Short-Chain Fatty Acids Promote Mycobacterium avium subsp. hominissuis Growth in Nutrient-Limited Environments and Influence Susceptibility to Antibiotics

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Culture
2.2. Static Biofilm Formation
2.3. Screening the Capacity of MAH104 Cells to Utilize a Range of Metabolites in Planktonic and Biofilm States
2.4. Testing the Effect of SCFA and Glycerol on Growth of MAH104 Planktonic Cultures in Nutrient-Restricted Media
2.5. Testing the Effect of SCFA and Glycerol on MAH104 Biofilm Cultures
2.6. Antibiotic Treatment of MAH Biofilms
2.7. MAH104 Infection and Antibiotic Treatment of Human Macrophages
2.8. Statistical Analysis
3. Results
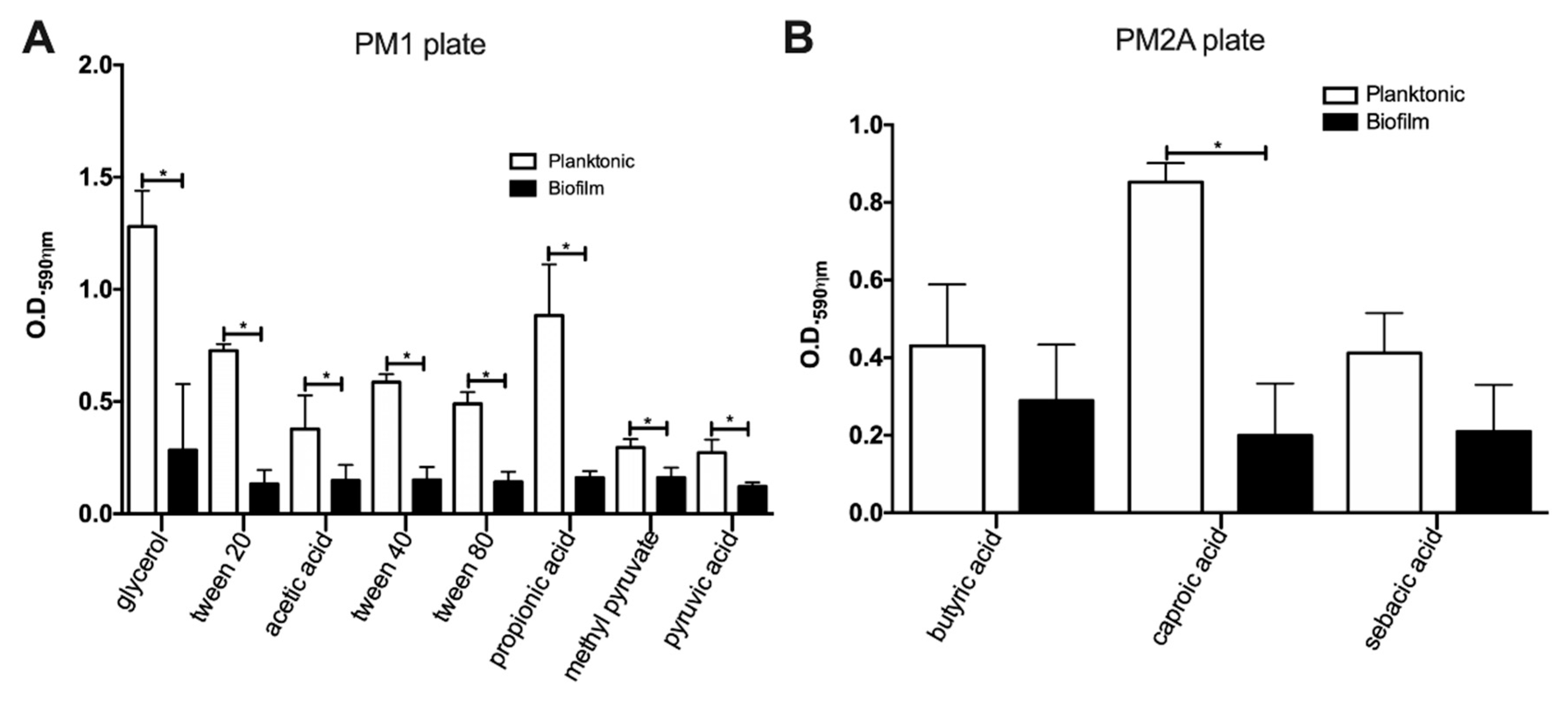
3.1. MAH104 Displays a Decreased Capacity for Processing of Carbon Substrates in Biofilms
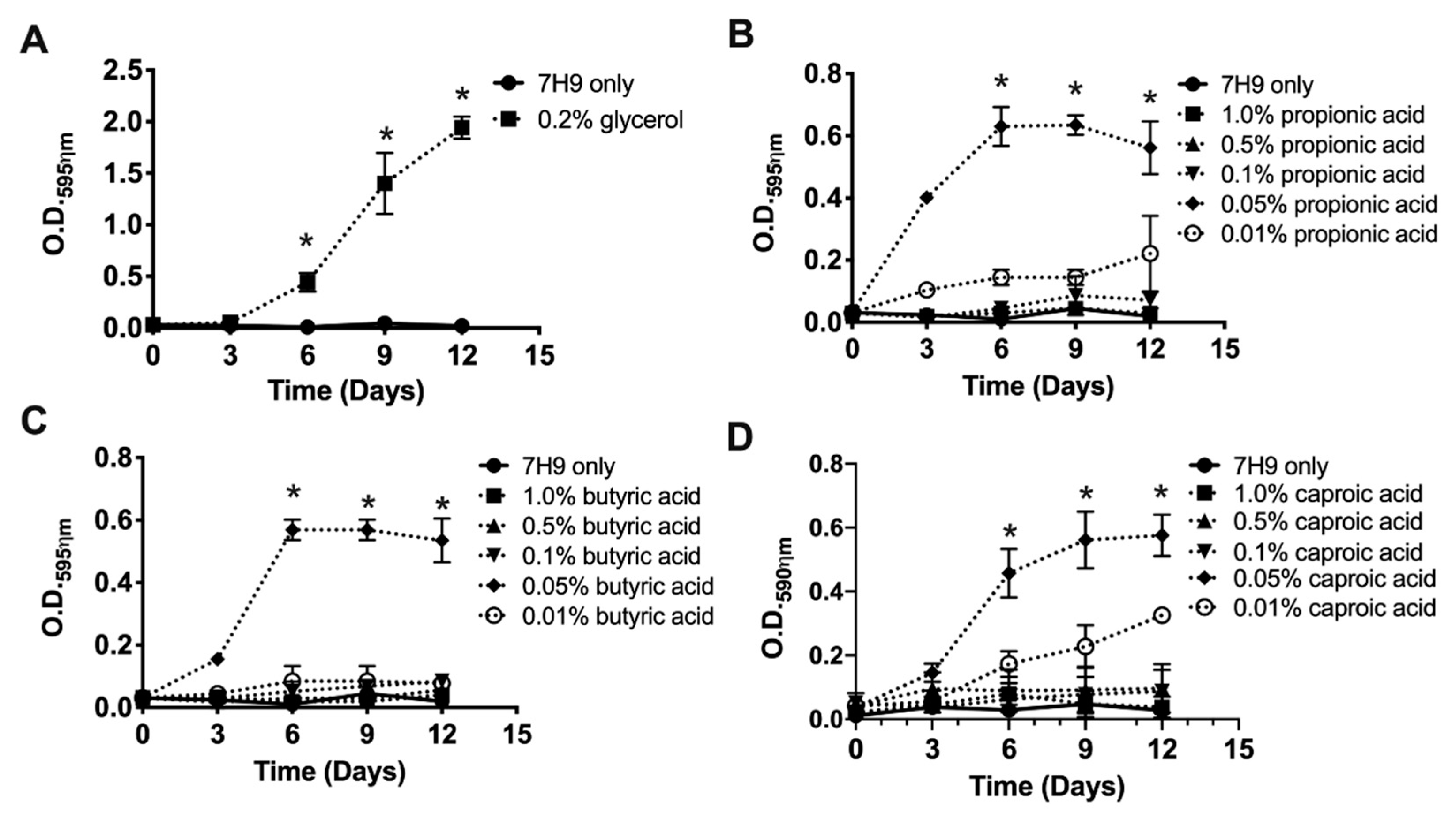
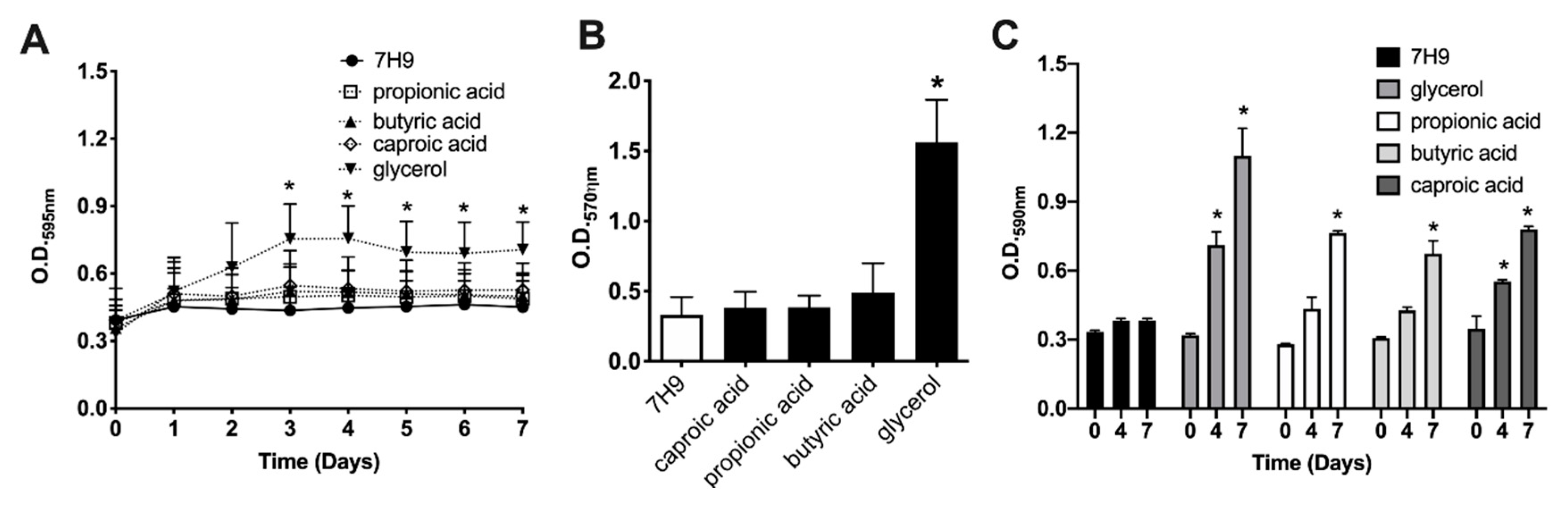
3.2. Glycerol and SCFA Support the Growth of Planktonic MAH104 in Nutrient-Limited Media
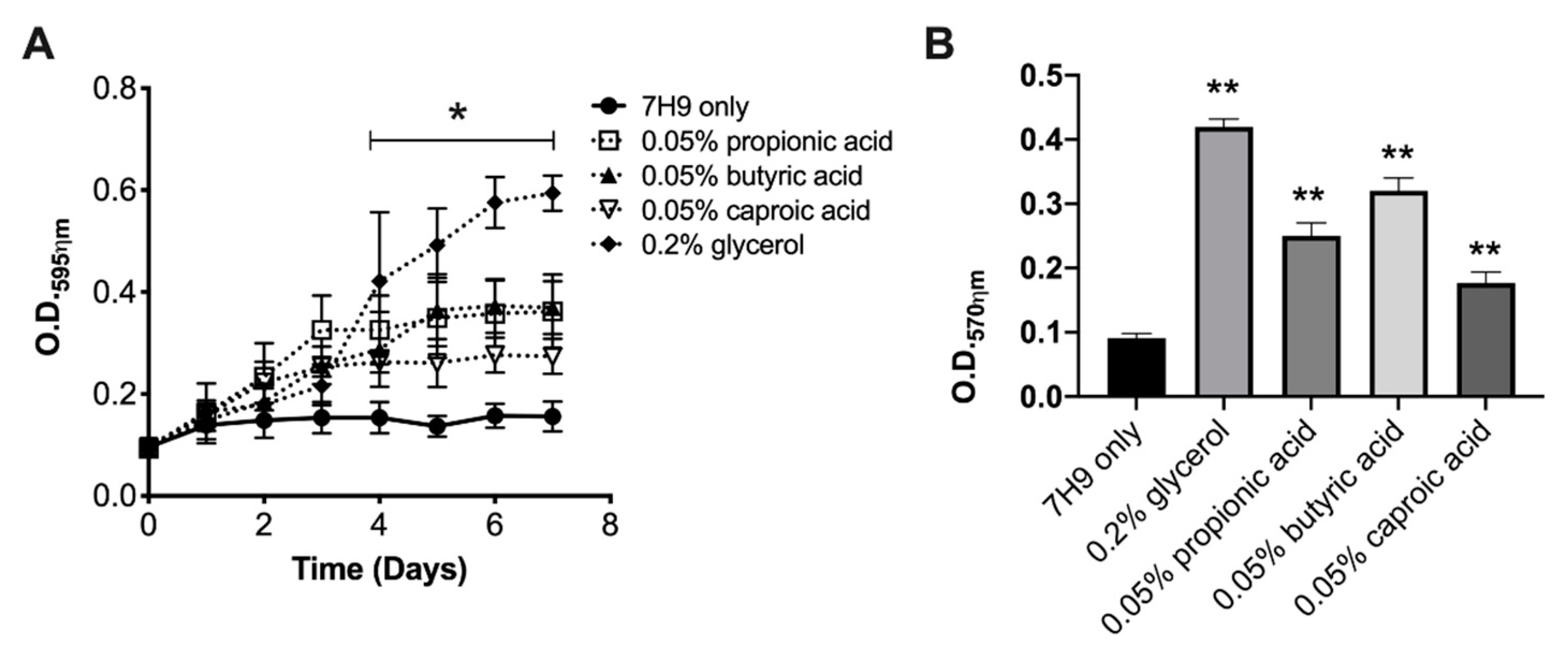
3.3. The Sessile MAH104 of Planktonic State Displays Higher Capacity for Growth and Biofilm Formation when Incubated with Glycerol or SCFA
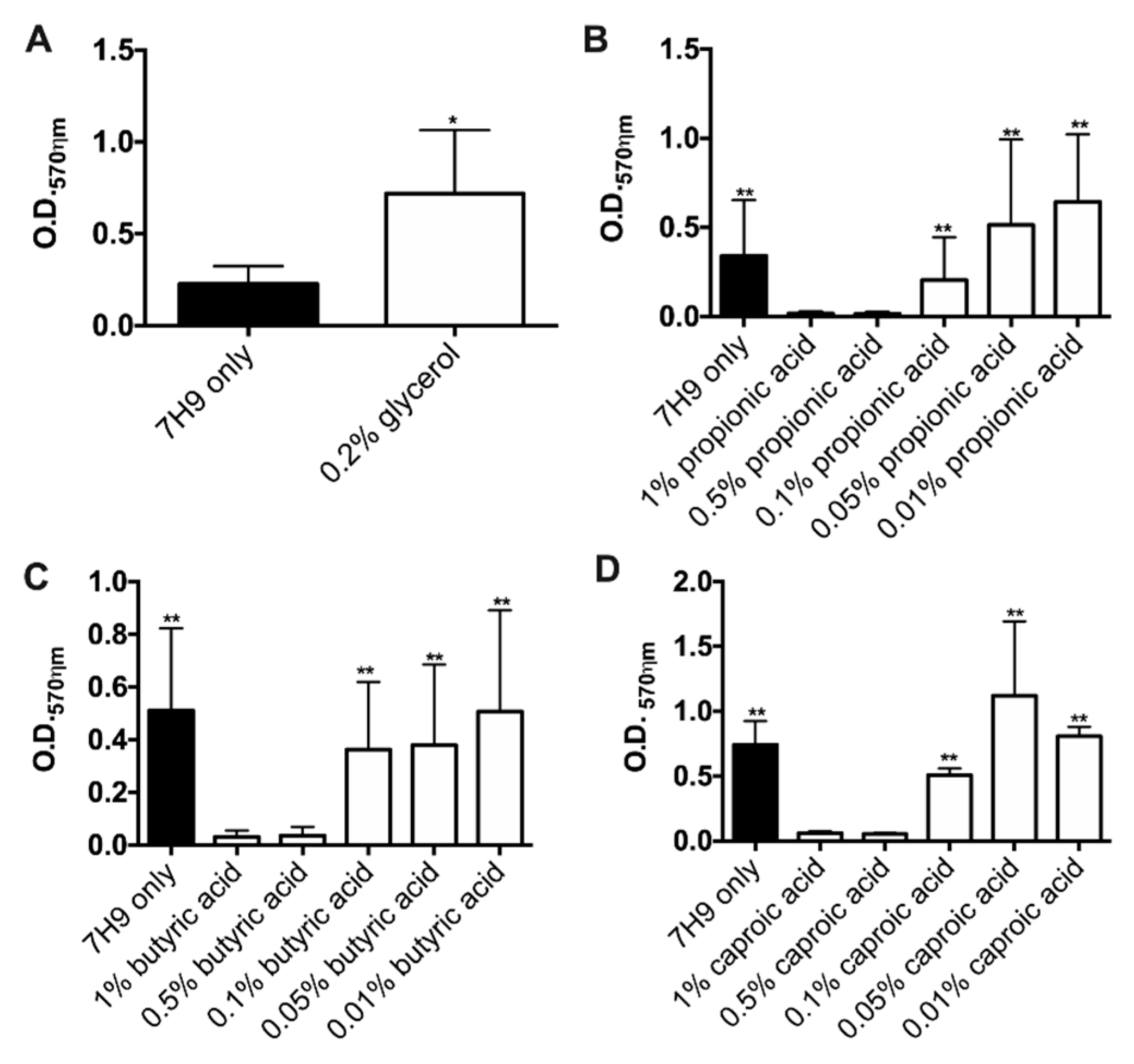
3.4. Glycerol but Not SCFA Promote MAH104 Growth in Biofilms
3.5. Glycerol and SCFA Enhance Antibiotic Efficacy against MAH104 in Biofilms and in Cultured Macrophages
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bodle, E.E.; Cunningham, J.A.; Della-Latta, P.; Schluger, N.W.; Saiman, L. Epidemiology of nontuberculous mycobacteria in patients without HIV infection, New York City. Emerg. Infect. Dis. 2008, 14, 390–396. [Google Scholar] [CrossRef]
- Honda, J.R.; Knight, V.; Chan, E.D. Pathogenesis and risk factors for nontuberculous mycobacterial lung disease. Clin. Chest Med. 2015, 36, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Catherinot, E.; Roux, A.L.; Vibet, M.A.; Bellis, G.; Ravilly, S.; Lemonnier, L.; Le Roux, E.; Bernède-Bauduin, C.; Le Bourgeois, M.; Herrmann, J.L.; et al. Mycobacterium avium and Mycobacterium abscessus complex target distinct cystic fibrosis patient subpopulations. J. Cyst. Fibros. Off. J. Eur. Cyst. Fibros. Soc. 2013, 12, 74–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnatunga, C.N.; Lutzky, V.P.; Kupz, A.; Doolan, D.L.; Reid, D.W.; Field, M.; Bell, S.C.; Thomson, R.M.; Miles, J.J. The Rise of Non-Tuberculosis Mycobacterial Lung Disease. Front. Immunol. 2020, 11, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwak, N.; Park, J.; Kim, E.; Lee, C.H.; Han, S.K.; Yim, J.J. Treatment Outcomes of Mycobacterium avium Complex Lung Disease: A Systematic Review and Meta-analysis. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 65, 1077–1084. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Koh, W.J.; Daley, C.L. Treatment of Mycobacterium avium Complex Pulmonary Disease. Tuberc. Respir. Dis. 2019, 82, 15–26. [Google Scholar] [CrossRef]
