miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis
by

Snježana Židovec Lepej
1,2, 
Maja Matulić
2,
Paula Gršković
2,
Mirjana Pavlica
2,
Leona Radmanić
1 and
Petra Korać
2,* 1
Department of Immunological and Molecular Diagnostics, University Hospital for Infectious Diseases “Dr. Fran Mihaljević”, 10000 Zagreb, Croatia
2
Division of Molecular Biology, Department of Biology, Faculty of Science, University of Zagreb, 10000 Zagreb, Croatia
*
Author to whom correspondence should be addressed.
Pathogens 2020, 9(5), 353; https://doi.org/10.3390/pathogens9050353
Submission received: 7 April 2020
/
Revised: 25 April 2020
/
Accepted: 4 May 2020
/
Published: 8 May 2020
(This article belongs to the Section Human Pathogens)
Abstract
:Epstein-Barr virus (EBV) or human herpesvirus 4 (HHV-4) is a ubiquitous human oncogenic virus, and the first human virus found to express microRNAs (miRNAs). Its genome contains two regions encoding more than 40 miRNAs that regulate expression of both viral and human genes. There are numerous evidences that EBV miRNAs impact immune response, affect antigen presentation and recognition, change T- and B-cell communication, drive antibody production during infection, and have a role in cell apoptosis. Moreover, the ability of EBV to induce B-cell transformation and take part in mechanisms of oncogenesis in humans is well known. Although EBV infection is associated with development of various diseases, the role of its miRNAs is still not understood. There is abundant data describing EBV miRNAs in nasopharyngeal carcinoma and several studies that have tried to evaluate their role in gastric carcinoma and lymphoma. This review aims to summarize so far known data about the role of EBV miRNAs in altered regulation of gene expression in human cells in EBV-associated diseases.
1. Introduction
Epstein-Barr virus (EBV) or human herpesvirus 4 (HHV-4) is a ubiquitous human oncogenic virus that belongs to the family Herpesviridae, subfamily Gammaherpesvirinae, and genus Lymphocryptovirus [1]. Since its first description in 1964 and subsequent recognition of its ability to induce B-cell transformation in vitro, EBV has been extensively used as a model for research focusing on fundamental mechanisms of oncogenesis in humans. It is also used as a model in devising diagnostic, therapeutic, and prevention strategies in malignant diseases [2,3,4,5]. Approximately 95% of the human population is infected with EBV and will remain carriers of the virus for the rest of their lives. By modulating its own transcriptional patterns during lytic and latent stages of infection, EBV establishes a lifelong persistence in both immunocompetent and immunocompromised hosts and modifies the host’s immune system effector mechanisms [6]. However, in the context of immunosuppression, regardless of the specific cause, the immune system fails to efficiently control EBV replication [7]. Subsequently, latent infection with EBV may be associated with development of various malignancies originating from epithelial cells, lymphocytes, and mesenchymal cells, including posttransplant B-cell lymphomas, Hodgkin’s and non-Hodgkin’s lymphomas, diffuse large B-cell lymphoma, Burkitt’s lymphoma, natural killer (NK)/T-cell lymphoma, nasopharyngeal carcinoma (NPC), and gastric carcinoma (GC) [7,8].
MicroRNAs (miRNA) are small, non-coding RNA molecules of cellular or viral origin that consist of 18–22 nucleotides and play an important role in regulation of gene expression. Consequently, they affect key events in cell biology such as proliferation, apoptosis, and lipid metabolism. By binding to messenger RNAs (mRNAs), miRNAs induce degradation of mRNAs or inhibition of translation, thus reducing levels of expression of target genes. Virus-encoded miRNAs (v-miRNAs) are considered an important non-immunogenic tool for post-transcriptional regulation of both host and viral gene expression in infected cells [9]. The majority of literature data on v-miRNAs, particularly in the context of tumorigenesis, focuses on v-miRNAs in EBV, Kaposi’s sarcoma-associated herpesvirus (KSHV/HHV8), human papillomaviruses (HPV), hepatitis C virus (HCV), hepatitis B virus (HBV), and Merkel Cell Polyomavirus (MCPyV) [9,10,11]. EBV was the first virus shown to express miRNAs [12]. EBV-encoded miRNAs (EBV miRNAs) target both viral and cellular mRNAs in infected cells, extending their role beyond regulating various stages of the EBV replication cycle. They influence cellular proliferation and apoptosis and play a part in driving diverse molecular pathways of oncogenesis and evading innate and adaptive immune responses. The aim of this review is to summarize current views on the role of EBV miRNAs in altered gene expression associated with immune evasion and tumorigenesis.
