
Antimicrobial and Immunomodulating Activities of Two Endemic Nepeta Species and Their Major Iridoids Isolated from Natural Sources

 ,
,  , , , , , ,
, , , , , ,
Abstract
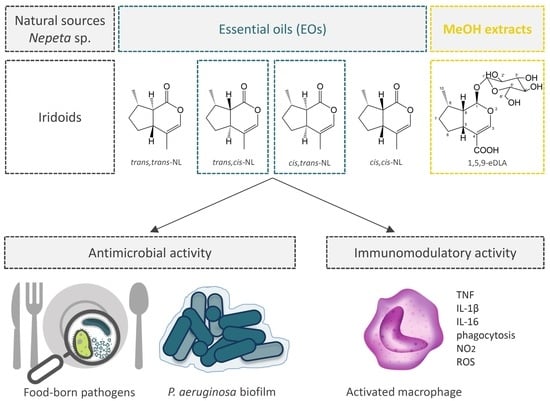
:
1. Introduction
2. Results and Discussion
2.1. Isolation of cis,trans- and trans,cis-Nepetalactone
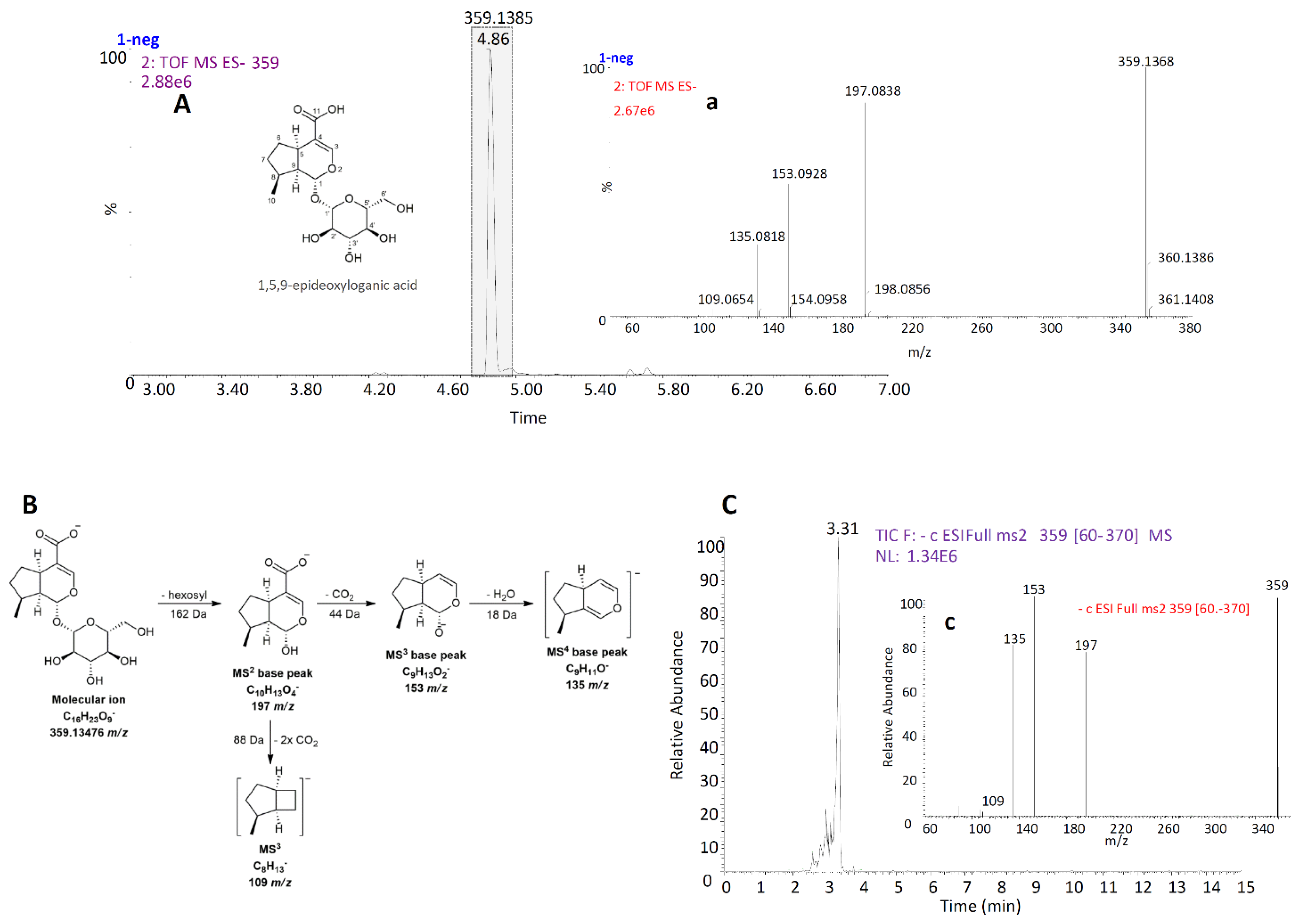
2.2. Isolation of 1,5,9-Epideoxyloganic Acid
2.3. Chemical Composition of N. rtanjensis and N. argolica Subsp. argolica Methanol Extracts
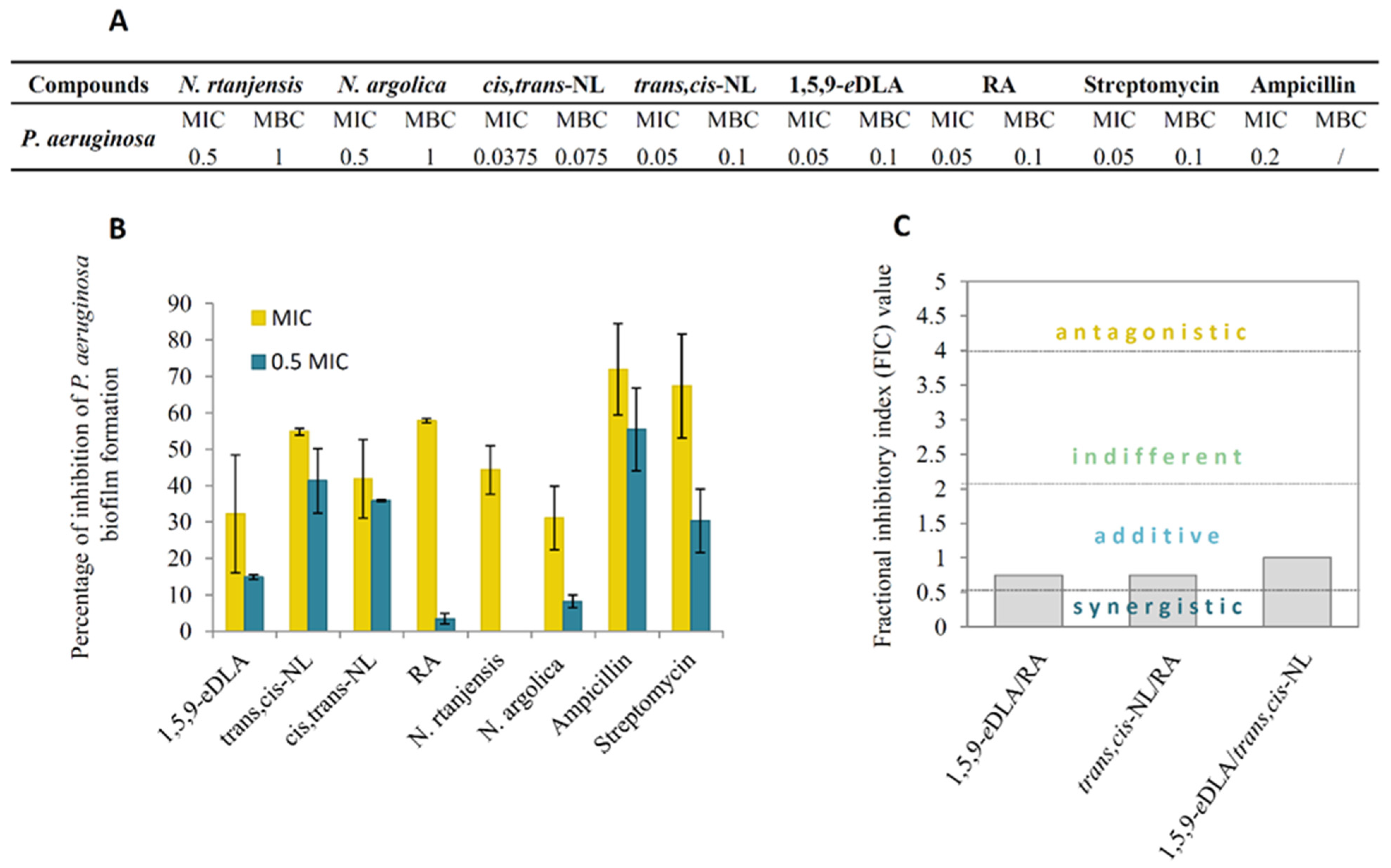
2.4. Antimicrobial Activity
2.5. Immunomodulating Activity
2.6. In Conclusion
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Isolation of cis,trans- and trans,cis-Nepetalactone
3.2.1. Origin of Essential Oils
3.2.2. Semi-Preparative HPLC/DAD Fractionation and Isolation of cis,trans- and trans,cis-Nepetalactone
3.2.3. GC-MS Identification of Nepetalactone Diastereoisomers in Isolated Fractions
3.2.4. UHPLC-MS2 Analysis of Isolated Fractions
3.2.5. NMR Analysis for the Structural Characterization of Isolated cis,trans- and trans,cis-Nepetalactone
3.3. Isolation of 1,5,9-Epideoxyloganic Acid
3.3.1. Plant Material
3.3.2. Preparation of Methanol Extracts
3.3.3. HRMS Analysis of N. rtanjensis Methanol Extract
3.3.4. Fractionation of N. rtanjensis Methanol Extract
3.3.5. Preparative HPLC/DAD Isolation of 1,5,9-Epideoxyloganic Acid
3.3.6. NMR Structure Confirmation of 1,5,9-Epideoxyloganic Acid
3.4. Phytochemical Characterization of Methanol Extracts of Nepeta rtanjensis and N. argolica subsp. argolica Grown In Vitro
3.4.1. Plant Material
3.4.2. Preparation of Methanol Extracts
3.4.3. Identification of Phytochemicals in Methanol Extracts Using UHPLC-LTQ OrbiTrap XL
3.4.4. UHPLC/qqqMS2 Quantification of Major Iridoids in Methanol Extracts
3.4.5. UHPLC/qqqMS2 Quantification of Major Phenolics in Methanol Extracts
3.5. Antimicrobial Activity of Two Nepeta Species Methanol Extracts and Their Major Iridoids
3.5.1. Microorganisms and Culture Conditions
3.5.2. Microdilution Method
3.5.3. Antibiofilm Activity
3.5.4. Antimicrobial Interaction between trans,cis-NL, 1,5,9-eDLA, and RA
3.6. Immunomodulating Activity of Two Nepeta Species Methanol Extracts and Isolated Nepetalactones
3.6.1. Cells and Cell Cultures
3.6.2. Cell Viability Assays
3.6.3. ELISA Test for Determination of Cytokines
3.6.4. Detection of Phagocytosis
3.6.5. Detection of ROS and NO Production
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salehi, B.; Valussi, M.; Jugran, A.K.; Martorell, M.; Ramírez-Alarcón, K.; Stojanović-Radić, Z.Z.; Antolak, H.; Kręgiel, D.; Mileski, K.S.; Sharifi-Rad, M.; et al. Nepeta species: From farm to food applications and phytotherapy. Trends Food Sci. Technol. 2018, 80, 104. [Google Scholar] [CrossRef]
- Bellahsene, C.; Bendahou, M.; Khadir, A.; Zenati, F.; Benbelaïd, F.; Aissaoui, N. Antimicrobial activity and chemical composition of essential oil and hydrosol extract of Nepeta nepetella subsp. amethystina (Poir.) Briq. from Algeria. J. Appl. Pharm. Sci. 2015, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Kalpoutzakis, E.; Aligiannis, N.; Mentis, A.; Mitaku, S.; Charvala, C. Composition of the essential oil of two Nepeta species and in vitro evaluation of their activity against Helicobacter pylori. Planta Med. 2001, 67, 880. [Google Scholar] [CrossRef]
