
In Vitro Activity of the Arylaminoartemisinin GC012 against Helicobacter pylori and Its Effects on Biofilm

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Results
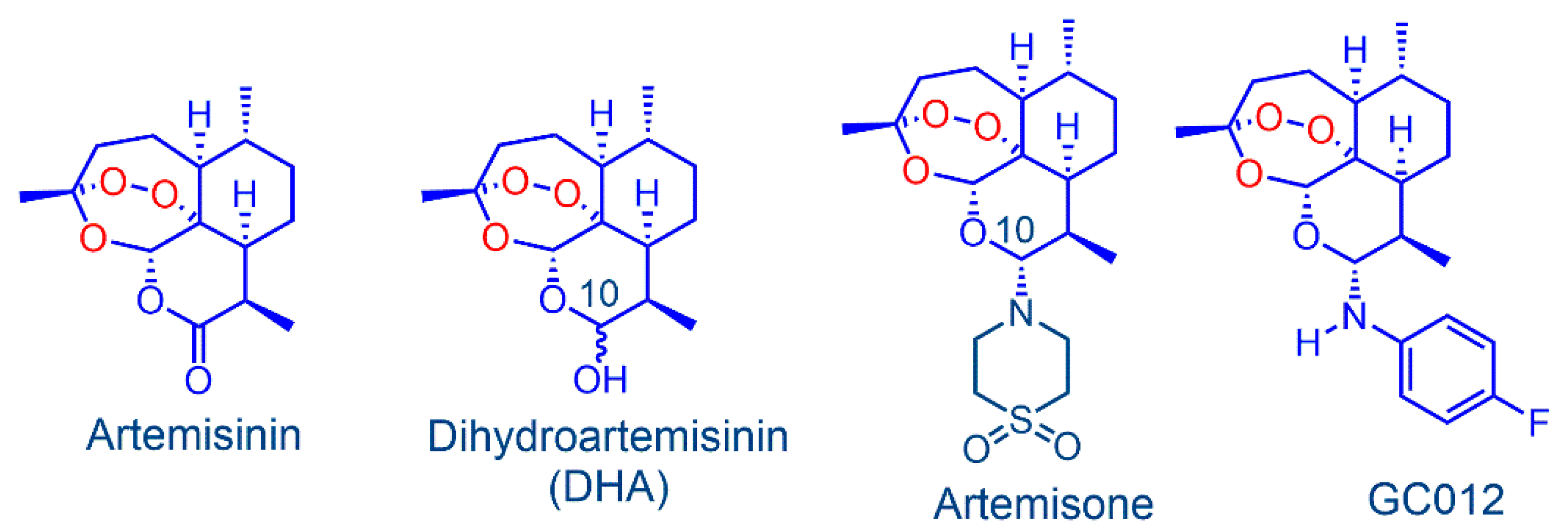
2.1. Antimicrobial Activity of GC012
2.2. Effect of pH on the Stability of GC012
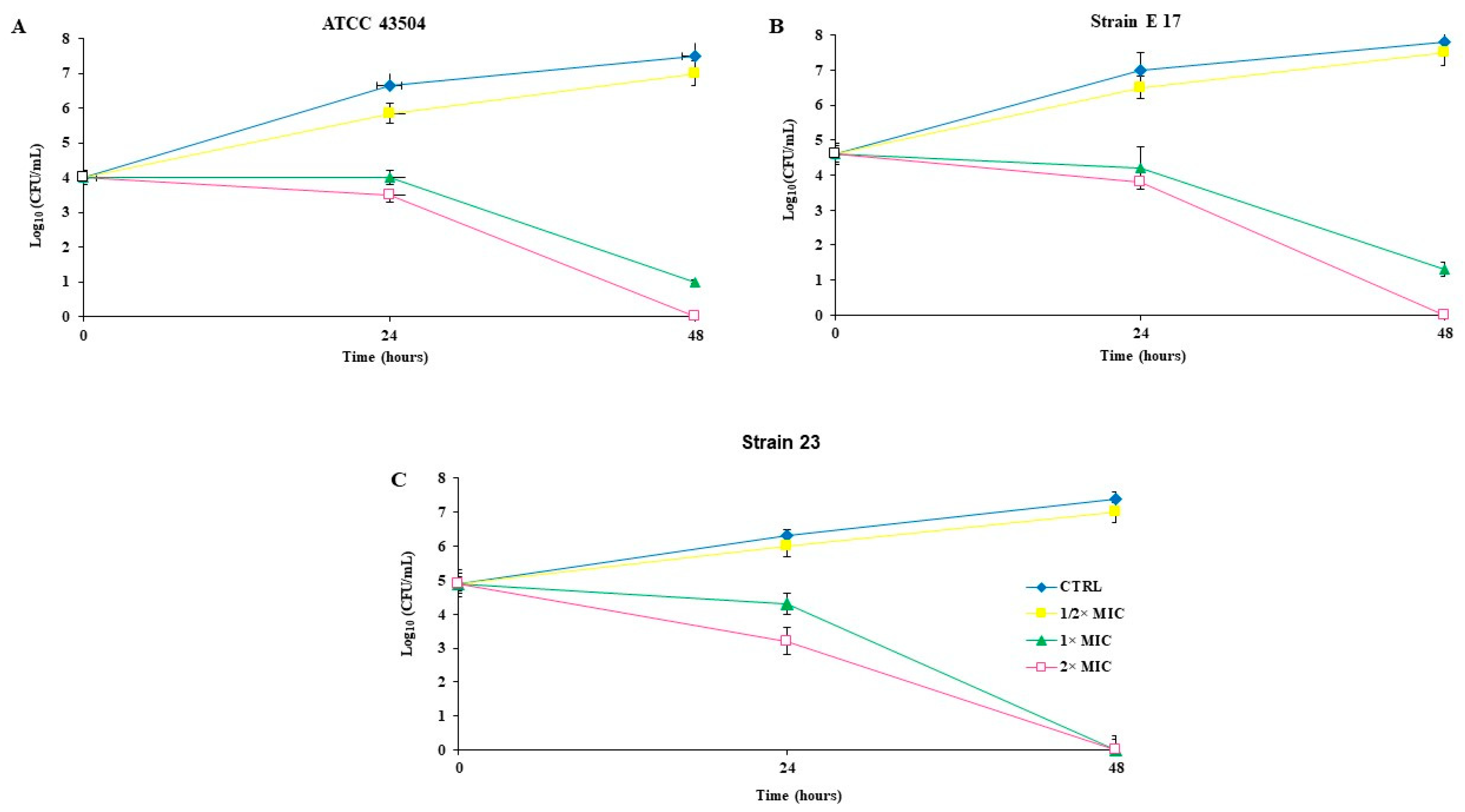
2.3. Killing Kinetics
2.4. Checkerboard Assay
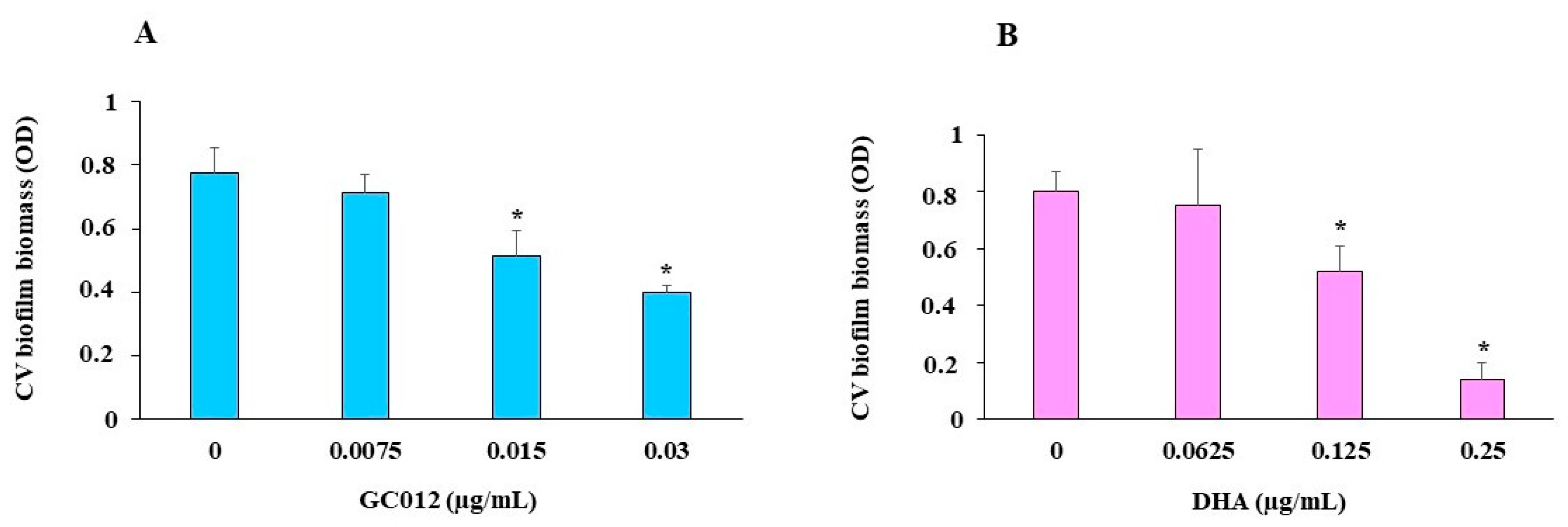
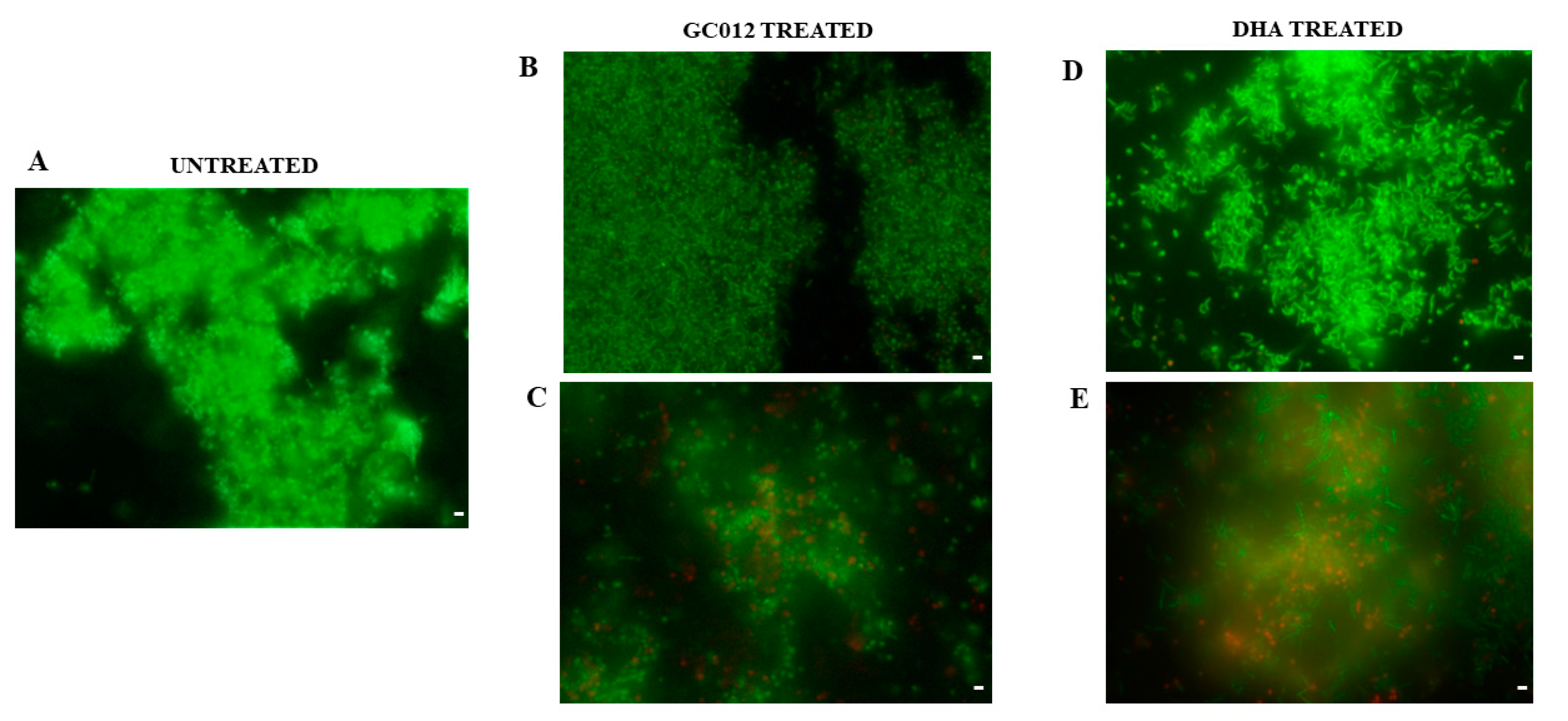
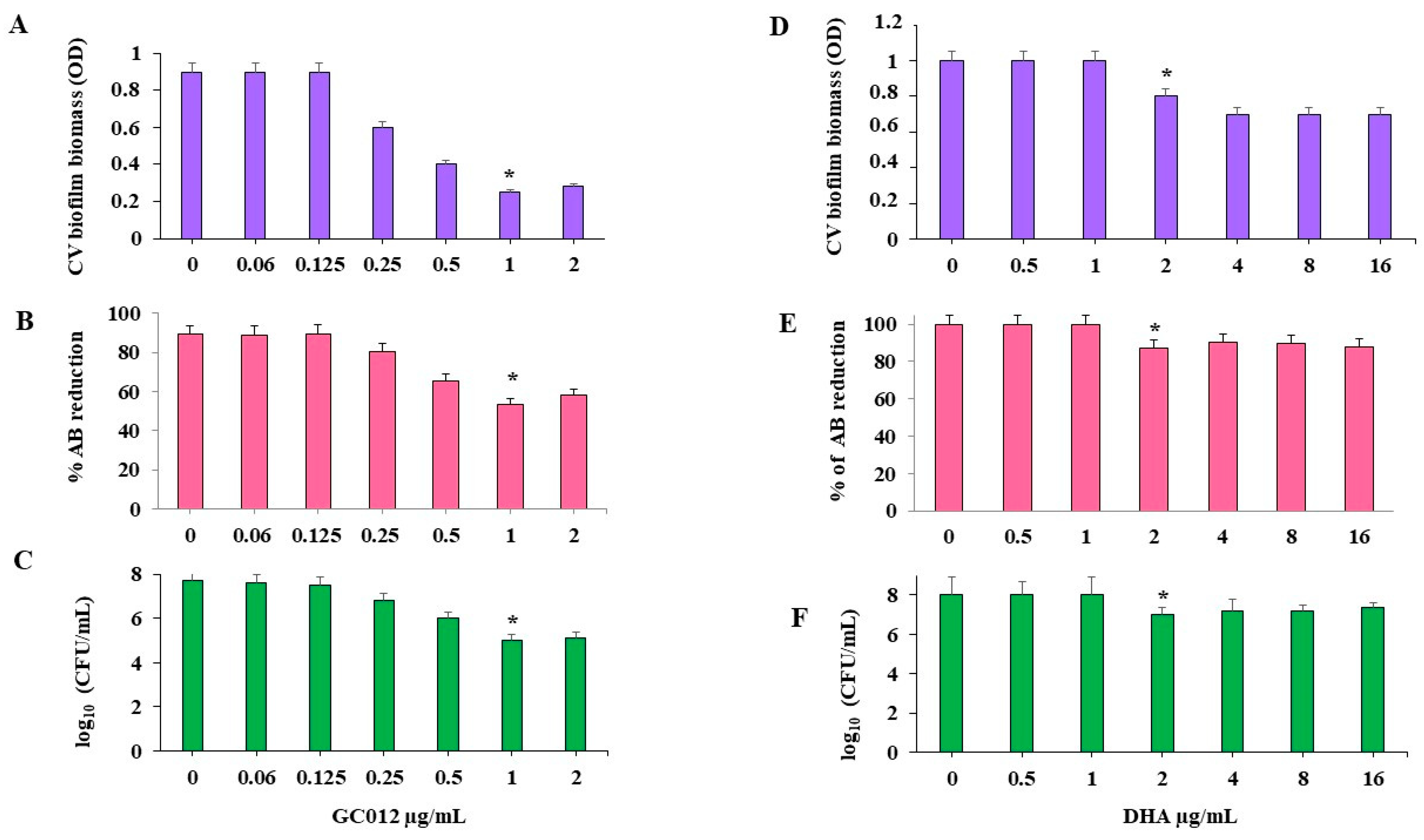
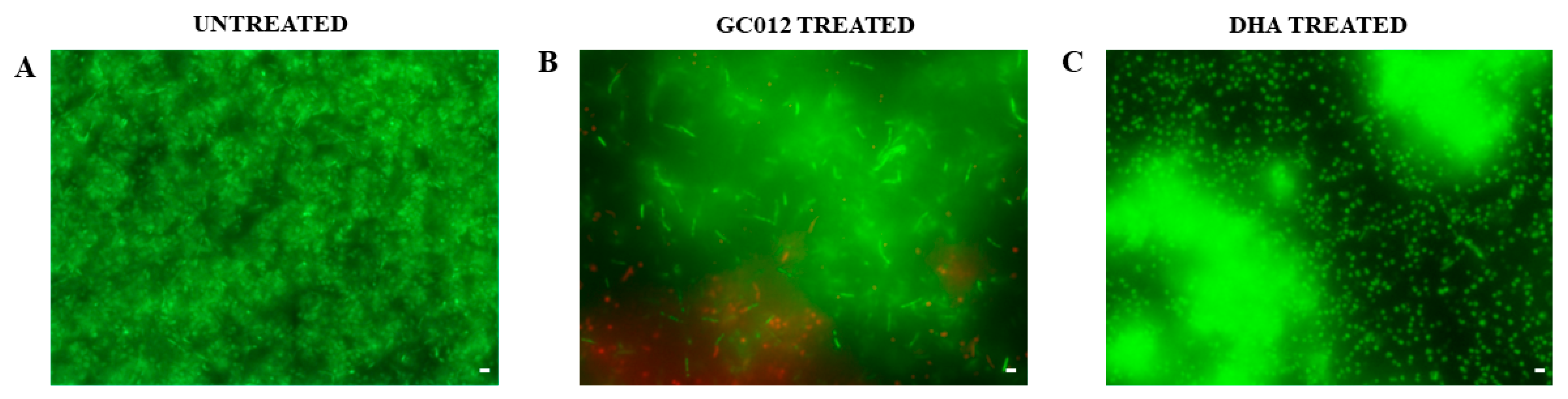
2.5. Prevention of Biofilm Formation
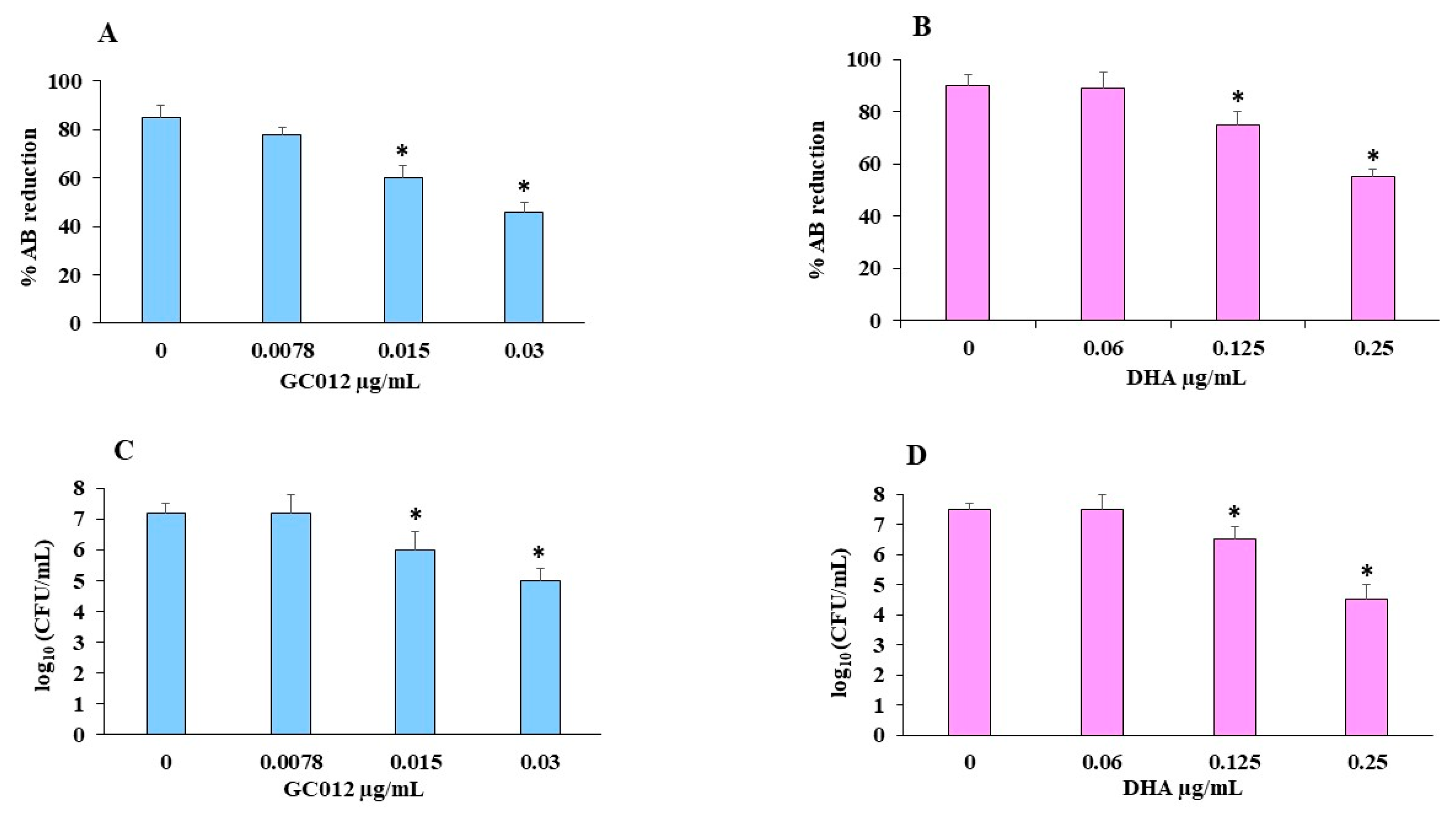
2.6. Eradication of Mature Biofilm
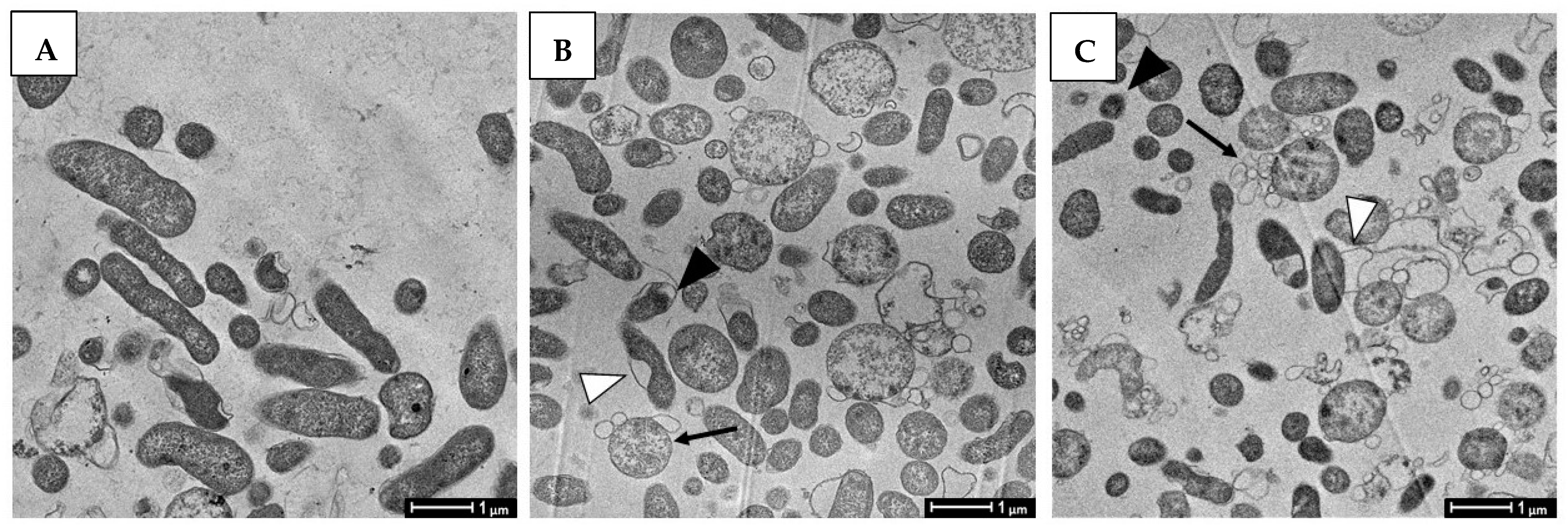
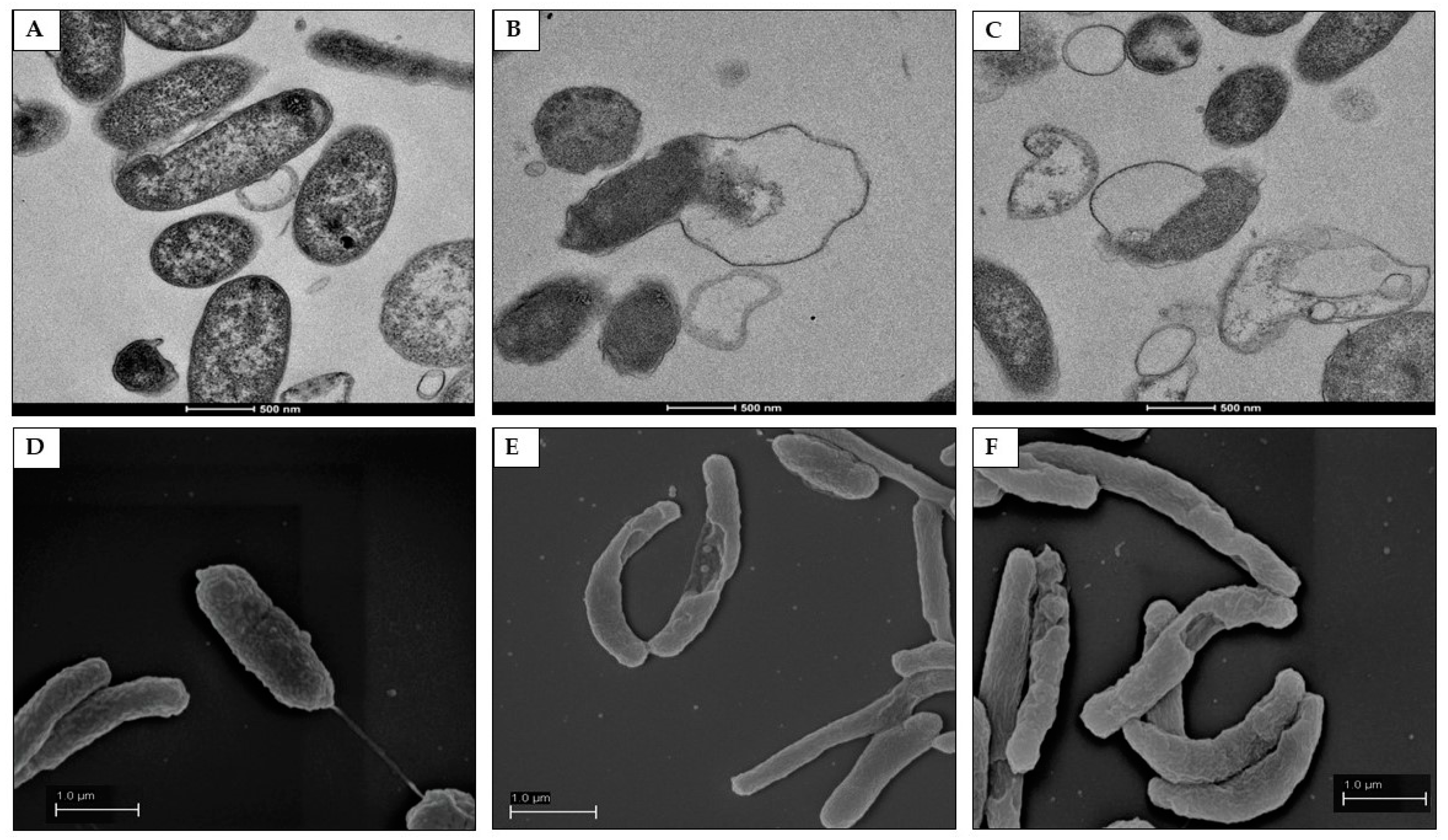
2.7. Morphological and Ultra-Structural Modifications of DHA- and GC012-Treated H. pylori Cells as Visualized with TEM and SEM
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains Culture and Cell Culture
4.2. Reagents
4.3. Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC) Determination
4.4. Determination of Killing Kinetics of GC012
