
Culture Supernatant of Enterococcus faecalis Promotes the Hyphal Morphogenesis and Biofilm Formation of Candida albicans
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
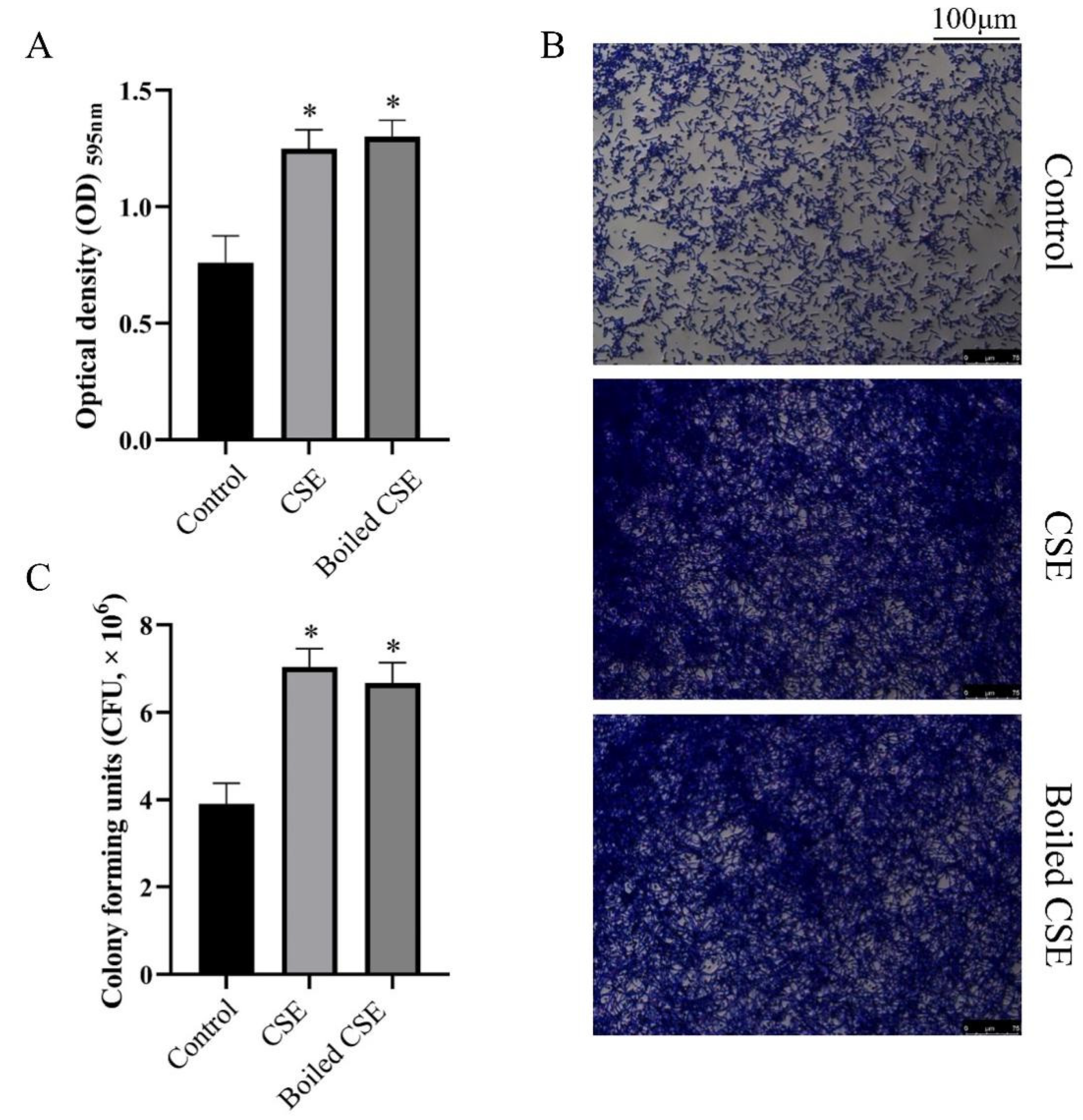
2.1. Culture Supernatant of E. faecalis (CSE) Could Promote the Biofilm Formation of C. albicans
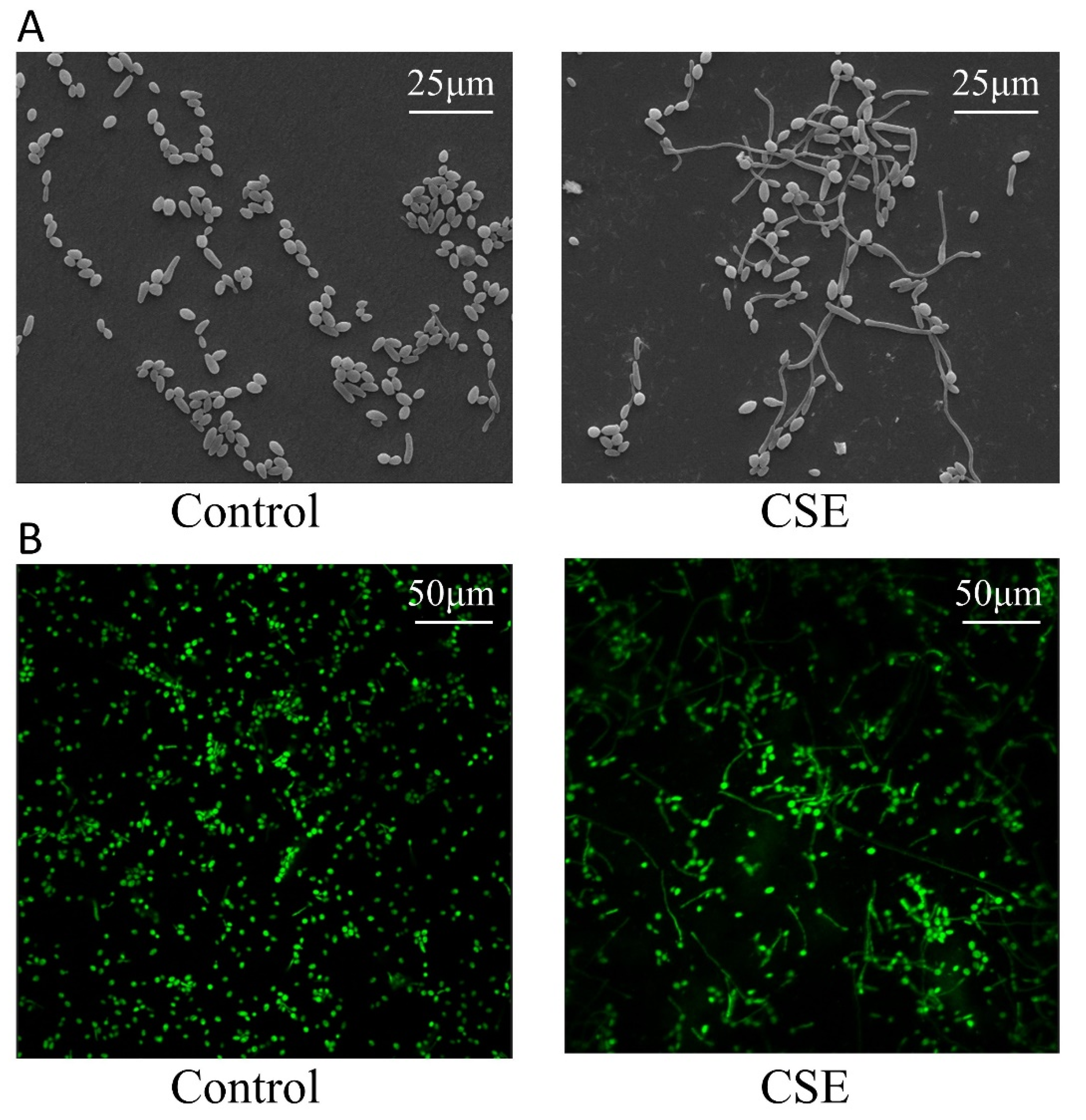
2.2. CSE Promoted Hyphal Morphogenesis of C. albicans
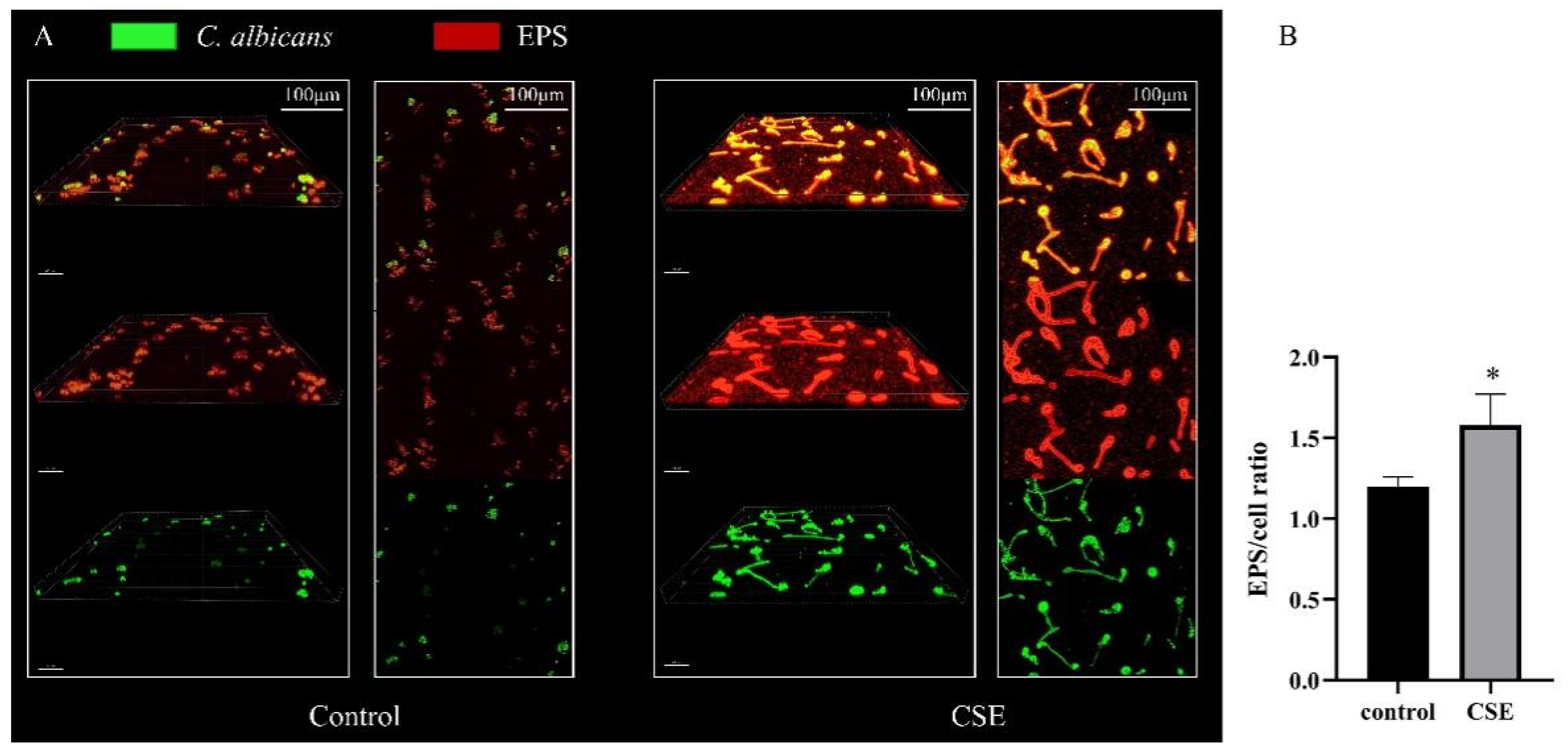
2.3. CSE Increased the EPS Production of C. albicans Biofilms
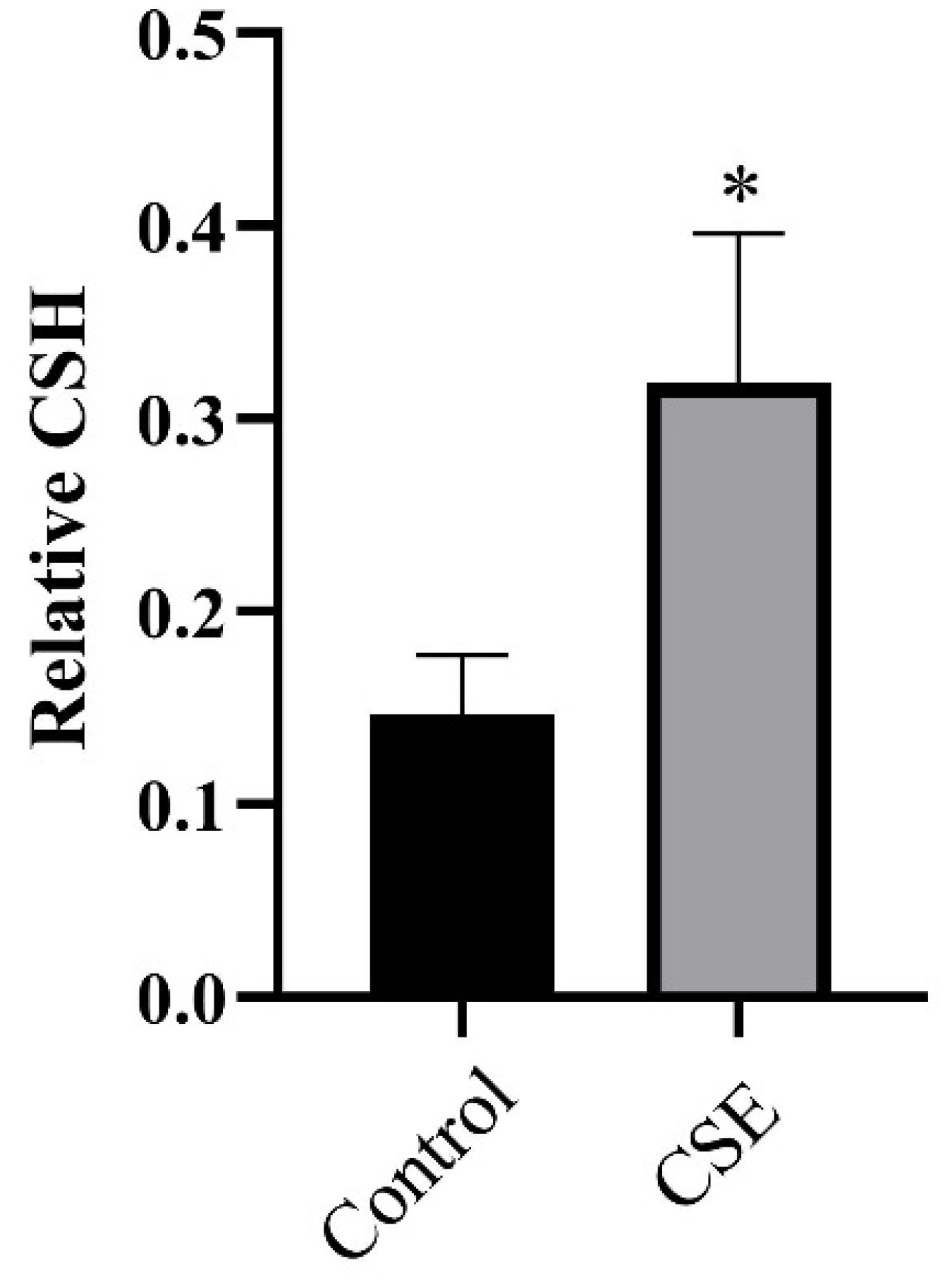
2.4. CSE Altered CSH of C. albicans
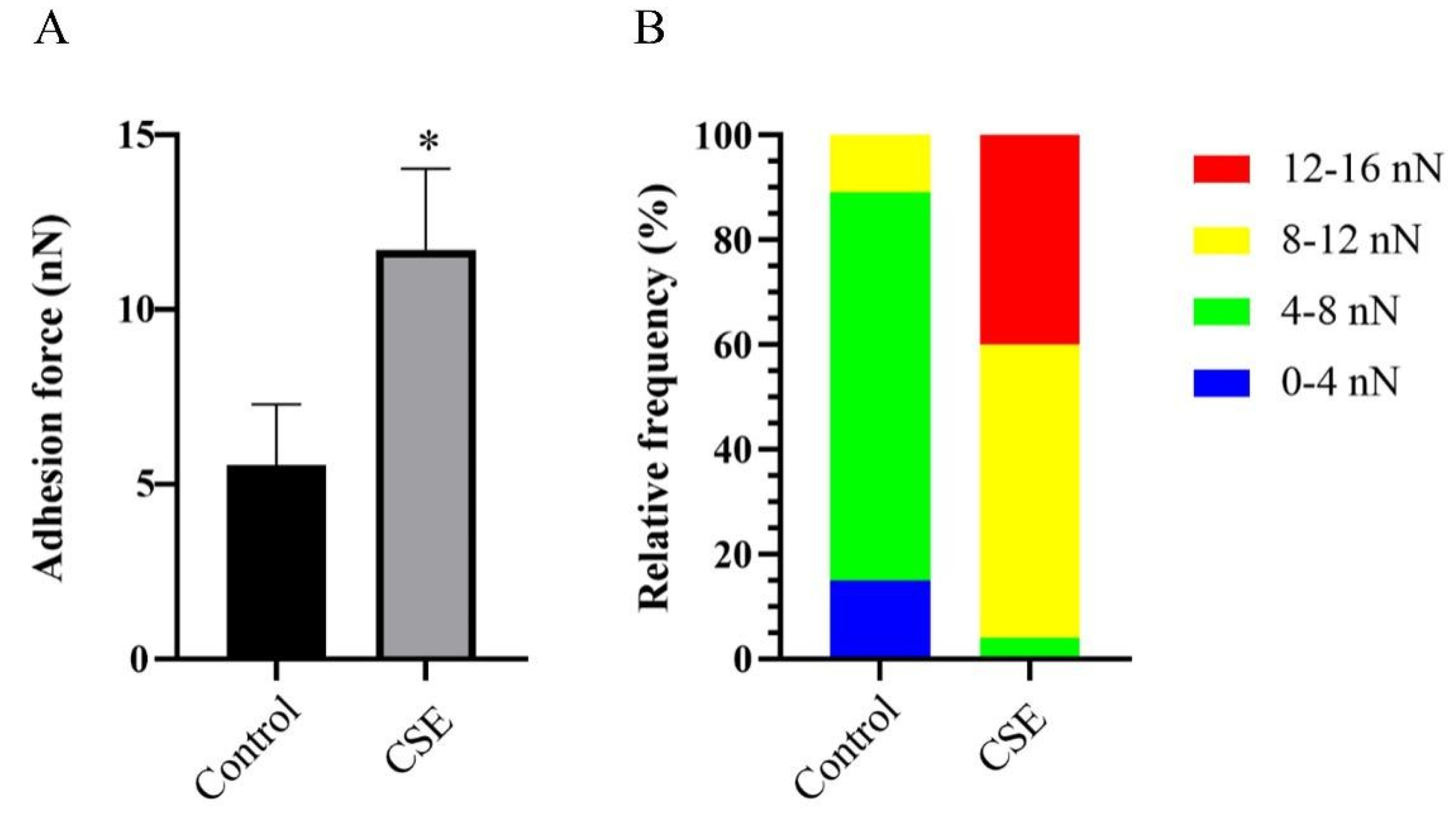
2.5. CSE Increased the Adhesion Force of C. albicans Biofilms
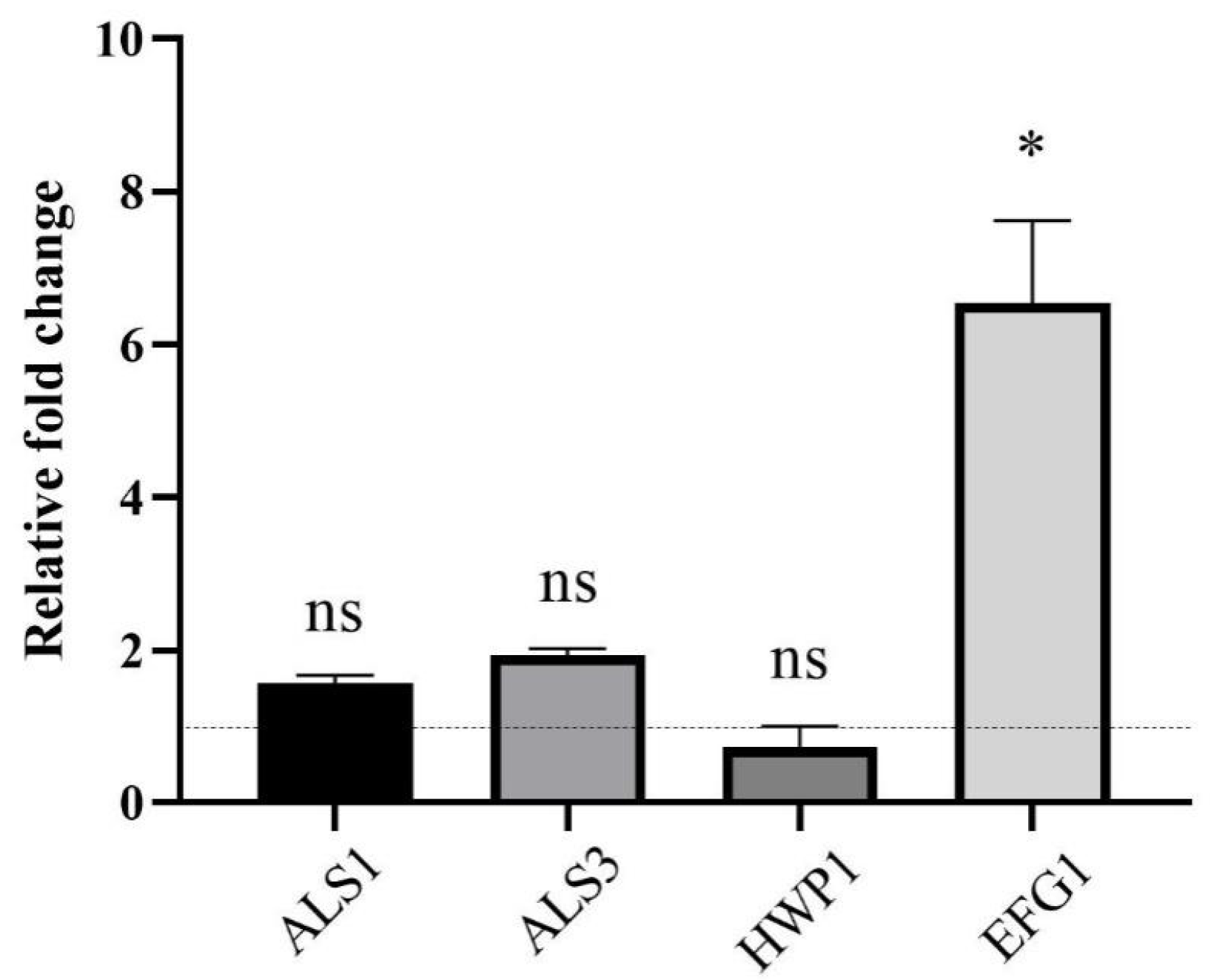
2.6. CSE Altered Gene Expression in C. albicans
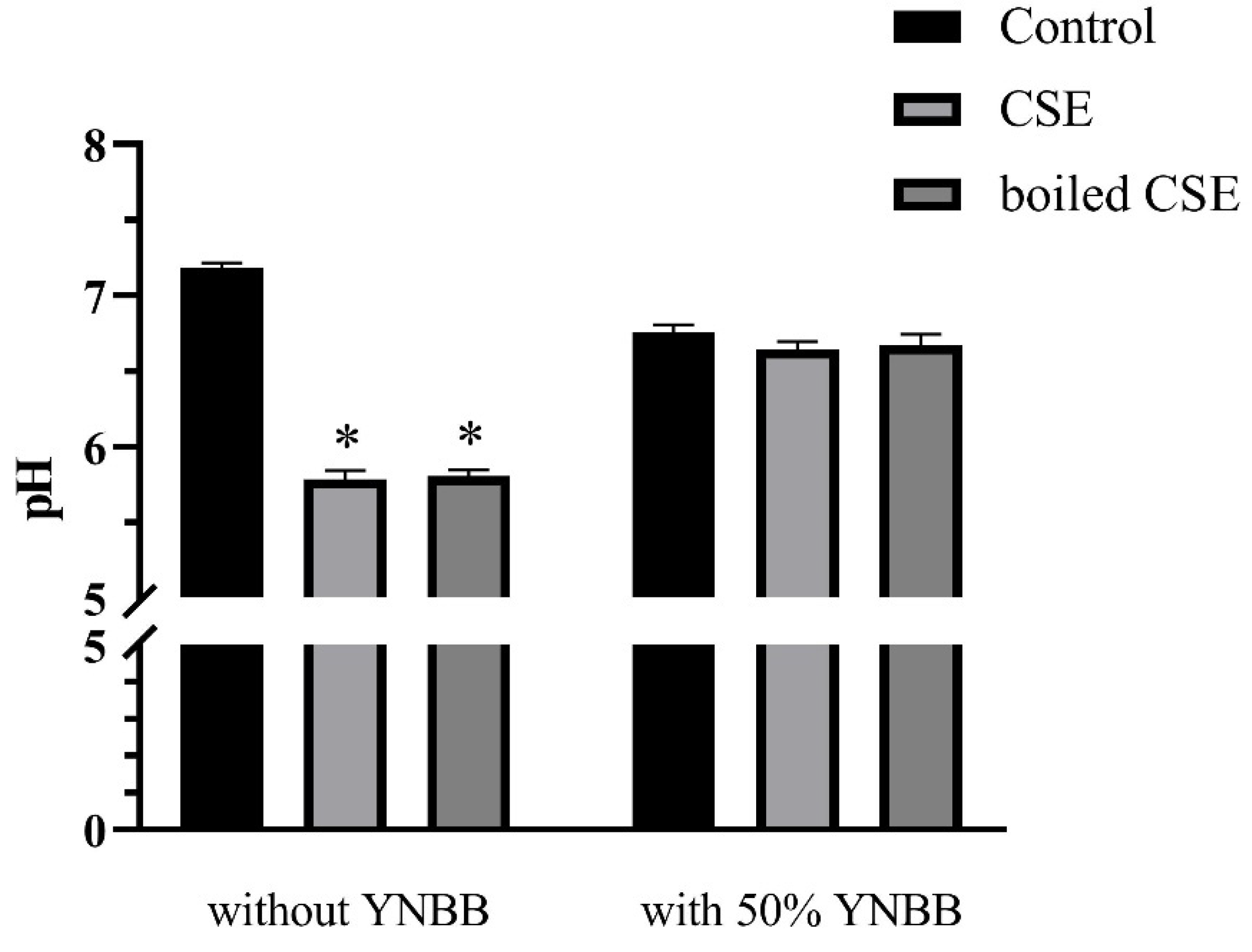
2.7. The pH Values of CSE and Boiled CSE
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Condition
4.2. Supernatant Collection and pH Measurement
4.3. Biofilm Formation and Biomass Analysis
4.4. CFU Counting
4.5. Scanning Electron Microscopy (SEM)
4.6. Confocal Laser Scanning Microscopy
4.7. Cellular Surface Hydrophobicity (CSH) Assay
4.8. Atomic Force Microscopy (AFM)
4.9. Real-Time Quantitative PCR (RT-qPCR)
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bouillaguet, S.; Manoil, D.; Girard, M.; Louis, J.; Gaia, N.; Leo, S.; Schrenzel, J.; Lazarevic, V. Root Microbiota in Primary and Secondary Apical Periodontitis. Front. Microbiol. 