Replication of Crohn’s Disease Mucosal E. coli Isolates inside Macrophages Correlates with Resistance to Superoxide and Is Dependent on Macrophage NF-kappa B Activation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
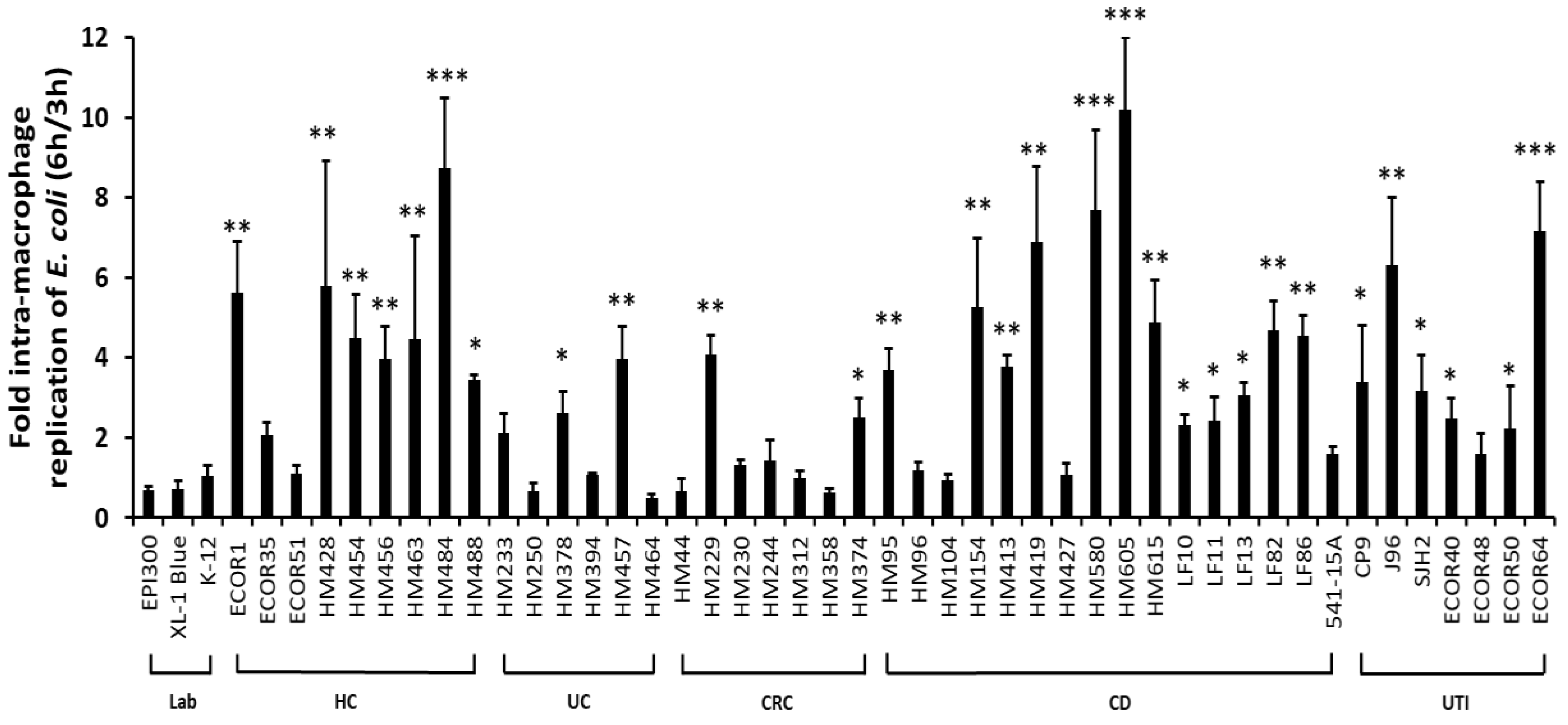
2.1. Ability to Survive and Replicate within Macrophages Is Not Confined Specifically to CD Mucosal E. coli Strains
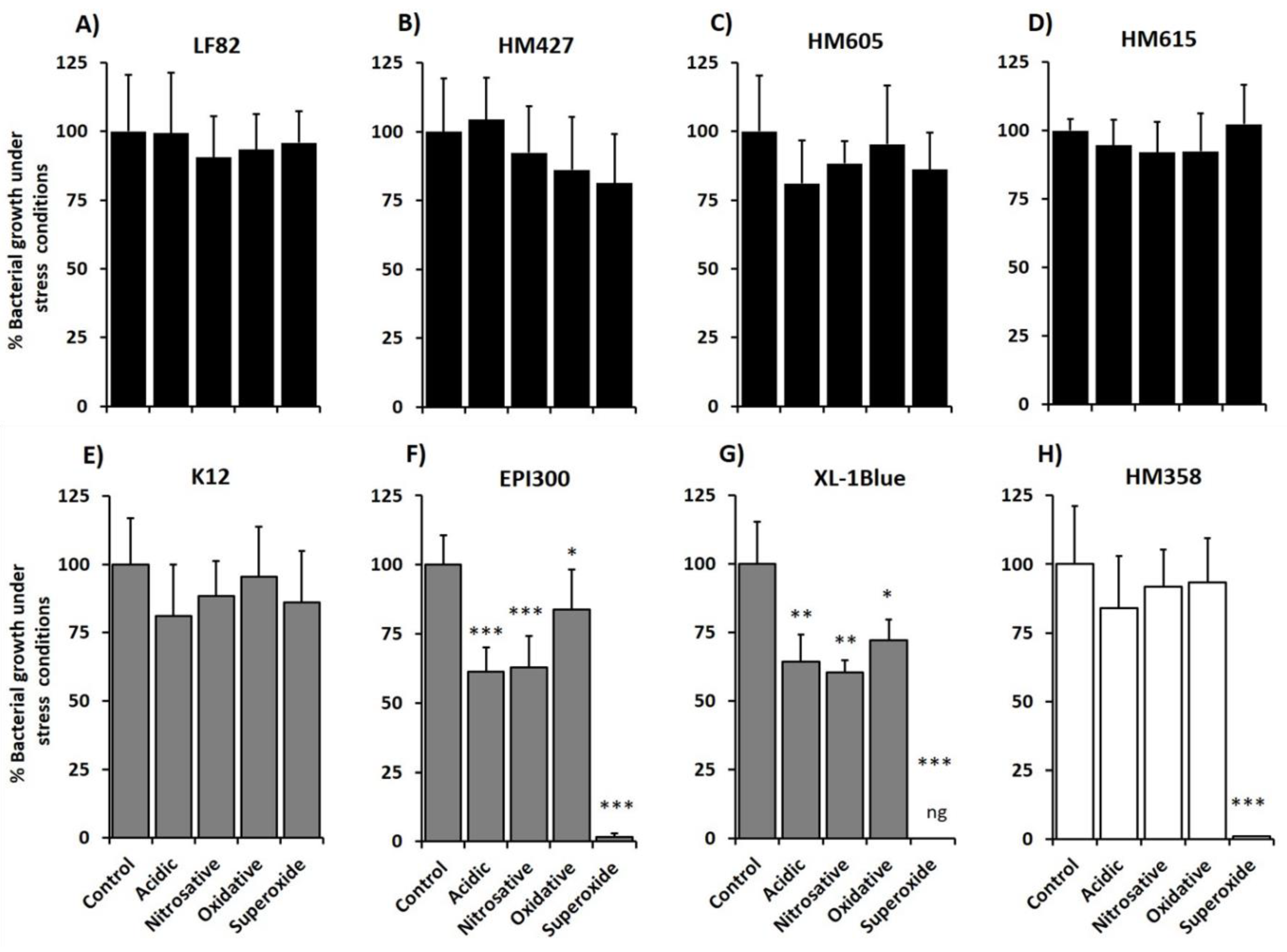
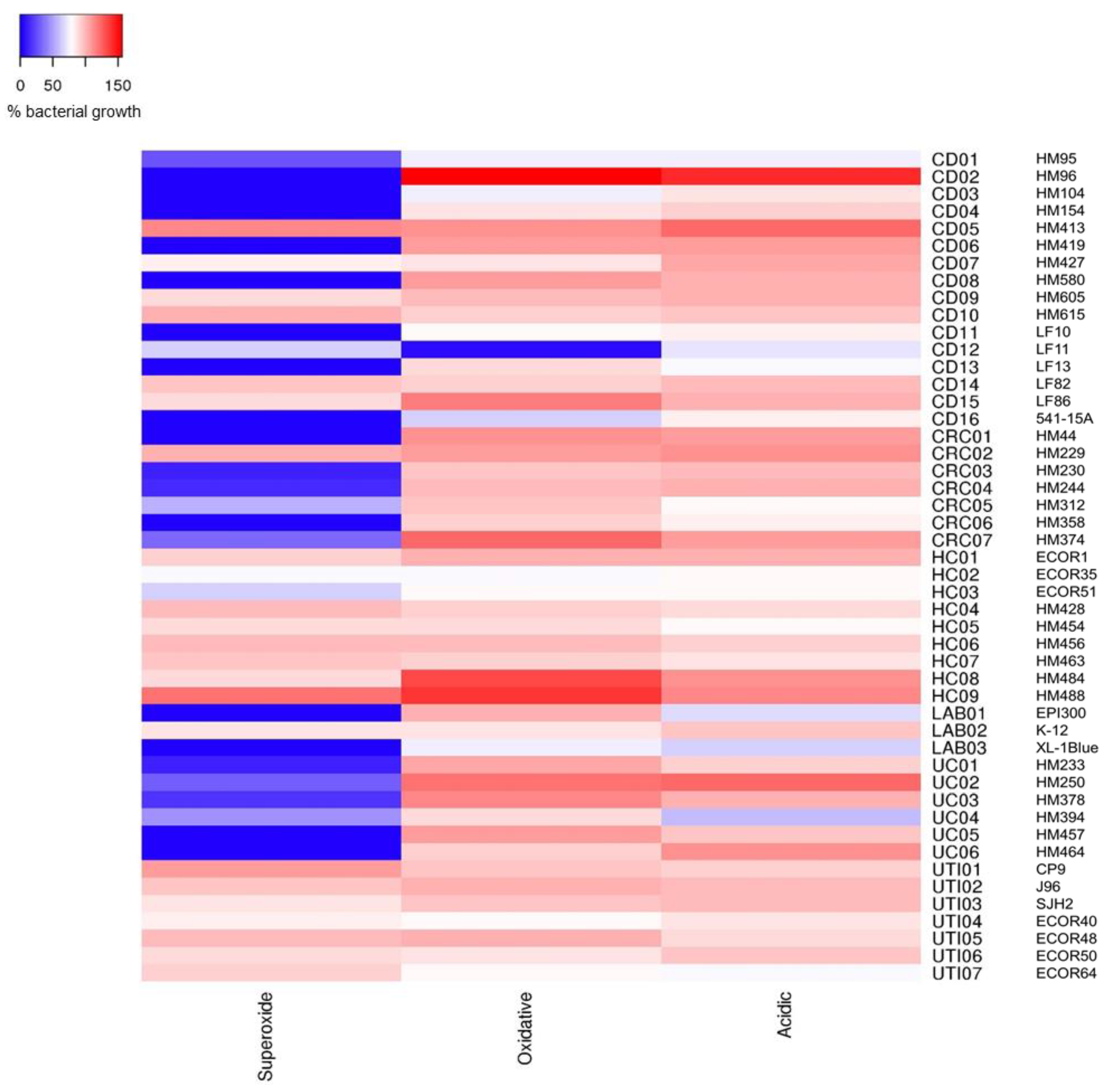
2.2. Crohn’s Disease Mucosa-Associated E. coli Strains Show Ability to Tolerate Stress Conditions that Mimic the Phagolysosome Environment, in Particular Superoxide Stress
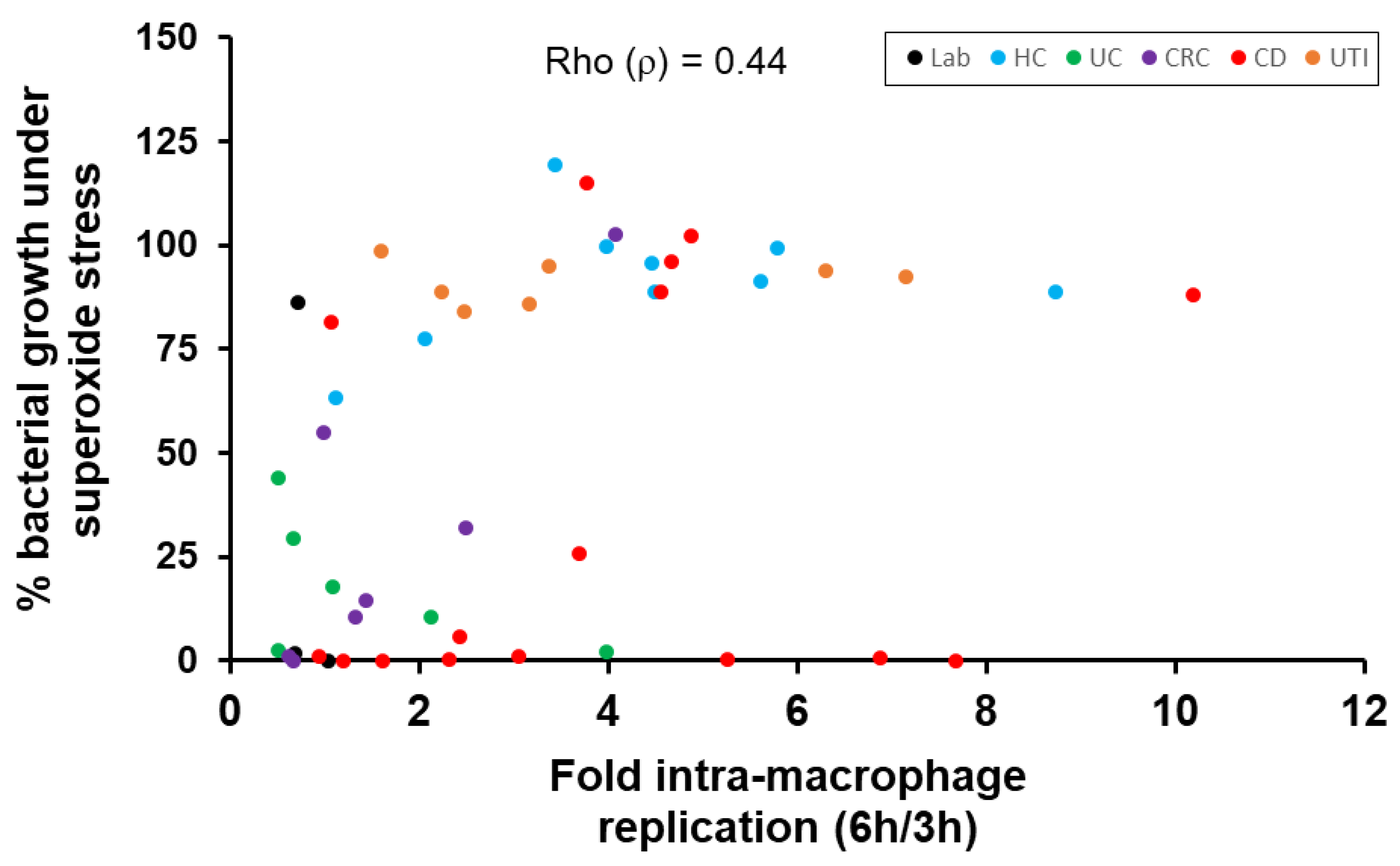
2.3. Correlation of E. coli Tolerance to Methyl Viologen Induced Superoxide Stress with Ability to Replicate within Macrophages
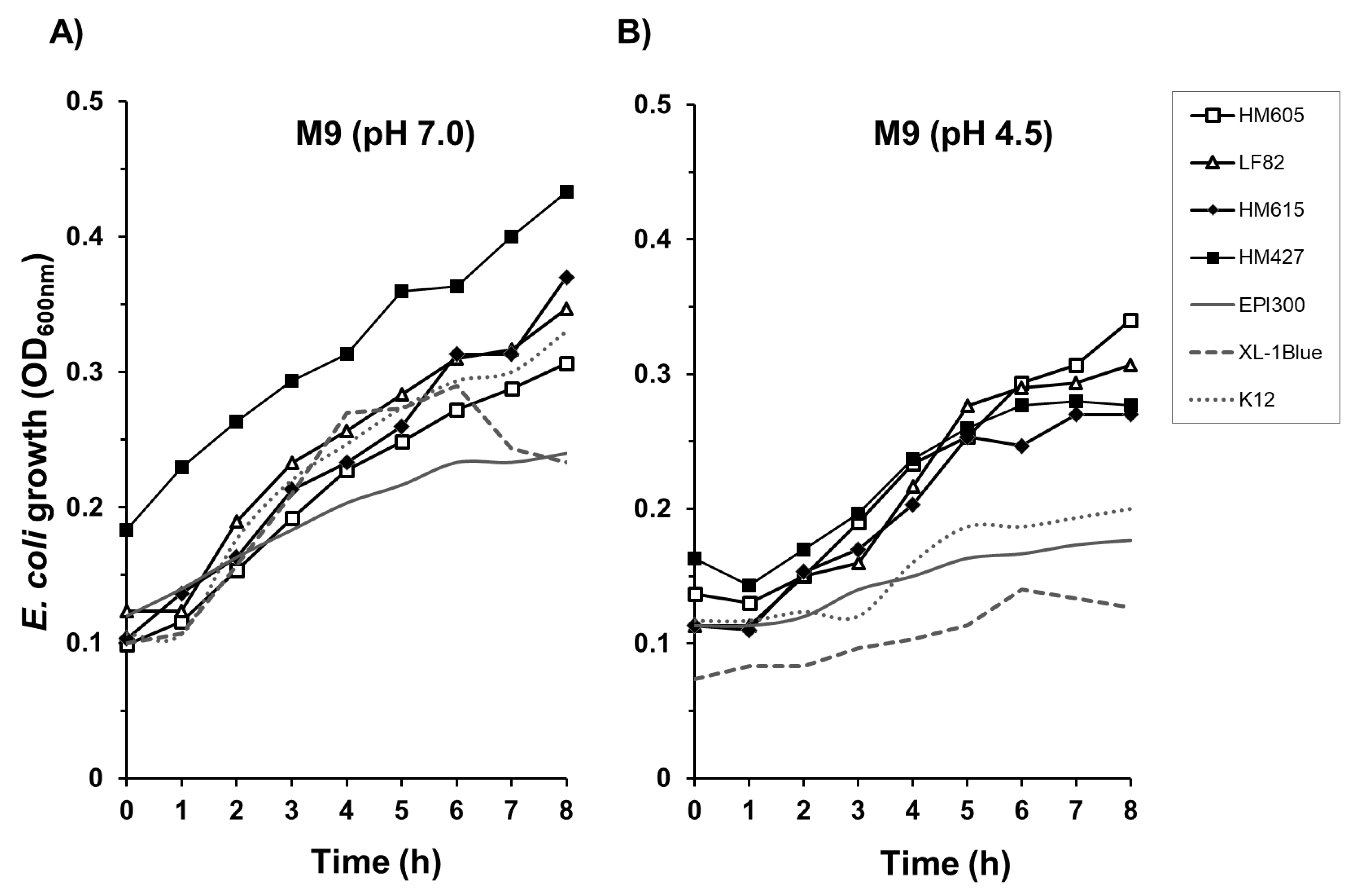
2.4. Crohn’s Disease E. coli Isolates Are Able to Tolerate a Low-Nutrient, Acidic Environment
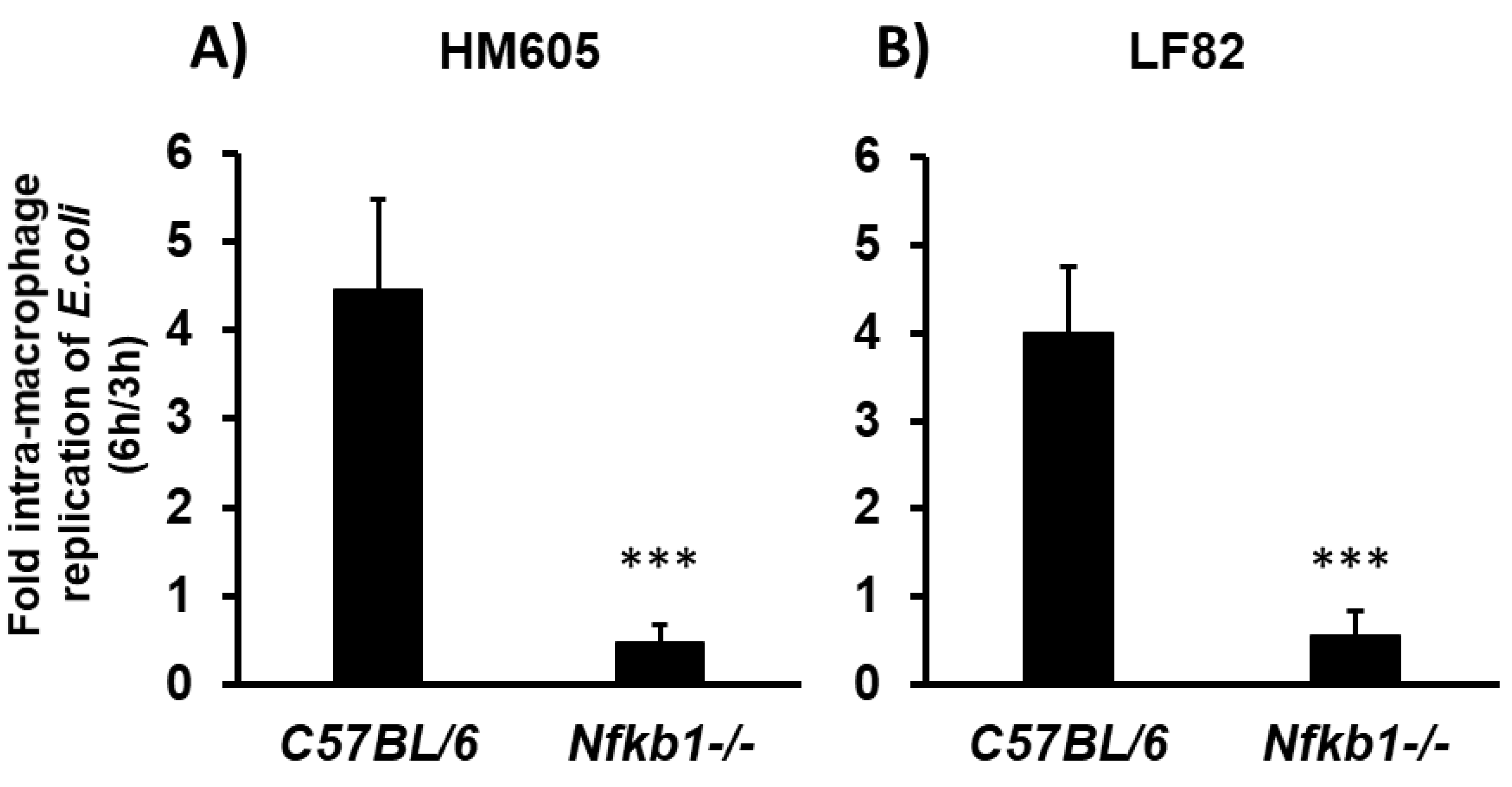
2.5. Crohn’s Disease Mucosal E. coli HM605 and LF82 Replicate inside C57BL/6 Murine Bone Marrow-Derived Macrophages (BMDM), but Are Unable to Survive within Nfκb1-Deficient BMDM
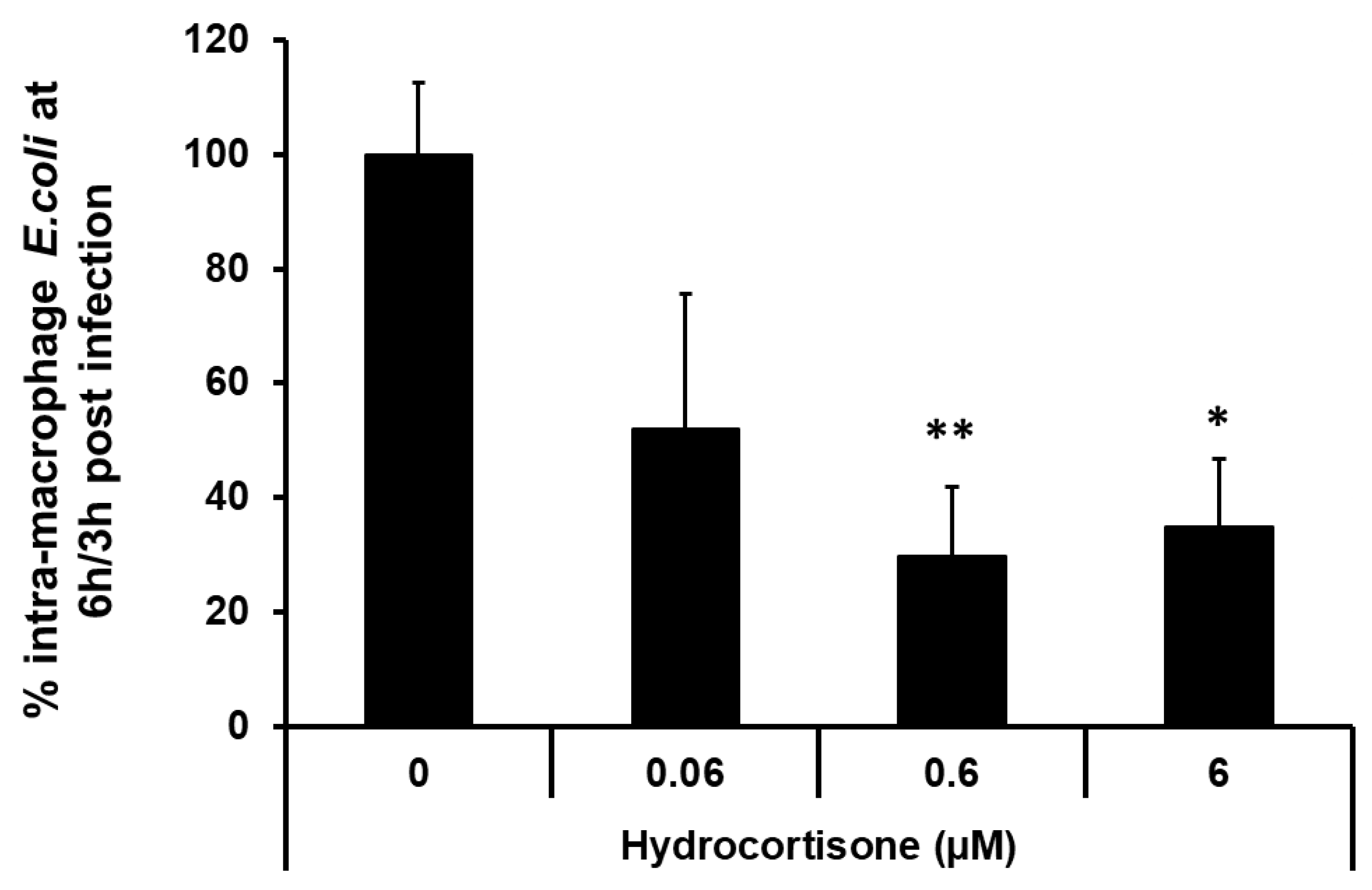
2.6. Macrophage Pre-Incubation with the Corticosteroid Hydrocortisone Inhibits Intra-Macrophage Replication of a Crohn’s Disease Mucosa-Associated E. coli
3. Discussion
4. Materials and Methods
4.1. Murine Bone Marrow Isolation
4.2. Bacteria and Culture Conditions
4.2.1. CD E. coli Strains
4.2.2. UC E. coli Strains
4.2.3. CRC E. coli Strains
4.2.4. Other E. coli Strains
4.3. Bacteria Stress Tolerance Tests
4.4. Bacteria Survival and Growth in Acidic Nutrient-poor M9 Medium
4.5. Intra-Macrophage Replication Assays
4.6. Effect of Hydrocortisone Pre-Treatment of Macrophages on Intracellular Replication of E. coli
4.7. Data Analysis