- Nestorović, J.; Mišić, D.; Šiler, B.; Soković, M.; Glamočlija, J.; Ćirić, A.; Maksimović, V.; Grubišić, D. Nepetalactone content in shoot cultures of three endemic Nepeta species and the evaluation of their antimicrobial activity. Fitoterapia 2010, 81, 621. [Google Scholar] [CrossRef]
- Formisano, C.; Rigano, D.; Senatore, F. Chemical Constituents and Biological Activities of Nepeta Species. Chem. Biodiver. 2011, 8, 1783–1818. [Google Scholar] [CrossRef] [PubMed]
- Bates, R.B.; Sigel, C.W. Terpenoids. Cis-trans- and trans-cis-Nepetalactones. Experientia 1963, 19, 564–565. [Google Scholar] [CrossRef]
- Hardie, J.; Peace, L.; Pickett, J.A.; Smiley, D.W.M.; Storer, J.R.; Wadhams, L.J. Sex pheromone stereochemistry and purity affect field catches of male aphids. J. Chem. Ecol. 1997, 23, 2547–2554. [Google Scholar] [CrossRef]
- Peterson, C.J.; Nemetz, L.T.; Jones, L.M.; Coats, J.R. Behavioral activity of catnip (Lamiaceae) essential oil components to the german cockroach (Blattodea: Blattellidae). Entomol. Soc. Am. 2002, 95, 377–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birkett, M.A.; Hassanali, A.; Hoglund, S.; Pettersson, J.; Pickett, J.A. Repellent activity of catmint, Nepeta cataria, and iridoid nepetalactone isomers against Afro-tropical mosquitoes, ixodid ticks and red poultry mites. Phytochemistry 2011, 72, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gkinis, G.; Tzakou, O.; Iliopoulou, D.; Roussis, V. Chemical Composition and Biological Activity of Nepeta parnassica Oils and Isolated Nepetalactones. Z. Naturforsch. C. J. Biosci. 2003, 58, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Mathela, C.S.; Tewari, G.; Singh, D. Antifungal activity of Nepeta elliptica Royle ex Benth. oil and its major constituent (7R)-trans,trans-nepetalactone: A comparative study. Ind. Crop. Prod. 2014, 55, 70–74. [Google Scholar] [CrossRef]
- Murai, F.; Tagawa, M.; Damtoft, S.; Jensen, S.R.; Nielsen, B.J. (1R, 5R, 8S, 9S)-Deoxyloganic acid from Nepeta cataria. Chem. Pharm. Bull. 1984, 32, 2809–2814. [Google Scholar] [CrossRef] [Green Version]
- Takeda, Y.; Ooiso, Y.; Masuda, T.; Honda, G.; Otsuka, H.; Sezik, E.; Yesilada, E. Iridoid and eugenol glycosides from Nepeta cadmea. Phytochemistry 1998, 49, 787–791. [Google Scholar] [CrossRef]
- Kökdil, G.; Yalçin, S.M.; Topçu, G. Nepetalactones and other constituents of Nepeta nuda ssp. albiflora. Turk. J. Chem. 1999, 23, 99–104. [Google Scholar]
- Ahmed, A.A.; Hassan, H.E.; Hegazy, M.F.; Tzakou, O.; Couladis, M.; Mohamed, A.E.-H.H.; Abdella, M.A.; Pare, P. Argolic acid A and argolic methyl ester B, two new cyclopentano-monoterpenes diol from Nepeta argolica. Nat. Prod. Com. 2006, 1, 523–526. [Google Scholar] [CrossRef] [Green Version]
- Stojanović, G.; Radulović, N.; Lazarević, J.; Miladinović, D.; Đoković, D. Antimicrobial Activity of Nepeta rtanjensis Essential Oil. J. Essent. Oil Res. 2005, 17, 587–589. [Google Scholar] [CrossRef]
- Mišić, D.; Šiler, B.; Gašić, U.; Avramov, S.; Živković, S.; Nestorović Živković, J.; Milutinović, M.; Tešić, Ž. Simultaneous UHPLC/DAD/(+/−)HESI-MS/MS analysis of phenolic acids and nepetalactones in methanol extracts of Nepeta species: A possible application in chemotaxonomic studies. Phytochem. Anal. 2015, 26, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Aničić, N.; Matekalo, D.; Skorić, M.; Živković, J.N.; Petrović, L.; Dragićević, M.; Dmitrović, S.; Mišić, D. Alterations in nepetalactone metabolism during polyethylene glycol (PEG)-induced dehydration stress in two Nepeta species. Phytochemistry 2020, 174, 112340. [Google Scholar] [CrossRef]