2018, 9, 2374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taneja, S.; Kumari, M. Use of triple antibiotic paste in the treatment of large periradicular lesions. J. Investig. Clin. Dent. 2012, 3, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Fouad, A.F.; Zerella, J.; Barry, J.; Spangberg, L.S. Molecular detection of Enterococcus species in root canals of therapy-resistant endodontic infections. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2005, 99, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, J.F.; Rocas, I.N. Polymerase chain reaction-based analysis of microorganisms associated with failed endodontic treatment. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2004, 97, 85–94. [Google Scholar] [CrossRef]
- Haapasalo, M.; Orstavik, D. Invitro infection and disinfection of dentinal tubules. J. Dent. Res. 1987, 66, 1375–1379. [Google Scholar] [CrossRef]
- Sedgley, C.M.; Lennan, S.L.; Appelbe, O.K. Survival of Enterococcus faecalis in root canals ex vivo. Int. Endod. J. 2005, 38, 735–742. [Google Scholar] [CrossRef] [Green Version]
- Mergoni, G.; Percudani, D.; Lodi, G.; Bertani, P.; Manfredi, M. Prevalence of Candida Species in Endodontic Infections: Systematic Review and Meta-analysis. J. Endodont. 2018, 44, 1616–1625. [Google Scholar] [CrossRef]
- Sharma, M.; Prasad, R. The quorum-sensing molecule farnesol is a modulator of drug efflux mediated by ABC multidrug transporters and synergizes with drugs in Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 4834–4843. [Google Scholar] [CrossRef] [Green Version]
- Ten Oever, J.; Netea, M.G. The bacteriome-mycobiome interaction and antifungal host defense. Eur. J. Immunol. 2014, 44, 3182–3191. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Wall, G.; Montelongo-Jauregui, D.; Vidal Bonifacio, B.; Lopez-Ribot, J.L.; Uppuluri, P. Candida albicans biofilm growth and dispersal: Contributions to pathogenesis. Curr. Opin. Microbiol. 2019, 52, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Bernard, C.; Girardot, M.; Imbert, C. Candida albicans interaction with Gram-positive bacteria within interkingdom biofilms. J. Mycol. Med. 2020, 30, 100909. [Google Scholar] [CrossRef] [PubMed]
- Kean, R.; Rajendran, R.; Haggarty, J.; Townsend, E.M.; Short, B.; Burgess, K.E.; Lang, S.; Millington, O.; Mackay, W.G.; Williams, C.; et al. Candida albicans Mycofilms Support Staphylococcus aureus Colonization and Enhances Miconazole Resistance in Dual-Species Interactions. Front. Microbiol. 2017, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falsetta, M.L.; Klein, M.I.; Colonne, P.M.; Scott-Anne, K.; Gregoire, S.; Pai, C.-H.; Gonzalez-Begne, M.; Watson, G.; Krysan, D.J.; Bowen, W.H.; et al. Symbiotic Relationship between Streptococcus mutans and Candida albicans Synergizes Virulence of Plaque Biofilms In Vivo. Infect. Immun. 2014, 82, 1968–1981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ricker, A.; Vickerman, M.; Dongari-Bagtzoglou, A. Streptococcus gordonii glucosyltransferase promotes biofilm interactions with Candida albicans. J. Oral Microbiol. 2014, 6, 23419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentine, M.; Benade, E.; Mouton, M.; Khan, W.; Botha, A. Binary interactions between the yeast Candida albicans and two gut-associated Bacteroides species. Microb. Pathog. 2019, 135, 103619. [Google Scholar] [CrossRef]