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darfeuille-Michaud, A.; Neut, C.; Barnich, N.; Lederman, E.; Di Martino, P.; Desreumaux, P.; Gambiez, L.; Joly, B.; Cortot, A.; Colombel, J.F. Presence of adherent Escherichia coli strains in ileal mucosa of patients with Crohn’s disease. Gastroenterology 1998, 115, 1405–1413. [Google Scholar] [CrossRef]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.M.; Campbell, B.J.; Hart, C.A.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer. Gastroenterology 2004, 127, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Kotlowski, R.; Bernstein, C.N.; Sepehri, S.; Krause, D.O. High prevalence of Escherichia coli belonging to the B2+D phylogenetic group in inflammatory bowel disease. Gut 2007, 56, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G.; et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. Isme J. 2007, 1, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, M.; Aldeguer, X.; Lopez-Siles, M.; González-Huix, F.; López-Oliu, C.; Dahbi, G.; Blanco, J.E.; Blanco, J.; Garcia-Gil, J.L.; Darfeuille-Michaud, A. Molecular diversity of Escherichia coli in the human gut: New ecological evidence supporting the role of adherent-invasive E. coli (AIEC) in Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Khilkin, M.; Kerjaschki, D.; Schreiber, S.; Ortner, M.; Weber, J.; Lochs, H. Association between intraepithelial Escherichia coli and colorectal cancer. Gastroenterology 1998, 115, 281–286. [Google Scholar] [CrossRef]
- Sasaki, M.; Sitaraman, S.V.; Babbin, B.A.; Gerner-Smidt, P.; Ribot, E.M.; Garrett, N.; Alpern, J.A.; Akyildiz, A.; Theiss, A.L.; Nusrat, A.; et al. Invasive Escherichia coli are a feature of Crohn’s disease. Lab Investig. 2007, 87, 1042–1054. [Google Scholar] [CrossRef]
- Sepehri, S.; Khafipour, E.; Bernstein, C.N.; Coombes, B.K.; Pilar, A.V.; Karmali, M.; Ziebell, K.; Krause, D.O. Characterization of Escherichia coli isolated from gut biopsies of newly diagnosed patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 1451–1463. [Google Scholar] [CrossRef]
- Darfeuille-Michaud, A. Adherent-invasive Escherichia coli: A putative new E-coli pathotype associated with Crohn’s disease. Int. J. Med. Microbiol. 2002, 292, 185–193. [Google Scholar] [CrossRef]