- Sharma, A.; Cooper, R.; Bhardwaj, G.; Cannoo, D.S. The genus Nepeta: Traditional uses, phytochemicals and pharmacological properties. J. Ethnopharmacol. 2020, 268, 113679. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Cheng, K.W.; Wu, Q.; Simon, J.E. Quantification of nepetalactones in catnip (Nepeta cataria L.) by HPLC coupled with ultraviolet and mass spectrometric detection. Phytochem. Anal. 2007, 18, 157–160. [Google Scholar] [CrossRef]
- Goldansaz, S.M.; Festa, C.; Pagano, E.; De Marino, S.; Finamore, C.; Parisi, O.A.; Borrelli, F.; Sonboli, A.; D’Auria, M.V. Phytochemical and Biological Studies of Nepeta asterotricha Rech. f. (Lamiaceae): Isolation of Nepetamoside. Molecules 2019, 24, 1684. [Google Scholar] [CrossRef] [Green Version]
- Jamzad, Z.; Chase, M.W.; Ingrouille, M.; Simmonds, M.S.J.; Jalili, A. Phylogenetic relationships in Nepeta L. (Lamiaceae) and related genera based on ITS sequence data. Taxon 2003, 52, 21–32. [Google Scholar] [CrossRef]
- Fraga, B.M.; González-Coloma, A.; Alegre-Gómez, S.; López-Rodríguez, M.; Amador, L.J.; Díaz, C.E. Bioactive constituents from transformed root cultures of Nepeta teydea. Phytochemistry 2017, 133, 59–68. [Google Scholar] [CrossRef]
- Miceli, N.; Taviano, M.F.; Giuffrida, D.; Trovato, A.; Tzakou, O.; Galati, E.M. Anti-inflammatory activity of extract and fractions from Nepeta sibthorpii Bentham. J. Ethnopharmacol. 2005, 97, 261–266. [Google Scholar] [CrossRef]
- Kumar, V.; Sood, H.; Sharma, M.; Chauhan, R.S. A proposed biosynthetic pathway of picrosides linked through the detection of biochemical intermediates in the endangered medicinal herb Picrorhiza kurroa. Phytochem. Anal. 2013, 24, 598–602. [Google Scholar] [CrossRef]
- Huang, X.H.; Chen, J.; Xu, X.Q.; Zhang, W.T.; Zhao, C.C. A new phenolic compound from Schizonepeta tenuifolia. Chem. Nat. Compd. 2016, 52, 1005–1007. [Google Scholar] [CrossRef]
- Aničić, N.; Matekalo, D.; Skorić, M.; Pećinar, I.; Brkušanin, M.; Nestorović Živković, J.; Dmitrović, S.; Dajić Stevanović, Z.; Schulz, H.; Mišić, D. Trichome-specific and developmentally regulated biosynthesis of nepetalactones in leaves of cultivated Nepeta rtanjensis plants. Ind. Crops Prod. 2018, 117, 347–358. [Google Scholar] [CrossRef]
- Xu, Z.; Xie, J.; Soteyome, T.; Peters, B.M.; Shirtliff, M.E.; Liu, J.; Harro, J.M. Polymicrobial interaction and biofilms between Staphylococcus aureus and Pseudomonas aeruginosa: An underestimated concern in food safety. Curr. Opin. Food Sci. 2019, 26, 57–64. [Google Scholar] [CrossRef]
- Smiljković, M.; Dias, M.I.; Stojković, D.; Barros, L.; Bukvički, D.; Ferreira, I.C.F.R.; Soković, M. Characterization of phenolic compounds in tincture of edible Nepeta nuda: Development of antimicrobial mouthwash. Food Funct. 2018, 9, 5417–5425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousavi Nia, A.; Kalantaripour, T.P.; Basiri, M.; Vafaee, F.; Asadi-Shekaari, M.; Eslami, A.; Zadeh, F.D. Nepeta dschuparensis Bornm. extract moderates COX-2 and IL-1β proteins in a rat model of cerebral ischemia. Iran. J. Med. Sci. 2017, 42, 179–186. [Google Scholar]