- Graham, C.E.; Cruz, M.R.; Garsin, D.A.; Lorenz, M.C. Enterococcus faecalis bacteriocin EntV inhibits hyphal morphogenesis, biofilm formation, and virulence of Candida albicans. Proc. Natl. Acad. Sci. USA 2017, 114, 4507–4512. [Google Scholar] [CrossRef] [Green Version]
- Krishnamoorthy, A.L.; Lemus, A.A.; Solomon, A.P.; Valm, A.M.; Neelakantan, P. Interactions between Candida albicans and Enterococcus faecalis in an Organotypic Oral Epithelial Model. Microorganisms 2020, 8, 1771. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Ellepola, A.N.B.; Samaranayake, L.P. The effect of limited exposure to antimycotics on the relative cell-surface hydrophobicity and the adhesion of oral Candida albicans to buccal epithelial cells. Arch. Oral Biol. 1998, 43, 879–887. [Google Scholar] [CrossRef]
- Sakko, M.; Tjaderhane, L.; Rautemaa-Richardson, R. Microbiology of Root Canal Infections. Prim. Dent. J. 2016, 5, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Ponde, N.O.; Lortal, L.; Ramage, G.; Naglik, J.R.; Richardson, J.P. Candida albicans biofilms and polymicrobial interactions. Crit. Rev. Microbiol. 2021, 47, 91–111. [Google Scholar] [CrossRef] [PubMed]
- Nogueira Vicosa, G.; Vieira Botelho, C.; Botta, C.; Bertolino, M.; Fernandes de Carvalho, A.; Nero, L.A.; Cocolin, L. Impact of co-cultivation with Enterococcus faecalis over growth, enterotoxin production and gene expression of Staphylococcus aureus in broth and fresh cheeses. Int. J. Food Microbiol. 2019, 308, 108291. [Google Scholar] [CrossRef] [PubMed]
- Vylkova, S.; Carman, A.J.; Danhof, H.A.; Collette, J.R.; Zhou, H.; Lorenz, M.C. The fungal pathogen Candida albicans autoinduces hyphal morphogenesis by raising extracellular pH. MBio 2011, 2, e00055-00011. [Google Scholar] [CrossRef] [Green Version]