- Glasser, A.L.; Boudeau, J.; Barnich, N.; Perruchot, M.H.; Colombel, J.F.; Darfeuille-Michaud, A. Adherent invasive Escherichia coli strains from patients with Crohn’s disease survive and replicate within macrophages without inducing host cell death. Infect. Immun. 2001, 69, 5529–5537. [Google Scholar] [CrossRef] [PubMed]
- Bringer, M.A.; Glasser, A.L.; Tung, C.H.; Meresse, S.; Darfeuille-Michaud, A. The Crohn’s disease-associated adherent-invasive Escherichia coli strain LF82 replicates in mature phagolysosomes within J774 macrophages. Cell Microbiol. 2006, 8, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Mpofu, C.M.; Campbell, B.J.; Subramanian, S.; Marshall–Clarke, S.; Hart, C.A.; Cross, A.; Roberts, C.L.; McGoldrick, A.; Edwards, S.W.; Rhodes, J.M. Microbial mannan inhibits bacterial killing by macrophages: A possible pathogenic mechanism for Crohn’s disease. Gastroenterology 2007, 133, 1487–1498. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Roberts, C.L.; Hart, C.A.; Martin, H.M.; Edwards, S.W.; Rhodes, J.M.; Campbell, B.J. Replication of colonic Crohn’s disease mucosal Escherichia coli isolates within macrophages and their susceptibility to antibiotics. Antimicrob Agents Ch. 2008, 52, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Meconi, S.; Vercellone, A.; Levillain, F.; Payré, B.; Al Saati, T.; Capilla, F.; Desreumaux, P.; Darfeuille-Michaud, A.; Altare, F. Adherent-invasive Escherichia coli isolated from Crohn’s disease patients induce granulomas in vitro. Cell Microbiol. 2007, 9, 1252–1261. [Google Scholar] [CrossRef] [PubMed]
- Lissner, D.; Schumann, M.; Batra, A.; Kredel, L.I.; Kühl, A.A.; Erben, U.; May, C.; Schulzke, J.D.; Siegmund, B. Monocyte and M1 Macrophage-induced Barrier Defect Contributes to Chronic Intestinal Inflammation in IBD. Inflamm. Bowel Dis. 2015, 21, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.A.; Cutler, D.J.; Okou, D.; Dodd, A.; Aronow, B.J.; Haberman, Y.; Stevens, C.; Walters, T.D.; Griffiths, A.; Baldassano, R.N.; et al. Genetic variants and pathways implicated in a pediatric inflammatory bowel disease cohort. Genes Immun. 2018, 20, 131. [Google Scholar] [CrossRef]
