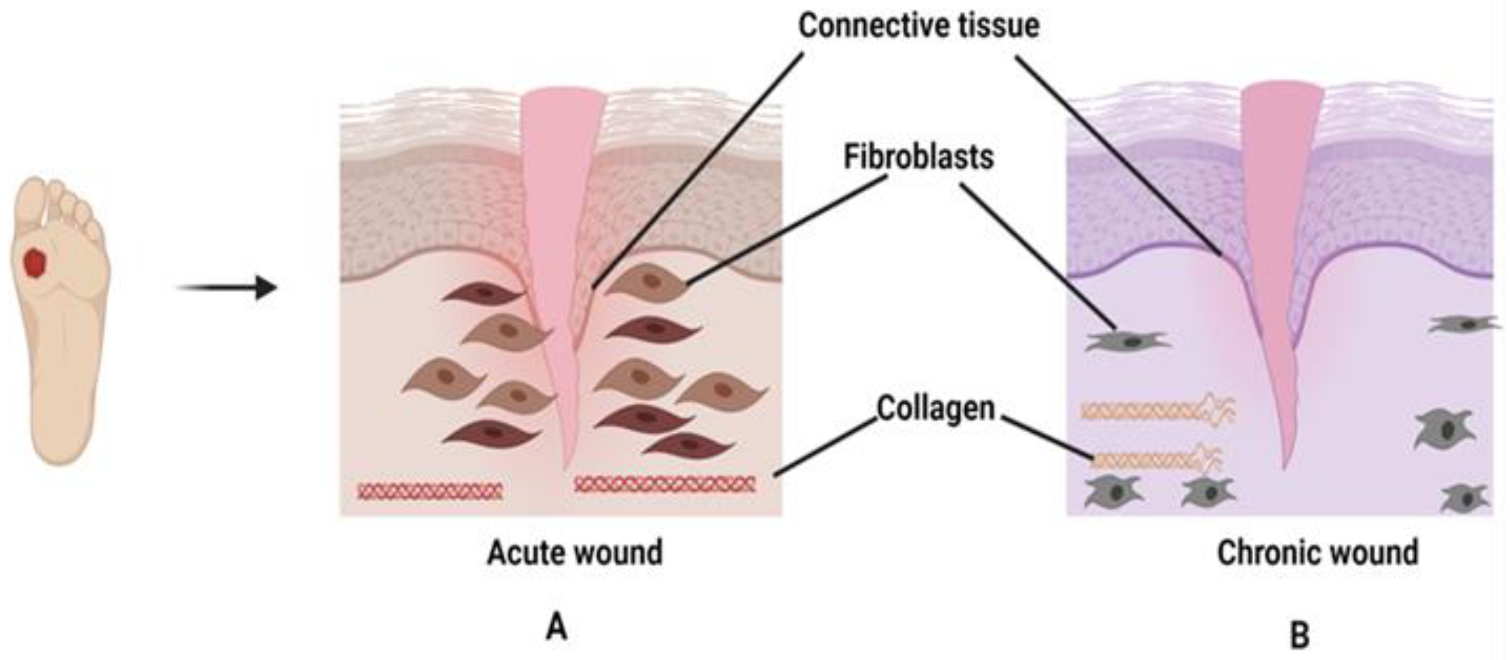{kind=link}