- Skorić, M.; Gligorijević, N.; Čavić, M.; Ristić, M.; Mišić, D.; Radulović, S. Cytotoxic activity of Nepeta rtanjensis Diklić and Milojević essential oil and and its mode of action. Ind. Crop. Prod. 2017, 100, 163–170. [Google Scholar] [CrossRef]
- Banjanac, T.; Dragićević, M.; Šiler, B.; Gašić, U.; Bohanec, B.; Nestorović Živković, J.; Trifunović, S.; Mišić, D. Chemodiversity of two closely related tetraploid Centaurium species and their hexaploid hybrid: Metabolomic search for high-resolution taxonomic classifiers. Phytochemistry 2017, 140, 27–44. [Google Scholar] [CrossRef]
- Čolić, S.D.; Fotirić Akšić, M.M.; Lazarević, K.B.; Zec, G.N.; Gašić, U.M.; Dabić Zagorac, D.; Natić, M.M. Fatty acid and phenolic profiles of almond grown in Serbia. Food Chem. 2017, 234, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Kartsev, V.; Lichitsky, B.; Geronikaki, A.; Petrou, A.; Smiljkovic, M.; Kostic, M.; Radanovic, O.; Soković, M. Design, synthesis and antimicrobial activity of usnic acid derivatives. MedChemComm 2019, 10, 180. [Google Scholar] [CrossRef] [Green Version]
- Soković, M.; van Griensven, L.J.L.D. Antimicrobial activity of essential oils and their components against the three major pathogens of the cultivated button mushroom, Agaricus bisporus. Eur. J. Plant Pathol. 2006, 116, 211–224. [Google Scholar] [CrossRef]
- Sirakanyan, S.; Kartsev, V.; Spinelli, D.; Geronikaki, A.; Petrou, A.; Ivanov, M.; Glamočlija, J.; Soković, M.; Hakobyan, E.; Hovakimyan, A. Synthesis and antimicrobial activity of new 2-piperazin-1-yl-N-1,3-thiazol-2-ylacetamides of cyclopenta[c]pyridines and pyrano[3,4-c]pyridines. Archiv Der Pharmazie 2021, 354, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nikolić, M.M.; Jovanović, K.K.; Marković, T.L.; Marković, D.L.; Gligorijević, N.N.; Radulović, S.S.; Kostić, M.; Glamočlija, J.M.; Soković, M.D. Antimicrobial synergism and cytotoxic properties of Citrus limon L., Piper nigrum L. and Melaleuca alternifolia (Maiden and Betche) Cheel essential oils. J. Pharm. Pharmacol. 2017, 69, 1606–1614. [Google Scholar] [CrossRef] [PubMed]
- Jevtić, B.; Djedović, N.; Stanisavljević, S.; Gašić, U.; Mišić, D.; Despotović, J.; Samardžić, J.; Miljković, D.; Timotijević, G. Anti-encephalitogenic effects of cucumber leaf extract. J. Funct. Foods 2017, 37, 249–262. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound Name | TR, Min | Molecular Formula, [M–H]− | Calculated Mass, [M–H]− | Exact Mass, [M–H]− | Δ mDa | MS2 Fragments, (% Base Peak) | MS3 Fragments, (% Base Peak) | MS4 Fragments, (% Base Peak) | NA | NR |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenolic Acid Derivatives | |||||||||||
| 1 | DihyDroxybenZoic Acid Hexoside | 5.76 | C13H15O9− | 315.07216 | 315.06858 | 3.58 | 108 (8), 109 (11), 152 (41), 153 (100), 163 (8), 165 (14), 269 (7) | 109 (100) | ND | + | + |
| 2 | Syringic Acid Hexoside | 6.17 | C15H19O10− | 359.09837 | 359.09405 | 4.32 | 182 (4), 197 (100), 198 (8), 313 (7) | 138 (8), 153 (18), 182 (100) | 137 (3), 138 (4), 167 (100) | + | – |
| 3 | 3-O-CafeOylquinic Acid | 6.38 | C16H17O9− | 353.08781 | 353.08319 | 4.62 | 135 (8), 179 (37), 191 (100), 192 (4) | 85 (100), 93 (52), 111 (32), 127 (91), 171 (21), 173 (59) | 57 (100) | + | + |
