
The YfkO Nitroreductase from Bacillus Licheniformis on Gold-Coated Superparamagnetic Nanoparticles: Towards a Novel Directed Enzyme Prodrug Therapy Approach
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Enzyme Reactivity with CB1954
2.3. CB1954 Kinetics
2.4. HPLC Analysis
2.5. Cell Viability Assays
2.6. Synthesis and Purification of AuMNPs
2.7. Conjugation of Cys-Tagged Enzymes to Au-MNPs
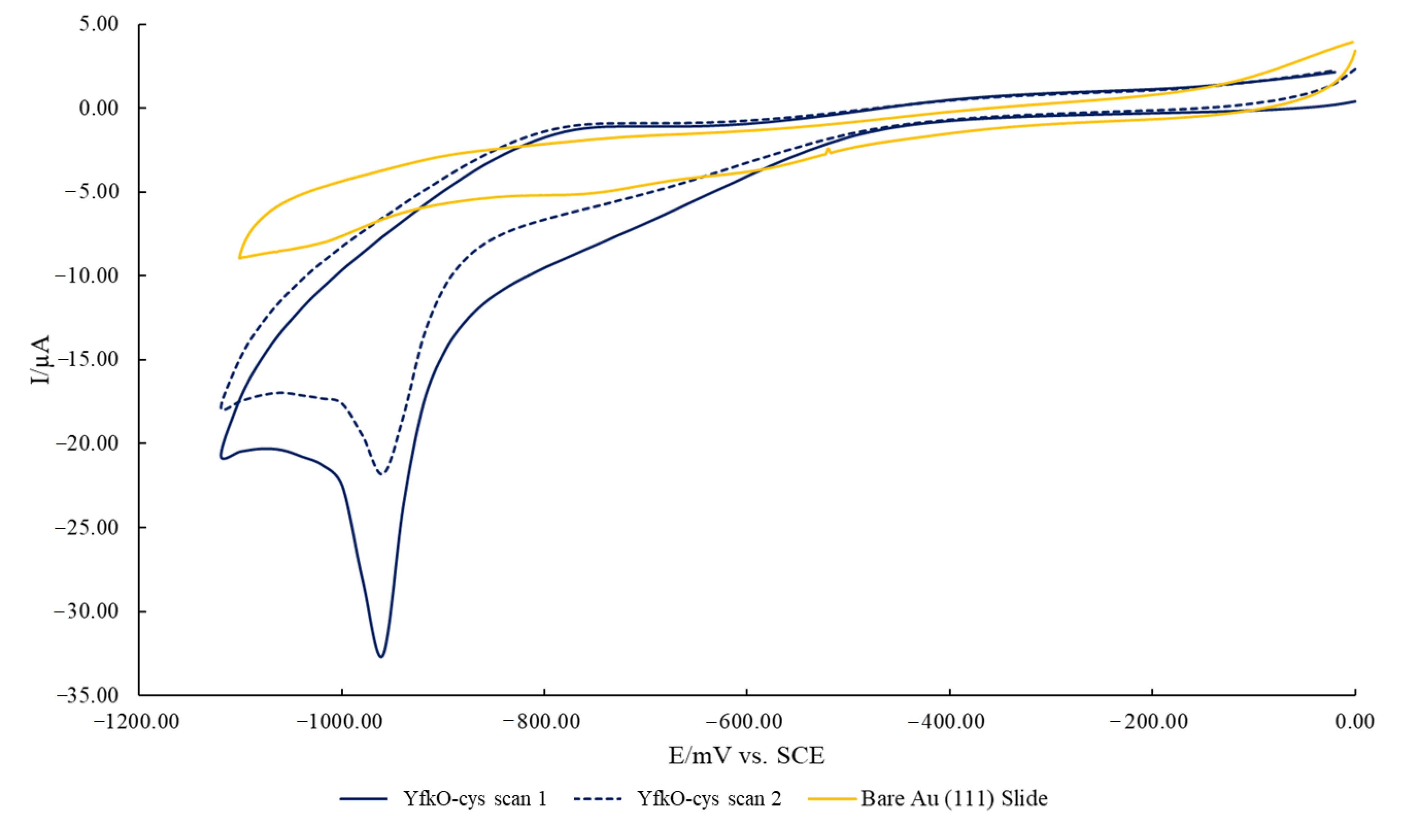
2.8. Characterisation of AuMNPs and Subsequent Conjugates
3. Results
3.1. Protein Expression and Purification
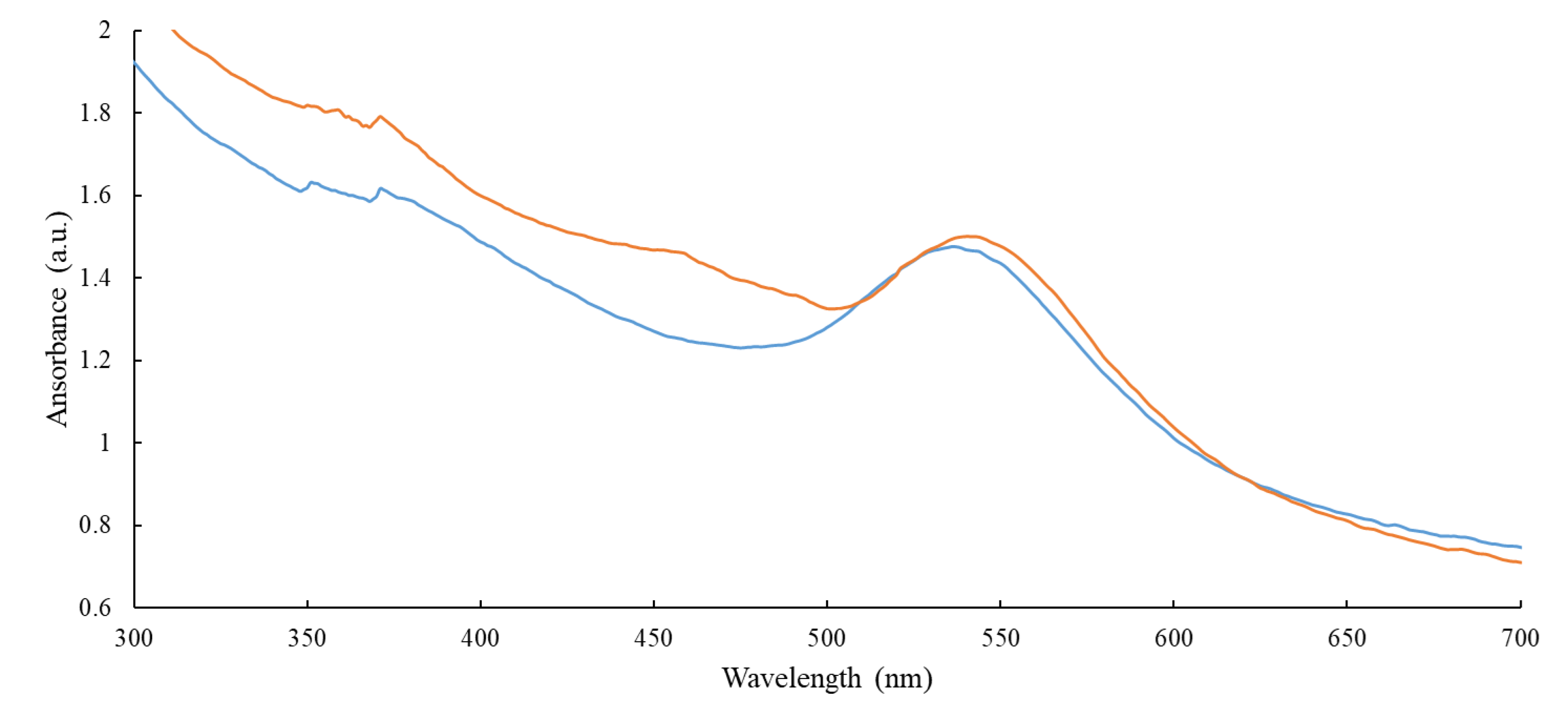
3.2. Characterization of AuMNPs and Subsequent Conjugates
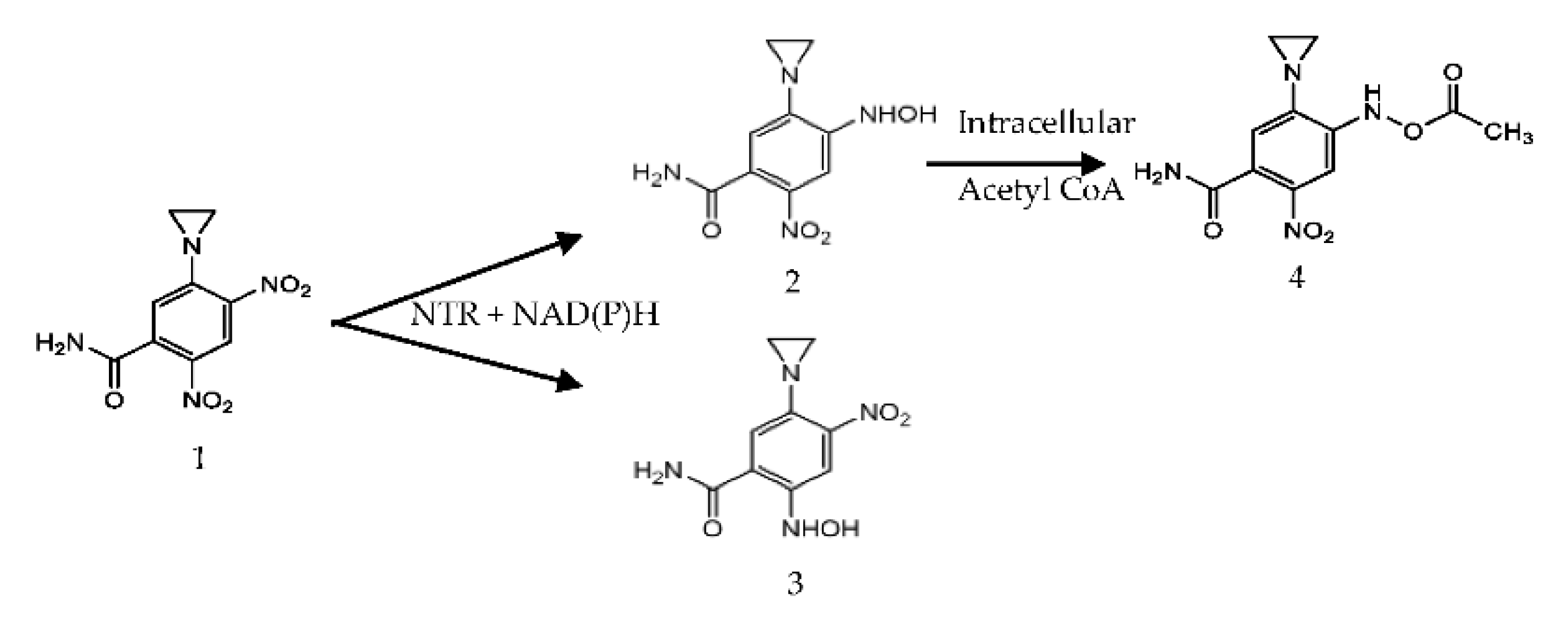
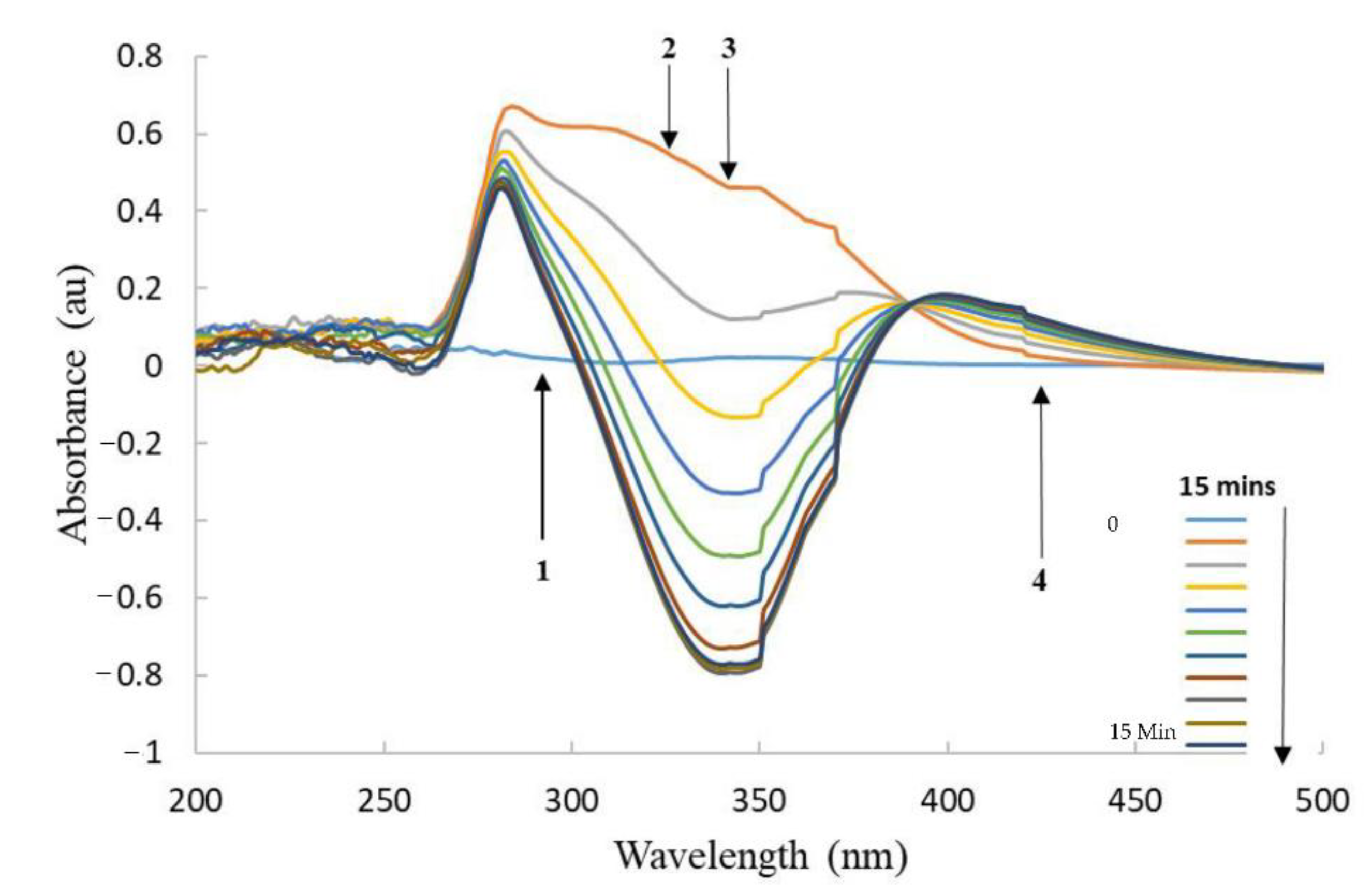
3.3. Enzyme Reactivity with CB1954