4.5. Combination Effect of Antibiotics with GC012
4.6. Antibiofilm Assay
4.7. Cristal Violet (CV) Staining
4.8. Determination of Minimal Biofilm Inhibitory Concentration (MBIC) and Minimal Biofilm Eradication Concentration (MBEC) Biofilm
4.9. Live/Dead Staining
4.10. MTT Assay
4.11. Urease Inhibitory Effect
4.12. Transmission Electron Microscopy (TEM) and Scanning Electron Microscope (SEM)
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sjomina, O.; Pavlova, J.; Niv, Y.; Leja, M. Epidemiology of Helicobacter pylori infection. Helicobacter 2018, 23 (Suppl. S1), e12514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Working Group on the Evaluation of Carcinogenic Risks to Humans. Schistosomes, Liver Flukes and Helicobacter pylori. IARC Working Group on the Evaluation of Carcinogenic Risks to Human. Lyon, 7–14 June 1994. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 61, 1. [Google Scholar]
- Smyth, E.C.; Nilsson, M.; Grabsch, H.I.; van Grieken, N.C.T.; Lordick, F. Gastric cancer. Lancet 2020, 396, 635–648. [Google Scholar] [CrossRef]
- Kosunen, T.U.; Pukkala, E.; Sarna, S.; Seppala, K.; Aromaa, A.; Knek, P.; Rautelin, H. Gastric cancers in Finnish patients after cure of Helicobacter pylori infection: A cohort study. Int. J. Cancer 2011, 128, 433–439. [Google Scholar] [CrossRef]
- Thung, I.; Aramin, H.; Vavinskaya, V.; Gupta, S.; Park, J.Y.; Crowe, S.E.; Valasek, M.A. The global emergence of Helicobacter pylori antibiotic resistance. Aliment. Pharmacol. Ther. 2016, 43, 514–533. [Google Scholar] [CrossRef] [Green Version]
- Kuo, Y.T.; Liou, J.M.; El-Omar, E.M.; Wu, J.Y.; Leow, A.H.R.; Goh, K.L.; Das, R.; Lu, H.; Lin, J.T.; Tu, Y.K.; et al. Primary antibiotic resistance in Helicobacter pylori in the Asia-Pacific region: A systematic review and meta-analysis. Lancet Gastroenterol. Hepatol. 2017, 2, 707–715. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of Antibiotic Resistance in Helicobacter pylori: A Systematic Review and Metanalysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382. [Google Scholar] [CrossRef] [Green Version]