| 4 | DihyDroxybenZoic Acid Hexoside Isomer | 6.57 | C13H15O9− | 315.07216 | 315.06872 | 3.44 | 109 (5), 135 (4), 153 (100), 154 (7) | 109 (50), 135 (100) | 91 (100) | + | + |
| 5 | HydroxybenZoic Acid Hexoside | 6.80 | C13H15O8− | 299.07724 | 299.07395 | 3.29 | 93(3), 137 (100) | 93 (100) | ND | + | + |
| 6 | Caffeic Acid Hexoside | 6.88 | C15H17O9− | 341.08781 | 341.08369 | 4.12 | 135 (10), 179 (100), 180 (9), 181 (3), 251 (6), 281 (8), 293 (3) | 135 (100) | ND | + | + |
| 7 | Sinapic Acid a | 6.93 | C11H11O5− | 223.06120 | 223.05872 | 2.48 | 164 (22), 165 (3), 177 (12), 179 (39), 180 (6), 208 (100), 209 (11) | 135 (4), 149 (16), 163 (10), 164 (100), 193(8) | 134 (5), 135 (42), 149 (100) | + | + |
| 8 | TrihyDroxy-CinnamoylQuinic Acid | 7.03 | C16H17O10− | 369.08272 | 369.07675 | 5.97 | 149 (14), 173 (62), 191 (100), 193 (28), 194 (23), 195 (23), 245 (36) | 85 (100), 93 (71), 109 (29), 111 (29), 127 (92), 173 (64) | ND | + | + |
| 9 | 5-O-Café Oylquinic Acid a | 7.04 | C16H17O9− | 353.08781 | 353.08303 | 4.78 | 179 (9), 191 (100) | 85 (100), 93 (56), 109 (23), 111 (34), 127 (91), 173 (57) | 57 (100) | + | + |
| 10 | 5-O-CafeOylquinic Acid Isomer | 7.49 | C16H17O9− | 353.08781 | 353.08321 | 4.60 | 179 (7), 191 (100) | 85 (100), 93 (51), 111 (29), 127 (84), 171 (25), 173 (54) | 57 (100) | + | + |
| 11 | Danshensuan C | 7.53 | C18H17O9− | 377.08781 | 377.08261 | 5.20 | 359 (100), 360 (3) | 133 (5), 161 (100), 179 (19), 197 (21), 223 (8) | 133 (100) | – | + |
| 12 | 5-O-CaffeOylshikimic Acid | 7.74 | C16H15O8− | 335.07724 | 335.07371 | 3.53 | 135 (24), 161 (4), 179 (100), 180 (7) | 135 (100) | ND | + | + |
| 13 | Caffeic Acid a | 7.78 | C9H7O4− | 179.03498 | 179.03311 | 1.87 | 135 (100), 136 (14), 161 (7) | 79 (37), 107 (80), 117 (100) | ND | + | + |
| 14 | Feruloylquinic Acid | 8.01 | C17H19O9− | 367.10346 | 367.09822 | 5.24 | 173 (5), 191 (100), 192 (7), 193 (4) | 85 (100), 93 (49), 109 (29), 111 (41), 127 (78), 173 (66) | ND | + | – |
| 15 | DicaffeoyLquinic Acid | 8.67 | C25H23O12− | 515.11950 | 515.11446 | 5.04 | 173 (18), 179 (12), 191 (7), 203 (7), 335 (17), 353 (100), 354 (14) | 135 (13), 173 (100), 179 (73), 191 (46) | 59 (10), 93 (100), 111 (66), 127 (13), 155 (28) | + | – |
| 16 | Dicaffeoylquinic acid isomer | 9.07 | C25H23O12− | 515.11950 | 515.11437 | 5.13 | 173 (8), 179 (6), 203 (15), 255 (6), 299 (10), 353 (100), 354 (15) | 135 (11), 173 (100), 179 (66), 191 (38) | 71 (15), 93 (100), 109 (10), 111 (44), 155 (18) | – | + |
| 17 | Rosmarinic acid a | 9.33 | C18H15O8− | 359.07724 | 359.07265 | 4.59 | 133 (3), 161 (100), 162(6), 179 (14), 197 (14), 223 (6) | 133 (100) | 77 (41), 89 (29), 105 (100), 115 (40), 133 (60) | + | + |
| 18 | Nepetoidin A or B | 9.38 | C17H13O6− | 313.07176 | 313.06826 | 3.50 | 161 (100), 179 (10), 203 (10), 267 (15), 268 (16), 269 (26) | 133 (100) | ND | + | + |
| 19 | Clinopodic acid A | 10.04 | C18H15O7− | 343.08233 | 343.07838 | 3.95 | 135 (12), 145 (15), 161 (100), 179 (23), 197 (11), 297 (73), 298 (19) | 133 (100) | ND | + | + |
| 20 | Methyl rosmarinate | 10.21 | C19H17O8− | 373.09289 | 373.08847 | 4.42 | 135 (100), 161 (47), 175 (26), 179 (83), 197 (30), 311 (17), 355 (18) | 79 (26), 91 (24), 107 (57), 135 (100) | ND | + | + |