- Field, S.K.; Fisher, D.; Cowie, R.L. Mycobacterium avium complex pulmonary disease in patients without HIV infection. Chest 2004, 126, 566–581. [Google Scholar] [CrossRef] [PubMed]
- Banaschewski, B.; Hofmann, T. Inhaled Antibiotics for Mycobacterial Lung Disease. Pharmaceutics 2019, 11, 352. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, Y.; Danelishvili, L.; Wu, M.; Hidaka, E.; Katsuyama, T.; Stang, B.; Petrofsky, M.; Bildfell, R.; Bermudez, L.E. The ability to form biofilm influences Mycobacterium avium invasion and translocation of bronchial epithelial cells. Cell. Microbiol. 2006, 8, 806–814. [Google Scholar] [CrossRef]
- Babrak, L.; Danelishvili, L.; Rose, S.J.; Kornberg, T.; Bermudez, L.E. The environment of “Mycobacterium avium subsp. Hominissuis” microaggregates induces synthesis of small proteins associated with efficient infection of respiratory epithelial cells. Infect. Immun. 2015, 83, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Balcazar, J.L.; Subirats, J.; Borrego, C.M. The role of biofilms as environmental reservoirs of antibiotic resistance. Front. Microbiol. 2015, 6, 1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babrak, L.; Danelishvili, L.; Rose, S.J.; Bermudez, L.E. Microaggregate-associated protein involved in invasion of epithelial cells by Mycobacterium avium subsp. hominissuis. Virulence 2015, 6, 694–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.J.; Bermudez, L.E. Identification of Bicarbonate as a Trigger and Genes Involved with Extracellular DNA Export in Mycobacterial Biofilms. mBio 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.J.; Bermudez, L.E. Mycobacterium avium biofilm attenuates mononuclear phagocyte function by triggering hyperstimulation and apoptosis during early infection. Infect. Immun. 2014, 82, 405–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojony, R.; Martin, M.; Campeau, A.; Wozniak, J.M.; Gonzalez, D.J.; Jaiswal, P.; Danelishvili, L.; Bermudez, L.E. Quantitative analysis of Mycobacterium avium subsp. hominissuis proteome in response to antibiotics and during exposure to different environmental conditions. Clin. Proteom. 2019, 16, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, S.J.; Babrak, L.M.; Bermudez, L.E. Mycobacterium avium Possesses Extracellular DNA that Contributes to Biofilm Formation, Structural Integrity, and Tolerance to Antibiotics. PLoS ONE 2015, 10, e0128772. [Google Scholar] [CrossRef] [Green Version]
- Gollan, B.; Grabe, G.; Michaux, C.; Helaine, S. Bacterial Persisters and Infection: Past, Present, and Progressing. Annu. Rev. Microbiol. 2019, 73, 359–385. [Google Scholar] [CrossRef]
- Anderl, J.N.; Zahller, J.; Roe, F.; Stewart, P.S. Role of nutrient limitation and stationary-phase existence in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrob. Agents Chemother. 2003, 47, 1251–1256. [Google Scholar] [CrossRef] [Green Version]
- Greendyke, R.; Byrd, T.F. Differential antibiotic susceptibility of Mycobacterium abscessus variants in biofilms and macrophages compared to that of planktonic bacteria. Antimicrob. Agents Chemother. 2008, 52, 2019–2026. [Google Scholar] [CrossRef] [Green Version]