2. EBV miRNAs
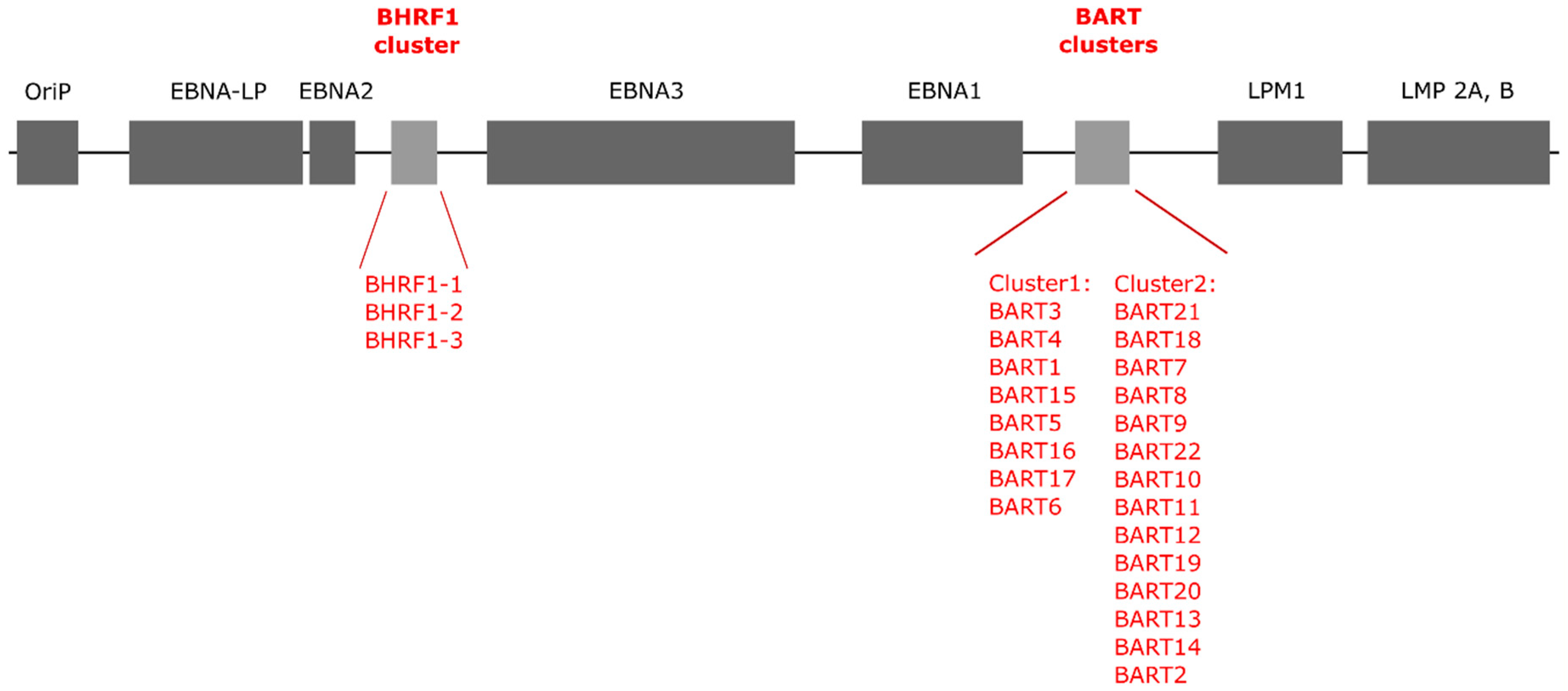
EBV genome is a double-stranded DNA molecule that consists of 175 kbp containing nearly 100 genes and coding for 44 microRNAs [13,14,15]. It was first sequenced in 1984 by using M13 libraries made from viral EcoR I and BamH I fragments gathered after sonification [16]. Today, after more than 100 EBV genomes from tumor cells samples and healthy individuals’ tissue have been sequenced, the complete EBV genome can be found in the NCBI GeneBank [17]. In 2004, Pfeffer et al. first described two clusters in the EBV genome responsible for production of EBV miRNAs [12]. The first cluster was found in the sequence for BamH I fragment H rightward open reading frame 1 (BHRF1) mRNA. It produces three miRNA precursors (ebv-miR-BHRF1-1, -2, and -3) and, subsequently, four mature miRNAs [18]. The second cluster is BamH I-A region rightward transcript (BART) and it encodes 22 miRNA precursors (ebv-miR-BART1-22) of 40 mature miRNA molecules [12,19] (Figure 1).
Expression of BHRF1 miRNAs is latency stage-dependent (they are mainly expressed in type III latency), while BART miRNAs are transcribed in all latency stages [12]. Despite the coordinated expression of each miRNA cluster, significant differences in the expression levels of individual EBV miRNAs in the same type of human cells (as high as 50-fold) have been observed [12,22]. It has been suggested that different genotypes as well as genomic variants of EBV could be associated with different patterns of individual miRNA expression in infected cells [23,24,25]. A number of studies on EBV miRNA biosynthesis (reviewed by Wang et al., 2018) suggest that EBV gene products are not necessary for v-miRNA production. This proposition indicates that the molecular mechanisms responsible for the synthesis of v-miRNAs and cellular miRNAs in host cells are similar [20]. Analysis of EBV genomic sequences in gastric carcinoma and EBV-associated lymphoma has showed that despite the genetic diversity in almost the entire EBV genome, the regions encoding miRNAs are highly conserved [26,27].
3. The Role of EBV miRNAs in Immune Evasion
Alongside immune evasion strategies mediated by host miRNAs and viral glycoproteins, EBV miRNAs are another means by which the virus successfully avoids effector mechanisms of the host’s immune system [28]. The main strategies for immune evasion used by EBV miRNAs are the following:
3.1. Pattern-Recognition Receptor-Mediated Signaling Pathways and Interferons
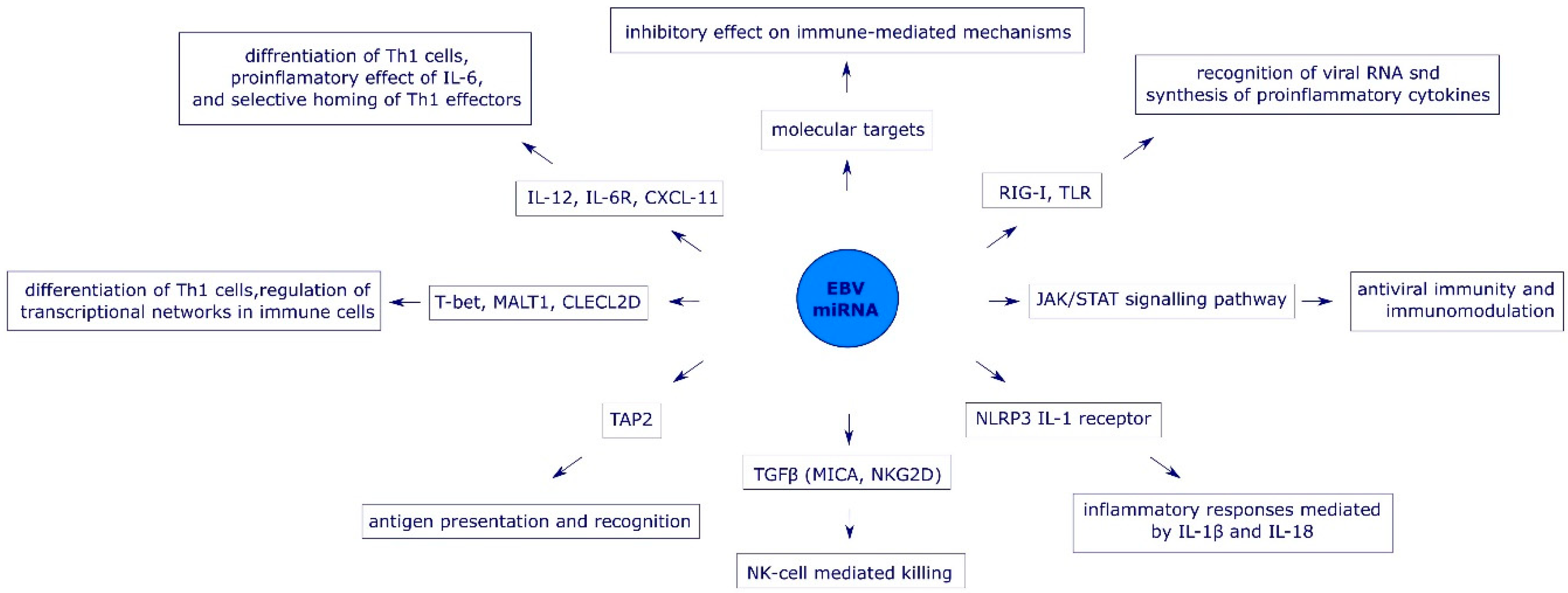
EBV interferes with efficient initiation of innate immune response at the very first step, e.g., by targeting expression of pattern recognition receptors (PRR). The impaired expression of PRR affects subsequent signal transduction as well as cytokine synthesis [29]. The two main targets for EBV miRNAs are retinoic acid-inducible protein 1 (RIG-I) receptors (mediated by miR-BART6-3p) and Toll-like receptors [20,30]. The lack of EBV molecular pattern recognition by PRR is associated with the absence of the JAK (Janus kinase)-STAT (Signal Transducer and Activator of Transcription)-mediated transduction pathway. The ineffectiveness of this signaling pathway leads to the impaired synthesis of type I interferons (IFNs) and other pro-inflammatory cytokines that are essential for innate immune responses [31]. Studies on nasal NK-cell lymphoma (NNL) cells showed that EBV miRNAs (miR-BART20-5p and miR-BART8) impact the signal transduction pathway for IFN-γ by targeting STAT1, which enables the virus to avoid antiviral activity of both type I and type II IFNs [32] (Figure 2).