3.4. Michaelis-Menten Kinetics
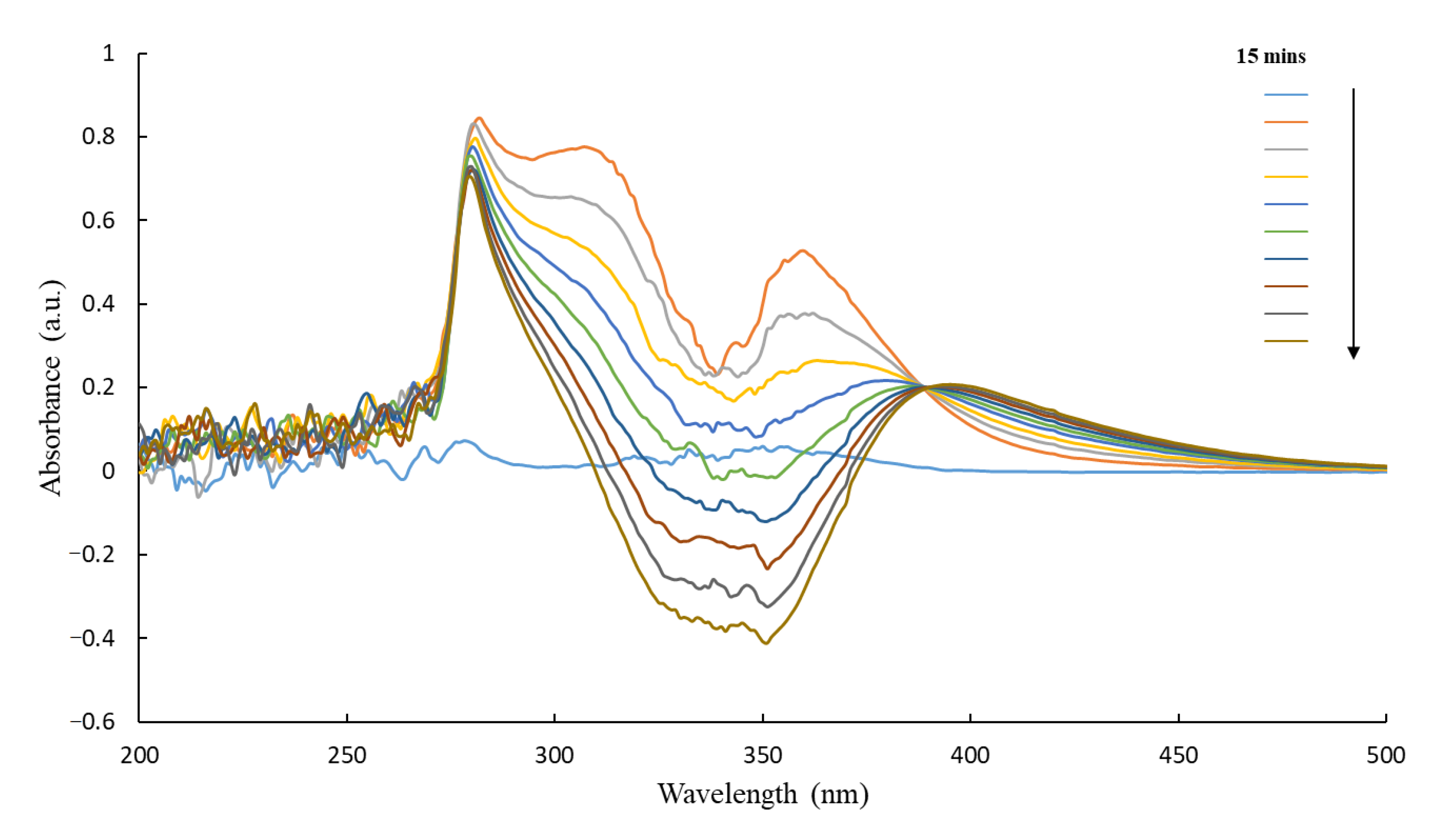
3.5. HPLC of CB1954 Products
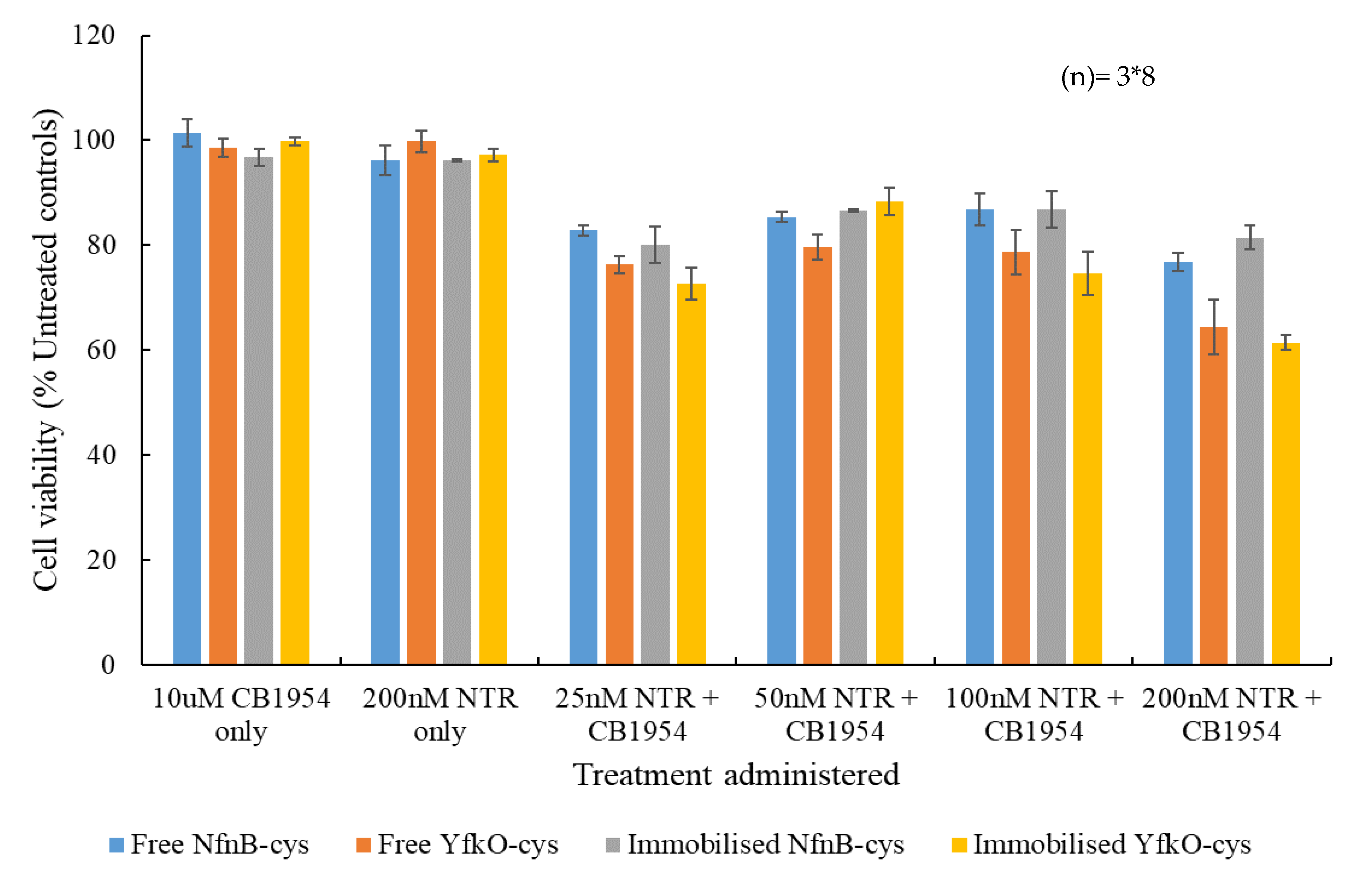
3.6. Cell Viability Assays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The World Health Organization. The World Health Organization’s Fight against Cancer: Strategies that Prevent, Cure and Care; WHO: Geneva, Switzerland, 2007. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics. Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- Albert, A. Chemical Aspects of Selective Toxicity. Nat. Cell Biol. 1958, 182, 421–423. [Google Scholar] [CrossRef]
- Sinhababu, A.K.; Thakker, D.R. Prodrugs of anticancer agents. Adv. Drug Deliv. Rev. 1996, 19, 241–273. [Google Scholar] [CrossRef]
