
The p38 MAPK/PMK-1 Pathway Is Required for Resistance to Nocardia farcinica Infection in Caenorhabditis elegance

1
State Key Laboratory for Conservation and Utilization of Bio-Resource in Yunnan, Yunnan University, Kunming 650091, China
2
School of Basic Medical Sciences, Guizhou Medical University, Guiyang 550025, China
3
Yunnan Provincial Key Laboratory of Molecular Biology for Sinomedicine, Yunnan University of Chinese Medicine, Kunming 650500, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pathogens 2022, 11(10), 1071; https://doi.org/10.3390/pathogens11101071
Submission received: 24 April 2022
/
Revised: 8 September 2022
/
Accepted: 9 September 2022
/
Published: 21 September 2022
(This article belongs to the Special Issue Microbe-Nematode Interactions)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Nocardia farcinica is an opportunistic pathogen that causes nocardiosis primarily in patients with compromised immune systems. In this study, we used the genetically tractable organism Caenorhabditis elegans as a model to study the innate immune responses to N. farcinica infection. We found that unlike other pathogenic bacteria such as Pseudomonas aeruginosa and Staphylococcus aureus, N. farcinica failed to kill adult worms. In another words, adult worms exposed to N. farcinica exhibited a normal lifespan, compared with those fed the standard laboratory food bacterium Escherichia coli OP50. Interestingly, deletion of three core genes (pmk-1, nsy-1 and sek-1) in the p38 MAPK/PMK-1 pathway reduced the survival of worm exposure to N. farcinica, highlighting a crucial role of this pathway for C. elegans in resistance to N. farcinica. Furthermore, our results revealed that N. farcinica exposure up-regulated the level of PMK-1 phosphorylation. The activation of PMK-1 promoted nuclear translocation of a transcription factor SKN-1/Nrf2, which in turn mediated N. farcinica infection resistance in C. elegans. Our results provide an excellent example that the integrity of immune system is key aspect for counteract with pathogenesis of N. farcinica.
1. Introduction
Nocardia farcinica, is an opportunistic pathogen under the genus of Actinomycetes widely distributed in the soil and rotten substances, causing lung infection through inhalation of bacterial particles [1]. Since the first case of pulmonary nocardiosis was reported in the USA in 1898, thousands of cases of pulmonary nocardiosis have been reported annually, primarily in patients with compromised immune systems [2]. Nocardia not only cause lung infections, but also erode other organs by entering blood vessels, causing localized or systematical infection in central nervous system, skin, subcutaneous tissue, retina, kidney, joint, bone, and heart [1]. Nocardia are also associated with Parkinson’s syndrome, as subcutaneous injection of Nocardia induce symptoms similar to Parkinson’s syndrome clinically and pathologically in mice [3]. A recent study reveals that Nocobactin NA-a and Nocobactin NA-b, two compounds isolated from N. farcinica, suppress immune responses by inhibiting Notch signal pathway in RAW264.7 macrophages [4]. Despite several studies have revealed the virulence of N. farcinica in mammal. The mechanism of pathogenesis in N. farcinica infection is not fully understood.
Caenorhabditis elegans is a free-living, bacterivorous nematode that lives in the soil, well known as invertebrate without adaptive immune system as well as specific immune cells [5]. In wild worms, a large microbial fauna is observed in the gut [6]. In the natural environment, C. elegans often encounters a variety of pathogens, thus it exerts a strong selective pressure to evolve and develop an effective innate immune system. C. elegans therefore represents a powerful invertebrate model to study the innate immune responses to nosocomial bacterial pathogens such as Pseudomonas aeruginosa, Staphylococcus aureus, Salmonella enterica, Salmonella typhimurium, and Serratia marcescens [7,8,9,10], and fungal species such as Cryptococcus neoformans [11] and Candida albicans [12].
During the last two decade, studies using C. elegans have revealed a variety of the evolutionarily conserved signaling pathways in the innate immunity, such as the DAF-2 insulin-like signaling pathway [13], the p38 mitogen-activated protein kinase (MAPK) pathway [14], and the ERK MAPK pathway [15], the JNK MAPK pathway [16], the protein kinase D pathway [17], the G protein coupled receptor FSHR-1 pathway [18], the G protein EGL-30 pathway [19], and the transcription factor TFEB pathway [20]. Of these signaling pathways, the p38 MAPK/PMK-1 pathway, which consists of a core cassette of NSY-1/MAPKKK, SEK-1/MAPKK, and PMK-1/MAPK, act in the intestine of C. elegans to regulate innate immunity [14,21]. PMK-1 mediates the expression of a set of immune-related genes, such as C-type lectins, ShK toxins, lysozymes, and antimicrobial peptides, which are involved in fighting off infection [22]. Meanwhile, a couple of the downstream transcription factors of PMK-1 that promote pathogen resistance have been identified. For instance, PMK-1 activates the IRE-1–XBP-1-dependent unfolded protein response (UPR) in C. elegans upon P. aeruginosa infection [23]. Interestingly, the role for XBP-1-mediated UPR is to protect worms against the lethal endoplasmic reticulum stress during activation of innate immunity. Furthermore, the transcription factor SKN-1 is activated by PMK-1, thereby conferring resistance P. aeruginosa infection [24]. Under normal conditions, the transcription factor ATF-7 inhibits PMK-1 regulated immune-related gene expression. However, the activation of PMK-1 by bacterial infection can phosphorylates ATF-7, which switches the transcription factor from a repressor to an activator of PMK-1–dependent immune gene expression [25].
In this study, we screened the major signaling pathways that are involved in defense against N. farcinica in C. elegans. We discovered that the PMK-1/p38 MAPK signaling was required for the survival of worms upon N. farcinica exposure. Further studies indicated that PMK-1/p38 MAPK activated the transcription factor SKN-1, which in turn conferred resistance to N. farcinica infection in C. elegans.
2. Results
2.1. The Lifespan of C. elegans Is Unaffected when Fed on N. farcinica
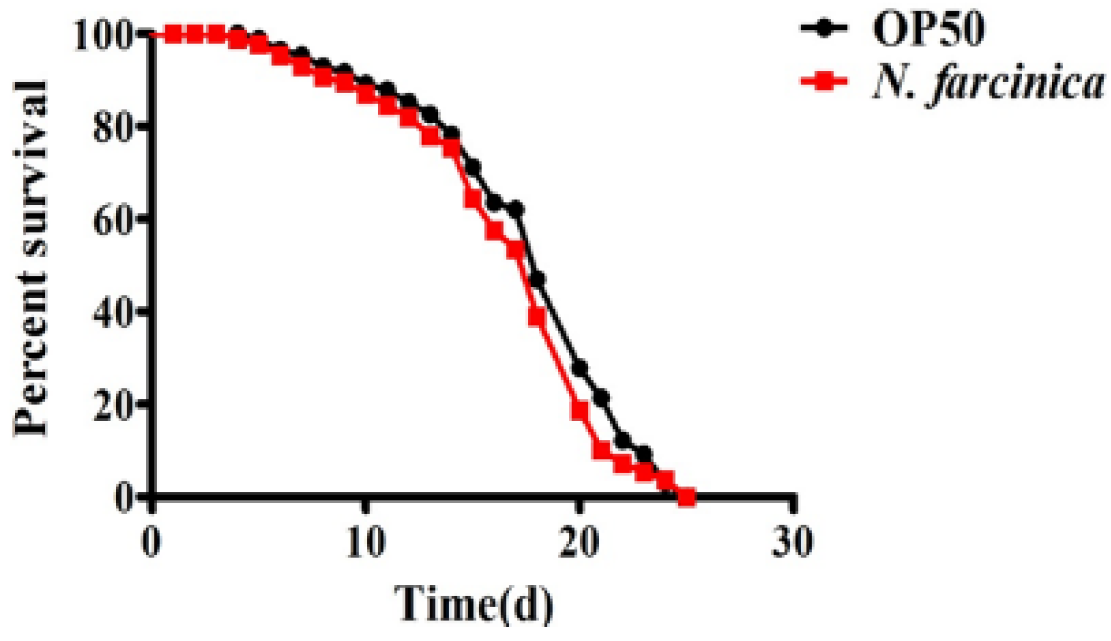
We first investigated the pathogenicity of N. farcinica in nematodes. However, we found that unlike worms exposed to P. aeruginosa, S. aureus, and E. faecium [26,27], wild-type (WT) worms exposed to N. farcinica exhibited a similar lifespan as those fed E. coli OP50, the standard laboratory food (Figure 1). Thus, C. elegans feeding on N. farcinica exhibits a normal lifespan.