- Konno, N.; Ishii, M.; Nagai, A.; Watanabe, T.; Ogasawara, A.; Mikami, T.; Matsumoto, T. Mechanism of Candida albicans transformation in response to changes of pH. Biol. Pharm. Bull. 2006, 29, 923–926. [Google Scholar] [CrossRef] [Green Version]
- Abbe, K.; Carlsson, J.; Takahashi-Abbe, S.; Yamada, T. Oxygen and the sugar metabolism in oral streptococci. Proc. Finn. Dent. Soc. Suom. Hammaslaak. Toim. 1991, 87, 477–487. [Google Scholar]
- Concha, M.L.; Castillo, A.; Liébana, J.; Gutiérrez, J.; Garcia-Mendoza, A. Initial pH as a determining factor of glucose consumption and lactic and acetic acid production in oral streptococci. Microbios 1996, 87, 207–216. [Google Scholar]
- Phan, Q.T.; Myers, C.L.; Fu, Y.; Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Ibrahim, A.S.; Edwards, J.E., Jr.; Filler, S.G. Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol. 2007, 5, 543–557. [Google Scholar] [CrossRef] [Green Version]
- Allonsius, C.N.; van den Broek, M.F.L.; De Boeck, I.; Kiekens, S.; Oerlemans, E.F.M.; Kiekens, F.; Foubert, K.; Vandenheuvel, D.; Cos, P.; Delputte, P.; et al. Interplay between Lactobacillus rhamnosus GG and Candida and the involvement of exopolysaccharides. Microb. Biotechnol. 2017, 10, 1753–1763. [Google Scholar] [CrossRef]
- Ma, S.; Ge, W.; Yan, Y.; Huang, X.; Ma, L.; Li, C.; Yu, S.; Chen, C. Effects of Streptococcus sanguinis Bacteriocin on Deformation, Adhesion Ability, and Young’s Modulus of Candida albicans. Biomed. Res. Int. 2017, 2017, 5291486. [Google Scholar] [CrossRef] [Green Version]