- Turpin, W.; Goethel, A.; Bedrani, L.; Croitoru Mdcm, K. Determinants of IBD Heritability: Genes, Bugs, and More. Inflamm Bowel Dis. 2018, 24, 1133–1148. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.M.; Rahman, F.Z.; Hayee, B.H.; Graham, S.J.; Marks, D.J.; Sewell, G.W.; Palmer, C.D.; Wilde, J.; Foxwell, B.M.; Gloger, I.S.; et al. Disordered macrophage cytokine secretion underlies impaired acute inflammation and bacterial clearance in Crohn’s disease. J. Exp. Med. 2009, 206, 1883–1897. [Google Scholar] [CrossRef]
- Segal, A.W.; Loewi, G. Neutrophil dysfunction in Crohn’s disease. Lancet 1976, 2, 219–221. [Google Scholar] [CrossRef]
- Segal, A.W. The role of neutrophils in the pathogenesis of Crohn’s disease. Eur. J. Clin. Investig. 2018, 48 (Suppl. 2), e12983. [Google Scholar] [CrossRef]
- Morain, C.O.; Segal, A.A.; Walker, D.; Levi, A.J. Abnormalities of neutrophil function do not cause the migration defect in Crohn’s disease. Gut 1981, 22, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, J.M.; Jewell, D.P. Motility of neutrophils and monocytes in Crohn’s disease and ulcerative colitis. Gut 1983, 24, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Flanagan, P.K.; Chiewchengchol, D.; Wright, H.L.; Edwards, S.W.; Alswied, A.; Satsangi, J.; Subramanian, S.; Rhodes, J.M.; Campbell, B.J. Killing of Escherichia coli by Crohn’s Disease Monocyte-derived Macrophages and Its Enhancement by Hydroxychloroquine and Vitamin D. Inflamm. Bowel Dis. 2015, 21, 1499–1510. [Google Scholar] [CrossRef] [PubMed]
- Elliott, T.R.; Hudspith, B.N.; Rayment, N.B.; Prescott, N.J.; Petrovska, L.; Hermon-Taylor, J.; Brostoff, J.; Boussioutas, A.; Mathew, C.G.; Sanderson, J.D. Defective macrophage handling of Escherichia coli in Crohn’s disease. J. Gastroenterol. Hepatol. 2015, 30, 1265–1274. [Google Scholar] [CrossRef]