- Tshibangu-Kamamba, E.; Yamaoka, Y. Helicobacter pylori infection and antibiotic resistance-From biology to clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; O’Morain, C.A.; Gisbert, J.P.; Kuipers, E.J.; Axon, A.T.; Bazzoli, F.; Gasbarrini, A.; Atherton, J.; Graham, D.Y.; et al. Management of Helicobacter pylori infection—the Maastricht V/Florence Consensus Report. Gut 2017, 66, 6–30. [Google Scholar] [CrossRef] [Green Version]
- Chey, W.D.; Leontiadis, G.L.; Howden, C.W.; Moss, S.F. ACG clinical guideline: Treatment of Helicobacter pylori infection. Am. J. Gastroenterol. 2017, 112, 212–238. [Google Scholar] [CrossRef]
- Fallone, C.A.; Chiba, N.; Veldhuyzen van Zanten, S.; Fischbach, L.; Gisbert, J.P.; Hunt, R.H.; Jones, N.L.; Render, C.; Leontiadis, G.I.; Moayyedi, P.; et al. The Toronto Consensus for the treatment of Helicobacter pylori infection in adults. Gastroenterology 2016, 151, 51–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, H.; Suzuki, H. Role of acid suppression in acid-related diseases: Proton pump inhibitor and potassium-competitive acid blocker. J. Neurogastroenterol. Motil. 2019, 25, 6–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stollman, N. Helicobacter pylori Infection in the Era of Antibiotic Resistance. Gastroenterol. Hepatol. 2016, 12, 122–125. [Google Scholar]
- Hunt, R.H.; Xiao, S.D.; Megraud, F.; Leon-Barua, R.; Bazzoli, F.; van der Merwe, S.; Vaz Coelho, L.G.; Fock, M.; Fedail, S.; Cohen, H.; et al. Helicobacer pylori in developing countries. World Gastroenterology Organisation Global Guideline. J. Gastrointestin. Liver Dis. 2011, 20, 299–304. [Google Scholar]
- Suerbaum, S.; Smith, J.M.; Bapumia, K.; Morelli, G.; Smith, N.H.; Kunstmann, E.; Dyrek, I.; Achtmant, M. Free recombination within Helicobacter pylori. Proc. Natl. Acad. Sci. USA 1998, 95, 12619–12624. [Google Scholar] [CrossRef] [Green Version]
- Björkholm, B.; Sjölund, M.; Falk, P.G.; Berg, O.G.; Engstrand, L.; Andersson, D. Mutation frequency and biological cost of antibiotic resistance in Helicobacter pylori. Proc. Natl. Acad. Sci. USA 2001, 98, 14607–14612. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, J. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Review on Antimicrobial Resistance; Wellcome Trust and HM Government. 2016. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 21 May 2022).
- WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. Available online: https://www.who.int/news/item/27-02-2017-who-publishes-list-of-bacteria-for-which-new-antibiotics-are-urgently-needed (accessed on 21 May 2022).
- Hathroubi, S.; Servetas, S.L.; Windham, I.; Merrell, D.S.; Ottemann, K.M. Helicobacter pylori Biofilm Formation and Its Potential Role in Pathogenesis. Microbiol. Mol. Biol. Rev. 2018, 82, e00001-18. [Google Scholar] [CrossRef] [Green Version]
- Hathroubi, S.; Zerebinski, J.; Ottemann, K.M. Helicobacter pylori Biofilm Involves a Multigene Stress-Biased Response, Including a Structural Role for Flagella. mBio 2018, 9, e01973-18. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.L.; Hassanbhai, A.M.; Chen, H.; Huang, Z.Y.; Lin, T.L.; Wu, S.H.; Ho, B. Proteomannans in biofilm of Helicobacter pylori ATCC 43504. Helicobacter 2011, 16, 89–98. [Google Scholar] [CrossRef]
- Grande, R.; Di Marcantonio, M.C.; Robuffo, I.; Pompilio, A.; Celia, C.; Di Marzio, L.; Hall-Stoodley, L. Helicobacter pylori ATCC 43629/NCTC 11639 Outer Membrane Vesicles (OMVs) from biofilm and planktonic phase associated with extracellular DNA (eDNA). Front. Microbiol. 2015, 6, 1369. [Google Scholar] [CrossRef] [Green Version]
- Yonezawa, H.; Osaki, T.; Hanawa, T.; Kurata, S.; Ochiai, K.; Kamiya, S. Impact of Helicobacter pylori biofilm formation on clarithromycin susceptibility and generation of resistance mutations. PLoS ONE 2013, 8, e73301. [Google Scholar] [CrossRef]
- Yonezawa, H.; Osaki, T.; Kamiya, S. Biofilm formation by Helicobacter pylori and its involvement for antibiotic resistance. Biomed. Res. Int. 2015, 2015, 914791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klayman, D.L. Qinghaosu (artemisinin): An antimalarial drug from China. Science 1985, 228, 1049–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Li, Y.; Li, Z.; Zeng, M. Artemisinin-Based and Other Antimalarials: Detailed Account of Studies by Chinese Scientists Who Discovered and Developed Them; Academic Press: Cambridge, MA, USA; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 978-0-12-813133-6. [Google Scholar]
- Eastman, R.T.; Fidock, D.A. Artemisinin-based combination therapies: A vital tool in efforts to eliminate malaria. Nat. Rev. Microbiol. 2009, 7, 864–874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, T.N.C.; Alonso, P.L.; Gutteridge, W.E. New medicines to improve control and contribute to the eradication of malaria. Nat. Rev. Drug Disc. 2009, 8, 879–891. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.K.; Chaudhary, S. Artemisinin-derived antimalarial endoperoxides from bench-side to bed-side: Chronological advancements and future challenges. Med. Res. Rev. 2020, 40, 1220–1275. [Google Scholar] [CrossRef]
- Guo, Z. Artemisinin anti-malarial drugs in China. Acta Pharm. Sin. B 2016, 6, 115–124. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Cao, J.; Huang, G.; Zhao, Q.; Shen, J. Biological activities of artemisinin derivatives beyond malaria. Curr. Top. Med. Chem. 2019, 19, 205–222. [Google Scholar] [CrossRef]
- D’Alessandro, S.; Scaccabarozzi, D.; Signorini, L.; Perego, F.; Ilboudo, D.P.; Ferrante, P.; Delbue, S. The Use of Antimalarial Drugs against Viral Infection. Microorganisms 2020, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Meng, Y.; Ma, N.; Lyu, H.; Wong, Y.K.; Zhang, X.; Zhu, Y.; Gao, P.; Sun, P.; Song, Y.; Lin, L.; et al. Recent pharmacological advances in the repurposing of artemisinin drugs. Med. Res. Rev. 2021, 41, 3156–3181. [Google Scholar] [CrossRef]
- Slade, D.; Galal, A.M.; Gul, W.; Radwan, M.M.; Ahmed, S.A.; Khana, S.I.; Tekwani, B.L.; Jacob, M.R.; Ross, S.A.; ElSohly, M.A. Antiprotozoal, anticancer and antimicrobial activities of dihydroartemisinin acetal dimers and monomers. Bioorg. Med. Chem. 2009, 17, 7949–7957. [Google Scholar] [CrossRef] [Green Version]
- Coertzen, D.; Reader, J.; van der Watt, M.; Nondaba, S.H.; Gibhard, L.; Wiesner, L.; Smith, P.; D’Alessandro, S.; Taramelli, D.; Wong, H.N.; et al. Artemisone and artemiside-potent pan-reactive antimalarial agents that also synergize redox imbalance in P. falciparum transmissible gametocyte stages. Antimicrob. Agents Chemother. 2018, 62, e02214-17. [Google Scholar] [CrossRef] [Green Version]
- Wong, H.N.; Padín-Irizarry, V.; van der Watt, M.E.; Reader, J.; Liebenberg, W.; Wiesner, L.; Smith, P.; Eribez, K.; Winzeler, E.A.; Kyle, D.E.; et al. Optimal 10-aminoartemisinins with potent transmission-blocking capabilities for new artemisinin combination therapies–activities against blood stage P. falciparum Including PfKI3 C580Y mutants and liver stage P. berghei parasites. Front. Chem. 2020, 7, 901. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.J.; Laing, L.; Gibhard, L.; Wong, H.N.; Haynes, R.K.; Wiesner, L. Towards new transmission-blocking combination therapies-pharmacokinetics of 10-amino-artemisinins and 11-aza-artemisinin, and comparison with dihydroartemisinin and artemether. Antimicrob. Agents Chemother. 2021, 65, e00990-21. [Google Scholar] [CrossRef] [PubMed]