- Walters, M.C., 3rd; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef] [Green Version]
- Borriello, G.; Werner, E.; Roe, F.; Kim, A.M.; Ehrlich, G.D.; Stewart, P.S. Oxygen limitation contributes to antibiotic tolerance of Pseudomonas aeruginosa in biofilms. Antimicrob. Agents Chemother. 2004, 48, 2659–2664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, E.M.; Rowe, S.E.; O’Gara, J.P.; Conlon, B.P. Convergence of Staphylococcus aureus Persister and Biofilm Research: Can Biofilms Be Defined as Communities of Adherent Persister Cells? PLoS Pathog. 2016, 12, e1006012. [Google Scholar] [CrossRef] [PubMed]
- Pontes, M.H.; Groisman, E.A. Slow growth determines nonheritable antibiotic resistance in Salmonella enterica. Sci. Signal. 2019. [Google Scholar] [CrossRef] [PubMed]
- Archuleta, R.J.; Yvonne Hoppes, P.; Primm, T.P. Mycobacterium avium enters a state of metabolic dormancy in response to starvation. Tuberculosis (Edinb. Scotl.) 2005, 85, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Mayhall, C.G.; Apollo, E. Effect of storage and changes in bacterial growth phase and antibiotic concentrations on antimicrobial tolerance in Staphylococcus aureus. Antimicrob. Agents Chemother. 1980, 18, 784–788. [Google Scholar] [CrossRef] [Green Version]
- McKay, S.L.; Portnoy, D.A. Ribosome hibernation facilitates tolerance of stationary-phase bacteria to aminoglycosides. Antimicrob. Agents Chemother. 2015, 59, 6992–6999. [Google Scholar] [CrossRef] [Green Version]
- Martins, D.; McKay, G.; Sampathkumar, G.; Khakimova, M.; English, A.M.; Nguyen, D. Superoxide dismutase activity confers (p)ppGpp-mediated antibiotic tolerance to stationary-phase Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2018, 115, 9797–9802. [Google Scholar] [CrossRef] [Green Version]
- Weiss, G.; Schaible, U.E. Macrophage defense mechanisms against intracellular bacteria. Immunol. Rev. 2015, 264, 182–203. [Google Scholar] [CrossRef] [Green Version]
- Kedia-Mehta, N.; Finlay, D.K. Competition for nutrients and its role in controlling immune responses. Nat. Commun. 2019, 10, 2123. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Adams, K.N.; Takaki, K.; Connolly, L.E.; Wiedenhoft, H.; Winglee, K.; Humbert, O.; Edelstein, P.H.; Cosma, C.L.; Ramakrishnan, L. Drug tolerance in replicating mycobacteria mediated by a macrophage-induced efflux mechanism. Cell 2011, 145, 39–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobritz, M.A.; Belenky, P.; Porter, C.B.; Gutierrez, A.; Yang, J.H.; Schwarz, E.G.; Dwyer, D.J.; Khalil, A.S.; Collins, J.J. Antibiotic efficacy is linked to bacterial cellular respiration. Proc. Natl. Acad. Sci. USA 2015, 112, 8173–8180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micheva-Viteva, S.N.; Ross, B.N.; Gao, J.; Adikari, S.; Zhang, P.; Mourant, J.R.; Wu, T.H.; Werner, J.H.; Torres, A.G.; Hong-Geller, E. Increased Mortality in Mice following Immunoprophylaxis Therapy with High Dosage of Nicotinamide in Burkholderia Persistent Infections. Infect. Immun. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allison, K.R.; Brynildsen, M.P.; Collins, J.J. Metabolite-enabled eradication of bacterial persisters by aminoglycosides. Nature 2011, 473, 216–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meylan, S.; Porter, C.B.M.; Yang, J.H.; Belenky, P.; Gutierrez, A.; Lobritz, M.A.; Park, J.; Kim, S.H.; Moskowitz, S.M.; Collins, J.J. Carbon Sources Tune Antibiotic Susceptibility in Pseudomonas aeruginosa via Tricarboxylic Acid Cycle Control. Cell Chem. Biol. 2017, 24, 195–206. [Google Scholar] [CrossRef]
- Barraud, N.; Buson, A.; Jarolimek, W.; Rice, S.A. Mannitol enhances antibiotic sensitivity of persister bacteria in Pseudomonas aeruginosa biofilms. PLoS ONE 2013, 8, e84220. [Google Scholar] [CrossRef]
- Vilcheze, C.; Hartman, T.; Weinrick, B.; Jain, P.; Weisbrod, T.R.; Leung, L.W.; Freundlich, J.S.; Jacobs, W.R., Jr. Enhanced respiration prevents drug tolerance and drug resistance in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2017, 114, 4495–4500. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.; Jain, S.; Bhargava, P.; Hamblin, M.; Lobritz, M.A.; Collins, J.J. Understanding and Sensitizing Density-Dependent Persistence to Quinolone Antibiotics. Mol. Cell 2017, 68, 1147–1154.e1143. [Google Scholar] [CrossRef] [Green Version]
- Carter, G.; Wu, M.; Drummond, D.C.; Bermudez, L.E. Characterization of biofilm formation by clinical isolates of Mycobacterium avium. J. Med. Microbiol. 2003, 52, 747–752. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Jezek, P.; Pavlovic, M.; Svabic-Vlahovic, M. Influence of dynamic conditions on biofilm formation by staphylococci. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2001, 20, 502–504. [Google Scholar] [CrossRef]
- Sanchini, A.; Dematheis, F.; Semmler, T.; Lewin, A. Metabolic phenotype of clinical and environmental Mycobacterium avium subsp. hominissuis isolates. PeerJ 2017, 5, e2833. [Google Scholar] [CrossRef] [PubMed]
- Nash, K.A.; Brown-Elliott, B.A.; Wallace, R.J., Jr. A novel gene, erm(41), confers inducible macrolide resistance to clinical isolates of Mycobacterium abscessus but is absent from Mycobacterium chelonae. Antimicrob. Agents Chemother. 2009, 53, 1367–1376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenito, J.; Wu, B.; Sulaiman, I.; Kurz, S.G.; Li, Y.; Perez, L.; Franca, B.; Olsen, E.; Gonzalez, A.; Yie, K.; et al. Microbial Short Chain Fatty Acids Impair Mycobacterium Avium (MAC) Clearance by Alveolar Macrophages. In Proceedings of the American Thoracic Society 2019 International Conference, Dallas, TX, USA, 17–22 May 2019; p. A4242. [Google Scholar]
- Hicks, N.D.; Yang, J.; Zhang, X.; Zhao, B.; Grad, Y.H.; Liu, L.; Ou, X.; Chang, Z.; Xia, H.; Zhou, Y.; et al. Clinically prevalent mutations in Mycobacterium tuberculosis alter propionate metabolism and mediate multidrug tolerance. Nat. Microbiol. 2018, 3, 1032–1042. [Google Scholar] [CrossRef] [PubMed]