- Dale, D.C.; Boxer, L.; Liles, W.C. The phagocytes: Neutrophils and monocytes. Blood 2008, 112, 935–945. [Google Scholar] [CrossRef] [PubMed]
- Segal, A.W. How neutrophils kill microbes. Annu. Rev. Immunol. 2005, 23, 197–223. [Google Scholar] [CrossRef]
- Forman, H.J.; Torres, M. Signaling by the respiratory burst in macrophages. Iubmb Life 2001, 51, 365–371. [Google Scholar]
- Devasagayam, T.P.; Tilak, J.C.; Boloor, K.K.; Sane, K.S.; Ghaskadbi, S.S.; Lele, R.D. Free radicals and antioxidants in human health: Current status and future prospects. J. Assoc. Phys. India 2004, 52, 794–804. [Google Scholar]
- Rahman, K.; Sasaki, M.; Nusrat, A.; Klapproth, J.M. Crohn’s Disease-associated Escherichia coli Survive in Macrophages by Suppressing NFkappaB Signaling. Inflamm. Bowel Dis. 2014, 20, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Bringer, M.A.; Billard, E.; Glasser, A.L.; Colombel, J.F.; Darfeuille-Michaud, A. Replication of Crohn’s disease-associated AIEC within macrophages is dependent on TNF-alpha secretion. Lab. Investig. 2012, 92, 411–419. [Google Scholar] [CrossRef] [PubMed]
- O’brien, C.L.; Bringer, M.A.; Holt, K.E.; Gordon, D.M.; Dubois, A.L.; Barnich, N.; Darfeuille-Michaud, A.; Pavli, P. Comparative genomics of Crohn’s disease-associated adherent-invasive Escherichia coli. Gut 2017, 66, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Darfeuille-Michaud, A. The commensal microbiota and enteropathogens in the pathogenesis of inflammatory bowel diseases. Gastroenterology 2011, 140, 1720–1728. [Google Scholar] [CrossRef] [PubMed]
- Dogan, B.; Suzuki, H.; Herlekar, D.; Sartor, R.B.; Campbell, B.J.; Roberts, C.L.; Stewart, K.; Scherl, E.J.; Araz, Y.; Bitar, P.P.; et al. Inflammation-associated adherent-invasive Escherichia coli are enriched in pathways for use of propanediol and iron and M-cell translocation. Inflamm. Bowel Dis. 2014, 20, 1919–1932. [Google Scholar] [CrossRef] [PubMed]