| Flavonoid Glycosides and Aglycones | |||||||||||
| 21 | Quercetin 3-O-rutinoside a | 8.17 | C27H29O16− | 609.14611 | 609.14136 | 4.75 | 179 (3), 255 (5), 271 (7), 300 (37), 301 (100), 343 (11), 463 (7) | 107 (7), 151 (77), 179 (100), 257 (12), 271 (19), 273 (20) | 151 (100) | + | + |
| 22 | Quercetin 3-O-glucoside a | 8.42 | C21H19O12− | 463.08820 | 463.08279 | 5.41 | 300 (22), 301 (100) | 107 (6), 151 (81), 179 (100), 257 (14), 271 (19), 273 (18) | 151 (100) | + | + |
| 23 | Luteolin 7-O-hexuronide | 8.50 | C21H17O12− | 461.07255 | 461.06743 | 5.12 | 285 (100), 286 (14), 327 (3), 357 (4), 381 (3) | 151 (36), 175 (86), 199 (68), 217 (67), 241 (100), 243 (56) | 185 (32), 197 (53), 198 (100), 199 (85), 213 (33) | + | + |
| 24 | Kaempferol 7-O-rutinoside | 8.51 | C27H29O15− | 593.15119 | 593.14990 | 1.29 | 229 (3), 257 (4), 285 (100), 286 (12), 327 (3), 547 (11), 548 (3) | 197 (26), 213 (30), 229 (52), 241 (43), 257 (100), 267 (52) | 163 (62), 185 (13), 213 (21), 229 (100), 239 (27) | + | + |
| 25 | Kaempferol 3-O-glucoside a | 8.84 | C21H19O11− | 447.09329 | 447.08843 | 4.85 | 255 (19), 284 (100), 285 (85), 327 (20) | 227 (14), 255 (100), 256 (21) | 211 (67), 227 (100), 255 (10) | + | + |
| 26 | Apigenin 7-O-glucoside a | 9.11 | C21H17O11− | 445.07764 | 445.07283 | 4.80 | 175 (13), 269 (100), 270 (14), 399 (9), 401 (4) | 149 (43), 151 (27), 181 (29), 197 (34), 201 (55), 225 (100) | ND | + | + |
| 27 | Thymusin | 11.91 | C17H13O7− | 329.06668 | 329.06286 | 3.82 | 299 (6), 314 (100), 315 (17), 316 (3) | 241 (4), 284 (3), 285 (45), 286 (4), 296 (6), 299 (100) | 199 (23), 227 (33), 243 (25), 255 (58), 271 (100) | + | + |
| 28 | Cirsimaritin a | 12.91 | C17H13O6− | 313.07176 | 313.06803 | 3.73 | 183 (25), 184 (3), 283 (6), 297 (3), 298 (100), 299 (14), 311 (3) | 150 (3), 225 (3), 268 (3), 269 (29), 270 (4), 283 (100) | 163 (8), 211 (5), 227 (9), 239 (6), 255 (100) | + | + |
| 29 | Ladanein | 13.22 | C17H13O6− | 313.07176 | 313.06810 | 3.66 | 183 (46), 184 (6), 198 (3), 283 (5), 298 (100), 299 (15), 311 (4) | 269 (20), 270 (3), 283 (100) | 163 (9), 211 (5), 227 (12), 239 (5), 255 (100) | + | + |
| 30 | Xanthomicrol | 13.55 | C18H15O7− | 343.08233 | 343.07811 | 4.22 | 313 (6), 328 (100), 329 (19), 330 (3) | 313 (100) | 193 (5), 270 (13), 285 (38), 295 (24), 298 (100) | + | + |
| 31 | Acacetin a | 13.81 | C16H11O5− | 283.06120 | 283.05807 | 3.13 | 268 (100), 269 (11) | 200 (6), 212 (6), 239 (18), 240 (49), 268 (100), 269 (21) | 172 (100), 196 (80), 211 (78), 212 (78), 240 (52) | + | + |
| 32 | Kumatakenin | 14.11 | C17H13O6− | 313.07176 | 313.06784 | 3.92 | 183 (59), 184 (8), 185 (3), 198 (5), 298 (100), 299 (16), 311 (5) | 255 (12), 270 (19), 283 (100), 284 (8) | 255 (100) | + | + |
| Iridoid Glycosides | |||||||||||
| 33 | Ajugol | 6.21 | C15H23O9− | 347.13476 | 347.13036 | 4.40 | 123 (9), 167 (47), 185 (100), 186 (7), 281 (7), 308 (6), 310 (8) | 71 (18), 123 (27), 125 (10), 139 (100), 141 (86), 167 (86) | 81 (15), 95 (100), 110 (17), 121 (69), 124 (31) | + | + |
| 34 | Bartsioside | 6.24 | C15H21O8− | 329.12419 | 329.11905 | 5.14 | 129 (8), 156 (8), 161 (23), 167 (100), 285 (23), 299 (9) | 149 (100), 123 (10), 121 (15) | ND | + | + |
| 35 | Lamiol | 6.34 | C16H25O10− | 377.14532 | 377.14004 | 5.28 | 151 (9), 153 (100), 179 (7), 195 (7), 197 (63), 201 (8), 215 (9) | 109 (100), 135 (7) | ND | – | + |
