
Common Microbial Genital Infections and Their Impact on the Innate Immune Response to HPV in Cervical Cells
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Group and Sample Collection
2.2. HPV DNA Detection and Genotyping
2.3. BV- and STI-Detection
2.4. Gene Expression Quantification and Data Analysis
3. Results
3.1. HPV, BV, and STI Distribution in the Study Group
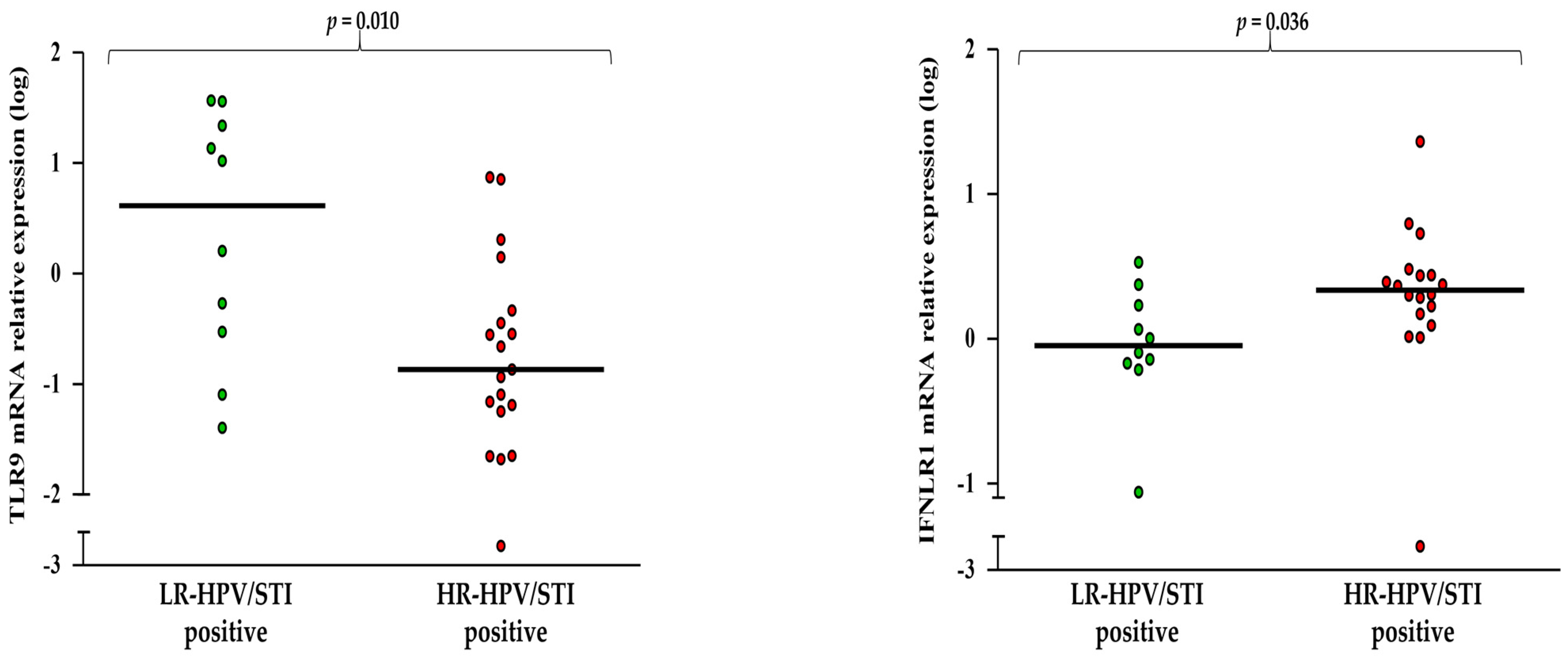
3.2. How Gene Expression Levels Vary According to HPV, BV, and STI Status
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burchell, A.N.; Winer, R.L.; de Sanjosé, S.; Franco, E. Chapter 6: Epidemiology and transmission dynamics of genital HPV infection. Vaccine 2006, 24 (Suppl. S3), S52–S61. [Google Scholar] [CrossRef]
- Muñoz, N.; Bosch, F.X.; de Sanjosé, S.; Herrero, R.; Castellsagué, X.; Shah, K.V.; Snijders, P.J.F.; Meijer, C.J.; International Agency for Research on Cancer Multicenter Cervical Cancer Study Group. Epidemiologic Classification of Human Papillomavirus Types Associated with Cervical Cancer. N. Engl. J. Med. 2003, 348, 518–527. [Google Scholar] [CrossRef] [Green Version]
- IARC Working Group. IARC Monographs–100B: Human Papillomaviruses; WHO Press, International Agency for Research on Cancer: Lyon, France, 2012; Available online: http://monographs.iarc.fr/ENG/Monographs/vol100B/mono100B-11.pdf (accessed on 1 September 2022).