2.2. The p38 MAPK Pathway Is Critical for C. elegans to Resist N. farcinica
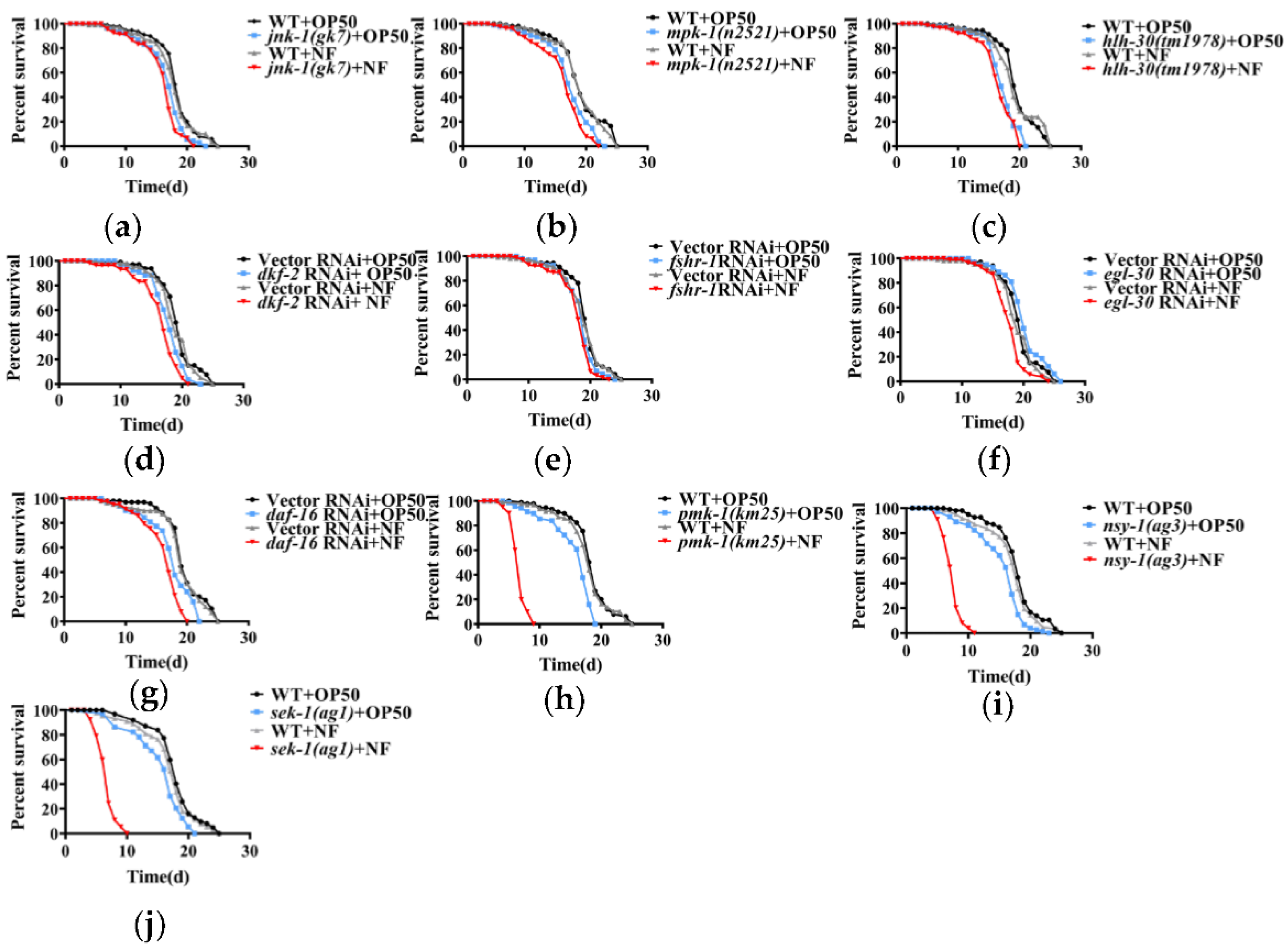
Although C. elegans does not have a canonical adaptive immune system, it has evolved an innate immune mechanism to counteract bacterial infection. However, as a bacterial pathogen, N. farcinica had no effect on lifespan in worms. The fact that N. farcinica mainly infects immunoincompetent patients prompted a hypothesis that C. elegans relies on its innate immune defenses against N. farcinica infection. To verify this hypothesis, we screened the major innate immune pathways in worms, such as ERK MAPK/MPK-1 [28], JNK MAPK/JNK-1 [16], p38 MAPK/PMK-1 [14], DAF-2/DAF-16 [13], G protein coupled receptor FSHR-1 [18], protein kinase PKD/DKF-2 [17], transcription factor TFEB/HLH-30 [20], and G protein EGL-30 [19]. We found that mpk-1, jnk-1, hlh-30 mutant strains which fed on either E. coli OP50 or N. farcinica had comparable lifespans (Figure 2a–c). Meanwhile, RNAi knockdown of fshr-1, dkf-2, egl-30 or daf-16 failed to affect lifespans when exposed to either E. coli OP50 or N. farcinica (Figure 2d–g). However, we observed slight lifespan shortness in mutant strains and RNAi knockdown worms compared to WT strains or subjected to empty vector RNAi, respectively (p ≤ 0.01). In contrast, pmk-1(km25) mutants exhibited marked decrease in lifespan when exposed to N. farcinica, compared to worms fed on E. coli OP50 and WT (Figure 2h). NSY-1/MAPKKK and SEK-1/MAPKK are kinases located upstream of PMK-1/p38 MAPK, which are involved in its activation through phosphorylation [14]. In the current study, the nsy-1(ag3) and sek-1(ag1) mutants exposed to N. farcinica also showed a significant reduction in lifespan, compared to E. coli OP50 diet and WT (Figure 2i,j). These results suggested that the p38 MAPK pathway is essential for survival of worms exposed to N. farcinica.
2.3. C. elegans Depended on PMK-1 Activation to Resist N. farcinica Infection
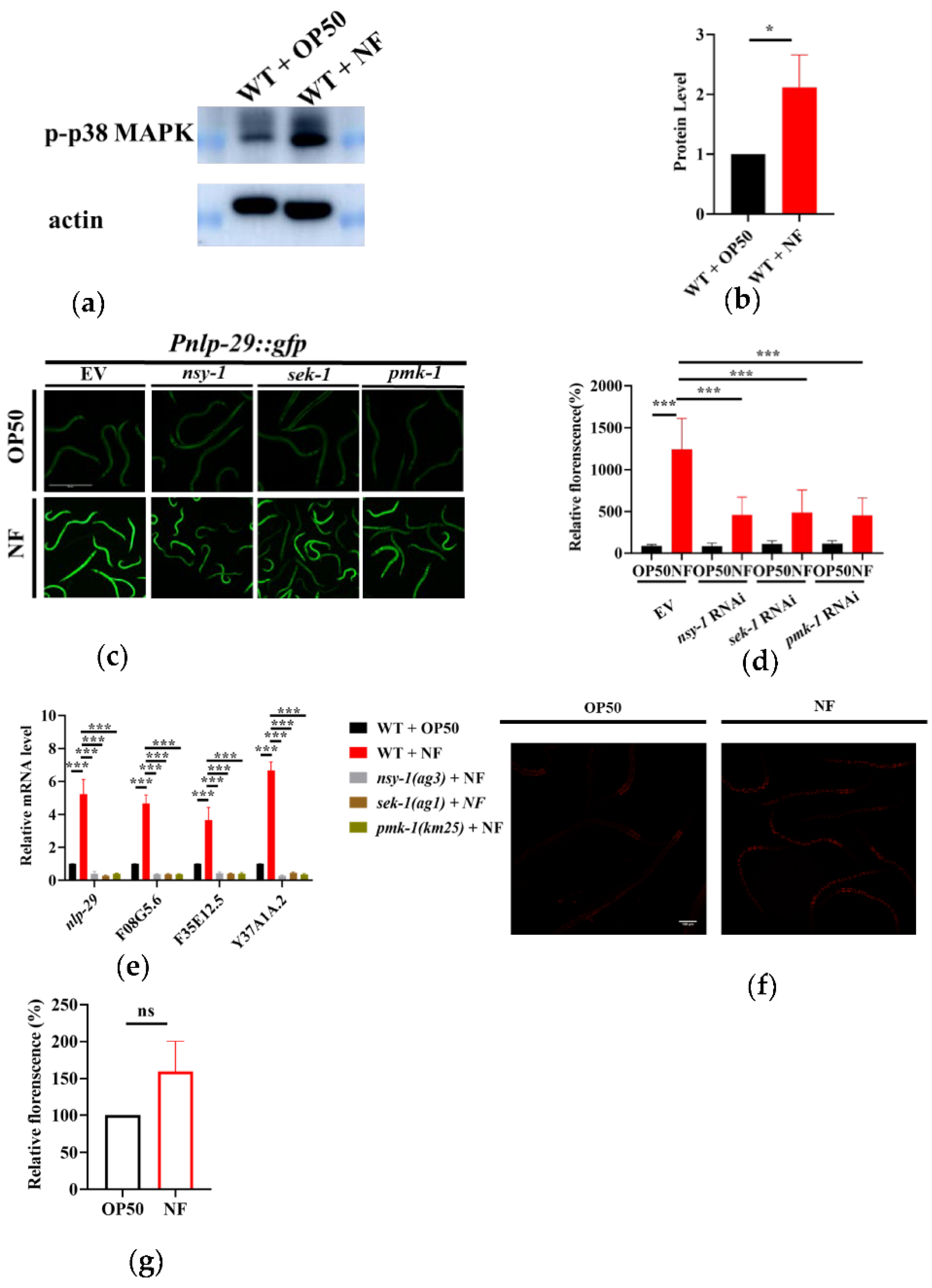
We then asked if N. farcinica activated PMK-1 by determining the phosphorylation level of PMK-1. We found that the phosphorylation levels of PMK-1 were significantly increased in worms exposed to N. farcinica for 24 hours, compared to those fed E. coli OP50 (Figure 3a,b). The increase in phosphorylation level of PMK-1 has also been reported when C. elegans was challenged by pathogens including Pseudomonas aeruginosa PA14 [29] and Cutibacterium acnes [30]. A previous study has demonstrated that the up-regulation of antimicrobial peptide gene nlp-29 is mediated by PMK-1 [31]. We thus asked if PMK-1 activation upon exposure to N. farcinica can up-regulate the nlp-29 expression (Figure 3c,d). As F08G5.6, F35E12.5 and Y37A1A.2 have been reported as PMK-1-regulated genes [22], we then verified the mRNA levels of these genes after N. farcinica exposure. The expressions of F08G5.6, F35E12.5 and Y37A1A.2 were significantly up-regulated after exposure to N. farcinica. In contrast, mutations in nsy-1(ag3), sek-1(ag1), and pmk-1(km25) suppressed their mRNA levels (Figure 3e). Taken together, these results suggested that N. farcinica exposure activates the p38 MAPK pathway in worms. In previous study, several gens like alkylhydroperoxide reductase (ahpC), catalase (katG), and superoxide dismutase (sodF) are predicted with high expressivity in N. farcinica, which could defense against reactive oxygen species (ROS) [32]. We thus checked the ROS level in worms through DHE stain. We found that the level of ROS was only increased slightly after N. farcinica infection. Thus, ROS are unlikely to be involved in the activation of the p38 MAPK pathway in worms.