3.2. Natural Killer (NK) Cells
The role of EBV miRNAs in the evasion of NK-cell-mediated responses has been analyzed in nasopharyngeal carcinoma cells in vitro. Interaction between the natural killer group 2 member D (NKG2D) receptor on NK cells and major histocompatibility complex class I chain-related peptide A (MICA) is considered the key step in the recognition and killing of cancer cells. Expression of MICA in NPC is positively regulated by transforming growth factor β-1 (TGF-β1). Wong et al. (2018) showed that miR-BART7 reduces the expression of TGF-β1 in NPC cells and impairs the NK-cell-mediated recognition of virus-infected cells [33] (Figure 2).
3.3. Inflammasome
Immune evasion strategies employed by EBV also target NLR family pyrin domain-containing 3 (NLRP3) inflammasomes, which are responsible for inflammatory responses mediated by IL-1β and IL-18 upon recognition of viral antigens. The activity of inflammasomes is targeted directly at the level of NLRP3 expression (via miR-BART15), or indirectly, by targeting the IL-1 receptor itself (via miR-BHRF1-2-5p) [34,35] (Figure 2).
3.4. Cytokines and Chemokines
miR-BART1, miR-BART2, miR-BART10, and miR-BART22 suppress efficient CD8+ T-cell-mediated antiviral immune response by targeting IL-12, a pro-inflammatory cytokine responsible for differentiation of naive type 1 helper T-cell (Th1 cells) to mature Th1 cells, increased synthesis of IFN-γ, activation of NK- and T-cells as well as inhibition of angiogenesis [36,37]. In addition, miR-BART6-3p interferes with biological activity of IL-6 by targeting expression of the IL-6 receptor [38]. EBV miRNAs also enable the virus to evade Th1-mediated antiviral immunity by modulating expression of chemokines. For example, miR-BHRF1-3 targets an IFN-inducible chemokine CXCL11 (CXC-chemokine ligand 11) responsible for the selective homing of Th1 effector cells and NK-cells to the sites of infection [20,39] (Figure 2).
3.5. Antigen Presentation
EBV miRNAs impair mechanisms of specific immunity by affecting adequate antigen recognition at the level of antigen processing (reduced expression of lysosomal enzymes), transport of processed antigenic peptides to major histocompatibility complex (MHC) molecules (by targeting peptide transporter subunit TAP2), and antigen presentation (reduction of lymphocyte antigen 75 expression on dendritic cells) [20,37,40,41,42] (Figure 2).
3.6. Specific Cellular Immunity
T-cell-mediated immunity can maintain long term immune control over EBV replication (for >50 years in some individuals) while clinical consequences associated with EBV infection in persons with impairment of T-cell development or function are shown to be very severe. This suggests that virus-specific T-cell responses represent the main means of protective immunity in EBV infection [37]. EBV miRNAs specifically target host genes coding for key regulators of T-cell responses including T-bet (miR-BART20-5p), Mucosa-associated lymphoid tissue lymphoma transport protein 1 (MALT1) (miR-BHRF1-2-5p), and C type lectin superfamily 2 member D (CLEC2D) (miR-BART1-3p and miR-BART3-3p) [43] (Figure 2).
T-bet belongs to the T-box family of transcription factors that are the main enhancers of the Th1 differentiation pathway and subsequent Th1 cell-specific IFN-γ synthesis, which are important for efficient antiviral immunity. Inhibition of T-bet translation (with subsequent suppression of p53) by EBV miRNAs, originally shown in invasive nasal NK/T-cell lymphoma cells, is supposedly associated with the inhibition of Th1 differentiation pathways. As a result, the control of EBV replication is less efficient [43].
However, T-bet also regulates transcriptional networks that are common among other types of immune cells (including dendritic cells and innate lymphoid cells) and is currently considered to have an important role in bridging innate and adaptive immunity (for review see Lazarevic et al., 2017) [44].
In addition, T-bet acts as a selective repressor of transcriptional pathways associated with type I IFNs subsequent to IFN-γ-induced signaling [44,45]. Therefore, EBV miRNAs-mediated inhibition of T-bet’s biological activity may have a significantly broader effect on the evasion of antiviral immunity in EBV infection than originally thought.
4. The Role of EBV miRNAs in Tumorigenesis
EBV association with development and progression of malignant tumors is well known, especially its frequent infection of lymphoma and carcinoma cells. Nevertheless, the role of EBV miRNAs in tumorigenesis has only recently come into focus.
4.1. EBV miRNAs in Lymphoma
EBV miRNAs have been linked to development and progression of various types of lymphoma, such as acquired immunodeficiency syndrome-related diffuse large B-cell lymphoma (AIDS-related DLBCL), EBV-positive DLBCL, Burkitt lymphoma (BL), nasal NK/T-cell lymphoma (NNL), Hodgkin’s lymphoma (HL), posttransplant lymphoproliferative disorder (PTLD), and others [23].
miR-BHRF1-3 was found to be highly expressed in AIDS-related DLBCL compared to EBV-positive BL and EBV-negative DLBCL [39]. miR-BHRF-1-1 was detected in all samples of EBV-associated primary central nervous system PTLD (pCNS PTLD) and mir-BHRF-1-2 was found in about 50% of the same pCNS PTLD patients [46].