- Camprubí-Font, C.; Ewers, C.; Lopez-Siles, M.; Martinez-Medina, M. Genetic and phenotypic features to screen for putative Adherent-Invasive Escherichia coli. Front Microbiol. 2019, 10, 108. [Google Scholar] [CrossRef] [PubMed]
- Buc, E.; Dubois, D.; Sauvanet, P.; Raisch, J.; Delmas, J.; Darfeuille-Michaud, A.; Pezet, D.; Bonnet, R. High prevalence of mucosa-associated E. coli producing cyclomodulin and genotoxin in colon cancer. PLoS ONE 2013, 8, e56964. [Google Scholar] [CrossRef]
- Prorok-Hamon, M.; Friswell, M.K.; Alswied, A.; Roberts, C.L.; Song, F.; Flanagan, P.K.; Knight, P.; Codling, C.; Marchesi, J.R.; Winstanley, C.; et al. Colonic mucosa-associated diffusely adherent afaC+ Escherichia coli expressing lpfA and pks are increased in inflammatory bowel disease and colon cancer. Gut 2014, 63, 761–770. [Google Scholar] [CrossRef]
- Bokil, N.J.; Totsika, M.; Carey, A.J.; Stacey, K.J.; Hancock, V.; Saunders, B.M.; Ravasi, T.; Ulett, G.C.; Schembri, M.A.; Sweet, M.J. Intramacrophage survival of uropathogenic Escherichia coli: Differences between diverse clinical isolates and between mouse and human macrophages. Immunobiology 2011, 216, 1164–1171. [Google Scholar] [CrossRef]
- Mavromatis, C.; Bokil, N.J.; Totsika, M.; Kakkanat, A.; Schaale, K.; Cannistraci, C.V.; Ryu, T.; Beatson, S.A.; Ulett, G.C.; Schembri, M.A.; et al. The co-transcriptome of uropathogenic Escherichia coli-infected mouse macrophages reveals new insights into host-pathogen interactions. Cell Microbiol. 2015, 17, 730–746. [Google Scholar] [CrossRef]
- Loui, C.; Chang, A.C.; Lu, S. Role of the ArcAB two-component system in the resistance of Escherichia coli to reactive oxygen stress. BMC Microbiol. 2009, 9, 183. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, C.; Veeger, C. The effects of pH and redox potential on the hydrogen production activity of the hydrogenase from Megasphaera elsdenii. Eur. J. Biochem. FEBS 1981, 114, 209–219. [Google Scholar] [CrossRef]