2.4. N. farcinica Exposure Activates the Transcription Factor SKN-1 in a p38 MAPK Dependent Manner
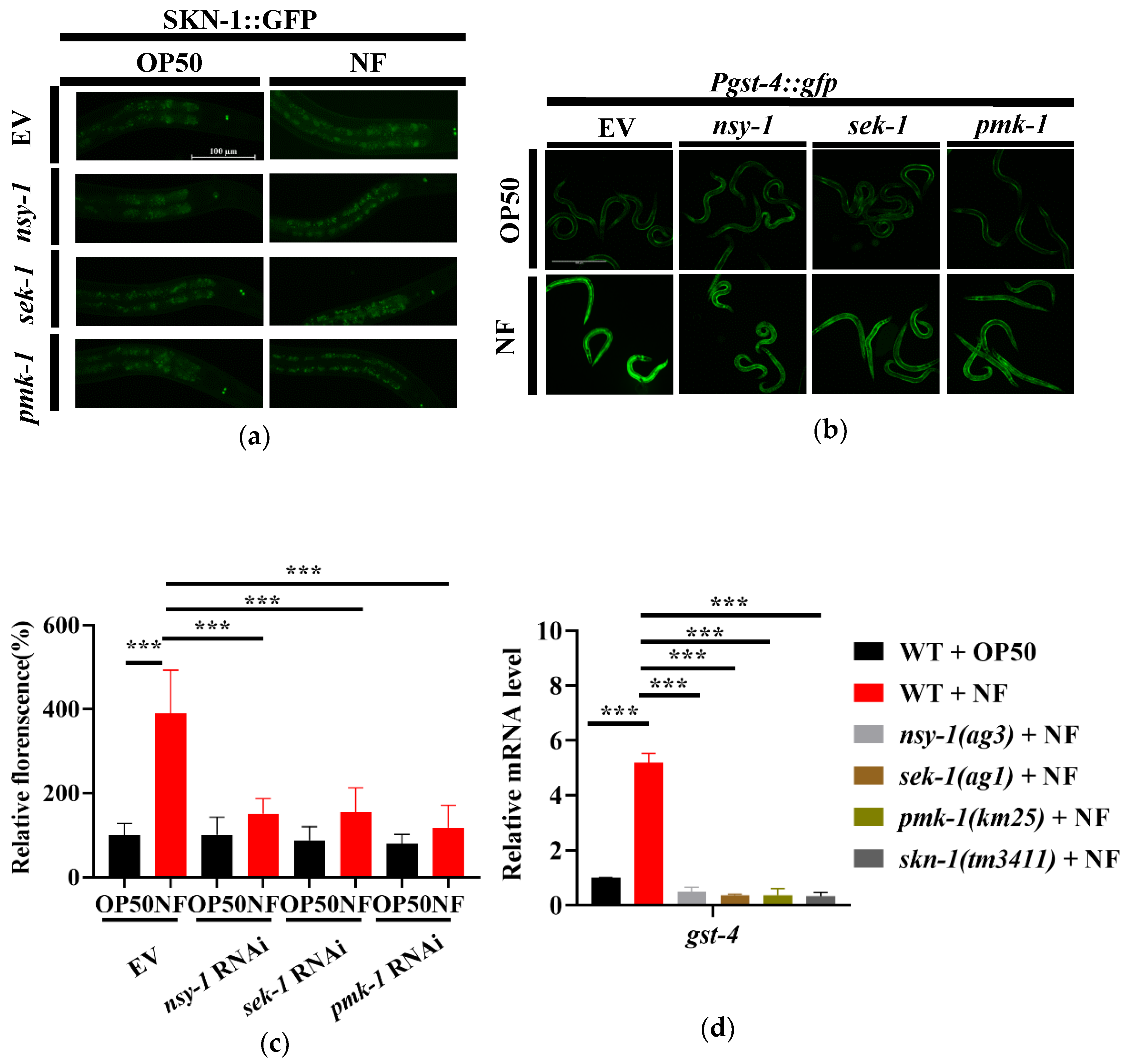
The transcription factor SKN-1/Nrf2 is a key downstream molecule of the p38 MAPK pathway [33]. PMK-1 has been reported to activate SKN-1 under both oxidative stress [34] and pathogenic infection conditions [24]. In C. elegans, SKN-1 confers resistance to Gram-negative bacteria P. aeruginosa PA14 and Gram-positive bacteria E. faecalis [33]. To determine whether SKN-1 was activated after N. farcinica infection, we used transgenic worms expressing SKN-1b/c::GFP+rol-6 to detect the nuclear translocation of SKN-1, which is an indicator of its activation [35]. Under standard growth conditions, SKN-1 was mainly located in the cytoplasm in all tissues (Figure 4a). By contrast, the SKN-1 nuclear translocation was observed in the nucleus of intestine upon 24 hours of N. farcinica exposure. However, knockdown of nsy-1, sek-1 or pmk-1 by RNAi significantly reduced the nuclear translocation of SKN-1 in worms exposed to N. farcinica (Figure 4a). The second phase detoxification gene gst-4 (encoding Glutathione S-transferase-4) is a target gene of SKN-1 [36,37]. We thus used transgenic worms expressing the Pgst-4::gfp reporter gene to further demonstrate the activation of SKN-1 by N. farcinica infection. We found that the expression level of Pgst-4::gfp was dramatically increased in worms exposed to N. farcinica for 24 hours, The increase is abolished when worms subjected to skn-1, nsy-1, sek-1 or pmk-1 RNAi (Figure 4b,c). qPCR analysis showed that mutations in skn-1(tm3411), nsy-1(ag3), sek-1(ag1), and pmk-1(km25) suppressed up-regulation of gst-4 expression induced by N. farcinica infection (Figure 4d). Taken together, our study indicated that N. farcinica infection activates SKN-1via the p38 MAPK pathway.
2.5. SKN-1 Confers Resistance to N. farcinica Infection
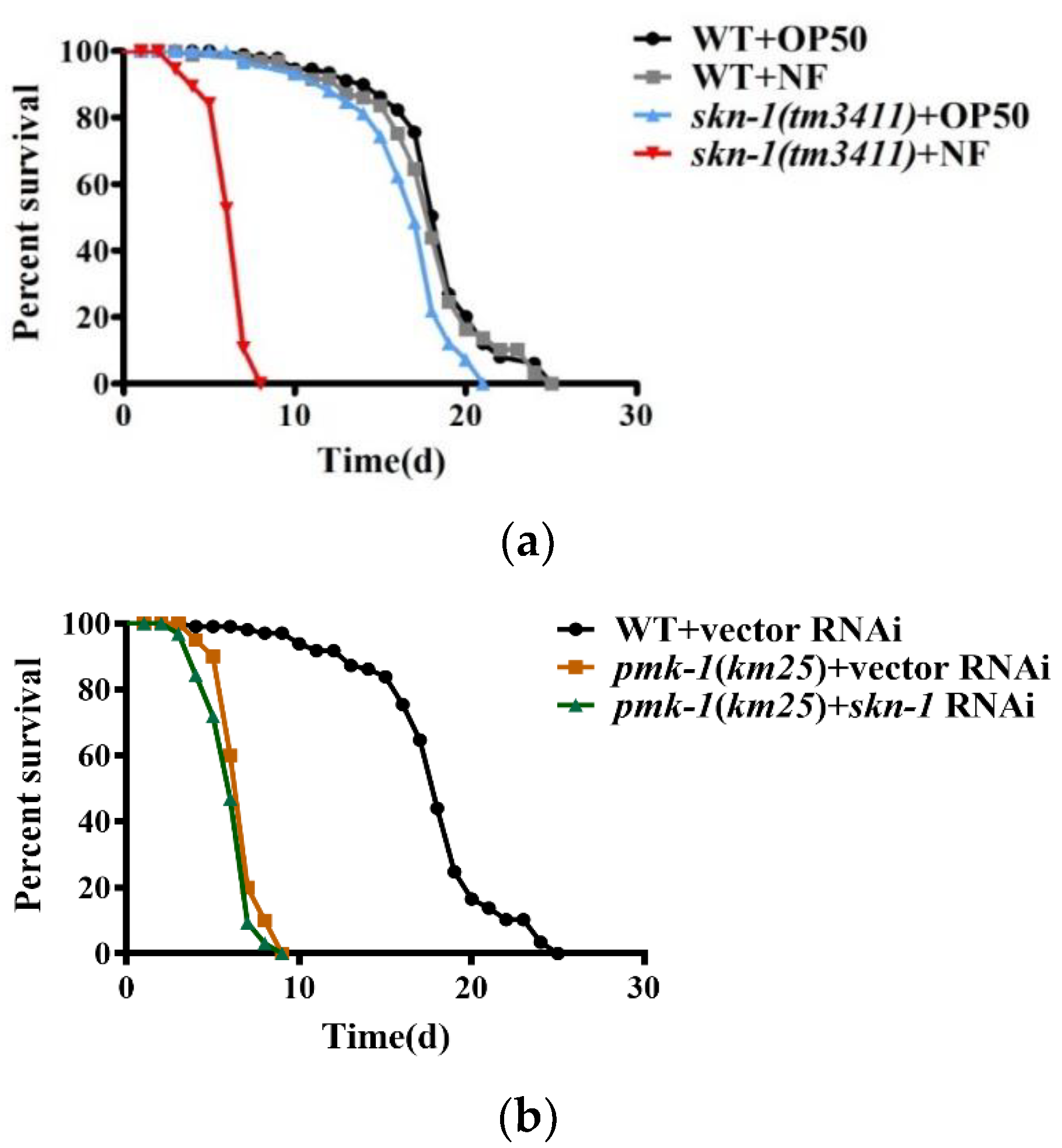
skn-1 RNAi, indicating that skn-1 is required to resist this pathogen. However, the survival rate of the pmk-1(km25) mutants was comparable to that of the pmk-1(km25) mutant subjected to skn-1 RNAi upon N. farcinica infection (Figure 5b), indicating that skn-1 is epistatic to pmk-1. Taken together, these results suggest that the p38 MAPK pathway mediates resistance to N. farcinica infection by activating SKN-1.