EBV-miRNAs belonging to BART clusters were found in pCNS PTLD, but with varying expression of different BART miRNAs [46]. Furthermore, miR-BART7, miR-BART22, miR-BART10, miR-BART11-5p, and miR-BART16 were found to be most prominently expressed in EBV-positive DLBCL not associated with AIDS [47]. Only EBV miRNAs from BART clusters were found in endemic BL (eBL) [48]. miR-BART7, miR-BART5, miR-BART11-5p, miR-BART1-5p, and miR-BART19-3p were found in NNL, as well as miR-BART21 and miR-BART22, previously found in EBV-positive carcinomas [49,50]. Expression of BART miRNAs have also been observed in HL [12].
The presence of diverse EBV miRNAs in different types of lymphoma was analyzed not only in patient samples, but it was also researched through cell line-based studies. In BL41/95 cell line, derived from BL, BHRF miRNAs were detected [12]. The study by Ambrosio et al. in which BL-derived cell line Akata was used as a model, revealed that the expression of PTEN and IL6 receptor subunits was lower in the presence of miR-BART6-3p and was restored if the cells were simultaneously transfected with miR-BART6-3p inhibitor [48]. Zhou et al. found that Ramos cell line (derived from EBV-negative BL) transfected simultaneously with oligonucleotides mimicking cellular miRNA-142 and miBART6-3p, displayed lower expression of PTEN compared to negative control and cells transfected with the same oligonucleotides separately. This suggests that miR-BART6-3p downregulates PTEN, a tumor suppressor that regulates the PI3K/Akt pathway, in cooperation with cellular miRNA-142 [51]. Moreover, in two cell lines derived from lymphomas of NK cell origin (YT and NK92), it was shown that miR-BART20-5p was responsible for downregulation of T-bet, and therefore p53, and IFN-γ [43].
4.2. EBV miRNAs in Carcinoma
EBV is also known to be associated with the development of NPC and GC. Recently, the role of EBV miRNAs was thoroughly studied in these malignancies. BART miRNAs were first found in NPC, xenografted and propagated in nude mice [52,53], and subsequently in NPC patient biopsies [54]. They were found to be highly expressed in NPC and GC, but BHRF1 miRNAs were generally not present in those entities. At least 105 host genes were shown to be regulated by BART miRNAs during carcinoma development [55], but genes recognized as EBV miRNAs’ targets in B-cell lymphomas were not confirmed in carcinomas. Increased expression of BART miRNA clusters and individual BART miRNAs correlated with higher tumor grade and poor patient survival in NPC and GC.
Generally, it is believed that BART miRNAs in carcinoma act in synergy or obtain significant effects by combining their individual activities, in cooperation or competition with cellular miRNAs. It was shown that BART miRNAs target numerous transcripts of different genes, thus deregulating various downstream molecules and signaling pathways [56]. Consequently, BART miRNAs allow infected cells to avoid apoptosis by inactivating different pro-apoptotic molecules, influence cell proliferation, inhibit the expression of regulatory tumor suppressors, mediate cancer metabolism, stimulate cell migration and metastasis, inhibit cell differentiation, and manage immune evasion and regulation of the virus latency through coordination of cellular and viral signaling pathways [49,57,58] (Table 1).
BART miRNAs also modulate host cell pathways by mimicking cellular miRNAs. Although this feature is still not explored enough, several BART miRNAs were shown to have “seed” sequences similar to those of cellular miRNAs: miR-BART5 compared to miR-18a and miR-18b [66,79], miR-BART9 to miR-200a and miR-141 [72], miR-BART15-3p to miR-223-3p, and miR-BART18-5p to miR-26a [35,80].
Overall, BART miRNAs are highly expressed in NPC and GC types associated with EBV infection where they act synergistically and have redundant activities, but also possibly differ in their target genes in different intracellular milieus.
5. Conclusions
We envision that further detection of cellular processes affected and regulated by EBV miRNAs will contribute to better understanding of the role of viral non-coding RNAs in the development of virus-induced cancers in humans. In case of EBV, despite extensive research, there are currently no antiviral drugs or EBV-vaccines approved for use in humans. In years to come, better evaluation and understanding of the viral miRNA mechanisms might reveal new biomarkers and potential therapeutic targets.
Author Contributions
Conceptualization, S.Ž.L. and P.K.; writing—original draft preparation, S.Ž.L., P.K., M.M., P.G., M.P., and L.R.; writing—review and editing, S.Ž.L., P.K., M.M., P.G., M.P., and L.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- International Committee on Taxonomy of Viruses ICTV (2019): ICTV Taxonomy History: Human Gammaherpesvirus 4. Available online: https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=20171486 (accessed on 1 October 2019).
- Epstein, M.A.; Achong, B.G.; Barr, Y.M. Virus particles in cultured lymphoblasts from Burkitt’s lymphoma. Lancet 1964, 1, 702–703. [Google Scholar] [CrossRef]
- Pope, J.H. Establishment of Cell Lines from Peripheral Leucocytes in Infectious Mononucleosis. Nature 1967, 216, 810–811. [Google Scholar] [CrossRef]
- Henle, W.; Diehl, V.; Kohn, G.; Hausen, H.Z.; Henle, G. Herpes-type virus and chromosome marker in normal leukocytes after growth with irradiated Burkitt cells. Science 1967, 157, 1064–1065. [Google Scholar] [CrossRef]
- Young, L.S.; Yap, F.L.; Murray, P.G. Epstein-Barr virus: More than 50 years old and still providing surprises. Nat. Rev. Cancer 2016, 16, 789–802. [Google Scholar] [CrossRef]
- Thorley-Lawson, D.A. EBV Persistence-Introducing the Virus. Curr. Top. Microbiol. Immunol. 2015, 390 Pt 1, 151–209. [Google Scholar]
- Saha, A.; Robertson, E.S. Mechanisms of B-Cell Oncogenesis Induced by Epstein-Barr Virus. J. Virol. 2019, 93, e00238–e00319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsueh, C.Y.; Yang, C.F.; Gau, J.P.; Kuan, E.C.; Ho, C.Y.; Chiou, T.J.; Hsiao, L.T.; Lin, T.A.; Lan, M.Y. Nasopharyngeal lymphoma: A 22-year review of 35 cases. J. Clin. Med. 2019, 8, 1604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, R.; Kumar, A.; Ingle, H.; Kumar, H. The Interplay Between Viral-Derived miRNAs and Host Immunity During Infection. Front. Immunol. 2020, 10, 3079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shigeyoshi, F.; Nakamura, H. Animal Models for Gammaherpesvirus Infections: Recent Development in the Analysis of Virus-Induced Pathogenesis. Pathogens 2020, 9, 116. [Google Scholar]
- Gallo, A.; Miceli, V.; Bulati, M.; Iannolo, G.; Contino, F.; Conaldi, P.G. Viral miRNAs as Active Players and Participants in Tumorigenesis. Rev. Cancers 2020, 12, 358. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.; Zavolan, M.; Grässer, F.A.; Chien, M.; Russo, J.J.; Ju, J.; John, B.; Enright, A.J.; Marks, D.; Sander, C.; et al. Identification of virus-encoded microRNAs. Science 2004, 304, 734–736. [Google Scholar] [CrossRef] [PubMed]
- Mui, U.N.; Haley, C.T.; Tyring, S.K. Viral Oncology: Molecular Biology and Pathogenesis. J. Clin. Med. 2017, 6, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, S.; Endo, K.; Wakisaka, N.; Aga, M.; Kano, M.; Seishima, N.; Imoto, T.; Kobayashi, E.; Moriyama-Kita, M.; Nakanishi, Y.; et al. Expression of interferon regulatory factor 7 correlates with the expression of Epstein-Barr Virus latent membrane protein 1 and cervical lymph node metastasis in nasopharyngeal cancer. Pathol. Int. 2017, 67, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Chen, J.N.; Qiu, X.M.; Pan, Y.H.; Zhang, Z.G.; Shao, C.K. Comparative analysis of 22 Epstein-Barr virus genomes from diseased and healthy individuals. J. Gen. Virol. 2017, 98, 96–107. [Google Scholar] [CrossRef] [PubMed]
- Baer, R.; Bankier, A.T.; Biggin, M.D.; Deininger, P.L.; Farrell, P.J.; Gibson, T.J.; Hatfull, G.; Hudson, G.S.; Satchwell, S.C.; Séguin, C.; et al. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature 1984, 310, 207–211. [Google Scholar] [CrossRef]
- Human Gammaherpesvirus 4, Complete Genome. Available online: https://www.ncbi.nlm.nih.gov/nuccore/NC_007605 (accessed on 1 October 2019).
