
Interrelation between Stress Management and Secretion Systems of Ralstonia solanacearum: An In Silico Assessment
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Mining and Categorization of Rs Stress-Responsive Genes
2.1.1. MSR
2.1.2. OSR
2.1.3. OxSR
2.1.4. NxSR
2.1.5. DdSR
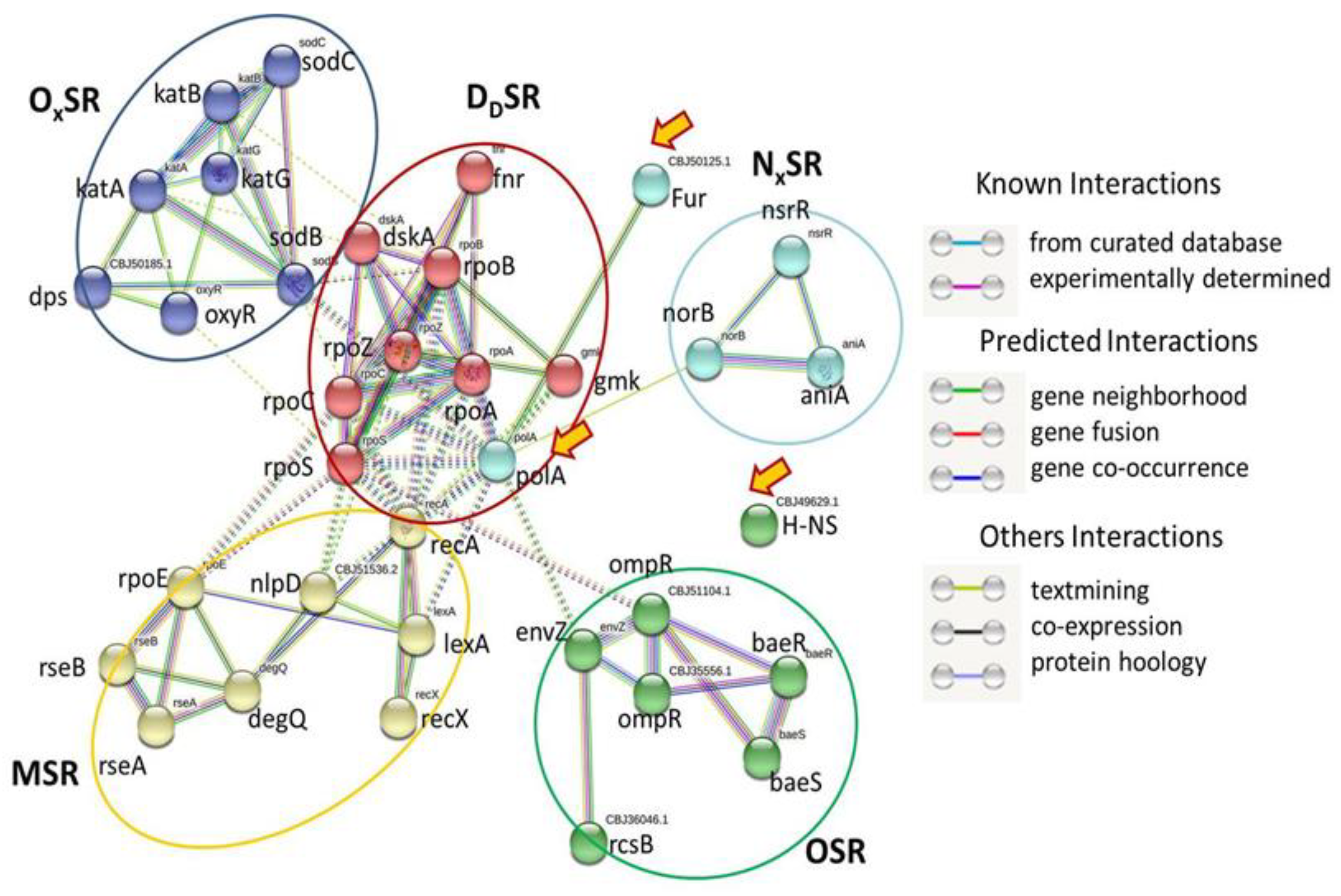
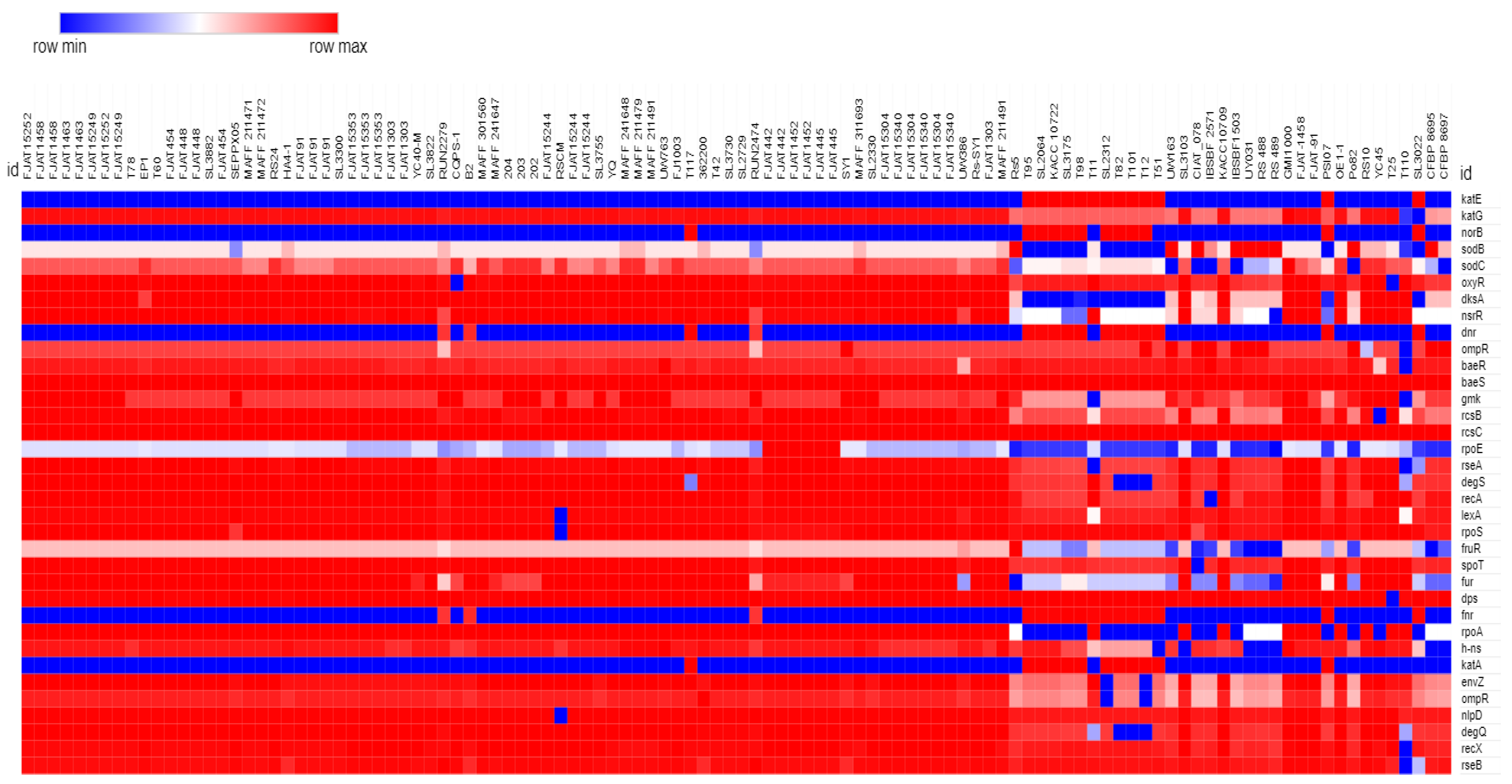
2.2. Homogeneous Distribution of SRGs across Rs Genomes
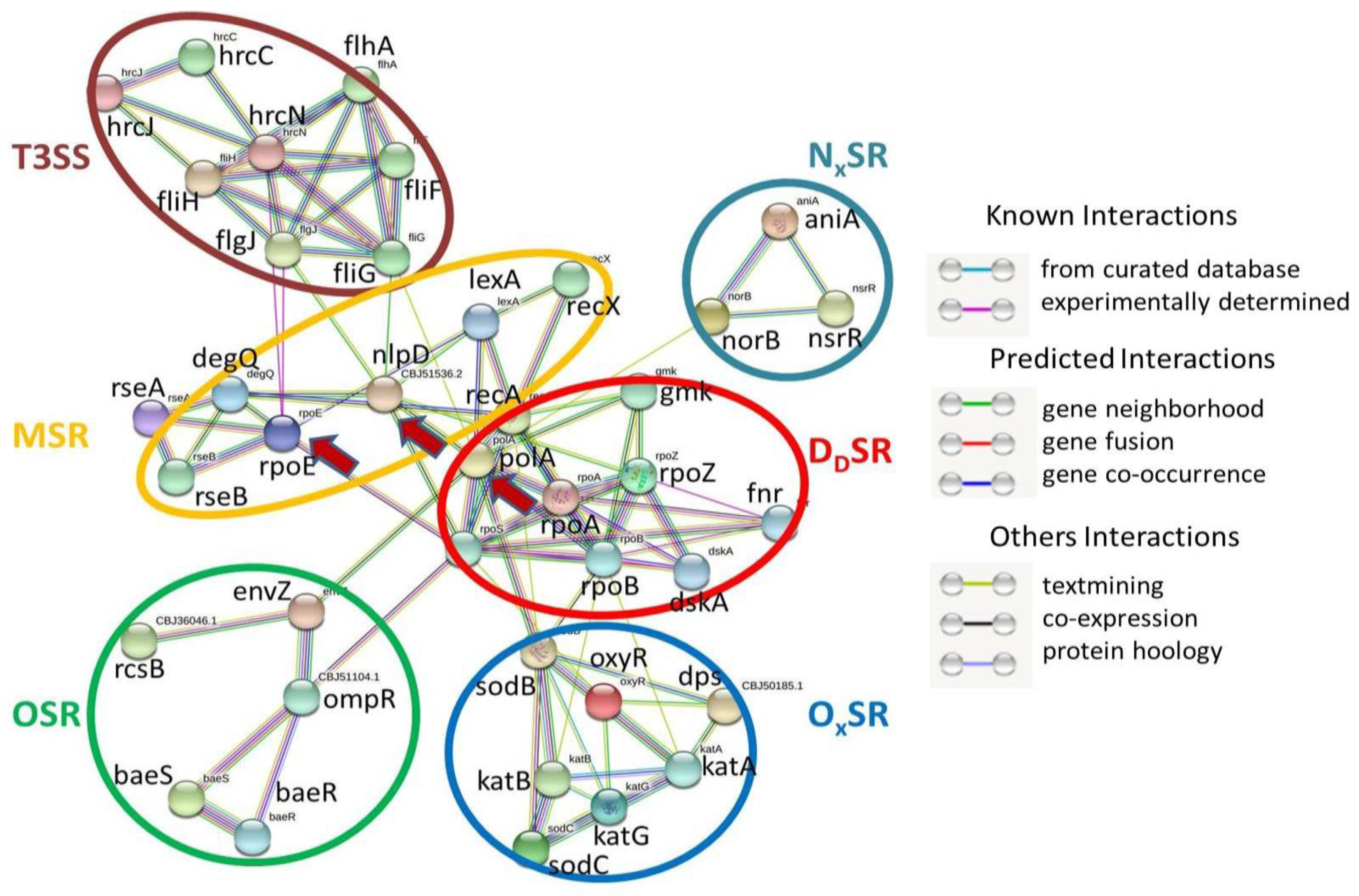
2.3. SRGs and Secretion Systems of Rs
2.4. SRGs and T3SS
2.5. SRGs and GSPS
2.6. SRGs and Pili Systems
3. Discussion
3.1. MSR SRGs of Rs
3.2. OSR SRGs of Rs
3.3. OxSR SRGs of Rs
3.4. NxSR SRGs of Rs
3.5. DdSR SRGs of Rs
3.6. Global Strategy in Stress Response of Rs
3.7. Interactions among T3SS and SRGs of Rs
3.8. Interactions among GSPS and SRGs of Rs
3.9. Interactions among Different Pili Systems and SRGs of Rs
3.10. Network of SRGs and Bacterial Secretion Systems
4. Materials and Methods
4.1. Data Mining
4.2. Mining of Bacterial Genes Coding for Stress-Responsive Enzymes and Stress Regulators
4.3. Mining of Bacterial Secretion System Genes
4.4. Protein–Protein Interaction Network (PIN) Study
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schandry, N.; de Lange, O.; Prior, P.; Lahaye, T. TALE-like Effectors Are an Ancestral Feature of the Ralstonia solanacearum Species Complex and Converge in DNA Targeting Specificity. Front. Plant Sci. 2016, 7, 1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanoubat, M.; Genin, S.; Artiguenave, F.; Gouzy, J.; Mangenot, S.; Arlat, M.; Billault, A.; Brottier, P.; Camus, J.C.; Cattolico, L.; et al. Genome sequence of the plant pathogen Ralstonia solanacearum. Nature 2002, 415, 497–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, P.; Banerjee, G.; Handique, P.J. Use of an abscisic acid-producing Bradyrhizobium japonicum isolate as biocontrol agent against bacterial wilt disease caused by Ralstonia solanacearum. J. Plant Dis. Prot. 2022, 1–11. [Google Scholar] [CrossRef]