- Lachmandas, E.; van den Heuvel, C.N.; Damen, M.S.; Cleophas, M.C.; Netea, M.G.; van Crevel, R. Diabetes Mellitus and Increased Tuberculosis Susceptibility: The Role of Short-Chain Fatty Acids. J. Diabetes Res. 2016, 2016, 6014631. [Google Scholar] [CrossRef]
- Maji, A.; Misra, R.; Dhakan, D.B.; Gupta, V.; Mahato, N.K.; Saxena, R.; Mittal, P.; Thukral, N.; Sharma, E.; Singh, A.; et al. Gut microbiome contributes to impairment of immunity in pulmonary tuberculosis patients by alteration of butyrate and propionate producers. Environ. Microbiol. 2018, 20, 402–419. [Google Scholar] [CrossRef]
- Segal, L.N.; Clemente, J.C.; Li, Y.; Ruan, C.; Cao, J.; Danckers, M.; Morris, A.; Tapyrik, S.; Wu, B.G.; Diaz, P.; et al. Anaerobic Bacterial Fermentation Products Increase Tuberculosis Risk in Antiretroviral-Drug-Treated HIV Patients. Cell Host Microbe 2017, 21, 530–537.e534. [Google Scholar] [CrossRef] [Green Version]
- Goring, S.M.; Wilson, J.B.; Risebrough, N.R.; Gallagher, J.; Carroll, S.; Heap, K.J.; Obradovic, M.; Loebinger, M.R.; Diel, R. The cost of Mycobacterium avium complex lung disease in Canada, France, Germany, and the United Kingdom: A nationally representative observational study. BMC Health Serv. Res. 2018, 18, 700. [Google Scholar] [CrossRef] [Green Version]
- Faria, S.; Joao, I.; Jordao, L. General Overview on Nontuberculous Mycobacteria, Biofilms, and Human Infection. J. Pathog. 2015, 2015, 809014. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.H.; Falkinham, J.O., 3rd. Microaerobic growth and anaerobic survival of Mycobacterium avium, Mycobacterium intracellulare and Mycobacterium scrofulaceum. Int. J. Mycobacteriology 2015, 4, 25–30. [Google Scholar] [CrossRef] [Green Version]
- Esteban, J.; García-Coca, M. Mycobacterium Biofilms. Front. Microbiol. 2017, 8, 2651. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.X.; Shen, M.J.; Liu, W.B.; Ye, B.C. GlnR-Mediated Regulation of Short-Chain Fatty Acid Assimilation in Mycobacterium smegmatis. Front. Microbiol. 2018, 9, 1311. [Google Scholar] [CrossRef] [PubMed]
- Camargo, E.E.; Wagner, H.N., Jr. Radiometric studies on the oxidation of [1-14C]fatty acids and [U-14C]L-amino acids by mycobacteria. Int. J. Radiat. Appl. Instrum. Part B Nucl. Med. Biol. 1987, 14, 43–49. [Google Scholar] [CrossRef]
- Hampshire, T.; Soneji, S.; Bacon, J.; James, B.W.; Hinds, J.; Laing, K.; Stabler, R.A.; Marsh, P.D.; Butcher, P.D. Stationary phase gene expression of Mycobacterium tuberculosis following a progressive nutrient depletion: A model for persistent organisms? Tuberculosis (Edinb. Scotl.) 2004, 84, 228–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhargava, P.; Collins, J.J. Boosting bacterial metabolism to combat antibiotic resistance. Cell Metab. 2015, 21, 154–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, B.; Su, Y.B.; Li, H.; Han, Y.; Guo, C.; Tian, Y.M.; Peng, X.X. Exogenous alanine and/or glucose plus kanamycin kills antibiotic-resistant bacteria. Cell Metab. 2015, 21, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Fux, C.A.; Wilson, S.; Stoodley, P. Detachment characteristics and oxacillin resistance of Staphyloccocus aureus biofilm emboli in an in vitro catheter infection model. J. Bacteriol. 2004, 186, 4486–4491. [Google Scholar] [CrossRef] [Green Version]