- Sisto, F.; Scaltrito, M.M.; Masia, C.; Bonomi, A.; Coccè, V.; Marano, G.; Haynes, R.K.; Miani, A.; Farronato, G.; Taramelli, D. In vitro activity of artemisone and artemisinin derivatives against extracellular and intracellular Helicobacter pylori. Int. J. Antimicrob. Agents 2016, 48, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.K.; Chan, H.W.; Ho, W.Y.; Ko, C.K.F.; Gerena, L.; Kyle, D.E.; Peters, W.; Robinson, B.L. Convenient access both to highly antimalaria active 10-arylaminoartemisinins, alkyl ethers including artemether, arteether, and artelinate. Chem. Bio. Chem. 2005, 6, 659–667. [Google Scholar] [CrossRef]
- Müller, J.; Balmer, V.; Winzer, P.; Rahmana, M.; Manser, V.; Haynes, R.K.; Hemphill, A. In vitro effects of new artemisinin derivatives in Neospora caninum-infected human fibroblasts. Int. J. Antimicrob. Agents 2015, 46, 88–93. [Google Scholar] [CrossRef]
- Bahari, H.M.; Ross, I.N.; Turnberg, L.A. Demonstration of a pH gradient across the mucus layer on the surface of human gastric mucosa in vitro. Gut 1982, 23, 513–516. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.R.; Rabbee, M.F.; Roy, A.; Chowdhury, R.; Banik, A.; Kubra, K.; Mehadi, M.; Chowdhury, H.; Baek, K.H. Therapeutic Promises of Medicinal Plants in Bangladesh and Their Bioactive Compounds against Ulcers and Inflammatory Diseases. Plants 2021, 10, 1348. [Google Scholar] [CrossRef]
- Baker, D.A. Plants against Helicobacter pylori to combat resistance: An ethnopharmacological review. Biotechnol. Rep. 2020, 26, e00470. [Google Scholar] [CrossRef]
- Xie, J.; Lin, Z.; Xian, Y.; Kong, S.; Lai, Z.; Ip, S.; Chen, H.; Guo, H.; Su, Z.; Yang, X.; et al. (−)-Patchouli alcohol protects against Helicobacter pylori urease-induced apoptosis, oxidative stress and inflammatory response in human gastric epithelial cells. Int. Immunopharmacol. 2016, 35, 43–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Monte, C.; Bizzarri, B.; Gidaro, M.C.; Carradori, S.; Mollica, A.; Luisi, G.; Granese, A.; Alcaro, S.; Costa, G.; Basilico, N.; et al. Bioactive compounds of Crocus sativus L. and their semi-synthetic derivatives as promising anti-Helicobacter pylori, anti-malarial and anti-leishmanial agents. J. Enzyme Inhib. Med. Chem. 2015, 30, 1027–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vegaa, A.E.; Wendel, G.H.; Maria, A.O.M.; Pelzer, L. Antimicrobial activity of Artemisia douglasiana and dehydroleucodine against Helicobacter pylori. J. Ethnopharmacol. 2009, 124, 653–655. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Juárez, I.; González, V.; Jaime-Aguilar, H.; Martínez, G.; Linares, E.; Bye, R.; Romero, I. Anti-Helicobacter pylori activity of plants used in Mexican traditional medicine for gastrointestinal disorders. J. Ethnopharmacol. 2009, 122, 402–405. [Google Scholar] [CrossRef] [PubMed]
- Palacios-Espinosa, J.F.; Núñez-Aragón, P.N.; Gomez-Chang, E.; Linares, E.; Bye, R.; Romero, I. Anti-Helicobacter pylori Activity of Artemisia ludoviciana subsp. mexicana and two of Its Bioactive Components, Estafiatin and Eupatilin. Molecules 2021, 26, 3654. [Google Scholar] [CrossRef] [PubMed]
- Goswami, S.; Bhakuni, R.S.; Chinniah, A.; Pal, A.; Kar, S.K.; Dasa, P.K. Anti-Helicobacter pylori Potential of Artemisinin and Its Derivatives. Antimicrob. Agents Chemother. 2012, 56, 4594–4607. [Google Scholar] [CrossRef] [Green Version]
- Carron, M.A.; Tran, V.R.; Sugawa, C.; Coticchia, J.M. Identification of Helicobacter pylori biofilms in human gastric mucosa. J. Gastrointest. Surg. 2006, 10, 712–717. [Google Scholar] [CrossRef]
- Krzyżek, P.; Grande, R.; Migdał, P.; Paluch, E.; Gościniak, G. Biofilm Formation as a Complex Result of Virulence and Adaptive Responses of Helicobacter pylori. Pathogens 2020, 9, 1062. [Google Scholar] [CrossRef]
- Cherian, S.; Forbes, D.; Sanfilippo, F.; Cook, A.; Burgner, D. The epidemiology of Helicobacter pylori infection in African refugee children resettled in Australia. Med. J. Aust. 2008, 189, 438–441. [Google Scholar] [CrossRef]
- Lage, T.C.A.; Maciel, T.M.S.; Mota, Y.C.C.; Sisto, F.; Sabino, J.R.; Santos, J.C.C.; Figueiredo, I.M.; Masia, C.; de Fátima, Â.; Fernandes, S.A.; et al. In vitro inhibition of Helicobacter pylori and interaction studies of lichen natural products with jack bean urease. New J. Chem. 2018, 42, 5356–5366. [Google Scholar] [CrossRef] [Green Version]
- Sisto, F.; Carradori, S.; Guglielmi, P.; Traversi, C.B.; Spano, M.; Sobolev, A.P.; Secci, D.; Di Marcantonio, M.C.; Haloci, E.; Grande, R.; et al. Synthesis and Biological Evaluation of Carvacrol-Based Derivatives as Dual Inhibitors of H. pylori Strains and AGS Cell Proliferation. Pharmaceuticals 2020, 13, 405. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standard Institute (CLSI). Development of In Vitro Susceptibility Testing Criteria and Quality Control Parameters: Approved Guideline, 3rd ed.; Clinical and Laboratory Standard Institute: Wayne, PA, USA, 2008; Volume 28. [Google Scholar]
- Sisto, F.; Scaltrito, M.M.; Russello, G.G.; Bonomi, A.; Dubini, F. Antimicrobial susceptibility testing of Helicobacter pylori determined by microdilution method using a new medium. Curr. Microbiol. 2009, 58, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Clinical and Laboratory Standard Institute(CLSI). Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts: Approved Standard, 3rd ed.; CLSI Document M27-A3; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008. [Google Scholar]