2.6. PMK-1 and SKN-1 Are Critical in Elimination of N. farcinica
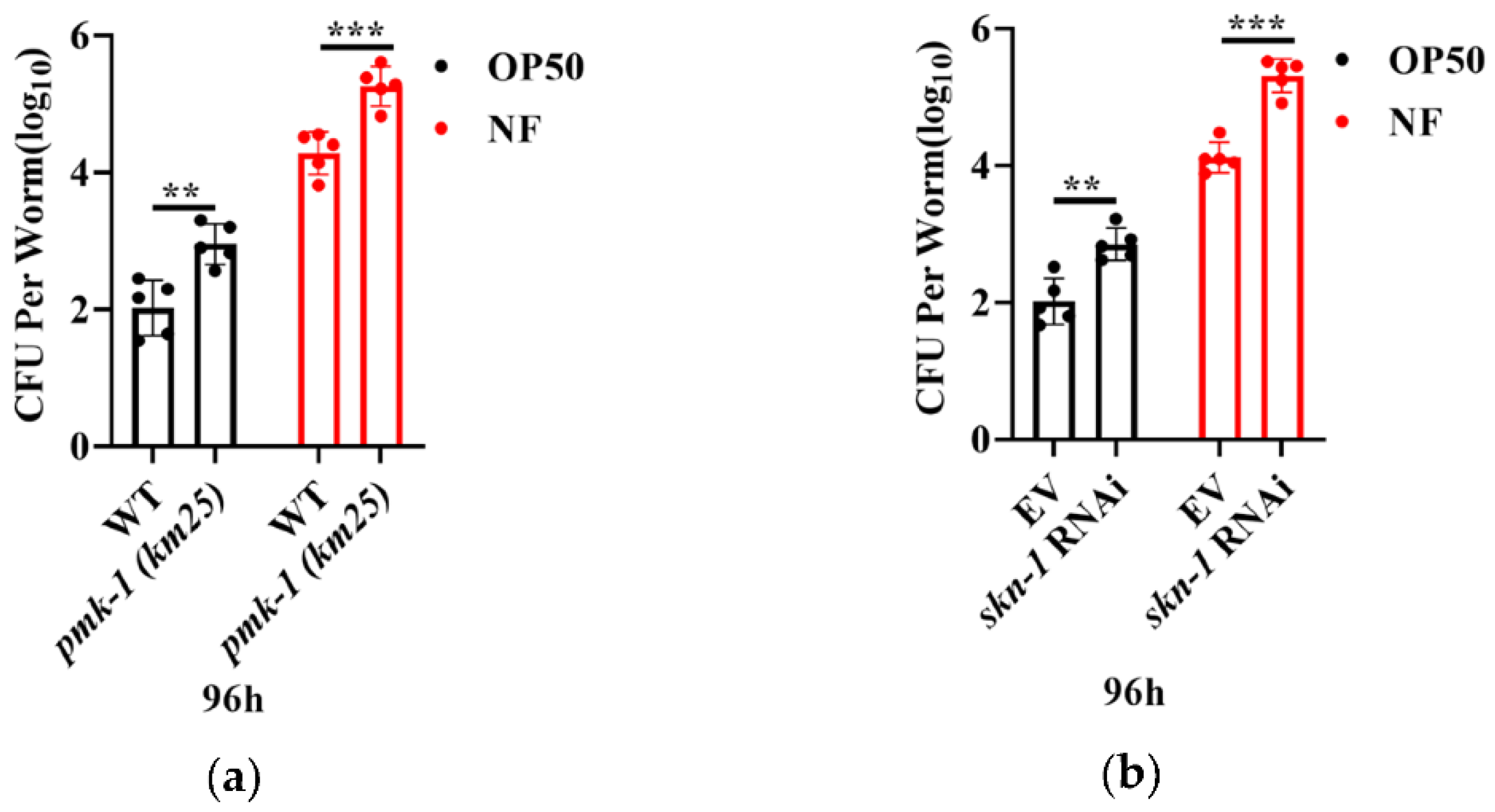
The successful colonization of bacteria is known as a key step in conducing infection. To ask if N. farcinica colonized in worms, young adult worms were exposed with either E. coli OP50 or N. farcinica on BHI plates for 96 hours. We found that RNAi knockdown of pmk-1 and skn-1 significantly increased the colony forming units (CFU) of N. farcinica and E. coli OP50 in the body of worms (Figure 6a,b). These results suggest that PMK-1 and SKN-1 are required for limitation of bacterial accumulation in the intestine of worms.
2.7. PMK-1 and SKN-1 Protects C. elegans from Tissue Damage upon N. farcinica Infection
To determine whether N. farcinica induced tissue damage after long time infection in the worms, we observed the DIC images of pmk-1 (km25) worms, skn-1 (RNAi) worms, and control group worms. As black arrows pointing out, genetic inactivation of pmk-1 and skn-1 caused necrosis in the head of worms exposed to N. farcinica, but not to E. coli OP50 (Figure 7a,b). These results suggest that PMK-1 and SKN-1 plays a protective role in tissue damages resulting from N. farcinica infection.
3. Discussion
Our study demonstrated that the PMK-1/p38 MAPK pathway confers resistance to N. farcinica infection in C. elegans. This pathway is identified by screening eight signaling pathways required for innate immune responses to bacterial infections. Furthermore, our results indicated that N. farcinica exposure mediated the phosphorylation of p38 MAPK increased, which in turn activates SKN-1/Nrf2. The transcription factor is involved in C. elegans defense against N. farcinica.
In C. elegans, the PMK-1/p38 MAPK pathway plays a central role in innate immune response to infection with pathogenic bacteria, such as P. aeruginosa [14,38] S. aureus [39], Salmonella [7], Yersinia pestis [40], and Mycobacteria [41]. The p38 MAPK pathway mediates the resistance to bacterial infection and bacterial toxins, such as the toxin Cry5B [16] secreted by Bacillus thuringiensis and Shiga-like toxin of enterohemorrhagic E. coli O157:H7 [42]. In mammals, the p38 MAPK pathway is also involved in defense against pathogens [43,44], suggesting that this pathway may represent an ancient feature of metazoan innate immunity. Interestingly, N. farcinica can activate dendritic cells via the p38 MAPK pathway, leading to release of interleukin-12 (IL-12/23p40), known as key immune factor, mediates resistance to Mycobacterium. spp [45,46]. These results implicate that p38 MAPK is a conserved pathway mediates host resistance to N. farcinica in organisms ranging from C. elegans to mammals.