- Stanley, M. Host defence and persistent human papillomavirus infection. Curr. Opin. Virol. 2021, 51, 106–110. [Google Scholar] [CrossRef]
- Medzhitov, R. Toll-like receptors and innate immunity. Nat. Rev. Immunol. 2001, 1, 135–145. [Google Scholar] [CrossRef]
- McClure, R.; Emassari, P. TLR-Dependent Human Mucosal Epithelial Cell Responses to Microbial Pathogens. Front. Immunol. 2014, 5, 386. [Google Scholar] [CrossRef] [Green Version]
- Daud, I.I.; Scott, M.E.; Ma, Y.; Shiboski, S.; Farhat, S.; Moscicki, A.-B. Association between toll-like receptor expression and human papillomavirus type 16 persistence. Int. J. Cancer 2011, 128, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Pierangeli, A.; Degener, A.; Ferreri, M.; Riva, E.; Rizzo, B.; Turriziani, O.; Luciani, S.; Scagnolari, C.; Antonelli, G. Interferon-Induced Gene Expression in Cervical Mucosa during Human Papillomavirus Infection. Int. J. Immunopathol. Pharmacol. 2011, 24, 217–223. [Google Scholar] [CrossRef]
- Sommereyns, C.; Paul, S.; Staeheli, P.; Michiels, T. IFN-Lambda (IFN-λ) Is Expressed in a Tissue-Dependent Fashion and Primarily Acts on Epithelial Cells In Vivo. PLOS Pathog. 2008, 4, e1000017. [Google Scholar] [CrossRef]
- Cannella, F.; Scagnolari, C.; Selvaggi, C.; Stentella, P.; Recine, N.; Antonelli, G.; Pierangeli, A. Interferon lambda 1 expression in cervical cells differs between low-risk and high-risk human papillomavirus-positive women. Med. Microbiol. Immunol. 2014, 203, 177–184. [Google Scholar] [CrossRef]
- Chensue, S.W. Molecular machinations: Chemokine signals in host-pathogen interactions. Clin. Microbiol. Rev. 2001, 14, 821–835. [Google Scholar] [CrossRef]
- Cremel, M.; Berlier, W.; Hamzeh, H.; Cognasse, F.; Lawrence, P.; Genin, C.; Bernengo, J.-C.; Lambert, C.; Dieu-Nosjean, M.-C.; Delézay, O. Characterization of CCL20 secretion by human epithelial vaginal cells: Involvement in Langerhans cell precursor attraction. J. Leukoc. Biol. 2005, 78, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Guess, J.C.; McCance, D.J. Decreased migration of Langerhans precursor-like cells in response to human keratinocytes expressing human papillomavirus type 16 E6/E7 is related to reduced macrophage inflammatory protein-3alpha production. J. Virol. 2005, 79, 14852–14862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, B.; Xue, M. Correlation of E6 and E7 levels in high-risk HPV16 type cervical lesions with CCL20 and Langerhans cells. Genet Mol. Res. 2015, 14, 10473–10481. [Google Scholar] [CrossRef] [PubMed]
- Verteramo, R.; Pierangeli, A.; Mancini, E.; Calzolari, E.; Bucci, M.; Osborn, J.; Nicosia, R.; Chiarini, F.; Antonelli, G.; Degener, A.M. Human Papillomaviruses and genital co-infections in gynaecological outpatients. BMC Infect. Dis. 2009, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Sieving, R.E.; O’Brien, J.R.G.; Saftner, M.A.; Argo, T.A. Sexually Transmitted Diseases Among US Adolescents and Young Adults: Patterns, Clinical Considerations, and Prevention. Nurs. Clin. N. Am. 2019, 54, 207–225. [Google Scholar] [CrossRef]