- Li, D.D.; Zhao, L.X.; Mylonakis, E.; Hu, G.H.; Zou, Y.; Huang, T.K.; Yan, L.; Wang, Y.; Jiang, Y.Y. In vitro and in vivo activities of pterostilbene against Candida albicans biofilms. Antimicrob. Agents Chemother. 2014, 58, 2344–2355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.; Shang, Z.; Li, T.; Yang, M.; Kong, L. In Vitro Antibiofilm Activity of Eucarobustol E against Candida albicans. Antimicrob. Agents Chemother. 2017, 61, e02707-16. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Hu, D.D.; Hu, G.H.; Su, J.; Bi, S.; Zhang, Z.E.; Wang, Z.; Zhang, R.L.; Xu, Z.; Jiang, Y.Y.; et al. Activity of Sanguinarine against Candida albicans Biofilms. Antimicrob. Agents Chemother. 2017, 61, e02259-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudbery, P.E. Growth of Candida albicans hyphae. Nat. Rev. Microbiol. 2011, 9, 737–748. [Google Scholar] [CrossRef]
- Araújo, D.; Henriques, M.; Silva, S. Portrait of Candida Species Biofilm Regulatory Network Genes. Trends Microbiol. 2017, 25, 62–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Qiu, W.; Zhang, K.; Zhou, X.; Ren, B.; He, J.; Xu, X.; Cheng, L.; Li, M. Nicotine Enhances Interspecies Relationship between Streptococcus mutans and Candida albicans. Biomed. Res. Int. 2017, 2017, 7953920. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Wei, Y.; Zhou, X.; Zhang, K.; Peng, X.; Ren, B.; Chen, V.; Cheng, L.; Li, M. Function of alanine racemase in the physiological activity and cariogenicity of Streptococcus mutans. Sci. Rep. 2018, 8, 5984. [Google Scholar] [CrossRef] [Green Version]
- Luo, W.; Huang, Y.; Zhou, X.; Han, Q.; Peng, X.; Ren, B.; Li, J.; Li, M.; Cheng, L. The effect of disaggregated nano-hydroxyapatite on oral biofilm in vitro. Dent. Mater. 2020, 36, e207–e216. [Google Scholar] [CrossRef]