As a transcription factor, SKN-1 is involved in resistance to oxidative stress by upregulating a set of the Phase II detoxification genes [37]. SKN-1 also confers resistance to infections with pathogenic bacteria, such as P. aeruginosa, E. faecalis [24], Mycobacterium [41], and Aeromonas dhakensis [47]. In the current study, SKN-1/Nrf2 is required for innate immune responses to N. farcinica infection. It has been reported that PMK-1 can phosphorylate SKN-1 at Ser-74 and Ser-340 [34]. The phosphorylation of SKN-1 promotes its nuclear translocation and activation. Like bacterial pathogens including P. aeruginosa, E. faecalis and N. farcinica infection activates SKN-1 in a p38 MAPK-dependent manner. These results suggest that SKN-1 is one of downstream effectors of p38 MAPK that mediates C. elegans innate immune responses to both Gram-negative and Gram-positive pathogens. The molecular mechanism underlying SKN-1-mediated innate immunity remains unclear. Previously, we and others have demonstrated that autophagy plays an important role in C. elegans defense against a variety of pathogenic bacteria by repairing organismal insults [15,20,48]. SKN-1 up-regulates a set of autophagic genes in the germline-deficient glp-1(e2141ts) mutants [49]. Meanwhile, SKN-1 is involved in activation of autophagy in long-lived hyl-1; lagr-1 mutants [50]. Thus, SKN-1 is likely to limit the damage induced by N. farcinica infection via promoting autophagy. The opportunistic infections followed immunologic deficiency have mainly been seen in HIV infected patients, revealing that the environmental “harmless” microorganisms could be life threaten, such as Penicillium marneffei, known as the third common opportunistic pathogen that causes penicilliosis in HIV patients. Likewise, N. farcinica is an opportunistic pathogen, with the majority of infections occurring in immunocompromised patients. In the current study, WT worms do not have a reduced lifespan when exposed to N. farcinica. These data suggested that like humans, WT worms with an intact innate immune system can resist to N. farcinica infection, but lacking key genes in the PMK-1/p38 MAPK pathway become susceptible to N. farcinica infection. In our studies, we also noticed that lack of p38 MAPK pathway and SKN-1 leads to significant increases accumulation of N. farcinica and obvious enlarged vacuoles in the head of worms. Thus, our finding emphasizes that the integrity of immune system is crucial for defense against N. farcinica infection in C. elegans.
What are the critical signal molecules from N. farcinica to activate the p38 MAPK/PMK-1 pathway?However, our data demonstrate that ROS are not the signal component produced by N. farcinica. Several virulence factors of N. farcinica have been reported when the pathogen infected mammals. For instance, Nfa34810 protein of N. farcinica can stimulate macrophages to produce tumor necrosis factor alpha (TNF-α) and other immune related factors through the TLR4 pathway [51]. Besides, N. farcinica IFM 11523, which isolated from Japanese patient, secretes two pathogenesis factors nocobactin NA-a (compound 1) and nocobactin NA-b (compound 2), which act as on the notch signaling inhibitors [4]. Thus, the signal molecules that activate the p38 MAPK/PMK-1 pathway need be investigated further in light of our present study.
4. Materials and Methods
4.1. C. elegans and Bacteria Strains
The C. elegans strains were maintained on NGM medium containing E. coli OP50 under standard condition. Strain used in this study include C. elegans Bristol N2, pmk-1(km25), mpk-1(n2521), let-60(ga89), jnk-1(gk7), hlh-30(tm1978), skn-1(tm3411), nsy-1(ag3), sek-1(ag1), skn-1b/c::gfp+rol-6(su1006), nlp-29p::gfp+col-12p::DsRed, gst-4p::gfp::NLS(pAF15) were kindly provided by the Caenorhabditis Genetics Center (CGC), which is funded by NIH Office of Research Infrastructure Programs (P40 OD010440). Bristol N2 strain of C. elegans is used as non-genetic modified control in experiments, and labeled as wild-type (WT) in our study [52,53].
4.2. RNA Interference
Bacterial strains contain RNAi targeting genes were obtained from Ahringer library [54]. E. coli HT115 (DE3) strain carrying vector L4440 express dsRNA corresponding to targeted genes to perform double-stranded (ds)RNA-mediated RNA interference (RNAi). As experimental control, E. coli HT115 carrying empty vector (no dsRNA expression) were used, labelled as EV. All strains were grown overnight in LB medium with 100 μg/mL ampicillin at 37 °C. Cultured E. coli strains spread onto NGM plates contained 100 μg/mL ampicillin and 1 mM isopropyl 1-thio-β-D-galactopyranoside (IPTG). RNAi assay were performed according to pervious study [55], in brief, synchronized L1 larvae (Bristol N2 strain of C. elegans) fed on RNAi bacteria strains at 20 °C until reach to young adult. Then young adult worms were transferred on the BHI plates for further studies.
4.3. C. elegans Survival Assays
The C. elegans strains were cultured on NGM plate with E. coli OP50 at 20 °C following the standard maintenance procedures [52]. Worms were synchronized by alkaline hypochlorite solution [56], eggs were incubated in M9 buffer at 20 °C, overnight for hatching. Hatched L1 larvae were transferred onto NGM plates seeded with live E. coli OP50 for further assays. To examine the effect of N. farcinica on worms’ lifespan, synchronized L1 larvae were cultivated fed E. coli OP50 at 20 °C until the young adult stage. Then 50–60 worms were fed with either N. farcinica or laboratory standard diet E. coli OP50 on BHI medium containing 5′-fluoro- 2′-deoxyuridine (FUdR) (75 μg/mL) at 25 °C. The survival curves were plotted by scoring the dead worm at 24 h interval. Immobile worms unresponsive to touch were scored as dead. Three plates were analyzed per assay and all experiments were performed three times.
4.4. SKN-1 Nuclear Localization Assays
After young adult worms expressing skn-1b/c::gfp were fed with either N. farcinica or laboratory standard diet E. coli OP50 on BHI plates for 24 hours. Briefly, worms were mounted on slides from plates and imaged by using Nikon e800 fluorescence microscope. At least 30 worms were examined under each condition in three independent experiments.
4.5. Western Blot
After washed with M9 buffer, worms were homogenized in liquid nitrogen. Then the homogenate was lysed on ice for 30 min in lysis buffer RIPA (Beyotime Institute of Biotechnology, Haimen, China). After centrifuged at 12,000 rpm for 15 min at 4 °C, the supernatant was obtained and used for Western blot analysis. The total protein extraction was loaded on 10% SDS-PAGE for electrophoresis. Proteins were then transferred to immobilon-PSQ transfer PVDF membrane (Millipore, Bedford, MA, USA). Primary antibodies were anti-phospho-p38 antibodies (1:1000 dilution; Promega Biotech Co.,Ltd, Beijing, China), and anti-α-tubulin antibodies (1:1000 dilution; Abcam, Cambridge, UK). The secondary antibodies were peroxidase-coupled anti-rabbit IgG (1:10,000 dilution; Abmart, Shanghai, China). Blots were developed using Super Signal chemiluminescence substrate (Thermo Fisher Scientific, Waltham, MA). An imaging system (Amersham Imager 600) was used for documentation of the Western blot results. Band intensities were measured using ImageJ software (NIH).
4.6. Quantitative PCR Analysis
Total RNA was isolated from worms with TRIzol Reagent (Invitrogen, Carlsbad, CA). Random-primed cDNAs were generated by reverse transcription of the total RNA samples with SuperScript II (Invitrogen). A real time-PCR analysis was conducted using SYBR® Premix-Ex TagTM (Takara, Dalian, China) on a Roche LightCycler 480® System (Roche Applied Science, Penzberg, Germany). act-1 gene was used for an internal control. The primers used for PCR are listed in Table S2.
4.7. Fluorescence Microscopic Analysis
For detecting fluorescence in worms, analysis, synchronized young adult worms expressing either nlp-29p::gfp or gst-4p::gfp were exposed to N. farcinica or E. coli OP50 for 24 h. Then the worms were mounted in M9 onto microscope slides. The slides were imaged using a Nikon e800 fluorescence microscope. Fluorescence intensity was quantified by using the ImageJ software (NIH). Mean value and standard errors were calculated based on more than 30 worms under each condition in three independent experiments.
4.8. ROS Analysis
ROS was detected by transferring worms in to M9 buffer with DHE (3 μM) and stained for 3 h before mounting in M9 buffer onto microscope slide, examined by fluorescence microscope (Zeiss Axioskop 2 Plus) [57].
4.9. The Colony Forming Units (CFU) Analysis
To ask if N. farcinica colonized in worms, we fed young adult worms with either E. coli OP50 or N. farcinica for 96 h on BHI plates, then worms were transferred in to M9 buffer containing 25 mM levamisole hydrochloride (Sangon Biotech Co., Shanghai, China), 50 μg/mL kanamycin and 100 μg/mL carbenicillin ( Sangon Biotech Co., Shanghai, China), and soaking for 30 min before washing with M9 buffer for three times. Worms were collected and grinded in PBS with 0.1% Triton, and serial diluted before placing on BHI ager for incubation, then bacterial colonies were counted to measure CFU.
4.10. Statistical Analysis
The statistical significance of differences in gene expression and fluorescence intensity was assessed by performing a one-way ANOVA followed by a Student-Newman-Keuls test and two-tailed unpaired t-test. Differences in survival rates were analyzed using the log-rank test. Difference of CFU counting was assessed by two-tailed unpaired t-test. Data were analyzed using SPSSsoftware, version 26.0 (SPSS Inc., Chicago, Illinois, USA) and Graphpad Prism 8.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/pathogens11101071/s1, Table S1: Lifespan assay, Table S2: Primers of QPCR analysis.
Author Contributions
Conceptualization, Q.W. and C.Z.; methodology, Q.W. and R.Y.; software, R.Y.; validation, R.Y., Y.K. and J.D.; formal analysis, R.Y. and J.D.; investigation, R.Y., Q.W. and J.D.; resources, Y.K.; data curation, J.D. and Y.K.; writing—original draft preparation, Q.W.; writing—review and editing, C.Z.; visualization, R.Y.; supervision, C.Z.; project administration, Q.W.; funding acquisition, Q.W. and C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the grants (31770060 and U1802233) from the National Natural Science Foundation of China and grant 202001AU070140 from Yunnan Science and Technology Program Project.