- Barth, S.; Meister, G.; Grasser, F.A. EBV-encoded miRNAs. Biochim. Biophys. Acta 2011, 1809, 631–640. [Google Scholar] [CrossRef]
- Cai, X.; Schäfer, A.; Lu, S.; Bilello, J.P.; Desrosiers, R.C.; Edwards, R.; Raab-Traub, N.; Cullen, B.R. Epstein-Barr virus microRNAs are evolutionarily conserved and differentially expressed. PLoS Pathog. 2006, 2, e23. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Yu, F.; Wu, W.; Wang, Y.; Ding, H.; Qian, L. Epstein-Barr virus-encoded microRNAs as regulators in host immune responses. Int. J. Biol. Sci. 2018, 14, 565–576. [Google Scholar] [CrossRef] [Green Version]
- Zanella, L.; Riquelme, I.; Buchegger, K.; Abanto, M.; Ili, C.; Brebi, P. A reliable Epstein-Barr Virus classification based on phylogenomic and population analyses. Sci. Rep. 2019, 9, 9829. [Google Scholar] [CrossRef] [Green Version]
- Pratt, Z.L.; Kuzembayeva, M.; Sengupta, S.; Sugden, B. The microRNAs of Epstein-Barr Virus are expressed at dramatically differing levels among cell lines. Virology 2009, 386, 387–397. [Google Scholar] [CrossRef] [Green Version]
- Navari, M.; Etebari, M.; Ibrahimi, M.; Leoncini, L.; Piccaluga, P.P. Pathobiologic Roles of Epstein–Barr Virus-Encoded MicroRNAs in Human Lymphomas. Int. J. Mol. Sci. 2018, 19, 1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.; Lin, X.; Shumilov, A.; Bernhardt, K.; Feederle, R.; Poirey, R.; Kopp-Schneider, A.; Pereira, B.; Almeida, R.; Delecluse, H.J. The biological properties of different Epstein-Barr virus strains explain their association with various types of cancers. Oncotarget 2017, 8, 10238–10254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correia, S.; Palser, A.; Elgueta Karstegl, C.; Middeldorp, J.M.; Ramayanti, O.; Cohen, J.I.; Hildesheim, A.; Fellner, M.D.; Wiels, J.; White, R.E.; et al. Natural Variation of Epstein-Barr Virus Genes, Proteins, and Primary MicroRNA. J. Virol. 2017, 91, e00375–e00417. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.N.; Zhou, L.; Qiu, X.M.; Yang, R.H.; Liang, J.; Pan, Y.H.; Li, H.F.; Peng, G.R.; Shao, C.K. Determination and genome-wide analysis of Epstein-Barr virus (EBV) sequences in EBV-associated gastric carcinoma from Guangdong, an endemic area of nasopharyngeal carcinoma. J. Med. Microbiol. 2018, 67, 1614–1627. [Google Scholar] [CrossRef]
- Sunagawa, K.; Hishima, T.; Fukumoto, H.; Hasegawa, H.; Katano, H. Conserved sequences of bart and bhrf regions encoding viral microRNAs in epstein–barr virus-associated lymphoma. BMC Res. Notes 2017, 10, 279. [Google Scholar] [CrossRef] [Green Version]
- Zuo, L.; Yue, W.; Du, S.; Xin, S.; Zhang, J.; Liu, L.; Li, G.; Lu, J. An update: Epstein-Barr virus and immune evasion via microRNA regulation. Virol. Sin. 2017, 32, 175–187. [Google Scholar] [CrossRef]
- Jangra, S.; Yuen, K.S.; Botelho, M.G.; Jin, D.Y. Epstein–Barr Virus and Innate Immunity: Friends or Foes? Microorganisms 2019, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Qin, Z.; Wang, J.; Zheng, X.; Lu, J.; Zhang, X.; Wei, L.; Peng, Q.; Zheng, Y.; Ou, C.; et al. Epstein-Barr virus miR-BART6-3p inhibits the RIG-I pathway. J. Innate Immun. 2017, 9, 574–586. [Google Scholar] [CrossRef]
- Owen, K.L.; Brockwell, N.K.; Parker, B.S. JAK-STAT Signaling: A Double-Edged Sword of Immune Regulation and Cancer Progression. Cancers 2019, 11, 2002. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.T.; Lin, C.W. EBV-encoded miR-BART20-5p and miR-BART8 inhibit the IFN-gamma-STAT1 pathway associated with disease progression in nasal NK-cell lymphoma. Am. J. Pathol. 2014, 184, 1185–1197. [Google Scholar] [CrossRef]
- Wong, T.S.; Chen, S.; Zhang, M.J.; Chan, J.Y.; Gao, W. Epstein-Barr virus-encoded microRNA BART7 downregulates major histocompatibility complex class I chain-related peptide A and reduces the cytotoxicity of natural killer cells to nasopharyngeal carcinoma. Oncol. Lett. 2018, 16, 2887–2892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, C.M.; Ivanov, N.S.; Barr, S.A.; Chen, Y.; Skalsky, R.L. An Epstein-Barr Virus MicroRNA Blocks Interleukin-1 (IL-1) Signaling by Targeting IL-1 Receptor 1. J. Virol. 2017, 91, e00530–e00617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O’Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1 production. J. Immunol. 2012, 189, 3795–3799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Cao, W.; Zhu, Y. Immunoregulatory Functions of the IL-12 Family of Cytokines in Antiviral Systems. Viruses 2019, 11, 772. [Google Scholar] [CrossRef] [Green Version]
- Albanese, M.; Tagawa, T.; Bouvet, M.; Maliqi, L.; Lutter, D.; Hoser, J.; Hastreiter, M.; Hayes, M.; Sugden, B.; Martin, L.; et al. Epstein-Barr virus microRNAs reduce immune surveillance by virus-specific CD8+ T cells. Proc. Natl. Acad. Sci. USA 2016, 113, e6467–e6475. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.M.; Yu, Y.; Zhao, H.P. EBVBART63p and cellular microRNA197 compromise the immune defense of host cells in EBV-positive Burkitt lymphoma. Mol. Med. Rep. 2017, 15, 1877–1883. [Google Scholar] [CrossRef]