- Gutarra, L.; Herrera, J.; Fernandez, E.; Kreuze, J.; Lindqvist-Kreuze, H. Diversity, pathogenicity, and current occurrence of bacterial wilt bacterium Ralstonia solanacearum in Peru. Front. Plant Sci. 2017, 8, 1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filloux, A. Bacterial Regulatory Networks; Caister Academic Press: London, UK, 2012. [Google Scholar]
- Hengge-Aronis, R. Signal Transduction and Regulatory Mechanisms Involved in Control of the σ S (RpoS) Subunit of RNA Polymerase. Microbiol. Mol. Biol. Rev. 2002, 66, 373–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colburn-Clifford, J.M.; Scherf, J.M.; Allen, C. Ralstonia solanacearum Dps Contributes to Oxidative Stress Tolerance and to Colonization of and Virulence on Tomato Plants. Appl. Environ. Microbiol. 2010, 76, 7392–7399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, S.; Kar, I.; Mukherjee, A.K.; Acharya, P. Elicitor-Induced Defense Responses in Solanum lycopersicumagainst against Ralstonia solanacearum. Sci. World J. 2013, 2013, 561056. [Google Scholar] [CrossRef] [Green Version]
- Flores-Cruz, Z.; Allen, C. Necessity of OxyR for the Hydrogen Peroxide Stress Response and Full Virulence in Ralstonia solanacearum. Appl. Environ. Microbiol. 2011, 77, 6426–6432. [Google Scholar] [CrossRef] [Green Version]
- Taylor, P.D.; Inchley, C.J.; Gallagher, M.P. The Salmonella typhimurium AhpC Polypeptide Is Not Essential for Virulence in BALB/c Mice but Is Recognized as an Antigen during Infection. Infect. Immun. 1998, 66, 3208–3217. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Torres, A. Redox Active Thiol Sensors of Oxidative and Nitrosative Stress. Antioxid. Redox Signal. 2012, 17, 1201–1214. [Google Scholar] [CrossRef] [Green Version]
- Karlinsey, J.E.; Bang, I.-S.; Becker, L.A.; Frawley, E.R.; Porwollik, S.; Robbins, H.F.; Thomas, V.C.; Urbano, R.; McClelland, M.; Fang, F.C. The NsrR regulon in nitrosative stress resistance of Salmonella enterica serovar Typhimurium. Mol. Microbiol. 