- Feng, S.-S.; Chien, S. Chemotherapeutic engineering: Application and further development of chemical engineering principles for chemotherapy of cancer and other diseases. Chem. Eng. Sci. 2003, 58, 4087–4114. [Google Scholar] [CrossRef]
- Kratz, F.; Müller, I.A.; Ryppa, C.; Warnecke, A. Prodrug Strategies in Anticancer Chemotherapy. ChemMedChem 2008, 3, 20–53. [Google Scholar] [CrossRef]
- A Denny, W. Prodrug strategies in cancer therapy. Eur. J. Med. Chem. 2001, 36, 577–595. [Google Scholar] [CrossRef]
- Anlezark, G.M.; Melton, R.G.; Sherwood, R.F.; Coles, B.; Friedlos, F.; Knox, R.J. The bioactivation of 5-(aziridin-1-yl)-2,4-dinitrobenzamide (CB1954)--I. Purification and properties of a nitroreductase enzyme from Escherichia coli--a potential enzyme for antibody-directed enzyme prodrug therapy (ADEPT). Biochem. Pharmacol. 1992, 44, 2289–2295. [Google Scholar] [CrossRef]
- Bagshawe, K. Antibody-Directed Enzyme Prodrug Therapy. In Prodrugs; Springer: London, UK, 2007; pp. 527–540. [Google Scholar]
- Palmer, D.H.; Mautner, V.; Mirza, D.F.; Oliff, S.; Gerritsen, W.R.; Van Der Sijp, J.R.; Hubscher, S.G.; Reynolds, G.M.; Bonney, S.; Rajaratnam, R.; et al. Virus-Directed Enzyme Prodrug Therapy: Intratumoral Administration of a Replication-Deficient Adenovirus Encoding Nitroreductase to Patients with Resectable Liver Cancer. J. Clin. Oncol. 2004, 22, 1546–1552. [Google Scholar] [CrossRef]
- Searle, P.F.; Chen, M.-J.; Hu, L.; Race, P.R.; Lovering, A.L.; I Grove, J.; Guise, C.; Jaberipour, M.; James, N.D.; Mautner, V.; et al. NITROREDUCTASE: A PRODRUG-ACTIVATING ENZYME FOR CANCER GENE THERAPY. Clin. Exp. Pharmacol. Physiol. 