- Gradelski, E.; Kolek, B.; Bonner, D.; Fung-Tomc, J. Bactericidal mechanism of gatifloxacin compared with other quinolones. J. Antimicrob. Chemother. 2002, 49, 185–188. [Google Scholar] [CrossRef] [Green Version]
- Herbert, D.; Paramasivan, C.N.; Venkatesan, P.; Kubendiran, G.; Prabhakar, R.; Mitchison, D.A. Bactericidal action of ofloxacin, sulbactam-ampicillin, rifampin, and isoniazid on logarithmic- and stationary-phase cultures of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 1996, 40, 2296–2299. [Google Scholar] [CrossRef] [Green Version]
- Rojony, R.; Danelishvili, L.; Campeau, A.; Wozniak, J.M.; Gonzalez, D.J.; Bermudez, L.E. Exposure of Mycobacterium abscessus to Environmental Stress and Clinically Used Antibiotics Reveals Common Proteome Response among Pathogenic Mycobacteria. Microorganisms 2020, 8, 698. [Google Scholar] [CrossRef]
- Adams, K.N.; Verma, A.K.; Gopalaswamy, R.; Adikesavalu, H.; Singhal, D.K.; Tripathy, S.; Ranganathan, U.D.; Sherman, D.R.; Urdahl, K.B.; Ramakrishnan, L.; et al. Diverse Clinical Isolates of Mycobacterium tuberculosis Develop Macrophage-Induced Rifampin Tolerance. J. Infect. Dis. 2019, 219, 1554–1558. [Google Scholar] [CrossRef] [Green Version]
- Helaine, S.; Cheverton, A.M.; Watson, K.G.; Faure, L.M.; Matthews, S.A.; Holden, D.W. Internalization of Salmonella by macrophages induces formation of nonreplicating persisters. Science 2014, 343, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Kåhrström, C.T. Bacterial physiology: Tracking persisters in vivo. Nat. Rev. Microbiol. 2014, 12, 152–153. [Google Scholar] [CrossRef] [PubMed]
- Bellerose, M.M.; Baek, S.H.; Huang, C.C.; Moss, C.E.; Koh, E.I.; Proulx, M.K.; Smith, C.M.; Baker, R.E.; Lee, J.S.; Eum, S.; et al. Common Variants in the Glycerol Kinase Gene Reduce Tuberculosis Drug Efficacy. mBio 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safi, H.; Gopal, P.; Lingaraju, S.; Ma, S.; Levine, C.; Dartois, V.; Yee, M.; Li, L.; Blanc, L.; Ho Liang, H.P.; et al. Phase variation in Mycobacterium tuberculosis glpK produces transiently heritable drug tolerance. Proc. Natl. Acad. Sci. USA 2019, 116, 19665–19674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, A.M.; McKinney, J.D. Role of the methylcitrate cycle in propionate metabolism and detoxification in Mycobacterium smegmatis. Microbiology (Read. Engl.) 2007, 153, 3973–3982. [Google Scholar] [CrossRef] [Green Version]