Institutional Review Board Statement
Not applicated.
Informed Consent Statement
Not applicated.
Data Availability Statement
Not applicable.
Acknowledgments
We thank the Caenorhabditis Genetics Center for worm strains.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Beaman, B.L.; Beaman, L. Nocardia species: Host-parasite relationships. Clin. Microbiol Rev. 1994, 7, 213–264. [Google Scholar] [CrossRef] [PubMed]
- Goodall, L.; Faggian, F. Nocardia farcinica brain abscesses in an hiv-positive patient. Int. J. STD AIDS 2012, 23, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Kohbata, S.; Beaman, B.L. L-dopa-responsive movement disorder caused by nocardia asteroides localized in the brains of mice. Infect. Immun. 1991, 59, 181–191. [Google Scholar] [CrossRef]
- Arai, M.A.; Ebihara, I.; Makita, Y.; Hara, Y.; Ishibashi, M. Isolation of nocobactin nas as notch signal inhibitors from nocardia farcinica, a possibility of invasive evolution. J. Antibiot. 2021, 74, 255–259. [Google Scholar] [CrossRef]
- Fares, H.; Greenwald, I. Genetic analysis of endocytosis in caenorhabditis elegans: Coelomocyte uptake defective mutants. Genetics 2001, 159, 133–145. [Google Scholar] [CrossRef] [PubMed]
- Felix, M.A.; Duveau, F. Population dynamics and habitat sharing of natural populations of caenorhabditis elegans and c. Briggsae. BMC Biol. 2012, 10, 59. [Google Scholar] [CrossRef] [PubMed]
- Aballay, A.; Drenkard, E.; Hilbun, L.R.; Ausubel, F.M. Caenorhabditis elegans innate immune response triggered by salmonella enterica requires intact lps and is mediated by a mapk signaling pathway. Curr. Biol. 2003, 13, 47–52. [Google Scholar] [CrossRef]
- Aballay, A.; Yorgey, P.; Ausubel, F.M. Salmonella typhimurium proliferates and establishes a persistent infection in the intestine of caenorhabditis elegans. Curr. Biol. 2000, 10, 1539–1542. [Google Scholar] [CrossRef]
- Irazoqui, J.E.; Troemel, E.R.; Feinbaum, R.L.; Luhachack, L.G.; Cezairliyan, B.O.; Ausubel, F.M. Distinct pathogenesis and host responses during infection of c. Elegans by p. Aeruginosa and s. Aureus. PLoS Pathog. 2010, 6, e1000982. [Google Scholar] [CrossRef]
- Tan, M.-W.; Mahajan-Miklos, S. Killing of caenorhabditis elegans by pseudomonas aeruginosa used to model mammalian bacterial. Proc. Natl. Acad. Sci. USA 1999, 96, 715–720. [Google Scholar] [CrossRef] [Green Version]
- Mylonakis, E.; Ausubel, F.M.; Perfect, J.R.; Heitman, J.; Calderwood, S.B. Killing of caenorhabditis elegans by cryptococcus neoformans as a model of yeast pathogenesis. Proc. Natl. Acad. Sci. USA 2002, 99, 15675–15680. [Google Scholar] [CrossRef] [PubMed]
- Pukkila-Worley, R. Surveillance immunity: An emerging paradigm of innate defense activation in caenorhabditis elegans. PLoS Pathog. 2016, 12, e1005795. [Google Scholar] [CrossRef] [PubMed]
- Garsin, D.A.; Villanueva, J.M.; Begun, J.; Kim, D.H.; Sifri, C.D.; Calderwood, S.B.; Ruvkun, G.; Ausubel, F.M. Long-lived c. Elegans daf-2 mutants are resistant to bacterial pathogens. Science 2003, 300, 1921. [Google Scholar] [CrossRef]
- Kim, D.H.; Feinbaum, R.; Alloing, G.; Emerson, F.E.; Garsin, D.A.; Inoue, H.; Tanaka-Hino, M.; Hisamoto, N.; Matsumoto, K.; Tan, M.W.; et al. A conserved p38 map kinase pathway in caenorhabditis elegans innate immunity. Science 2002, 297, 623–626. [Google Scholar] [CrossRef]
- Zou, C.-G.; Ma, Y.-C.; Dai, L.-L.; Zhan, K.-Q. Autophagy protects c. Elegans against necrosis during pseudomonas aeruginosa infection. Proc. Natl. Acad. Sci. USA 2014, 111, 12480–12485. [Google Scholar] [CrossRef] [PubMed]
- Huffman, D.L.; Abrami, L.; Sasik, R.; Corbeil, J.; van der Goot, F.G.; Aroian, R.V. Mitogen-activated protein kinase pathways defend against bacterial pore-forming toxins. Proc. Natl. Acad. Sci. USA 2004, 101, 10995–11000. [Google Scholar] [CrossRef]
- Ren, M.; Feng, H.; Fu, Y.; Land, M.; Rubin, C.S. Protein kinase d is an essential regulator of c. Elegans innate immunity. Immunity 2009, 30, 521–532. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Kim, D.H.; Ausubel, F.M. The g protein-coupled receptor fshr-1 is required for the caenorhabditis elegans innate immune response. Proc. Natl. Acad. Sci. USA 2009, 106, 2782–2787. [Google Scholar] [CrossRef]
- Kawli, T.; He, F.; Tan, M.W. It takes nerves to fight infections: Insights on neuro-immune interactions from c. Elegans. Dis. Models Mech. 2010, 3, 721–731. [Google Scholar] [CrossRef]
- Visvikis, O.; Ihuegbu, N.; Labed, S.A.; Luhachack, L.G.; Alves, A.F.; Wollenberg, A.C.; Stuart, L.M.; Stormo, G.D.; Irazoqui, J.E. Innate host defense requires tfeb-mediated transcription of cytoprotective and antimicrobial genes. Immunity 2014, 40, 896–909. [Google Scholar] [CrossRef] [Green Version]
- Shivers, R.P.; Kooistra, T.; Chu, S.W.; Pagano, D.J.; Kim, D.H. Tissue-specific activities of an immune signaling module regulate physiological responses to pathogenic and nutritional bacteria in c. Elegans. Cell Host Microbe 2009, 6, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Troemel, E.R.; Chu, S.W.; Reinke, V.; Lee, S.S.; Ausubel, F.M.; Kim, D.H. P38 mapk regulates expression of immune response genes and contributes to longevity in c. Elegans. PLoS Genet. 2006, 2, e183. [Google Scholar] [CrossRef] [PubMed]
- Richardson, C.E.; Kooistra, T.; Kim, D.H. An essential role for xbp-1 in host protection against immune activation in c. Elegans. Nature 2010, 463, 1092–1095. [Google Scholar] [CrossRef] [PubMed]
- Hoeven, R.; McCallum, K.C.; Cruz, M.R.; Garsin, D.A. Ce-duox1/bli-3 generated reactive oxygen species trigger protective skn-1 activity via p38 mapk signaling during infection in c. Elegans. PLoS Pathog. 2011, 7, e1002453. [Google Scholar]
- Shivers, R.P.; Pagano, D.J.; Kooistra, T.; Richardson, C.E.; Reddy, K.C.; Whitney, J.K.; Kamanzi, O.; Matsumoto, K.; Hisamoto, N.; Kim, D.H. Phosphorylation of the conserved transcription factor atf-7 by pmk-1 p38 mapk regulates innate immunity in caenorhabditis elegans. PLoS Genet. 2010, 6, e1000892. [Google Scholar] [CrossRef] [PubMed]
- Irazoqui, J.E.; Urbach, J.M.; Ausubel, F.M. Evolution of host innate defence: Insights from caenorhabditis elegans and primitive invertebrates. Nat. Rev. Immunol. 2010, 10, 47–58. [Google Scholar] [CrossRef]
- Moy, T.I.; Mylonakis, E.; Calderwood, S.B.; Ausubel, F.M. Cytotoxicity of hydrogen peroxide produced by enterococcus faecium. Infect. Immun. 2004, 72, 4512. [Google Scholar] [CrossRef]
- Nicholas, H.R.; Hodgkin, J. The erk map kinase cascade mediates tail swelling and a protective response to rectal infection in c. Elegans. Curr. Biol. 2004, 14, 1256–1261. [Google Scholar] [CrossRef]
- Kim, D.H.; Liberati, N.T.; Mizuno, T.; Inoue, H.; Hisamoto, N.; Matsumoto, K.; Ausubel, F.M. Integration of caenorhabditis elegans mapk pathways mediating immunity and stress resistance by mek-1 mapk kinase and vhp-1 mapk phosphatase. Proc. Natl. Acad. Sci. USA 2004, 101, 10990–10994. [Google Scholar] [CrossRef]
- Huang, X.; Pan, W.; Kim, W.; White, A.; Li, S.; Li, H.; Lee, K.; Fuchs, B.B.; Zeng, K.; Mylonakis, E. Caenorhabditis elegans mounts a p38 mapk pathway-mediated defence to cutibacterium acnes infection. Cell. Microbiol. 2020, 22, e13234. [Google Scholar] [CrossRef]