2004, 31, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Race, P.R.; Lovering, A.L.; White, S.A.; Grove, J.I.; Searle, P.F.; Wrighton, C.W.; Hyde, E. Kinetic and Structural Characterisation of Escherichia coli Nitroreductase Mutants Showing Improved Efficacy for the Prodrug Substrate CB1954. J. Mol. Biol. 2007, 368, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Prosser, G.; Copp, J.; Syddall, S.; Williams, E.; Smaill, J.; Wilson, W.; Patterson, A.; Ackerley, D. Discovery and evaluation of Escherichia coli nitroreductases that activate the anti-cancer prodrug CB1954. Biochem. Pharmacol. 2010, 79, 678–687. [Google Scholar] [CrossRef]
- Prosser, G.A.; Copp, J.N.; Mowday, A.M.; Guise, C.P.; Syddall, S.P.; Williams, E.M.; Horvat, C.N.; Swe, P.M.; Ashoorzadeh, A.; Denny, W.A.; et al. Creation and screening of a multi-family bacterial oxidoreductase library to discover novel nitroreductases that efficiently activate the bioreductive prodrugs CB1954 and PR-104A. Biochem. Pharmacol. 2013, 85, 1091–1103. [Google Scholar] [CrossRef]
- Gwenin, V.V.; Gwenin, C.D.; Kalaji, M. Colloidal Gold Modified with a Genetically Engineered Nitroreductase: Toward a Novel Enzyme Delivery System for Cancer Prodrug Therapy. Langmuir 2011, 27, 14300–14307. [Google Scholar] [CrossRef]
- Gwenin, V.V.; Gwenin, C.D.; Kalaji, M. Drug Activation System, WO2011026898. 2009. Available online: https://patentscope.wipo.int/search/en/detail.jsf?docId=WO2011026898 (accessed on 11 May 2018).