- Kho, Z.Y.; Lal, S.K. The Human Gut Microbiome—A Potential Controller of Wellness and Disease. Front. Microbiol. 2018, 9, 1835. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.S.; Davies, S.S. Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 2016, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Canani, R.B.; Costanzo, M.D.; Leone, L.; Pedata, M.; Meli, R.; Calignano, A. Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 2011, 17, 1519–1528. [Google Scholar] [CrossRef]
- Ciarlo, E.; Heinonen, T.; Herderschee, J.; Fenwick, C.; Mombelli, M.; Le Roy, D.; Roger, T. Impact of the microbial derived short chain fatty acid propionate on host susceptibility to bacterial and fungal infections in vivo. Sci. Rep. 2016, 6, 37944. [Google Scholar] [CrossRef]
- Ghorbani, P.; Santhakumar, P.; Hu, Q.; Djiadeu, P.; Wolever, T.M.; Palaniyar, N.; Grasemann, H. Short-chain fatty acids affect cystic fibrosis airway inflammation and bacterial growth. Eur. Respir. J. 2015, 46, 1033–1045. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Feng, Y.; Wu, J.; Liu, F.; Zhang, Z.; Hao, Y.; Liang, S.; Li, B.; Li, J.; Lv, N.; et al. The Gut Microbiome Signatures Discriminate Healthy From Pulmonary Tuberculosis Patients. Front. Cell. Infect. Microbiol. 2019, 9, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guinan, J.; Wang, S.; Hazbun, T.R.; Yadav, H.; Thangamani, S. Antibiotic-induced decreases in the levels of microbial-derived short-chain fatty acids correlate with increased gastrointestinal colonization of Candida albicans. Sci. Rep. 2019, 9, 8872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Immerseel, F.; De Buck, J.; Boyen, F.; Bohez, L.; Pasmans, F.; Volf, J.; Sevcik, M.; Rychlik, I.; Haesebrouck, F.; Ducatelle, R. Medium-chain fatty acids decrease colonization and invasion through hilA suppression shortly after infection of chickens with Salmonella enterica serovar Enteritidis. Appl. Environ. Microbiol. 2004, 70, 3582–3587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul Rahim, M.B.H.; Chilloux, J.; Martinez-Gili, L.; Neves, A.L.; Myridakis, A.; Gooderham, N.; Dumas, M.E. Diet-induced metabolic changes of the human gut microbiome: Importance of short-chain fatty acids, methylamines and indoles. Acta Diabetol. 2019, 56, 493–500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelly, A.N.; Sato, Y.; Kearney, S.; Honda, K. Mining the microbiota for microbial and metabolite-based immunotherapies. Nat. Rev. Immunol. 2019, 19, 305–323. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plate Well | Chemical Compound | Planktonic | Biofilm | Abiotic Reaction |
|---|---|---|---|---|
| PM1 plate | ||||
| A1 | Negative control | - | - | - |
| B3 | Glycerol | + | + | - |
| C5 | Tween 20 | + | + | - |
| C8 | Acetic acid | + | + | - |
| D5 | Tween 40 | + | + | - |
| D6 | α-Ketoglutaric acid | + | - | - |
| D7 | α-Ketobutyric acid | + | - | - |
| E5 | Tween 80 | + | + | - |
| E7 | α-Hydroxybutyric acid | + | - | - |
| F7 | Propionic acid | + | + | - |
| G7 | Acetoacetic acid | + | - | - |
| G9 | Monomethyl succinate | + | - | - |
| G10 | Methyl pyruvate | + | + | - |
| H8 | Pyruvic acid | + | + | - |
| PM2A plate | ||||