| 36 | Geniposide isomer | 6.68 | C17H23O10− | 387.12967 | 387.12505 | 4.62 | 179 (100), 180 (13), 207 (11), 225 (50), 258 (8), 340 (14), 341 (33) | 146 (22), 161 (100), 164 (68) | 146 (100) | + | + |
| 37 | 6-O-Deoxylamioside | 7.20 | C18H27O10− | 403.16097 | 403.15625 | 4.72 | 137 (47), 181 (21), 191 (21), 199 (48), 247 (20), 343 (100), 361 (73) | 113 (4), 119 (5), 137 (100), 181 (44), 199 (55), 299 (3) | 80 (3), 107 (3), 119 (100) | + | + |
| 38 | Geniposide | 7.50 | C17H23O10− | 387.12967 | 387.12571 | 3.96 | 161 (100), 163 (3), 207 (5) | 105 (3), 133 (95), 143 (3), 146 (100), 161 (5) | 118 (100) | + | + |
| 39 | 1-O-Hexosyl-epideoxyloganic acid | 7.86 | C22H33O14− | 521.18758 | 521.18257 | 5.01 | 153 (18), 197 (92), 341 (50), 359 (100), 475 (34), 476 (29), 477 (23) | 109 (11), 135 (7), 153 (17), 197 (100), 315 (6) | 87 (5), 109 (22), 135 (6), 153 (100) | + | + |
| 40 | Epi-deoxyloganic acid isomer | 7.92 | C16H23O9− | 359.13476 | 359.12981 | 4.95 | 109 (5), 135 (24), 136 (5), 153 (63), 154 (9), 197 (100), 198 (16) | 109 (27), 135 (8), 153 (100) | 135 (100) | + | + |
| 41 | 1,5,9-Epi-deoxyloganic acida | 8.13 | C16H23O9− | 359.13476 | 359.12960 | 5.16 | 109 (3), 135 (20), 153 (49), 197 (100) | 109 (28), 135 (6), 153 (100) | 135 (100) | + | + |
| 42 | Boschnaloside | 8.25 | C16H23O8− | 343.13984 | 343.13512 | 4.72 | 101 (7), 113 (15), 143 (5), 161 (100), 181 (12) | 71(35), 87(20), 101 (100), 113(51), 143(15) | ND | + | + |
| 43 | Deoxyloganetic acid pentoside | 8.26 | C15H21O8− | 329.12419 | 329.12048 | 3.71 | 149 (9), 153 (44), 197 (100), 198 (10), 285 (16) | 109(9), 135(7), 153 (100) | 107 (11), 135 (100) | + | + |
| 44 | Aucubin | 8.43 | C16H25O8− | 345.15549 | 345.15103 | 4.46 | 101 (12), 113 (20), 119 (17), 143 (10), 161 (9), 183 (100) | 57 (33), 107 (6), 125 (34), 139 (65), 155 (17), 165 (100) | 55 (48), 95 (85), 107 (100), 109 (23), 137 (48) | + | + |
| 45 | Nepetariaside | 8.73 | C16H27O8− | 347.17114 | 347.16679 | 4.35 | 101 (40), 113 (47), 119 (50), 143 (16), 167 (75), 185 (100), 329 (17) | 57 (5), 139 (85), 141 (10), 167 (100) | 57 (92), 109 (100), 124 (3), 125 (5), 137 (7) | + | + |
| Other Compounds | |||||||||||
| 46 | Quinic acida | 1.59 | C7H11O6− | 191.05611 | 191.05418 | 1.93 | 173 (29), 171 (49), 153 (13), 127 (97), 111 (45), 93 (58), 85 (100) | 57 (100) | ND | + | + |
| 47 | 3,4-dihydroxyphenethyl alcohol 4-O-hexoside | 6.02 | C14H19O8− | 315.10854 | 315.10459 | 3.95 | 123 (8), 153 (100), 154 (7), 269 (4) | 109 (5), 123 (100) | 81 (11), 93 (12), 95 (100), 105 (46), 123 (6) | + | + |
| 48 | 12-O-HexOsyl-Jasmonate | 7.40 | C18H27O9− | 387.16606 | 387.16069 | 5.37 | 113 (4), 163 (73), 164 (8), 207 (100), 208 (10), 225 (5), 369 (16) | 163 (100) | 107 (18), 109 (100), 121 (4), 145 (8), 147 (8) | + | + |
| 49 | UmbelLiferone a | 9.33 | C9H5O3− | 161.02442 | 161.02294 | 1.48 | 113 (38), 115 (82), 119 (56), 131 (32), 133 (100), 134 (60), 141 (35) | 89 (7), 90 (4), 92 (3), 105 (18), 106 (4), 115 (100) | ND | + | + |
| Antibacterial Activity | N. rtanjensis | N. argolica | cis,trans-NL | trans,cis-NL | RA | 1,5,9-eDLA | Streptomycin |
|---|---|---|---|---|---|---|---|
| MIC-MBC | MIC-MBC | MIC-MBC | MIC-MBC | MIC-MBC | MIC-MBC | MIC-MBC | |
| [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | |
| B. cereus | 0.2000–0.4000 | 0.2000–0.4000 | 0.0400–0.0800 | 0.0800–0.1200 | 0.0100–0.0200 | 0.0600–0.0800 | 0.1000–0.2000 |
| S. aureus | 0.1500–0.2000 | 0.2000-0.4000 | 0.0100–0.0200 | 0.0100–0.0400 | 0.0075–0.0100 | 0.0050–0.0100 | 0.0500–0.1000 |
| M. flavus | 0.2000–0.4000 | 0.1500–0.4000 | 0.0300–0.0400 | 0.0300–0.0400 | 0.0300–0.040 | 0.6000–0.8000 | 0.1000–0.2000 |
| L. monicytogenes | 0.1500–0.2000 | 0.2000–0.4000 | 0.0025–0.0050 | 0.0100–0.0200 | 0.0150–0.0200 | 0.0200–0.0400 | 0.2000–0.3000 |
| E. faecalis | 0.4000–0.6000 | 0.3000–0.4000 | 0.01500–0.0400 | 0.0400–0.0800 | 0.0400–0.0800 | 0.0300–0.0400 | 0.1000–0.2000 |
| P. aeruginosa | 0.1000–0.2000 | 0.3000–0.4000 | 0.0100–0.0200 | 0.0300–0.0400 | 0.0100–0.0200 | 0.0400–0.0600 | 0.2000–0.3000 |
| E. coli | 0.2000–0.4000 | 0.1500–0.2000 | 0.0200–0.0400 | 0.0200–0.0400 | 0.0300–0.0400 | 0.0200–0.0400 | 0.1000–0.2000 |
| S. typhimurium | 0.2000–0.4000 | 0.2000–0.4000 | 0.0025–0.0050 | 0.0400–0.0800 | 0.0300–0.0400 | 0.0050–0.0100 | 0.2000–0.3000 |
| Antifungal Activity | N. rtanjensis | N. argolica | cis,trans-NL | trans,cis-NL | RA | 1,5,9-eDLA | Ketoconazole |
| MIC-MFC | MIC-MFC | MIC-MFC | MIC-MFC | MIC-MFC | MIC-MFC | MIC-MFC | |
| [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | [mg mL−1] | |
| A. fumigatus | 0.1000–0.2000 | 0.1000–0.2000 | 0.0100–0.0200 | 0.0100–0.0200 | 0.0200–0.0400 | 0.0400–0.0600 | 0.2500–0.5000 |
| A. ochraceus | 0.1000–0.2000 | 0.1000–0.2000 | 0.0050–0.0200 | 0.0025–0.005 | 0.0050–0.0100 | 0.0100–0.0200 | 0.1500–0.2000 |
| A. versicolor | 0.1000–0.2000 | 0.1000–0.4000 | 0.0200–0.0300 | 0.0050–0.0100 | 0.0100–0.0200 | 0.0400–0.0600 | 0.2000–0.5000 |
| A. niger | 0.0500–0.1000 | 0.1000–0.4000 | 0.0050–0.0200 | 0.0050–0.0200 | 0.0075–0.0100 | 0.0150–0.0400 | 0.2000–0.5000 |
| A. funiculosum | 0.2000–0.4000 | 0.2000–0.4000 | 0.0200–0.0300 | 0.0050–0.0100 | 0.0050–0.0100 | 0.0050–0.0400 | 0.2000–0.5000 |
| P. ochlochloron | 0.2000–0.4000 | 0.3000–0.4000 | 0.0050–0.0100 | 0.0050–0.0100 | 0.0100–0.0200 | 0.0600–0.0800 | 1.0000–1.5000 |
| P. v. cylopium | 0.3000–0.4000 | 0.3000–0.4000 | 0.0100–0.0300 | 0.0100–0.0200 | 0.0100–0.0200 | 0.0600–0.0800 | 0.2000–0.5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aničić, N.; Gašić, U.; Lu, F.; Ćirić, A.; Ivanov, M.; Jevtić, B.; Dimitrijević, M.; Anđelković, B.; Skorić, M.; Nestorović Živković, J.; et al. Antimicrobial and Immunomodulating Activities of Two Endemic Nepeta Species and Their Major Iridoids Isolated from Natural Sources. Pharmaceuticals 2021, 14, 414. https://doi.org/10.3390/ph14050414
Aničić N, Gašić U, Lu F, Ćirić A, Ivanov M, Jevtić B, Dimitrijević M, Anđelković B, Skorić M, Nestorović Živković J, et al. Antimicrobial and Immunomodulating Activities of Two Endemic Nepeta Species and Their Major Iridoids Isolated from Natural Sources. Pharmaceuticals. 2021; 14(5):414. https://doi.org/10.3390/ph14050414
Chicago/Turabian StyleAničić, Neda, Uroš Gašić, Feng Lu, Ana Ćirić, Marija Ivanov, Bojan Jevtić, Milena Dimitrijević, Boban Anđelković, Marijana Skorić, Jasmina Nestorović Živković, and et al. 2021. "Antimicrobial and Immunomodulating Activities of Two Endemic Nepeta Species and Their Major Iridoids Isolated from Natural Sources" Pharmaceuticals 14, no. 5: 414. https://doi.org/10.3390/ph14050414