2012, 85, 1179–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crack, J.; Green, J.; Thomson, A.J. Mechanism of Oxygen Sensing by the Bacterial Transcription Factor Fumarate-Nitrate Reduction (FNR). J. Biol. Chem. 2004, 279, 9278–9286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lobato, L.; Bouzhir-Sima, L.; Yamashita, T.; Wilson, M.T.; Vos, M.H.; Liebl, U. Dynamics of the heme-binding bacterial gas-sensing dissimilative nitrate respiration regulator (DNR) and activation barriers for ligand binding and escape. J. Biol. Chem. 2014, 289, 26514–26524. [Google Scholar] [CrossRef] [Green Version]
- Raivio, T.L. MicroReview: Envelope stress responses and Gram-negative bacterial pathogenesis. Mol. Microbiol. 2005, 56, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Humphreys, S.; Rowley, G.; Stevenson, A.; Anjum, M.F.; Woodward, M.J.; Gilbert, S.; Kormanec, J.; Roberts, M. Role of the Two-Component Regulator CpxAR in the Virulence of Salmonella enterica Serotype Typhimurium. Infect. Immun. 2004, 72, 4654–4661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humphreys, S.; Stevenson, A.; Bacon, A.; Weinhardt, A.B.; Roberts, M. The Alternative Sigma Factor, ς E, Is Critically Important for the Virulence of Salmonella typhimurium. Infect. Immun. 1999, 67, 1560–1568. [Google Scholar] [CrossRef]
- Fang, F.C.; Libby, S.J.; Buchmeier, N.A.; Loewen, P.C.; Switala, J.; Harwood, J.; Guiney, D.G. The alternative sigma factor katF (rpoS) regulates Salmonella virulence. Proc. Natl. Acad. Sci. USA 1992, 89, 11978–11982. [Google Scholar] [CrossRef] [Green Version]
- Shinagawa, H. SOS response as an adaptive response to DNA damage in prokaryotes. In Stress-Inducible Cellular Responses; Birkhäuser: Basel, Switzerland, 1996; pp. 221–235. [Google Scholar] [CrossRef]
- Waldor, M.K.; Friedman, D.I. Phage regulatory circuits and virulence gene expression. Curr. Opin. Microbiol. 2005, 8, 459–465. [Google Scholar] [CrossRef]
- Njoroge, J.W.; Nguyen, Y.; Curtis, M.M.; Moreira, C.G.; Sperandio, V. Virulence Meets Metabolism: Cra and KdpE Gene Regulation in Enterohemorrhagic Escherichia coli. mBio 2012, 3, e00280-12. [Google Scholar] [CrossRef] [Green Version]