- Ginocchio, C.C.; Chapin, K.; Smith, J.S.; Aslanzadeh, J.; Snook, J.; Hill, C.S.; Gaydos, C.A. Prevalence of Trichomonas vaginalis and coinfection with Chlamydia trachomatis and Neisseria gonorrhoeae in the United States as determined by the Aptima Trichomonas vaginalis nucleic acid amplification assay. J. Clin. Microbiol. 2012, 50, 2601–2608. [Google Scholar] [CrossRef] [Green Version]
- Leli, C.; Mencacci, A.; Latino, M.A.; Clerici, P.; Rassu, M.; Perito, S.; Castronari, R.; Pistoni, E.; Luciano, E.; De Maria, D.; et al. Prevalence of cervical colonization by Ureaplasma parvum, Ureaplasma urealyticum, Mycoplasma hominis and Mycoplasma genitalium in childbearing age women by a commercially available multiplex real-time PCR: An Italian observational multicentre study. J. Microbiol. Immunol. Infect. 2017, 51, 220–225. [Google Scholar] [CrossRef]
- Amabebe, E.; Anumba, D.O.C. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front. Med. 2018, 5, 181. [Google Scholar] [CrossRef] [Green Version]
- Fredricks, D.N.; Fiedler, T.L.; Marrazzo, J.M. Molecular identification of bacteria associated with bacterial vaginosis. N. Engl. J. Med. 2005, 353, 1899–1911. [Google Scholar] [CrossRef] [Green Version]
- Coudray, M.S.; Madhivanan, P. Bacterial vaginosis—A brief synopsis of the literature. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 245, 143–148. [Google Scholar] [CrossRef]
- Cherpes, T.L.; Meyn, L.A.; Krohn, M.A.; Lurie, J.G.; Hillier, S.L. Association between acquisition of herpes simplex virus type 2 in women and bacterial vaginosis. Clin. Infect. Dis. 2003, 37, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Masson, L.; Mlisana, K.; Little, F.; Werner, L.; Mkhize, N.N.; Ronacher, K.; Gamieldien, H.; Williamson, C.; Mckinnon, L.R.; Walzl, G.; et al. Defining genital tract cytokine signatures of sexually transmitted infections and bacterial vaginosis in women at high risk of HIV infection: A cross-sectional study. Sex. Transm. Infect. 2014, 90, 580–587. [Google Scholar] [CrossRef]
- Mao, C.; Hughes, J.P.; Kiviat, N.; Kuypers, J.; Lee, S.-K.; Adam, D.E.; Koutsky, L.A. Clinical findings among young women with genital human papillomavirus infection. Am. J. Obstet. Gynecol. 2003, 188, 677–684. [Google Scholar] [CrossRef]
- Nam, K.H.; Kim, Y.T.; Kim, S.R.; Kim, S.W.; Kim, J.W.; Lee, M.K.; Nam, E.J.; Jung, Y.W. Association between bacterial vaginosis and cervical intraepithelial neoplasia. J. Gynecol. Oncol. 2009, 20, 39–43. [Google Scholar] [CrossRef] [Green Version]
- Gillet, E.; Meys, J.F.; Verstraelen, H.; Bosire, C.; De Sutter, P.; Temmerman, M.; Broeck, D.V. Bacterial vaginosis is associated with uterine cervical human papillomavirus infection: A meta-analysis. BMC Infect. Dis. 2011, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Shannon, B.; Yi, T.J.; Perusini, S.; Gajer, P.; Ma, B.; Humphrys, M.S.; Thomas-Pavanel, J.; Chieza, L.; Janakiram, P.; Saunders, M.; et al. Association of HPV infection and clearance with cervicovaginal immunology and the vaginal microbiota. Mucosal Immunol. 2017, 10, 1310–1319. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, C.; Marrazzo, J. Bacterial vaginosis and the cervicovaginal immune response. Am. J. Reprod. Immunol. 2014, 71, 555–563. [Google Scholar] [CrossRef] [Green Version]