- Pujol, N.; Cypowyj, S.; Ziegler, K.; Millet, A.; Astrain, A.; Goncharov, A.; Jin, Y.; Chisholm, A.D.; Ewbank, J.J. Distinct innate immune responses to infection and wounding in the c. Elegans epidermis. Curr. Biol. 2008, 18, 481–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.; Nie, L.; Zhang, W. Predicted highly expressed genes in nocardia farcinica and the implication for its primary metabolism and nocardial virulence. Antonie Van Leeuwenhoek 2006, 89, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Papp, D.; Csermely, P.; Soti, C. A role for skn-1/nrf in pathogen resistance and immunosenescence in caenorhabditis elegans. PLoS Pathog. 2012, 8, e1002673. [Google Scholar] [CrossRef]
- Inoue, H.; Hisamoto, N.; An, J.H.; Oliveira, R.P.; Nishida, E.; Blackwell, T.K.; Matsumoto, K. The c. Elegans p38 mapk pathway regulates nuclear localization of the transcription factor skn-1 in oxidative stress response. Genes Dev. 2005, 19, 2278–2283. [Google Scholar] [CrossRef]
- An, J.H.; Blackwell, T.K. Skn-1 links c. Elegans mesendodermal specification to a conserved oxidative stress response. Genes Dev. 2003, 17, 1882–1893. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.P.; Porter Abate, J.; Dilks, K.; Landis, J.; Ashraf, J.; Murphy, C.T.; Blackwell, T.K. Condition-adapted stress and longevity gene regulation by caenorhabditis elegans skn-1/nrf. Aging Cell 2009, 8, 524–541. [Google Scholar] [CrossRef]
- Park, S.K.; Tedesco, P.M.; Johnson, T.E. Oxidative stress and longevity in caenorhabditis elegans as mediated by skn-1. Aging Cell 2009, 8, 258–269. [Google Scholar] [CrossRef]
- Liberati, N.T.; Fitzgerald, K.A.; Kim, D.H.; Feinbaum, R.; Golenbock, D.T.; Ausubel, F.M. Requirement for a conserved toll/interleukin-1 resistance domain protein in the caenorhabditis elegans immune response. Proc. Natl. Acad. Sci. USA 2004, 101, 6593–6598. [Google Scholar] [CrossRef]
- Sifri, C.D.; Begun, J.; Ausubel, F.M.; Calderwood, S.B. Caenorhabditis elegans as a model host for staphylococcus aureus pathogenesis. Infect. Immun. 2003, 71, 2208–2217. [Google Scholar] [CrossRef]
- Bolz, D.D.; Tenor, J.L.; Aballay, A. A conserved pmk-1/p38 mapk is required in caenorhabditis elegans tissue-specific immune response to yersinia pestis infection. J. Biol. Chem. 2010, 285, 10832–10840. [Google Scholar] [CrossRef]
- Galbadage, T.; Shepherd, T.F.; Cirillo, S.L.; Gumienny, T.L.; Cirillo, J.D. The caenorhabditis elegans p38 mapk gene plays a key role in protection from mycobacteria. MicrobiologyOpen 2016, 5, 436–452. [Google Scholar] [CrossRef] [PubMed]
- Chou, T.C.; Chiu, H.C.; Kuo, C.J.; Wu, C.M.; Syu, W.J.; Chiu, W.T.; Chen, C.S. Enterohaemorrhagic escherichia coli o157:H7 shiga-like toxin 1 is required for full pathogenicity and activation of the p38 mitogen-activated protein kinase pathway in caenorhabditis elegans. Cell. Microbiol. 2013, 15, 82–97. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Davis, R.J.; Flavell, R.A. Map kinases in the immune response. Annu. Rev. Immunol. 2002, 20, 55–72. [Google Scholar] [CrossRef] [PubMed]
- Matsuzawa, A.; Saegusa, K.; Noguchi, T.; Sadamitsu, C.; Nishitoh, H.; Nagai, S.; Koyasu, S.; Matsumoto, K.; Takeda, K.; Ichijo, H. Ros-dependent activation of the traf6-ask1-p38 pathway is selectively required for tlr4-mediated innate immunity. Nat. Immunol. 2005, 6, 587–592. [Google Scholar] [CrossRef]
- Eisenblatter, M.; Buchal, A.; Gayum, H.; Jasny, E.; Renner Viveros, P.; Ulrichs, T.; Schneider, T.; Schumann, R.R.; Zweigner, J.; Ignatius, R. Nocardia farcinica activates human dendritic cells and induces secretion of interleukin-23 (il-23) rather than il-12p70. Infect. Immun. 2012, 80, 4195–4202. [Google Scholar] [CrossRef]
- Cooper, A.M.; Solache, A.; Khader, S.A. Interleukin-12 and tuberculosis: An old story revisited. Curr. Opin. Immunol. 2007, 19, 441–447. [Google Scholar] [CrossRef]
- Chen, Y.W.; Wen-Chien, K.; Chen, C.S.; Chen, P.L. Riok-1 is a suppressor of the p38 mapk innate immune pathway in caenorhabditis elegans. Front. Immunol. 2018, 9, 774. [Google Scholar] [CrossRef]
- Ma, Y.C.; Dai, L.L.; Qiu, B.B.; Zhou, Y.; Zhao, Y.Q.; Ran, Y.; Zhang, K.Q.; Zou, C.G. Tor functions as a molecular switch connecting an iron cue with host innate defense against bacterial infection. PLoS Genet. 2021, 17, e1009383. [Google Scholar] [CrossRef]
- Steinbaugh, M.J.; Devi, N.S.; Stacey, R.S.; Moronetti, M.; Dreyfuss, J.M.; Hourihan, J.M.; Prashant, R.; Operaña, T.N.; Reza, E.; Keith, B.T. Lipid-mediated regulation of skn-1/nrf in response to germ cell absence. eLife 2015, 4, e07836. [Google Scholar]
- Mai-Britt, M.; Rikke, K.; Bang, H.E.; Braun, O.; Fernandez, G.S.; Kristian, H.; Ejsing, C.S.; Færgeman, N.J.; Junichi, S. Functional loss of two ceramide synthases elicits autophagy-dependent lifespan extension in c. Elegans. PLoS ONE 2013, 8, e70087. [Google Scholar]
- Ji, X.; Zhang, X.; Li, H.; Sun, L.; Hou, X.; Song, H.; Han, L.; Xu, S.; Qiu, X.; Wang, X.; et al. Nfa34810 facilitates nocardia farcinica invasion of host cells and stimulates tumor necrosis factor alpha secretion through activation of the nf-kappab and mitogen-activated protein kinase pathways via toll-like receptor 4. Infect. Immun. 2020, 88, e00831-19. [Google Scholar] [CrossRef]
- Brenner, S. The genetics of caenorhabditis elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef] [PubMed]
- Tejeda-Benitez, L.; Olivero-Verbel, J. Caenorhabditis elegans, a biological model for research in toxicology. Rev. Environ. Contam. Toxicol. 2016, 237, 1–35. [Google Scholar]
- Kamath, R.S.; Ahringer, J. Genome. Methods 2003, 30, 313–321. [Google Scholar] [CrossRef]
- Chen, Y.L.; Tao, J.; Zhao, P.J.; Tang, W.; Xu, J.P.; Zhang, K.Q.; Zou, C.G. Adiponectin receptor paqr-2 signaling senses low temperature to promote c. Elegans longevity by regulating autophagy. Nat. Commun. 2019, 10, 2602. [Google Scholar] [CrossRef]
- Girard, L.R.; Fiedler, T.J.; Harris, T.W.; Carvalho, F.; Antoshechkin, I.; Han, M.; Sternberg, P.W.; Stein, L.D.; Chalfie, M. Wormbook: The online review of caenorhabditis elegans biology. Nucleic Acids Res. 2007, 35, D472–D475. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Wu, Q.Y.; Ma, Y.C.; Chen, Y.L.; Zou, C.G. Antioxidant response is a protective mechanism against nutrient deprivation in c. Elegans. Sci. Rep. 2017, 7, 43547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
N. farcinica does not affect lifespan of adult C. elegans. Adult C. elegans was placed on BHI medium with either E. coli OP50 or N. farcinica at 25 °C, the lifespan was monitored at 24 h interval. The lifespan of WT worms was comparable when fed on E. coli OP50 and N. farcinica, respectively (log-rank p = 0.6). See Table S1 for details.
Figure 1.
N. farcinica does not affect lifespan of adult C. elegans. Adult C. elegans was placed on BHI medium with either E. coli OP50 or N. farcinica at 25 °C, the lifespan was monitored at 24 h interval. The lifespan of WT worms was comparable when fed on E. coli OP50 and N. farcinica, respectively (log-rank p = 0.6). See Table S1 for details.

Figure 2.
The role of the innate immune pathways in resistance to N. farcinica in worms. (a–j) Survival of C. elegans fed either N. farcinica (NF) or E. coli OP50 were monitored over time at 25 °C. The survival rates of jnk-1(gk7) (a), mpk-1(n2521) (b), hlh-30(tm1978) (c) mutants were comparable to worms in the presence of either E. coli OP50 or N. farcinica. RNAi knockdown of dkf-2 (d), fshr-1 (e), egl-30 (f), daf-16 (g) did not influence resistance to N. farcinica. Mutations in pmk-1(km25) (h), nsy-1(ag3) (i), sek-1(ag1) (j) enhanced susceptibility of worms to N. farcinica, (log-rank p < 0.001). See Table S1 for details.