- Yoon, H.; McDermott, J.E.; Porwollik, S.; McClelland, M.; Heffron, F. Coordinated Regulation of Virulence during Systemic Infection of Salmonella enterica Serovar Typhimurium. PLoS Pathog. 2009, 5, e1000306. [Google Scholar] [CrossRef] [Green Version]
- Troxell, B.; Sikes, M.L.; Fink, R.C.; Vazquez-Torres, A.; Jones-Carson, J.; Hassan, H.M. Fur Negatively Regulates hns and Is Required for the Expression of HilA and Virulence in Salmonella enterica Serovar Typhimurium. J. Bacteriol. 2011, 193, 497–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patzer, S.I.; Hantke, K. The Zinc-responsive Regulator Zur and Its Control of theznu Gene Cluster Encoding the ZnuABC Zinc Uptake System in Escherichia coli. J. Biol. Chem. 2000, 275, 24321–24332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinkel, T.L.; Roux, C.M.; Dunman, P.M.; Fang, F.C. The Staphylococcus aureus SrrAB Two-Component System Promotes Resistance to Nitrosative Stress and Hypoxia. mBio 2013, 4, e00696-13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Durand, S.; Braun, F.; Lioliou, E.; Romilly, C.; Helfer, A.-C.; Kuhn, L.; Quittot, N.; Nicolas, P.; Romby, P.; Condon, C. A Nitric Oxide Regulated Small RNA Controls Expression of Genes Involved in Redox Homeostasis in Bacillus subtilis. PLoS Genet. 2015, 11, e1004957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarez, A.F.; Rodriguez, C.; Georgellis, D. Ubiquinone and Menaquinone Electron Carriers Represent the Yin and Yang in the Redox Regulation of the ArcB Sensor Kinase. J. Bacteriol. 2013, 195, 3054–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sardiwal, S.; Kendall, S.L.; Movahedzadeh, F.; Rison, S.C.G.; Stoker, N.G.; Djordjevic, S. A GAF Domain in the Hypoxia/NO-inducible Mycobacterium tuberculosis DosS Protein Binds Haem. J. Mol. Biol. 2005, 353, 929–936. [Google Scholar] [CrossRef]
- Karlinsey, J.E.; Maguire, M.E.; Becker, L.A.; Crouch, M.-L.V.; Fang, F.C. The phage shock protein PspA facilitates divalent metal transport and is required for virulence of Salmonella enterica sv. Typhimurium. Mol. Microbiol. 2010, 78, 669–685. [Google Scholar] [CrossRef] [Green Version]