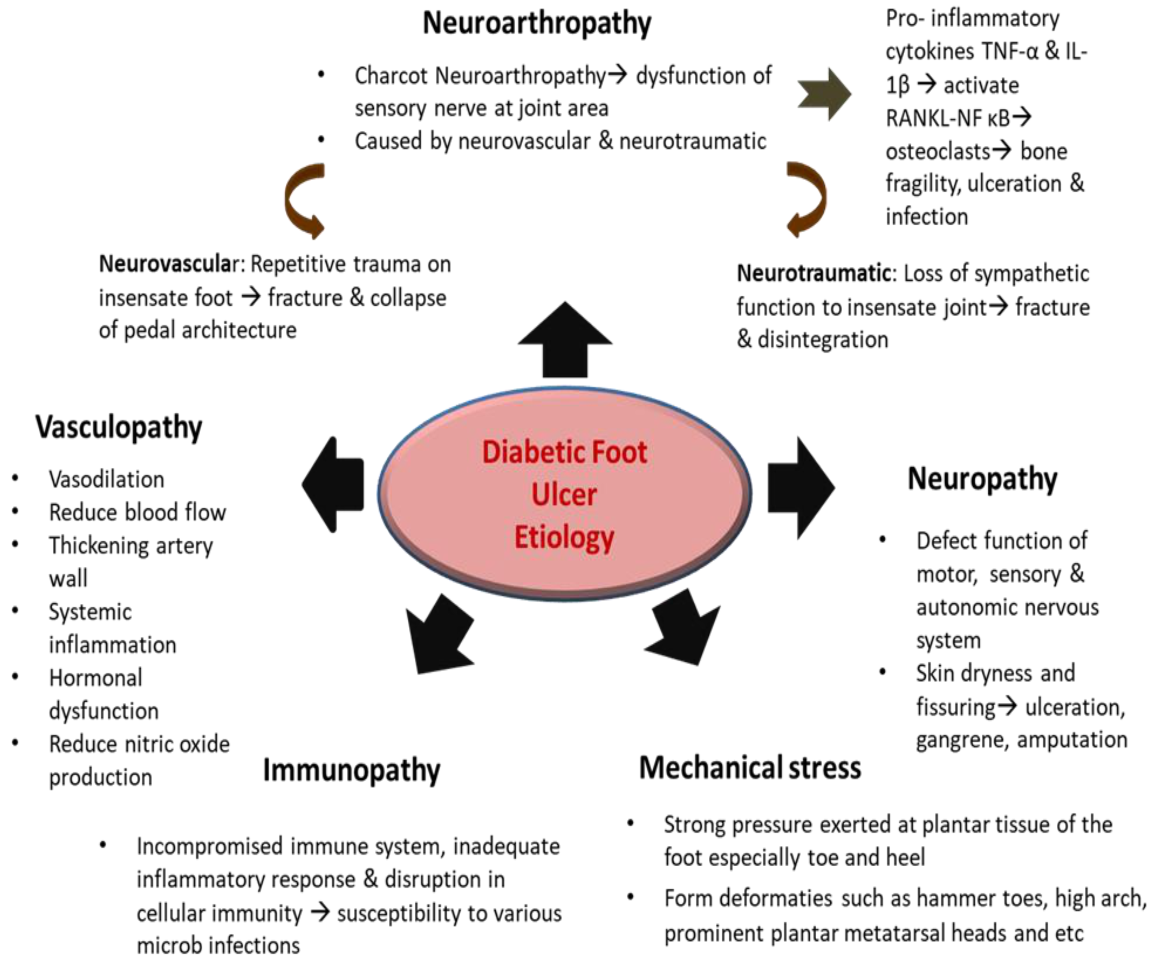{kind=link}
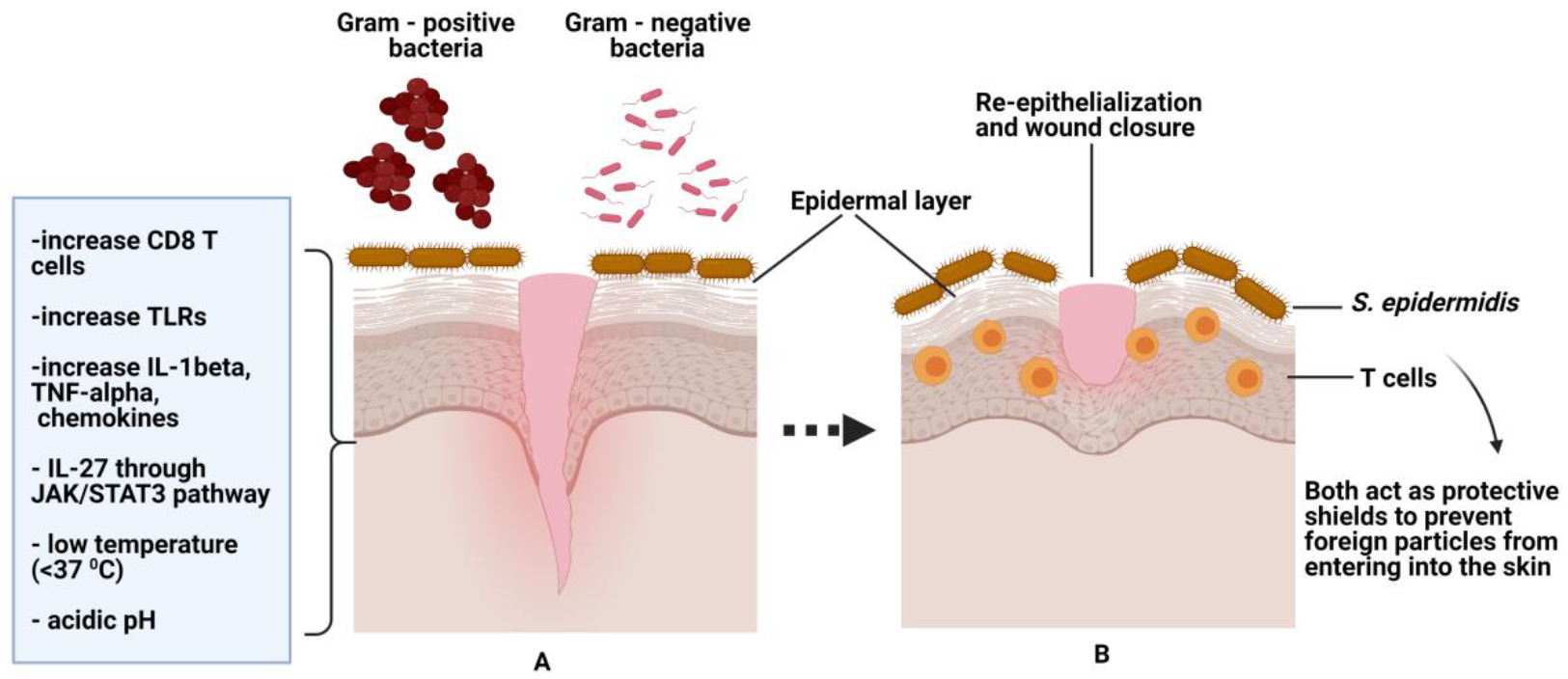{