- Xia, T.; O’Hara, A.; Araujo, I.; Barreto, J.; Carvalho, E.; Sapucaia, J.B.; Ramos, J.C.; Luz, E.; Pedroso, C.; Manrique, M.; et al. EBV microRNAs in primary lymphomas and targeting of CXCL-11 by ebv-mir-BHRF1-3. Cancer Res. 2008, 68, 1436–1442. [Google Scholar] [CrossRef] [Green Version]
- Tagawa, T.; Albanese, M.; Bouvet, M.; Moosmann, A.; Mautner, J.; Heissmeyer, V.; Zielinkski, C. Epstein-Barr viral miRNAs inhibit antiviral CD4+ T cell responses targeting IL-12 and peptide processing. J. Exp. Med. 2016, 213, 2065–2080. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Corcoran, D.L.; Gottwein, E.; Frank, C.L.; Kang, D.; Hafner, M.; Nusbaum, J.D. The viral and cellular microRNA targetome in lymphoblastoid cell lines. PLoS Pathog. 2012, 8, e1002484. [Google Scholar] [CrossRef]
- Gurer, C.; Strowig, T.; Brilot, F.; Pack, M.; Trumpfheller, C.; Arrey, F.; Park, C.G. Targeting the nuclear antigen 1 of Epstein-Barr virus to the human endocytic receptor DEC-205 stimulates protective T-cell responses. Blood 2008, 112, 1231–1239. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.C.; Liu, T.Y.; Hsu, S.M.; Lin, C.W. Epstein-Barr virus-encoded miR-BART20-5p inhibits T-bet translation with secondary suppression of p53 in invasive nasal NK/T-cell lymphoma. Am. J. Pathol. 2013, 182, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Lazarevic, V.; Szabo, S.; Glimcher, L.H. T-bet runs INTERFERence. Immunity 2017, 46, 968–970. [Google Scholar] [CrossRef] [PubMed]
- Lazarević, V.; Glimcher, L.H.; Lord, G.M. T-bet: A bridge between innate and adaptive immunity. Nat. Rev. Immunol. 2013, 13, 777–789. [Google Scholar] [CrossRef] [PubMed]
- Fink, S.E.K.; Gandhi, M.K.; Nourse, J.P.; Keane, C.; Jones, K.; Crooks, P.; Jöhrens, K.; Korfel, A.; Schmidt, H.; Neumann, S.; et al. A Comprehensive Analysis of the Cellular and EBV-Specific MicroRNAome in Primary CNS PTLD Identifies Different Patterns Among EBV-Associated Tumors. Am. J. Transplant. 2014, 14, 2577–2587. [Google Scholar] [CrossRef] [PubMed]
- Imig, J.; Motsch, N.; Zhu, J.Y.; Barth, S.; Okoniewski, M.; Reineke, T.; Tinguely, M.; Faggioni, A.; Trivedi, P.; Meister, G.; et al. microRNA profiling in Epstein–Barr virus-associated B-cell lymphoma. Nucleic Acids Res. 2011, 39, 1880–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosio, M.R.; Navari, M.; Di Lisio, L.; Leon, E.A.; Onnis, A.; Gazaneo, S.; Mundo, L.; Ulivieri, C.; Gomez, G.; Lazzi, S.; et al. The epstein–barr-encoded BART-6-3p microrna affects regulation of cell growth and immuno response in burkitt lymphoma. Infect. Agent Cancer 2014, 9, 12. [Google Scholar] [CrossRef] [Green Version]
- Motsch, N.; Alles, J.; Imig, J.; Zhu, J.; Barth, S.; Reineke, T.; Tinguely, M.; Cogliatti, S.; Dueck, A.; Meister, G.; et al. MicroRNA Profiling of Epstein-Barr Virus-Associated NK/T-Cell Lymphomas by Deep Sequencing. PLoS ONE 2012, 7, e42193. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Pfuhl, T.; Motsch, N.; Barth, S.; Nicholls, J.; Grässer, F.; Meister, G. Identification of Novel Epstein-Barr Virus MicroRNA Genes from Nasopharyngeal Carcinomas. J. Virol. 2009, 83, 3333–3341. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.; Bu, Y.; Liang, Y.; Zhang, F.; Zhang, H.; Li, S. Epstein-Barr Virus (EBV)-BamHI-A Rightward Transcript (BART)-6 and Cellular MicroRNA-142 Synergistically Compromise Immune Defense of Host Cells in EBV-Positive Burkitt Lymphoma. Med. Sci. Monit. 2016, 22, 4114–4120. [Google Scholar] [CrossRef] [Green Version]
- Gilligan, K.; Sato, H.; Rajadurai, P.; Busson, P.; Young, L.; Rickinson, A.; Tursz, T.; Raab-Traub, N. Novel transcription from the Epstein-Barr virus terminal EcoRI fragment, DIJhet, in a nasopharyngeal carcinoma. J. Virol. 1990, 64, 4948–4956. [Google Scholar] [CrossRef] [Green Version]
- Hitt, M.M.; Allday, M.J.; Hara, T.; Karran, L.; Jones, M.D.; Busson, P.; Tursz, T.; Ernberg, I.; Griffin, B.E. EBV gene expression in an NPC-related tumour. EMBO J. 1989, 8, 2639–2651. [Google Scholar] [CrossRef] [PubMed]
- Gilligan, K.J.; Rajadurai, P.; Lin, J.C.; Busson, P.; Abdel-Hamid, M.; Prasad, U.; Tursz, T.; Raab-Traub, N. Expression of the Epstein-Barr virus BamHI A fragment in nasopharyngeal carcinoma: Evidence for a viral protein expressed in vivo. J. Virol. 1991, 65, 6252–6259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, Z.; Shu, Y.; Zhou, H.; Wang, H.; Zhang, W. BART miRNAs: An unimaginable force in the development of nasopharyngeal carcinoma. Eur. J. Cancer Prev. 2017, 26, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wan, X.X.; Yi, H.; Qu, J.Q.; He, Q.Y.; Xiao, Z.Q. Integrated analysis of the differential cellular and EBV miRNA expression profiles in microdissected nasopharyngeal carcinoma and non-cancerous nasopharyngeal tissues. Oncol. Rep. 2015, 34, 2585–2601. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Z.; Huang, H.; Huang, L.; Sun, M.; Yan, Q.; Song, Y.; Wei, F.; Bo, H.; Gong, Z.; Zeng, Y.; et al. Regulation network and expression profiles of Epstein-Barr virus-encoded microRNAs and their potential target host genes in nasopharyngeal carcinomas. Sci. China Life Sci. 2014, 57, 315–326. [Google Scholar] [CrossRef] [Green Version]