- Torcia, M.G. Interplay among Vaginal Microbiome, Immune Response and Sexually Transmitted Viral Infections. Int. J. Mol. Sci. 2019, 20, 266. [Google Scholar] [CrossRef] [Green Version]
- Kero, K.; Rautava, J.; Syrjänen, K.; Grenman, S. Association of asymptomatic bacterial vaginosis with persistence of female genital human papillomavirus infection. Eur. J. Clin. Microbiol. 2017, 36, 2215–2219. [Google Scholar] [CrossRef]
- Usyk, M.; Zolnik, C.P.; Castle, P.E.; Porras, C.; Herrero, R.; Gradissimo, A.; Gonzalez, P.; Safaeian, M.; Schiffman, M.R.; Burk, R.D.; et al. Cervicovaginal microbiome and natural history of HPV in a longitudinal study. PLOS Pathog. 2020, 16, e1008376. [Google Scholar] [CrossRef]
- Nugent, R.P.; A Krohn, M.; Hillier, S.L. Reliability of diagnosing bacterial vaginosis is improved by a standardized method of gram stain interpretation. J. Clin. Microbiol. 1991, 29, 297–301. [Google Scholar] [CrossRef] [Green Version]
- Cannella, F.; Pierangeli, A.; Scagnolari, C.; Cacciotti, G.; Tranquilli, G.; Stentella, P.; Recine, N.; Antonelli, G. TLR9 is expressed in human papillomavirus-positive cervical cells and is overexpressed in persistent infections. Immunobiology 2015, 220, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Cigno, I.L.; Calati, F.; Albertini, S.; Gariglio, M. Subversion of Host Innate Immunity by Human Papillomavirus Oncoproteins. Pathogens 2020, 9, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, M.E.; Ma, Y.; Farhat, S.; Moscicki, A.-B. Expression of nucleic acid-sensing Toll-like receptors predicts HPV16 clearance associated with an E6-directed cell-mediated response. Int. J. Cancer 2014, 136, 2402–2408. [Google Scholar] [CrossRef] [Green Version]
- Halec, G.; Scott, M.E.; Farhat, S.; Darragh, T.M.; Moscicki, A.-B. Toll-like receptors: Important immune checkpoints in the regression of cervical intra-epithelial neoplasia 2. Int. J. Cancer 2018, 143, 2884–2891. [Google Scholar] [CrossRef] [Green Version]
- O’Hanlon, D.E.; Gajer, P.; Brotman, R.M.; Ravel, J. Asymptomatic Bacterial Vaginosis Is Associated with Depletion of Mature Superficial Cells Shed from the Vaginal Epithelium. Front. Cell Infect. Microbiol. 2020, 10, 106. [Google Scholar] [CrossRef]
- Moscicki, A.-B.; Shi, B.; Huang, H.; Barnard, E.; Li, H. Cervical-Vaginal Microbiome and Associated Cytokine Profiles in a Prospective Study of HPV 16 Acquisition, Persistence, and Clearance. Front. Cell Infect. Microbiol. 2020, 10, 569022. [Google Scholar] [CrossRef]
- Hasan, U.A.; Bates, E.; Takeshita, F.; Biliato, A.; Accardi, R.; Bouvard, V.; Mansour, M.; Vincent, I.; Gissmann, L.; Iftner, T.; et al. TLR9 expression and function is abolished by the cervical cancer-associated human papillomavirus type 16. J. Immunol. 2007, 178, 3186–3197. [Google Scholar] [CrossRef] [Green Version]