kind=link}
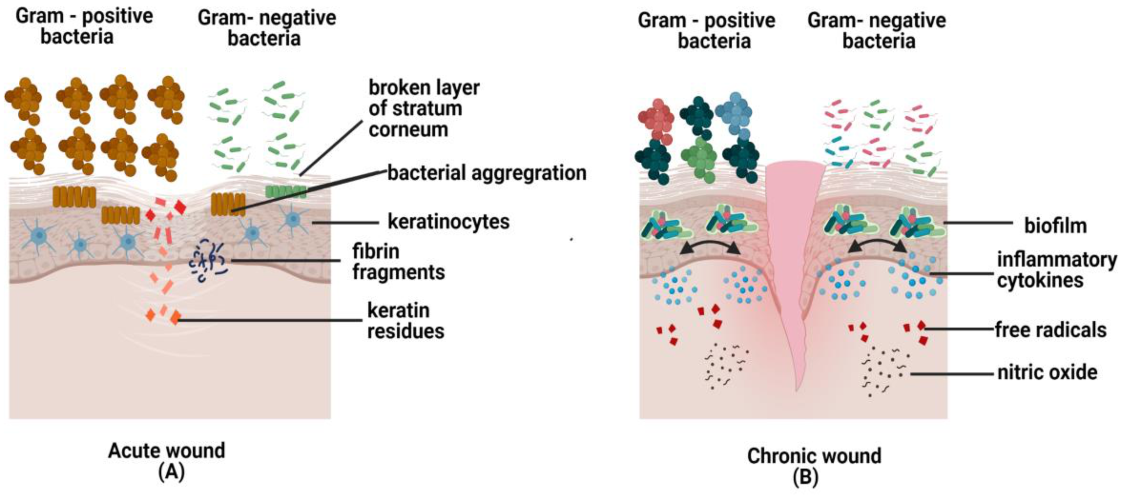{kind=link}
Table 1.