- Wong, A.M.; Kong, K.L.; Tsang, J.W.; Kwong, D.L.; Guan, X.Y. Profiling of Epstein-Barr virus-encoded microRNAs in nasopharyngeal carcinoma reveals potential biomarkers and oncomirs. Cancer 2012, 118, 698–710. [Google Scholar] [CrossRef]
- Cai, L.; Ye, Y.; Jiang, Q.; Chen, Y.; Lyu, X.; Li, J.; Wang, S.; Liu, T.; Cai, H.; Yao, K.; et al. Epstein-Barr virus-encoded microRNA BART1 induces tumour metastasis by regulating PTEN-dependent pathways in nasopharyngeal carcinoma. Nat. Commun. 2015, 6, 7353. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.M.; Lyu, X.M.; Luo, W.R.; Cui, X.F.; Ye, Y.F.; Yuan, C.C.; Peng, Q.X.; Wu, D.H.; Liu, T.F.; Wang, E.; et al. EBV-miR-BART7-3p promotes the EMT and metastasis of nasopharyngeal carcinoma cells by suppressing the tumor suppressor PTEN. Oncogene 2015, 34, 2156–2166. [Google Scholar] [CrossRef] [Green Version]
- Lyu, X.; Wang, J.; Guo, X.; Wu, G.; Jiao, Y.; Faleti, O.D.; Liu, P.; Liu, T.; Long, Y.; Chong, T.; et al. EBV-miR-BART1-5P activates AMPK/mTOR/HIF1 pathway via a PTEN independent manner to promote glycolysis and angiogenesis in nasopharyngeal carcinoma. PLoS Pathog. 2018, 14, e1007484. [Google Scholar] [CrossRef] [Green Version]
- Lo, A.K.; To, K.F.; Lo, K.W.; Lung, R.W.; Hui, J.W.; Liao, G.; Hayward, S.D. Modulation of LMP1 protein expression by EBV-encoded microRNAs. Proc. Natl. Acad. Sci. USA 2007, 104, 16164–16169. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.; Skalsky, R.L.; Cullen, B.R. EBV BART MicroRNAs Target Multiple Pro-apoptotic Cellular Genes to Promote Epithelial Cell Survival. PLoS Pathog. 2015, 11, e1004979. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Yuen, K.S.; Xu, R.; Tsao, S.W.; Chen, H.; Li, M.; Kok, K.H.; Jin, D.Y. Targeting of DICE1 tumor suppressor by Epstein-Barr virus-encoded miR-BART3* microRNA in nasopharyngeal carcinoma. Int. J. Cancer 2013, 133, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Shinozaki-Ushiku, A.; Kunita, A.; Isogai, M.; Hibiya, T.; Ushiku, T.; Takada, K.; Fukayama, M. Profiling of Virus-Encoded MicroRNAs in Epstein-Barr Virus-Associated Gastric Carcinoma and Their Roles in Gastric Carcinogenesis. J. Virol. 2015, 89, 5581–5591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choy, E.Y.; Siu, K.L.; Kok, K.H.; Lung, R.W.; Tsang, C.M.; To, K.F.; Kwong, D.L.; Tsao, S.W.; Jin, D.Y. An Epstein-Barr virus-encoded microRNA targets PUMA to promote host cell survival. J. Exp. Med. 2008, 205, 2551–2560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Wang, J.; Wei, L.; Peng, Q.; Gao, Y.; Fu, Y.; Lu, Y.; Qin, Z.; Zhang, X.; Lu, J.; et al. Epstein-Barr Virus MicroRNA miR-BART5-3p Inhibits p53 Expression. J. Virol. 2018, 92, e01022-18. [Google Scholar] [CrossRef] [Green Version]
- Lung, R.W.; Hau, P.M.; Yu, K.H.; Yip, K.Y.; Tong, J.H.; Chak, W.P.; Chan, A.W.; Lam, K.H.; Lo, A.K.; Tin, E.K.; et al. EBV-encoded miRNAs target ATM-mediated response in nasopharyngeal carcinoma. J. Pathol. 2018, 244, 394–407. [Google Scholar] [CrossRef]
- He, B.; Li, W.; Wu, Y.; Wei, F.; Gong, Z.; Bo, H.; Wang, Y.; Li, X.; Xiang, B.; Guo, C.; et al. Epstein-Barr virus-encoded miR-BART6-3p inhibits cancer cell metastasis and invasion by targeting long non-coding RNA LOC553103. Cell Death Dis. 2016, 7, e2353. [Google Scholar] [CrossRef]
- Iizasa, H.; Wulff, B.E.; Alla, N.R.; Maragkakis, M.; Megraw, M.; Hatzigeorgiou, A.; Iwakiri, D.; Takada, K.; Wiedmer, A.; Showe, L.; et al. Editing of Epstein-Barr virus-encoded BART6 microRNAs controls their dicer targeting and consequently affects viral latency. J. Biol. Chem. 2010, 285, 33358–33370. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Zong, J.; Lin, W.; Wang, M.; Xu, Y.; Zhou, R.; Lin, S.; Guo, Q.; Chen, H.; Ye, Y.; et al. EBV-miR-BART8-3p induces epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma cells through activating NF-κB and Erk1/2 pathways. J. Exp. Clin. Cancer Res. 2018, 37, 283. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Yi, Y.H.; Chang, K.P.; Chang, Y.S.; Chen, S.J.; Chen, H.C. The Epstein-Barr virus-encoded microRNA MiR-BART9 promotes tumor metastasis by targeting E-cadherin in nasopharyngeal carcinoma. PLoS Pathog. 2014, 10, e1003974. [Google Scholar] [CrossRef]
- Yan, Q.; Zeng, Z.; Gong, Z.; Zhang, W.; Li, X.; He, B.; Song, Y.; Li, Q.; Zeng, Y.; Liao, Q.; et al. EBV-miR-BART10-3p facilitates epithelial-mesenchymal transition and promotes metastasis of nasopharyngeal carcinoma by targeting BTRC. Oncotarget 2015, 6, 41766–41782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, K.; Lee, S.K. EBV miR-BART10-3p Promotes Cell Proliferation and Migration by Targeting DKK1. Int. J. Biol. Sci. 2019, 15, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, X.; Zeng, Z.; Li, Q.; Gong, Z.; Liao, Q.; Li, X.; Chen, P.; Xiang, B.; Zhang, W.; et al. Epstein-Barr virus encoded miR-BART11 promotes inflammation-induced carcinogenesis by targeting FOXP1. Oncotarget 2016, 7, 36783–36799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.J.; Zhou, R.; Zong, J.F.; Lin, W.S.; Tong, S.; Guo, Q.J.; Lin, C.; Lin, S.J.; Chen, Y.X.; Chen, M.R.; et al. Epstein-Barr virus-coded miR-BART13 promotes nasopharyngeal carcinoma cell growth and metastasis via targeting of the NKIRAS2/NF-κB pathway. Cancer Lett. 2019, 447, 33–40. [Google Scholar] [CrossRef]