- Scagnolari, C.; Cannella, F.; Pierangelli, A.; Pilgrim, R.M.; Antonelli, G.; Rowley, D.; Wong, M.; Best, S.; Xing, D.; Roden, R.B.S.; et al. Insights into the Role of Innate Immunity in Cervicovaginal Papillomavirus Infection from Studies Using Gene-Deficient Mice. J. Virol. 2020, 94, e00087-20. [Google Scholar] [CrossRef]
- Lee, J.-W.; Choi, J.-J.; Seo, E.S.; Kim, M.J.; Kim, W.Y.; Choi, C.H.; Kim, T.-J.; Kim, B.-G.; Song, S.Y.; Bae, D.-S. Increased toll-like receptor 9 expression in cervical neoplasia. Mol. Carcinog. 2007, 46, 941–947. [Google Scholar] [CrossRef]
- Hao, Y.; Yuan, J.-L.; Abudula, A.; Hasimu, A.; Kadeer, N.; Guo, X. TLR9 expression in uterine cervical lesions of Uyghur women correlate with cervical cancer progression and selective silencing of human papillomavirus 16 E6 and E7 oncoproteins in vitro. Asian Pac. J. Cancer Prev. 2014, 15, 5867–5872. [Google Scholar] [CrossRef] [Green Version]
- Baruah, P.; Bullenkamp, J.; Wilson, P.O.G.; Lee, M.; Kaski, J.C.; Dumitriu, I.E. TLR9 Mediated Tumor-Stroma Interactions in Human Papilloma Virus (HPV)-Positive Head and Neck Squamous Cell Carcinoma Up-Regulate PD-L1 and PD-L2. Front. Immunol. 2019, 10, 1644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Odendall, C.; Voak, A.A.; Kagan, J.C. Type III IFNs are commonly induced by bacteria-sensing TLRs and reinforce epithelial barriers during infection. J. Immunol. 2017, 199, 3270–3279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alphonse, N.; Dickenson, R.E.; Alrehaili, A.; Odendall, C. Functions of IFNλs in Anti-Bacterial Immunity at Mucosal Barriers. Front. Immunol. 2022, 13, 857639. [Google Scholar] [CrossRef] [PubMed]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda interferon (IFN-lambda), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. J. Virol. 2006, 80, 4501–4509. [Google Scholar] [CrossRef] [Green Version]
- Lazear, H.M.; Schoggins, J.W.; Diamond, M.S. Shared and Distinct Functions of Type I and Type III Interferons. Immunity 2019, 50, 907–923. [Google Scholar] [CrossRef]
- Broggi, A.; Granucci, F.; Zanoni, I. Type III interferons: Balancing tissue tolerance and resistance to pathogen invasion. J. Exp. Med. 2019, 217, e20190295. [Google Scholar] [CrossRef]
- Stanifer, M.L.; Pervolaraki, K.; Boulant, S. Differential Regulation of Type I and Type III Interferon Signaling. Int. J. Mol. Sci. 2019, 20, 1445. [Google Scholar] [CrossRef] [Green Version]
- Pierangeli, A.; Statzu, M.; Nenna, R.; Santinelli, L.; Petrarca, L.; Frassanito, A.; Gentile, M.; Antonelli, G.; Midulla, F.; Scagnolari, C. Interferon lambda receptor 1 (IFNL1R) transcript is highly expressed in rhinovirus bronchiolitis and correlates with disease severity. J. Clin. Virol. 2018, 102, 101–109. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Age Group | HPV NEG | LR HPV a | HR HPV a | p |
|---|---|---|---|---|
| 18–27 (N = 35) | 12 (34.3%) | 8 (22.8%) 6 [1]; 44 [1]; 54 [1]; 61 [2]; 81 [2]; 84 [1] | 15 (42.9%) 16 [3]; 18 [2]; 31 [1]; 33 [1]; 52 [1]: 53 [3]; 56 [1]; 58 [1]; 59 [1]; 66 [1] | 0.007 |