- Pagels, M.; Fuchs, S.; Pané-Farré, J.; Kohler, C.; Menschner, L.; Hecker, M.; McNamarra, P.J.; Bauer, M.C.; Von Wachenfeldt, C.; Liebeke, M.; et al. Redox sensing by a Rex-family repressor is involved in the regulation of anaerobic gene expression in Staphylococcus aureus. Mol. Microbiol. 2010, 76, 1142–1161. [Google Scholar] [CrossRef] [Green Version]
- Darwin, A.J.; Stewart, V. Nitrate and Nitrite Regulation of the Fnr-dependentaeg-46.5Promoter ofEscherichia coliK-12 is Mediated by Competition Between Homologous Response Regulators (NarL and NarP) for a Common DNA-binding Site. J. Mol. Biol. 1995, 251, 15–29. [Google Scholar] [CrossRef]
- Noriega, C.E.; Lin, H.-Y.; Chen, L.-L.; Williams, S.B.; Stewart, V. Asymmetric cross-regulation between the nitrate-responsive NarX-NarL and NarQ-NarP two-component regulatory systems from Escherichia coli K-12. Mol. Microbiol. 2010, 75, 394–412. [Google Scholar] [CrossRef] [Green Version]
- Kondoh, H.; Ball, C.B.; Adler, J. Identification of a methyl-accepting chemotaxis protein for the ribose and galactose chemoreceptors of Escherichia coli. Proc. Natl. Acad. Sci. USA 1979, 76, 260–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greer-Phillips, S.E.; Alexandre, G.; Taylor, B.L.; Zhulin, I.B. Aer and Tsr guide Escherichia coli in spatial gradients of oxidizable substrates. Microbiology 2003, 149, 2661–2667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Falconi, M.; Colonna, B.; Prosseda, G.; Micheli, G.; Gualerzi, C.O. Thermoregulation of Shigella and Escherichia coli EIEC pathogenicity. A temperature-dependent structural transition of DNA modulates accessibility of virF promoter to transcriptional repressor H-NS. EMBO J. 1998, 17, 7033–7043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, S.; Goldberg, M.D.; Olsson, T.; Esposito, D.; Hinton, J.C.D.; Ladbury, J.E. H-NS is a part of a thermally controlled mechanism for bacterial gene regulation. Biochem. J. 2005, 391, 203–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamp, H.D.; Higgins, D.E. A Protein Thermometer Controls Temperature-Dependent Transcription of Flagellar Motility Genes in Listeria monocytogenes. PLoS Pathog. 2011, 7, e1002153. [Google Scholar] [CrossRef]