- Choy, H.; Lee, H.; Kim, S.R.; Gho, Y.S.; Lee, S.K. Epstein-Barr virus-encoded microRNA BART15-3p promotes cell apoptosis partially by targeting BRUCE. J. Virol. 2013, 87, 8135–8144. [Google Scholar] [CrossRef] [Green Version]
- Kanda, T.; Miyata, M.; Kano, M.; Kondo, S.; Yoshizaki, T.; Iizasa, H. Clustered microRNAs of the Epstein-Barr virus cooperatively downregulate an epithelial cell-specific metastasis suppressor. J. Virol. 2015, 89, 2684–2697. [Google Scholar] [CrossRef] [Green Version]
- Babu, S.G.; Ponia, S.S.; Kumar, D.; Saxena, S. Cellular oncomiR orthologue in EBV oncogenesis. Comput. Biol. Med. 2011, 41, 891–898. [Google Scholar] [CrossRef]
- Skalsky, R.L.; Cullen, B.R. EBV Noncoding RNAs. Curr. Top. Microbiol. Immunol. 2015, 391, 181–217. [Google Scholar]
Figure 1.
Schematic presentation of the Epstein–Barr virus genome structure (based on References [12,20,21]).

Figure 2.
The role of EBV miRNAs in the immune evasion.


{kind=link}
{kind=link}
Table 1.
Targets of BART miRNAs and processes affected by their interactions in carcinoma cells.
| BART | BART Target | Affected Process | Study | Method |
|---|---|---|---|---|
| 1 | PTEN | migration, signaling | Cai et al. 2015a, Cai et al. 2015b [59,60] | reporter assay |
| 1-5p | α1subunit AMPK | metabolism | Lyu et al. 2018 [61] | reporter assay, biopsies, cell culture |
| 1-5p | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 3 | DICE1 | apoptosis | Kang et al. 2015, Lei et al. 2013 [63,64] | reporter assay, PAR-CLIP |
| 3 | CASZ1a | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 4-5p | BID | apoptosis | Shinozaki-Ushiku et al. 2015 [65] | reporter assay, biopsies |
| 5 | PUMA | apoptosis | Choy et al. 2008 [66] | reporter assay |
| 5-3p | p53 | apoptosis | Zheng et al. 2018 [67] | reporter assay |
| 5-5p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 6-3p | LOC353103 RNA | anti-migration | He et al. 2016 [69] | cell culture, reporter assay |
| 6 | OCT1 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 6 | Dicer | apoptosis | Kang et al. 2015, Iizasa et al. 2010 [63,70] | PAR-CLIP, reporter assay |
| 7-3p | PTEN | migration, signaling | Cai et al. 2015a, Cai et al. 2015b [59,60] | reporter assay |
| 7-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 8-3p | RNF38 | signaling, migration | Lin et al. 2018 [71] | biopsy sequencing, reporter assay |
| 8 | ARID2 | unknown | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 9 | E CAD | migration | Tsai et al. 2017, Hsu et al. 2014 [24,72] | reporter assay, biopsies |
| 9-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 10 | BTRC | signaling, migration | Zeng et al. 2014, Yan et al. 2015 [57,73] | reporter assay, biopsies |
| 10-3p | DKK1 | signaling, migration | Min et al. 2019 [74] | reporter assay |
| 11 | trFOXP1 | immune evasion, differentiation | Song et al. 2016 [75] | reporter assay, biopsies |
| 13 | NKIRAS2 | signaling | Xu et al. 2019 [76] | biopsies, cell culture |
| 14-3p | ATM | DNA repair | Lung et al. 2018 [68] | reporter assay |
| 15-3p | BRUCE | anti-apoptosis | Choy et al. 2013 [77] | reporter assay, WB |
| 16 | CRBBP | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | SH2B3 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | TOMM22 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 16 | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 17-5p | LMP1 | viral latency | Lo et al. 2007 [62] | reporter assay |
| 22 | PAK2 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 22 | TP53INP1 | apoptosis | Kang et al. 2015 [63] | PAR-CLIP, reporter assay |
| 22 and cluster II | NDRG1 | metastasis, differentiation | Kanda et al. 2015 [78] | microarray reporter assay |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Židovec Lepej, S.; Matulić, M.; Gršković, P.; Pavlica, M.; Radmanić, L.; Korać, P. miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis. Pathogens 2020, 9, 353. https://doi.org/10.3390/pathogens9050353
AMA Style
Židovec Lepej S, Matulić M, Gršković P, Pavlica M, Radmanić L, Korać P. miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis. Pathogens. 2020; 9(5):353. https://doi.org/10.3390/pathogens9050353
Chicago/Turabian StyleŽidovec Lepej, Snježana, Maja Matulić, Paula Gršković, Mirjana Pavlica, Leona Radmanić, and Petra Korać. 2020. "miRNAs: EBV Mechanism for Escaping Host’s Immune Response and Supporting Tumorigenesis" Pathogens 9, no. 5: 353. https://doi.org/10.3390/pathogens9050353
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.