- Bryant, D.W.; McCalla, D.R.; Leeksma, M.; Laneuville, P. Type I nitroreductases of Escherichia coli. Can. J. Microbiol. 1981, 27, 81–86. [Google Scholar] [CrossRef]
- Bryant, C.; Hubbard, L.; McElroy, W.D. Cloning, nucleotide sequence, and expression of the nitroreductase gene from Enterobacter cloacae. J. Biol. Chem. 1991, 266, 4126–4130. [Google Scholar] [CrossRef]
- Bryant, C.; DeLuca, M. Purification and characterization of an oxygen-insensitive NAD(P)H nitroreductase from Enterobacter cloacae. J. Biol. Chem. 1991, 266, 4119–4125. [Google Scholar] [CrossRef]
- Denny, W.A. Prodrugs for Gene-Directed Enzyme-Prodrug Therapy (Suicide Gene Therapy). J. Biomed. Biotechnol. 2003, 2003, 48–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, E.M.; Little, R.F.; Mowday, A.M.; Rich, M.H.; Chan-Hyams, J.V.; Copp, J.N.; Smaill, J.B.; Patterson, A.; Ackerley, D.F. Nitroreductase gene-directed enzyme prodrug therapy: Insights and advances toward clinical utility. Biochem. J. 2015, 471, 131–153. [Google Scholar] [CrossRef]
- Malekshah, O.M.; Chen, X.; Nomani, A.; Sarkar, S.; Hatefi, A. Enzyme/Prodrug Systems for Cancer Gene Therapy. Curr. Pharmacol. Rep. 2016, 2, 299–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knox, R.J.; Boland, M.P.; Friedlos, F.; Coles, B.; Southan, C.; Roberts, J.J. The nitroreductase enzyme in walker cells that activates 5-(aziridin-1-yl)-2,4-dinitrobenzamide (CB 1954) to 5-(aziridin-1-YL)-4-hydroxylamino-2-nitrobenzamide is a form of NAD(P)H dehydrogenase (quinone) (EC 1.6.99.2). Biochem. Pharmacol. 1988, 37, 4671–4677. [Google Scholar] [CrossRef]
- Knox, R.J.; Friedlos, F.; Marchbank, T.; Roberts, J.J. Bioactivation of CB 1954: Reaction of the active 4-hydroxylamino derivative with thioesters to form the ultimate DNA-DNA interstrand crosslinking species. Biochem. Pharmacol. 1991, 42, 1691–1697. [Google Scholar] [CrossRef]
- Prosser, G.; Patterson, A.; Ackerley, D. uvrB gene deletion enhances SOS chromotest sensitivity for nitroreductases that preferentially generate the 4-hydroxylamine metabolite of the anti-cancer prodrug CB1954. J. Biotechnol. 2010, 150, 190–194. [Google Scholar] [CrossRef]
- Weedon, S.J.; Green, N.K.; McNeish, I.A.; Gilligan, M.G.; Mautner, V.; Wrighton, C.J.; Mountain, A.; Young, L.S.; Kerr, D.J.; Searle, P.F. Sensitisation of human carcinoma cells to the prodrug CB1954 by adenovirus vector-mediated expression of E. coli nitroreductase. Int. J. Cancer 2000, 86, 848–854. [Google Scholar] [CrossRef]
- Patel, P.; Young, J.G.; Mautner, V.; Ashdown, D.; Bonney, S.; Pineda, R.G.; Collins, S.I.; Searle, P.F.; Hull, D.; Peers, E.; et al. A phase I/II clinical trial in localized prostate cancer of an adenovirus expressing nitroreductase with CB1954 [correction of CB1984]. Mol. Ther. 2009, 17, 1292–1299. [Google Scholar] [CrossRef]
- Emptage, C.D.; Knox, R.J.; Danson, M.J.; Hough, D.W. Nitroreductase from Bacillus licheniformis: A stable enzyme for prodrug activation. Biochem. Pharmacol. 2009, 77, 21–29. [Google Scholar] [CrossRef]
- Gwenin, C.D.; Kalaji, M.; Williams, P.A.; Kay, C.M. A kinetic analysis of three modified novel nitroreductases. Biogeochemistry 2010, 22, 463–474. [Google Scholar] [CrossRef]
- Gwenin, C.; Kalaji, M.; Williams, P.; Jones, R. The orientationally controlled assembly of genetically modified enzymes in an amperometric biosensor. Biosens. Bioelectron. 2007, 22, 2869–2875. [Google Scholar] [CrossRef] [PubMed]
- Gwenin, V.V.; Poornima, P.; Halliwell, J.; Ball, P.; Robinson, G.; Gwenin, C.D. Identification of novel nitroreductases from Bacillus cereus and their interaction with the CB1954 prodrug. Biochem. Pharmacol. 2015, 98, 392–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ball, P.; Thompson, E.; Anderson, S.; Gwenin, V.; Gwenin, C. Time dependent HPLC analysis of the product ratio of enzymatically reduced prodrug CB1954 by a modified and immobilised nitroreductase. Eur. J. Pharm. Sci. 2019, 127, 217–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.; Hobbs, R.; Gwenin, V.; Ball, P.; Bennie, L.; Coulter, J.; Gwenin, C. Cell-penetrating peptides as a tool for the ceullular uptake of a genetically modified nitroreductase for use in directed enzyme prodrug therapy. J. Funct. Biol. 2019, 10, 45. [Google Scholar] [CrossRef] [Green Version]