- Sampaio, A.A.; Souza, S.E.; Ricomini-Filho, A.P.; Del Bel Cury, A.A.; Cavalcanti, Y.W.; Cury, J.A. Candida albicans Increases Dentine Demineralization Provoked by Streptococcus mutans Biofilm. Caries Res. 2019, 53, 322–331. [Google Scholar] [CrossRef]
- Gunasegar, S.; Himratul-Aznita, W.H. Nicotine enhances the thickness of biofilm and adherence of Candida albicans ATCC 14053 and Candida parapsilosis ATCC 22019. FEMS Yeast Res. 2019, 19. [Google Scholar] [CrossRef]
- Qiu, W.; Zheng, X.; Wei, Y.; Zhou, X.; Zhang, K.; Wang, S.; Cheng, L.; Li, Y.; Ren, B.; Xu, X.; et al. d-Alanine metabolism is essential for growth and biofilm formation of Streptococcus mutans. Mol. Oral Microbiol. 2016, 31, 435–444. [Google Scholar] [CrossRef]
- Li, M.Y.; Huang, R.J.; Zhou, X.D.; Gregory, R.L. Role of sortase in Streptococcus mutans under the effect of nicotine. Int. J. Oral Sci. 2013, 5, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Nikoomanesh, F.; Roudbarmohammadi, S.; Khoobi, M.; Haghighi, F.; Roudbary, M. Design and synthesis of mucoadhesive nanogel containing farnesol: Investigation of the effect on HWP1, SAP6 and Rim101 genes expression of Candida albicans in vitro. Artif. Cells Nanomed. Biotechnol. 2019, 47, 64–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tome, F.M.; Paula Ramos, L.; Freire, F.; Pereira, C.A.; de Oliveira, I.C.B.; Junqueira, J.C.; Jorge, A.O.C.; Oliveira, L.D. Influence of sucrose on growth and sensitivity of Candida albicans alone and in combination with Enterococcus faecalis and Streptococcus mutans to photodynamic therapy. Lasers Med. Sci. 2017, 32, 1237–1243. [Google Scholar] [CrossRef]