- Rathman, M.; Sjaastad, M.D.; Falkow, S. Acidification of phagosomes containing Salmonella typhimurium in murine macrophages. Infect. Immun. 1996, 64, 2765–2773. [Google Scholar] [PubMed]
- Fabrega, A.; Vila, J. Salmonella enterica serovar Typhimurium skills to succeed in the host: Virulence and regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed]
- Vandal, O.H.; Nathan, C.F.; Ehrt, S. Acid resistance in Mycobacterium tuberculosis. J. Bacteriol. 2009, 191, 4714–4721. [Google Scholar] [CrossRef]
- Podinovskaia, M.; Lee, W.; Caldwell, S.; Russell, D.G. Infection of macrophages with Mycobacterium tuberculosis induces global modifications to phagosomal function. Cell Microbiol. 2013, 15, 843–859. [Google Scholar] [CrossRef] [PubMed]
- Bringer, M.A.; Barnich, N.; Glasser, A.L.; Bardot, O.; Darfeuille-Michaud, A. HtrA stress protein is involved in intramacrophagic replication of adherent and invasive Escherichia coli strain LF82 isolated from a patient with Crohn’s disease. Infect. Immun. 2005, 73, 712–721. [Google Scholar] [CrossRef]
- Bringer, M.A.; Rolhion, N.; Glasser, A.L.; Darfeuille-Michaud, A. The oxidoreductase DsbA plays a key role in the ability of the Crohn’s disease-associated adherent-invasive Escherichia coli strain LF82 to resist macrophage killing. J. Bacteriol. 2007, 189, 4860–4871. [Google Scholar] [CrossRef]
- Maurin, M.; Benoliel, A.M.; Bongrand, P.; Raoult, D. Phagolysosomes of Coxiella burnetii-infected cell lines maintain an acidic pH during persistent infection. Infect. Immun. 1992, 60, 5013–5016. [Google Scholar]
- Boulos, A.; Rolain, J.M.; Raoult, D. Antibiotic susceptibility of Tropheryma whipplei in MRC5 cells. Antimicrob Agents Chemother. 2004, 48, 747–752. [Google Scholar] [CrossRef]
- Fenollar, F.; Puechal, X.; Raoult, D. Whipple’s disease. N. Engl. J. Med. 2007, 356, 55–66. [Google Scholar] [CrossRef] [PubMed]
- US National Library of Medicine—ClinicalTrials.gov. Antibiotics and Hydroxychloroquine in Crohn’s (APRiCCOT). Available online: https://clinicaltrials.gov/ct2/show/NCT01783106 (accessed on 30 May 2019).