- Doern, C.D. When does 2 plus 2 equal 5? A review of antimicrobial synergy testing. J. Clin. Microbiol. 2014, 52, 4124–4128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronci, M.; Del Prete, S.; Puca, V.; Carradori, S.; Carginale, V.; Muraro, R.; Mincione, G.; Aceto, A.; Sisto, F.; Supuran, C.T.; et al. Identification and characterization of the α-CA in the outer membrane vesicles produced by Helicobacter pylori. J. Enzyme Inhib. Med. Chem. 2019, 34, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Grande, R.; Carradori, S.; Puca, V.; Vitale, I.; Angeli, A.; Nocentini, A.; Bonardi, A.; Gratteri, P.; Lanuti, P.; Bologna, G.; et al. Selective Inhibition of Helicobacter pylori Carbonic Anhydrases by Carvacrol and Thymol Could Impair Biofilm Production and the Release of Outer Membrane Vesicles. Int. J. Mol. Sci. 2021, 22, 11583. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic analyses, in vitro biological activities, and cytotoxicity of Cannabis sativa L. essential oil: A multidisciplinary study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef] [Green Version]
- Mossman, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Tanaka, T.; Kawase, M.; Tani, S. Urease inhibitory activity of simple α, β-unsaturated ketones. Life Sci. 2003, 73, 2985–2990. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Glowniak-Lipa, A.; Ludwiczuk, A.; Baj, T.; Malm, A. The In Vitro Activity of Essential Oils against Helicobacter pylori Growth and Urease Activity. Molecules 2020, 25, 586. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | MIC (µg/mL) | MBC (µg/mL) | |||
|---|---|---|---|---|---|
| Range | MIC50 | MIC90 | Range | MBC50 | |
| GC012 | 0.03–1 | 0.06 | 0.125 | 0.032–0.125 | 0.06 |
| DHA [38] | 0.5–2 | 1 | 2 | 1–4 | 2 |
| MNZ | 1–128 | 1 | 128 | 0.5–128 | 2 |
| CLR | 0.016–256 | 1 | 256 | 0.032–512 | 0.5 |
| AMX | 0.016–0.064 | 0.016 | 0.064 | 0.016–0.064 | 0.016 |
| MIC µg/mL * | |||||
|---|---|---|---|---|---|
| Strains | GC012 | DHA | CPX | MCZ | ECZ |
| Gram-negative bacteria | |||||
| E. coli ATCC 25922 | >128 | >128 | 0.0075 | n.d | n.d |
| K. pneumoniae clinical strain 80 | >128 | >128 | n.d | n.d | n.d |
| S. Typhimurium ATCC 14028 | >128 | >128 | n.d | n.d | n.d |
| P. aeruginosa ATCC 27853 | >128 | >128 | n.d | n.d | n.d |
| A. baumannii clinical strain 1/F1 | >128 | >128 | n.d | n.d | n.d |
| Gram-positive bacteria | |||||
| S. aureus ATCC 29213 | >128 | >128 | 0.25 | n.d | n.d |
| S. epidermidis clinical strain AS | >128 | >128 | n.d. | n.d | n.d |
| E. faecalis ATCC 29212 | >128 | >128 | n.d. | n.d | n.d |
| S. mutans clinical strain 6S8 | >128 | >128 | n.d. | n.d | n.d |
| L. acidophilus ATCC 4357 | >128 | >128 | n.d. | n.d | n.d |
| L. rhamnosus ATCC 53103 | >128 | >128 | n.d. | n.d | n.d |
| L. casei clinical strain 40 | >128 | >128 | n.d. | n.d | n.d |
| Yeasts | |||||
| C. albicans ATCC 90028 | >128 | >128 | n.d | <0.125 | <0.125 |
| C. krusei ATCC 6258 | >128 | >128 | n.d | <0.125 | <0.125 |
| Compounds | H. pylori Strains MIC/MBC (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| pH | ATCC 43504 | 102S | E34 | E17 | 190 | R5 | 23 (This Study) | |
| DHA [38] | 2 | 4/4 | 1/2 | 4/4 | 4/4 | 1/2 | 0.5/1 | 1/1 |
| 5 | 4/4 | 1/2 | 2/4 | 2/4 | 1/2 | 0.5/1 | 0.5/1 | |
| 7 | 2/2 | 0.5/0.5 | 2/2 | 2/2 | 1/1 | 0.5/0.5 | 0.5/0.5 | |
| GC012 | 2 | 0.125/0.125 | 0.06/0.06 | 0.015/0.015 | 0.06/0.06 | 0.03/0.03 | 0.06/0.06 | 0.064/0.064 |
| 5 | 0.5/0.5 | 0.03/0.03 | 0.03/0.03 | 0.06/0.06 | 0.03/0.03 | 0.06/0.06 | 0.064/0.064 | |
| 7 | 0.125/0.125 | 0.03/0.03 | 0.03/0.03 | 0.03/0.03 | 0.03/0.03 | 0.06/0.06 | 0.064/0.064 | |
| MNZ | 7 | 128 | 2 | 1 | 0.064 | 1 | 128 | 1 |
| CLR | 7 | 0.032 | 0.016 | 0.064 | 256 | 0.032 | 16 | 0.064 |
| Antimicrobial susceptibility | MNZR CLRS AMXS | MNZS CLRS AMXS | MNZS CLRS AMXS | MNZS CLRR AMXS | MNZS CLRS AMXS | MNZR CLRR AMXS | MNZS CLRS AMXS | |
| N° Isolates (%) | |||
|---|---|---|---|
| GC012 | |||
| FIC Index | CLR | MNZ | AMX |
| ≥0.5 (synergy) | 3 (100) | 3 (100) | 3 (100) |
| 0.5–1 (additive) | 0 (0) | 0 (0) | 0 (0) |
| 1–4 (neutral) | 0 (0) | 0 (0) | 0 (0) |
| >4 (antagonism) | 0 (0) | 0 (0) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sisto, F.; Carradori, S.; D’Alessandro, S.; Santo, N.; Lattuada, N.; Haynes, R.K.; Taramelli, D.; Grande, R. In Vitro Activity of the Arylaminoartemisinin GC012 against Helicobacter pylori and Its Effects on Biofilm. Pathogens 2022, 11, 740. https://doi.org/10.3390/pathogens11070740
Sisto F, Carradori S, D’Alessandro S, Santo N, Lattuada N, Haynes RK, Taramelli D, Grande R. In Vitro Activity of the Arylaminoartemisinin GC012 against Helicobacter pylori and Its Effects on Biofilm. Pathogens. 2022; 11(7):740. https://doi.org/10.3390/pathogens11070740
Chicago/Turabian StyleSisto, Francesca, Simone Carradori, Sarah D’Alessandro, Nadia Santo, Norma Lattuada, Richard K. Haynes, Donatella Taramelli, and Rossella Grande. 2022. "In Vitro Activity of the Arylaminoartemisinin GC012 against Helicobacter pylori and Its Effects on Biofilm" Pathogens 11, no. 7: 740. https://doi.org/10.3390/pathogens11070740