| A1 | Negative control | - | - | - |
| D12 | Butyric acid | + | + | - |
| E2 | Caproic acid | + | + | - |
| F8 | Sebacic acid | + | + | - |
| Treatment | CFU/mL at 14 Days | ||
|---|---|---|---|
| MAH104 | MAH3388 | MAH3393 | |
| None | 4.2 ± 0.5 × 108 | 9.4 ± 0.2 × 107 | 1.0 ± 0.8 × 108 |
| Clarithromycin | 8.9 ± 0.8 × 107 | - | - |
| Amikacin | 5.8 ± 0.5 × 107 | 5.4 ± 0.6 × 106 | 5.2 ± 0.6 × 107 |
| Glycerol | 1.1 ± 0.3 × 109 | n/a | n/a |
| Glycerol + clarithromycin | 1.7 ± 0.2 × 104 * | - | - |
| Glycerol + amikacin | 7.0 ± 0.2 × 105 * | n/a | n/a |
| Propionic acid | 5.5 ± 0.4 × 106 | 9.6 ± 0.4 × 107 | 1.7 ± 0.4 × 107 |
| Propionic acid + clarithromycin | 2.9 ± 0.3 × 103 * | - | - |
| Propionic acid + amikacin | 3.6 ± 0.6 × 103 * | 3.1 ± 0.5 × 103 * | 1.6 ± 0.3 × 103 * |
| Butyric acid | 3.4 ± 0.6 × 106 | 9.7 ± 0.2 × 107 | 6.2 ± 0.4 × 107 |
| Butyric acid + clarithromycin | 2.7 ± 0.5 × 103 * | - | - |
| Butyric acid + amikacin | 6.3 ± 0.7 × 103 * | 2.4 ± 0.5 × 103 * | 3.1 ± 0.5 × 103 * |
| Caproic acid | 6.4 ± 0.2 × 106 | 8.4 ± 0.7 × 107 | 4.7 ± 0.3 × 107 |
| Caproic acid + clarithromycin | 4.9 ± 0.6 × 103 * | - | - |
| Caproic acid + amikacin | 7.3 ± 0.4 × 103 * | 4.1 ± 0.4 × 103 * | 1.6 ± 0.5 × 103 * |
| Treatment | CFU/mL Cell Lysate | |
|---|---|---|
| 2 h | 4 Days | |
| None | 2.0 ± 0.4 × 105 | 8.2 ± 0.3 × 105 |
| Clarithromycin | 3.9 ± 0.3 × 104 * | |
| Amikacin | 4.8 ± 0.5 × 104 * | |
| Glycerol | 2.3 ± 0.2 × 105 | 9.3 ± 0.3 × 105 |
| Glycerol + clarithromycin | 8.3 ± 0.4 × 103 * | |
| Glycerol + amikacin | 9.1 ± 0.5 × 103 * | |
| Propionic acid | 3.1 ± 0.2 × 105 | 8.0 ± 0.2 × 105 |
| Propionic acid + clarithromycin | 1.1 ± 0.3 × 104 * | |
| Propionic acid + amikacin | 1.7 ± 0.3 × 104 * | |
| Butyric acid | 3.0 ± 0.3 × 105 | 8.4 ± 0.5 × 105 |
| Butyric acid + clarithromycin | 2.0 ± 0.4 × 104 * | |
| Butyric acid + amikacin | 2.0 ± 0.3 × 104 * | |
| Caproic acid | 2.4 ± 0.4 × 105 | 5.3 ± 0.4 × 105 |
| Caproic acid + clarithromycin | 3.6 ± 0.2 × 104 * | |
| Caproic acid + amikacin | 1.6 ± 0.4 × 104 * | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, C.A.d.M.e.; Rojony, R.; Bermudez, L.E.; Danelishvili, L. Short-Chain Fatty Acids Promote Mycobacterium avium subsp. hominissuis Growth in Nutrient-Limited Environments and Influence Susceptibility to Antibiotics. Pathogens 2020, 9, 700. https://doi.org/10.3390/pathogens9090700
Silva CAdMe, Rojony R, Bermudez LE, Danelishvili L. Short-Chain Fatty Acids Promote Mycobacterium avium subsp. hominissuis Growth in Nutrient-Limited Environments and Influence Susceptibility to Antibiotics. Pathogens. 2020; 9(9):700. https://doi.org/10.3390/pathogens9090700
Chicago/Turabian StyleSilva, Carlos Adriano de Matos e, Rajoana Rojony, Luiz E. Bermudez, and Lia Danelishvili. 2020. "Short-Chain Fatty Acids Promote Mycobacterium avium subsp. hominissuis Growth in Nutrient-Limited Environments and Influence Susceptibility to Antibiotics" Pathogens 9, no. 9: 700. https://doi.org/10.3390/pathogens9090700