| 28–37 (N = 57) | 36 (63.1%) | 5 (8.8%) 6 [2]; 11 [1]; 54 [1]; 84 [1] | 16 (28.1%) 16 [5]; 31 [3]; 53 [1]; 58 [3]; 66 [2]; 68 [1]; 70 [1] | |
| 38–47 (N = 28) | 21 (75.0%) | 4 (14.3%) 54 [2]; 61 [1]; 83 [1] | 3 (10.7%) 16 [1]; 18 [1]; 58 [1] |
| BV-Rt a | BV-Nu b NEG | BV-Nu b POS | HPV NEG | HPV POS | STI NEG | STI POS |
|---|---|---|---|---|---|---|
| BV-Rt-negative | 30 | 0 | 21 | 9 | 17 | 13 |
| BV-Rt-intermediate | 16 | 12 | 18 | 12 | 10 | 20 |
| BV-Rt-full positive | 6 | 47 | 27 | 29 | 13 | 43 |
| Vaginosis-Associated Bacteria a | HPV | |||
|---|---|---|---|---|
| Negative | Positive | p Value | ||
| Gardnerella vaginalis | Negative | 27 | 11 | 0.045 |
| Positive | 39 | 39 | ||
| Atopobium vaginae | Negative | 41 | 21 | 0.039 |
| Positive | 25 | 29 | ||
| Megasphaera type 1 | Negative | 54 | 37 | 0.365 |
| Positive | 12 | 13 | ||
| Bacteroides fragilis | Negative | 63 | 50 | 0.258 |
| Positive | 3 | 0 | ||
| Mobiluncus spp. | Negative | 58 | 46 | 0.550 |
| Positive | 8 | 4 | ||
| Bacterial vaginosis–associated bacteria 2 | Negative | 56 | 39 | 0.466 |
| Positive | 10 | 11 | ||
| Sexually Transmitted Infections a | HPV | BV | |||
|---|---|---|---|---|---|
| Negative | Positive | Negative | Positive | ||
| Ureaplasma parvum | Negative | 40 | 25 | 34 | 28 |
| Positive | 26 | 25 | 18 | 30 | |
| Ureaplasma urealiticum | Negative | 63 | 44 | 50 | 52 |
| Positive | 3 | 6 | 2 | 6 | |
| Mycoplasma genitalium | Negative | 65 | 49 | 51 | 58 |
| Positive | 1 | 1 | 1 | 0 | |
| Mycoplasma hominis | Negative | 57 | 44 | 46 | 51 |
| Positive | 9 | 6 | 6 | 7 | |
| Trichomonas vaginalis | Negative | 66 | 49 | 52 | 57 |
| Positive | 0 | 1 | 0 | 1 | |
| Chlamydia trachomatis | Negative | 63 | 47 | 52 | 52 |
| Positive | 3 | 3 | 0 | 6 | |
| Candida albicans/glabrata/krusei | Negative | 51 | 37 | 43 | 40 |
| Positive | 15 | 13 | 9 | 18 | |
| All STIs | Negative | 27 | 13 | 26 | 12 |
| Positive | 39 | 37 | 26 | 46 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fracella, M.; Oliveto, G.; Sorrentino, L.; Roberto, P.; Cinti, L.; Viscido, A.; Di Lella, F.M.; Giuffrè, F.; Gentile, M.; Pietropaolo, V.; et al. Common Microbial Genital Infections and Their Impact on the Innate Immune Response to HPV in Cervical Cells. Pathogens 2022, 11, 1361. https://doi.org/10.3390/pathogens11111361
Fracella M, Oliveto G, Sorrentino L, Roberto P, Cinti L, Viscido A, Di Lella FM, Giuffrè F, Gentile M, Pietropaolo V, et al. Common Microbial Genital Infections and Their Impact on the Innate Immune Response to HPV in Cervical Cells. Pathogens. 2022; 11(11):1361. https://doi.org/10.3390/pathogens11111361
Chicago/Turabian StyleFracella, Matteo, Giuseppe Oliveto, Leonardo Sorrentino, Piergiorgio Roberto, Lilia Cinti, Agnese Viscido, Federica Maria Di Lella, Federica Giuffrè, Massimo Gentile, Valeria Pietropaolo, and et al. 2022. "Common Microbial Genital Infections and Their Impact on the Innate Immune Response to HPV in Cervical Cells" Pathogens 11, no. 11: 1361. https://doi.org/10.3390/pathogens11111361