- Aguilar-Toala, J.E.; Astiazaran-Garcia, H.; Estrada-Montoya, M.C.; Garcia, H.S.; Vallejo-Cordoba, B.; Gonzalez-Cordova, A.F.; Hernandez-Mendoza, A. Modulatory Effect of the Intracellular Content of Lactobacillus casei CRL 431 Against the Aflatoxin B1-Induced Oxidative Stress in Rats. Probiotics Antimicrob. Proteins 2019, 11, 470–477. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ACT1 | GCGGTAGAGAGACTTGACCAACC | GACAATTTCTCTTTCAGCACTAGTAGTG |
| ALS1 | CCAAGTGTTCCAACAACTGAA | GAACCGGTTGTTGCTATGGT |
| ALS3 | CTAATGCTGCTACGTATAATT | CCTGAAATTGACATGTAGCA |
| HWP1 | TGGTGCTATTACTATTCCGG | CAATAATAGCAGCACCGAAG |
| EFG1 | TATGCCCCAGCAAACAACTG | TTGTTGTCCTGCTGTCTGTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Jing, Q.; Ren, B.; Cheng, L.; Zhou, X.; Lai, W.; He, J.; Li, M. Culture Supernatant of Enterococcus faecalis Promotes the Hyphal Morphogenesis and Biofilm Formation of Candida albicans. Pathogens 2022, 11, 1177. https://doi.org/10.3390/pathogens11101177
Jiang Q, Jing Q, Ren B, Cheng L, Zhou X, Lai W, He J, Li M. Culture Supernatant of Enterococcus faecalis Promotes the Hyphal Morphogenesis and Biofilm Formation of Candida albicans. Pathogens. 2022; 11(10):1177. https://doi.org/10.3390/pathogens11101177
Chicago/Turabian StyleJiang, Qingsong, Qi Jing, Biao Ren, Lei Cheng, Xuedong Zhou, Wenli Lai, Jinzhi He, and Mingyun Li. 2022. "Culture Supernatant of Enterococcus faecalis Promotes the Hyphal Morphogenesis and Biofilm Formation of Candida albicans" Pathogens 11, no. 10: 1177. https://doi.org/10.3390/pathogens11101177