Differences between acute and chronic wound healing.
| Items | Acute Wound Healing | Chronic Wound Healing |
|---|---|---|
| Growth factors | Normal degradation [40] | High degradation [40] |
| Neutrophils | Normal activation of neutrophils to phagocyte the pathogens or foreign particles during inflammation [41] | High activation of neutrophils with excessive secretion of reactive oxygen species and ECM degradation [41] |
| Macrophage | During inflammation, ability to transform from pro-inflammatory macrophage, M1 to anti-inflammatory macrophage, M2 [30] | During inflammation, poor transformation of macrophage from M1 to M2 [30] |
| Anti-inflammatory cytokines | Decrease production [42] | Increase production [42] |
| MMPs and inflammatory cytokines | Decrease secretion of MMPs and inflammatory cytokines [42] | High secretion of MMPs and inflammatory cytokines [42] |
| Wound healing contaction/ contracture and Types of wound | Wound contraction Types of wound-normal scar wound [42] | Wound Contracture Types of wound; (a) chronic non-healing wound-ulcer, (b) dehiscence-scanty of wound healing, (c) fibrosis (hypertropic scars and keloids)—uncontrollable wound healing [42] |
| Pathological condition of epidermal skin | Epidermal skin still present as normal skin [27] | Hyperkeratotic (thick keratin layer or parakeraotic (anucleated keratinocytes) [27] |
| Duration of wound healing | 3 months [43] | More than 3 months and up to 7 months (for active ulcer) [3] |
| Phases of wound | Normal four phases-hemostasis, inflammation, proliferation and remodeling [23] | Not normal phases—Prolonged inflammation, impaired proliferative and remodeling phase [23] |
| Cell mitosis | Takes place [41] | No cell mitosis [41] |
| Granulation tissue | Normal production [40] | Neoangiogeneis, less fibroblasts, low oxygen→ tissue hypoxia Tissue hypoxia—low production of granulation tissue [40] |
Table 2.
Types of antibiotic/antimicrobial therapy, route of therapy, targeted pathogens, and impact to wound.
Table 2.
Types of antibiotic/antimicrobial therapy, route of therapy, targeted pathogens, and impact to wound.
| Types of Antibiotic/ Antimicrobial Therapy | Methods of Application/ Route of Therapy | Targeted Pathogens | Impact of Therapy | References |
|---|---|---|---|---|
| Gentamicin-collagen sponge with systemic antibiotic therapy (levofloxacin with clindamycin or amoxicillin-clavulanate | Gentamicin collagen sponge–topical Antibiotic—oral or intravenous | S. aureus, Streptococci, E. coli and P. aeruginosa | Overall no significant improvement in healing No side effects caused by antibiotics | [175] |
| Cephalosporin agent (ceftaroline fosamil) | Intravenous | Gram-positive bacteria, MRSA | Effective in treatment Potential substitute for glycopeptide therapy | [58] |
| Amoxicillin—clavulonate or cefotaxime | Oral or parenteral | Anaerobic bacteria | Sensitive to the antibiotic treatment | [176] |
| Tobramycin | Oral or parenteral | Gram-negative bacilli | Effective in treatment | [177] |
| Linezolid | Oral or parenteral | Staphylococcus sp., methicillin-resistant staphylococcus (MRS), Enterococcus sp. | Effective in treatment | [177] |
| Pexiganan and nisin (dual Antimicrobial peptide-biogel) | Topical on collagen DFU 3-D model | S.aureus and P.aeruginosa | Eradication of S.aureus isolates in infected area Inhibitory activity of P.aeruginosa against the AMP | [178] |
| Collagen with gentimycin sulphate, doxycycline and vancomycine | Apply topically on patient’s infected wound | Enterococcus sp. | Wound healed with tissue formation and granulation | [179] |
| Metronidazole | Apply topically on patient’s infected wound | S. aureus and anaerobic bacteria | Wound healed and formation of scab over the large wound | [180] |
| Clindamycin | Intravenous | S. aureus, S. pyrogenes, polymicrobial (P. aeruginosa, Klebsiella and Proteus) | Effective in treatment (clearance of infection) from the wound | [181] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Maheswary, T.; Nurul, A.A.; Fauzi, M.B. The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review. Pharmaceutics 2021, 13, 981. https://doi.org/10.3390/pharmaceutics13070981
AMA Style
Maheswary T, Nurul AA, Fauzi MB. The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review. Pharmaceutics. 2021; 13(7):981. https://doi.org/10.3390/pharmaceutics13070981
Chicago/Turabian StyleMaheswary, Thambirajoo, Asma Abdullah Nurul, and Mh Busra Fauzi. 2021. "The Insights of Microbes’ Roles in Wound Healing: A Comprehensive Review" Pharmaceutics 13, no. 7: 981. https://doi.org/10.3390/pharmaceutics13070981
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.