Figure 2.
The role of the innate immune pathways in resistance to N. farcinica in worms. (a–j) Survival of C. elegans fed either N. farcinica (NF) or E. coli OP50 were monitored over time at 25 °C. The survival rates of jnk-1(gk7) (a), mpk-1(n2521) (b), hlh-30(tm1978) (c) mutants were comparable to worms in the presence of either E. coli OP50 or N. farcinica. RNAi knockdown of dkf-2 (d), fshr-1 (e), egl-30 (f), daf-16 (g) did not influence resistance to N. farcinica. Mutations in pmk-1(km25) (h), nsy-1(ag3) (i), sek-1(ag1) (j) enhanced susceptibility of worms to N. farcinica, (log-rank p < 0.001). See Table S1 for details.

Figure 3.
N. farcinica activates the p38 MAPK pathway in adult worms. (a) The phosphorylation of p38 MAPK was increase after feeding on N. farcinica (NF) for 24 h. (b) The level of phosphorylated p38 MAPK were quantified using Image J. The result is analyzed by t-test. * p < 0.05 versus E. coli OP50. (c) The expression of Pnlp-29::gfp was up-regulated in WT worms, but weaker in worms subjected to nsy-1, sek-1 and pmk-1 RNAi exposed to N.farcinica. Scale bar, 500 µm. (d) The fluorescent intensity of Pnlp-29::gfp. These results are means ± SD of three independent experiments (n = 35 worms per experiment). p values were calculated by one-way ANOVA followed by a Student-Newman-Keuls test and t-test. *** p < 0.001 versus E. coli OP50. (e) The mRNA levels of PMK-1 target genes (nlp-29, F08G5.6, F35E12.5 and Y37A1A.2) after feeding on either E. coli OP50 or N. farcinica, in WT or nsy-1(ag3), sek-1(ag1), or pmk-1(km25). *** p < 0.001 versus wild type. (f) DHE stain showed that N. farcinica induced slightly growing level of ROS in worms after 24 h exposure. Scale bar, 100 µm. (g) Quantification of relative fluorescent intensity of the DHE stain. n = 40, ns, not significant.
Figure 3.
N. farcinica activates the p38 MAPK pathway in adult worms. (a) The phosphorylation of p38 MAPK was increase after feeding on N. farcinica (NF) for 24 h. (b) The level of phosphorylated p38 MAPK were quantified using Image J. The result is analyzed by t-test. * p < 0.05 versus E. coli OP50. (c) The expression of Pnlp-29::gfp was up-regulated in WT worms, but weaker in worms subjected to nsy-1, sek-1 and pmk-1 RNAi exposed to N.farcinica. Scale bar, 500 µm. (d) The fluorescent intensity of Pnlp-29::gfp. These results are means ± SD of three independent experiments (n = 35 worms per experiment). p values were calculated by one-way ANOVA followed by a Student-Newman-Keuls test and t-test. *** p < 0.001 versus E. coli OP50. (e) The mRNA levels of PMK-1 target genes (nlp-29, F08G5.6, F35E12.5 and Y37A1A.2) after feeding on either E. coli OP50 or N. farcinica, in WT or nsy-1(ag3), sek-1(ag1), or pmk-1(km25). *** p < 0.001 versus wild type. (f) DHE stain showed that N. farcinica induced slightly growing level of ROS in worms after 24 h exposure. Scale bar, 100 µm. (g) Quantification of relative fluorescent intensity of the DHE stain. n = 40, ns, not significant.

Figure 4.
N. farcinica activates SKN-1 through the p38 MAPK pathway. (a) The nuclear translocation of SKN-1::GFP was increased in worms fed N. farcinica for 24 h. Scale bar, 100 µm. (b) The expression of Pgst-4::gfp in worms in the presence of E. coli OP50 or N. faricinica for 24 h. Scale bar, 500 µm. (c) Quantification of Pgst-4::gfp fluorescence intensity in worms. These results are means ± SD of three independent experiments (n = 40 worms per experiment). p values were calculated by one-way ANOVA followed by a Student-Newman-Keuls test and t-test. n = 40, *** p < 0.001. (d) qPCR of gst-4 mRNA level in worms exposed to either OP50 or N. farcinica for 24 h. *** p < 0.001.
Figure 4.
N. farcinica activates SKN-1 through the p38 MAPK pathway. (a) The nuclear translocation of SKN-1::GFP was increased in worms fed N. farcinica for 24 h. Scale bar, 100 µm. (b) The expression of Pgst-4::gfp in worms in the presence of E. coli OP50 or N. faricinica for 24 h. Scale bar, 500 µm. (c) Quantification of Pgst-4::gfp fluorescence intensity in worms. These results are means ± SD of three independent experiments (n = 40 worms per experiment). p values were calculated by one-way ANOVA followed by a Student-Newman-Keuls test and t-test. n = 40, *** p < 0.001. (d) qPCR of gst-4 mRNA level in worms exposed to either OP50 or N. farcinica for 24 h. *** p < 0.001.

Figure 5.
SKN-1 promotes worm resistance to N. farcinica. (a) Survival of wild type (WT) worms and skn-1(tm3411) mutants fed either E. coli OP50 or N. farcinica at 25 °C. skn-1(tm3411) worms were more sensitive to N. farcinica than WT worms. p < 0.001. (b) The susceptibility of pmk-1(km25) worms subjected to empty vector was similar to that of pmk-1(km25) worms subjected to skn-1 RNAi in the presence of N. farcinica, (log-rank p = 0.15).
Figure 5.
SKN-1 promotes worm resistance to N. farcinica. (a) Survival of wild type (WT) worms and skn-1(tm3411) mutants fed either E. coli OP50 or N. farcinica at 25 °C. skn-1(tm3411) worms were more sensitive to N. farcinica than WT worms. p < 0.001. (b) The susceptibility of pmk-1(km25) worms subjected to empty vector was similar to that of pmk-1(km25) worms subjected to skn-1 RNAi in the presence of N. farcinica, (log-rank p = 0.15).

Figure 6.
PMK-1 and SKN-1 are critical in elimination of N. farcinica. The colony forming units (CFU) of N. farcinica in young adult worms exposed to either E. coli OP50 or N. farcinica for 96 h. (a) pmk-1(km25) worms showed an increase in CFU of either E. coli OP50 or N. farcinica. ** p < 0.01 versus wild type, *** p < 0.001 versus wild type. (b) Inhibition of skn-1 by RNAi increased CFU of these two bacteria in worms. ** p < 0.01 versus vehicle, *** p < 0.001 versus vehicle. For each group, 50–60 worms were used in single measurement, and 6–10 independent experiments were carried out, p values were calculated using t-test.
Figure 6.
PMK-1 and SKN-1 are critical in elimination of N. farcinica. The colony forming units (CFU) of N. farcinica in young adult worms exposed to either E. coli OP50 or N. farcinica for 96 h. (a) pmk-1(km25) worms showed an increase in CFU of either E. coli OP50 or N. farcinica. ** p < 0.01 versus wild type, *** p < 0.001 versus wild type. (b) Inhibition of skn-1 by RNAi increased CFU of these two bacteria in worms. ** p < 0.01 versus vehicle, *** p < 0.001 versus vehicle. For each group, 50–60 worms were used in single measurement, and 6–10 independent experiments were carried out, p values were calculated using t-test.

Figure 7.
PMK-1 and SKN-1 prevent tissus damages upon N. farcinica exposure. DIC images of necrotic cells found in head of worms after 4 days of infection. (a) pmk-1(km25) worms showed enlarged vacuoles in the head of worms after N. farcinica infection. (b) Worms subjected to skn-1 RNAi exhibited enlarged vacuoles in the head of worms. Enlarged vacuoles are pointed out by black arrows. Scale bar, 100 µm.
Figure 7.
PMK-1 and SKN-1 prevent tissus damages upon N. farcinica exposure. DIC images of necrotic cells found in head of worms after 4 days of infection. (a) pmk-1(km25) worms showed enlarged vacuoles in the head of worms after N. farcinica infection. (b) Worms subjected to skn-1 RNAi exhibited enlarged vacuoles in the head of worms. Enlarged vacuoles are pointed out by black arrows. Scale bar, 100 µm.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, R.; Kang, Y.; Duan, J.; Zou, C.; Wu, Q. The p38 MAPK/PMK-1 Pathway Is Required for Resistance to Nocardia farcinica Infection in Caenorhabditis elegance. Pathogens 2022, 11, 1071. https://doi.org/10.3390/pathogens11101071
AMA Style
Yang R, Kang Y, Duan J, Zou C, Wu Q. The p38 MAPK/PMK-1 Pathway Is Required for Resistance to Nocardia farcinica Infection in Caenorhabditis elegance. Pathogens. 2022; 11(10):1071. https://doi.org/10.3390/pathogens11101071
Chicago/Turabian StyleYang, Ruiqiu, Yingqian Kang, Jiahong Duan, Chenggang Zou, and Qinyi Wu. 2022. "The p38 MAPK/PMK-1 Pathway Is Required for Resistance to Nocardia farcinica Infection in Caenorhabditis elegance" Pathogens 11, no. 10: 1071. https://doi.org/10.3390/pathogens11101071
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.