- Ball, P.; Halliwell, J.; Anderson, S.; Gwenin, V.; Gwenin, C. Evaluation of two xenobiotic reductases from Pseudomonas putida for their suitability for magnetic nanoparticle-directed enzyme prodrug therapy as a novel approach to cancer treatment. Microbiology 2020, 9, 1–10. [Google Scholar] [CrossRef]
- Haiss, W.; Thanh, N.T.K.; Aveyard, J.; Fernig, D.G. Determination of Size and Concentration of Gold Nanoparticles from UV—Vis Spectra Determination of Size and Concentration of Gold Nanoparticles from UV—Vis Spectra. Anal. Chem. 2007, 79, 4215–4221. [Google Scholar] [CrossRef]
- Haiss, W.; Lackey, D.; Sass, J.K.; Besocke, K.H. Atomic resolution scanning tunneling microscopy images of Au(111) surfaces in air and polar organic solvents. J. Chem. Phys. 1991, 95, 2193–2196. [Google Scholar] [CrossRef]
- Viana, A.; Kalaji, M.; Abrantes, L. Self-assembled monolayers of Vitamin B12 disulphide derivatives on gold. Electrochim. Acta 2002, 47, 1587–1594. [Google Scholar] [CrossRef]
- Chung-Faye, G.; Palmer, D.; Anderson, D.; Clark, J.; Downes, M.; Baddeley, J.; Hussain, S.; Murray, P.I.; Searle, P.; Seymour, L.; et al. Virus-directed Enzyme Prodrug Therapy with Nitroimidazole Reductase: A Phase I and Pharmacokinetic Study. Clin. Can. Res. 2001, 7, 2662–2668. [Google Scholar]
- McNeish, I.; Green, N.; Gilligan, M.; Ford, M.; Mautner, V.; Young, L.; Kerr, D.; Searle, P. Virus directed enzyme prodrug therapy for ovarian and pancreatic cancer using retrovirally delivered E. coli nitroreductase and CB1954. Gene Ther. 1998, 5, 1061–1069. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P. Hormesis Defined. Ageing Res. Rev. 2008, 7, 1–7. [Google Scholar] [CrossRef]
- Ying, W. NAD+/NADH and NADP+/NADPH in Cellular Functions and Cell Death: Regulation and Biological Consequences. Antioxid. Redox Signal. 2008, 10, 179–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Helsby, A.N.; Ferry, D.M.; Patterson, A.; Pullen, S.M.; Wilson, W.R. 2-Amino metabolites are key mediators of CB 1954 and SN 23862 bystander effects in nitroreductase GDEPT. Br. J. Cancer 2004, 90, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Chan-Hyams, J.V.; Copp, J.N.; Smaill, J.B.; Patterson, A.V.; Ackerley, D.F. Evaluating the abilities of diverse nitroaromatic prodrug metabolites to exit a model Gram negative vector for bacterial-directed enzyme-prodrug therapy. Biochem. Pharmacol. 2018, 158, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Vass, O.S.; Jarrom, D.; Wilson, W.R.; I Hyde, E.; Searle, P.F. E. coli NfsA: An alternative nitroreductase for prodrug activation gene therapy in combination with CB1954. Br. J. Cancer 2009, 100, 1903–1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Vmax (µM s−1) | Km (µM) | Kcat (s−1) | Kcat/Km (µM−1 s−1) |
|---|---|---|---|---|
| NfnB-cys | 19.4 ± 1.3 | 5000 ± 700 | 55.34 ± 0.57 | 0.0109 ± 3.8 × 10−3 |
| Immobilised NfnB-cys | 10.89 ± 1.2 | 1108 ± 300 | 61.85 ± 0.64 | 0.0558 ± 3.5 × 10−3 |
| YfkO-cys | 6.5 ± 1.2 | 830 ± 250 | 39.34 ± 0.89 | 0.0471 ± 3.6 × 10−3 |
| Immobilised YfkO-cys | 8.35 ± 1.4 | 1374 ± 400 | 50.30 ± 0.78 | 0.0366 ± 3.7 × 10−3 |
| Enzyme | Hydroxylamine Product Ratio (2-NHOH:4-NHOH) |
|---|---|
| NfnB-cys | 32:68 |
| Immobilised NfnB-cys | 13:87 |
| YfkO-cys | 4:96 |
| Immobilised YfkO-cys | 1:99 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ball, P.; Hobbs, R.; Anderson, S.; Thompson, E.; Gwenin, V.; Von Ruhland, C.; Gwenin, C. The YfkO Nitroreductase from Bacillus Licheniformis on Gold-Coated Superparamagnetic Nanoparticles: Towards a Novel Directed Enzyme Prodrug Therapy Approach. Pharmaceutics 2021, 13, 517. https://doi.org/10.3390/pharmaceutics13040517
Ball P, Hobbs R, Anderson S, Thompson E, Gwenin V, Von Ruhland C, Gwenin C. The YfkO Nitroreductase from Bacillus Licheniformis on Gold-Coated Superparamagnetic Nanoparticles: Towards a Novel Directed Enzyme Prodrug Therapy Approach. Pharmaceutics. 2021; 13(4):517. https://doi.org/10.3390/pharmaceutics13040517
Chicago/Turabian StyleBall, Patrick, Robert Hobbs, Simon Anderson, Emma Thompson, Vanessa Gwenin, Christopher Von Ruhland, and Christopher Gwenin. 2021. "The YfkO Nitroreductase from Bacillus Licheniformis on Gold-Coated Superparamagnetic Nanoparticles: Towards a Novel Directed Enzyme Prodrug Therapy Approach" Pharmaceutics 13, no. 4: 517. https://doi.org/10.3390/pharmaceutics13040517