- Mizgerd, J.P.; Lupa, M.M.; Kogan, M.S.; Warren, H.B.; Kobzik, L.; Topulos, G.P. Nuclear factor-kappaB p50 limits inflammation and prevents lung injury during Escherichia coli pneumonia. Am. J. Respir Crit. Care Med. 2003, 168, 810–817. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Sundaram, C.; Reuter, S.; Aggarwal, B.B. Inhibiting NF-kappaB activation by small molecules as a therapeutic strategy. Biochim. Biophys. Acta 2010, 1799, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Merga, Y.J.; O’Hara, A.; Burkitt, M.D.; Duckworth, C.A.; Probert, C.S.; Campbell, B.J.; Pritchard, D.M. Importance of the alternative NF-κB activation pathway in inflammation-associated gastrointestinal carcinogenesis. Am. J. Physiol. Gastrointest Liver Physiol. 2016, 310, G1081–G1090. [Google Scholar] [CrossRef] [PubMed]
- Krachler, A.M.; Woolery, A.R.; Orth, K. Manipulation of kinase signaling by bacterial pathogens. J. Cell Biol. 2011, 195, 1083–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alto, N.M.; Orth, K. Subversion of Cell Signaling by Pathogens. Csh Perspect. Biol. 2012, 4, a006114. [Google Scholar] [CrossRef] [PubMed]
- Meduri, G.U.; Kanangat, S.; Bronze, M.; Patterson, D.R.; Meduri, C.U.; Pak, C.; Tolley, E.A.; Schaberg, D.R. Effects of methylprednisolone on intracellular bacterial growth. Clin. Diagn Lab. Immunol. 2001, 8, 1156–1163. [Google Scholar] [CrossRef]
- Van der Goes, A.; Hoekstra, K.; van den Berg, T.K.; Dijkstra, C.D. Dexamethasone promotes phagocytosis and bacterial killing by human monocytes/macrophages in vitro. J. Leukoc. Biol. 2000, 67, 801–807. [Google Scholar] [CrossRef]
- Belchamber, K.B.; Thomas, C.M.; Dunne, A.E.; Barnes, P.J.; Donnelly, L.E. Comparison of fluticasone propionate and budesonide on COPD macrophage and neutrophil function. Int J. Chron Obs. Pulmon Dis. 2018, 13, 2883–2897. [Google Scholar] [CrossRef]
- Losito, A.; Williams, D.G.; Cooke, G.; Harris, L. The effects on polymorphonuclear leucocyte function of prednisolone and azathioprine in vivo and prednisolone, azathioprine and 6-mercaptopurine in vitro. Clin. Exp. Immunol. 1978, 32, 423–428. [Google Scholar]
- Subramanian, S.; Rhodes, J.M.; Hart, C.A.; Tam, B.; Roberts, C.L.; Smith, S.L.; Corkill, J.E.; Winstanley, C.; Virji, M.; Campbell, B.J. Characterization of epithelial IL-8 response to inflammatory bowel disease mucosal E. coli and its inhibition by mesalamine. Inflamm. Bowel Dis. 2008, 14, 162–175. [Google Scholar] [CrossRef] [PubMed]
- Finnie, I.A.; Campbell, B.J.; Taylor, B.A.; Milton, J.D.; Sadek, S.K.; Yu, L.G.; Rhodes, J.M. Stimulation of colonic mucin synthesis by corticosteroids and nicotine. Clin. Sci. 1996, 91, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.; Durrani, S.; Leiper, K.; Ellis, A.; Morris, A.I.; Rhodes, J.M. Effect of systemic corticosteroid therapy on risk for intra-abdominal or pelvic abscess in non-operated Crohn’s disease. Clin. Gastroenterol. Hepatol. 2005, 3, 1215–1220. [Google Scholar] [CrossRef]
- Rhodes, J.M.; Bartholomew, T.C.; Jewell, D.P. Inhibition of leucocyte motility by drugs used in ulcerative colitis. Gut 1981, 22, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Sha, W.C.; Liou, H.C.; Tuomanen, E.I.; Baltimore, D. Targeted disruption of the p50 subunit of NF-kappa B leads to multifocal defects in immune responses. Cell 1995, 80, 321–330. [Google Scholar] [CrossRef]
- Home Office. Animals (Scientific Procedures) Act 1986. Code of Practice for the Housing and Care of Animals Used in Scientific Procedures. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/516689/0107.pdf (accessed on 29 March 2019).
- Minshawi, F.; White, M.R.H.; Muller, W.; Humphreys, N.; Jackson, D.; Campbell, B.J.; Adamson, A.; Papoutsopoulou, S. Human TNF-Luc reporter mouse: A new model to quantify inflammatory responses. Sci Rep. 2019, 9, 193. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Boudeau, J.; Colombel, J.F.; Neut, C.; Desreumaux, P.; Joly, B.; Cortot, A.; Darfeuille-Michaud, A. Genetically related Escherichia coli strains associated with Crohn’s disease. Gut 2001, 48, 320–325. [Google Scholar] [CrossRef] [PubMed]
- ECOR group—E. coli Reference Collection of Natural Isolates. Available online: http://shigatox.net/new/reference-strains/ecor.html (accessed on 23 February 2019).
- Davis, J.M.; Carvalho, H.M.; Rasmussen, S.B.; O’Brien, A.D. Cytotoxic necrotizing factor type 1 delivered by outer membrane vesicles of uropathogenic Escherichia coli attenuates polymorphonuclear leukocyte antimicrobial activity and chemotaxis. Infect. Immun. 2006, 74, 4401–4408. [Google Scholar] [CrossRef]
- Simonsen, K.T.; Nielsen, G.; Vester Bjerrum, J.; Kruse, T.; Kallipolitis, B.H.; Møller-Jensen, J. A Role for the RNA Chaperone Hfq in Controlling Adherent-Invasive Escherichia coli Colonization and Virulence. PLoS ONE 2011, 6, e16387. [Google Scholar] [CrossRef]
- European Collection of Authenticated Cell Cultures (ECACC); Mouse BALB/c Monocyte Macrophage J774A.1 (ECACC 91051511). Available online: https://www.phe-culturecollections.org.uk/products/celllines/generalcell/detail.jsp?refId=91051511&collection=ecacc_gc (accessed on 30 May 2019).
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tawfik, A.; Knight, P.; Duckworth, C.A.; Pritchard, D.M.; Rhodes, J.M.; Campbell, B.J. Replication of Crohn’s Disease Mucosal E. coli Isolates inside Macrophages Correlates with Resistance to Superoxide and Is Dependent on Macrophage NF-kappa B Activation. Pathogens 2019, 8, 74. https://doi.org/10.3390/pathogens8020074
Tawfik A, Knight P, Duckworth CA, Pritchard DM, Rhodes JM, Campbell BJ. Replication of Crohn’s Disease Mucosal E. coli Isolates inside Macrophages Correlates with Resistance to Superoxide and Is Dependent on Macrophage NF-kappa B Activation. Pathogens. 2019; 8(2):74. https://doi.org/10.3390/pathogens8020074
Chicago/Turabian StyleTawfik, Ahmed, Paul Knight, Carrie A. Duckworth, D. Mark Pritchard, Jonathan M. Rhodes, and Barry J. Campbell. 2019. "Replication of Crohn’s Disease Mucosal E. coli Isolates inside Macrophages Correlates with Resistance to Superoxide and Is Dependent on Macrophage NF-kappa B Activation" Pathogens 8, no. 2: 74. https://doi.org/10.3390/pathogens8020074