- Prugnola, A.; Arico, B.; Manetti, R.; Rappuoli, R.; Scarlato, V. Response of the bvg regulon of Bordetella pertussis to different temperatures and short-term temperature shifts. Microbiology 1995, 141, 2529–2534. [Google Scholar] [CrossRef] [Green Version]
- Herbst, K.; Bujara, M.; Heroven, A.K.; Opitz, W.; Weichert, M.; Zimmermann, A.; Dersch, P. Intrinsic Thermal Sensing Controls Proteolysis of Yersinia Virulence Regulator RovA. PLoS Pathog. 2009, 5, e1000435. [Google Scholar] [CrossRef]
- Quade, N.; Mendonca, C.; Herbst, K.; Heroven, A.K.; Ritter, C.; Heinz, D.W.; Dersch, P. Structural Basis for Intrinsic Thermosensing by the Master Virulence Regulator RovA of Yersinia. J. Biol. Chem. 2012, 287, 35796–35803. [Google Scholar] [CrossRef] [Green Version]
- Reines, A.E.; Deller, A.T. Parsec-scale radio emission from the low-luminosity active galactic nucleus in the dwarf starburst galaxy Henize 2–10. Astrophys. J. 2012, 750, L24. [Google Scholar] [CrossRef] [Green Version]
- Chakraborty, S.; Mizusaki, H.; Kenney, L.J. A FRET-Based DNA Biosensor Tracks OmpR-Dependent Acidification of Salmonella during Macrophage Infection. PLoS Biol. 2015, 13, e1002116. [Google Scholar] [CrossRef] [Green Version]
- Marcus, E.A.; Sachs, G.; Wen, Y.; Scott, D.R. Phosphorylation-dependent and Phosphorylation-independent Regulation ofHelicobacter pyloriAcid Acclimation by the ArsRS Two-component System. Helicobacter 2015, 21, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gore, A.L.; Payne, S.M. CsrA and Cra Influence Shigella flexneri Pathogenesis. Infect. Immun. 2010, 78, 4674–4682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Converse, P.D.; DeShon, R.P. A tale of two tasks: Reversing the self-regulatory resource depletion effect. J. Appl. Psychol. 2009, 94, 1318–1324. [Google Scholar] [CrossRef] [PubMed]
- Boyd, J.; Oza, M.N.; Murphy, J.R. Molecular cloning and DNA sequence analysis of a diphtheria tox iron-dependent regulatory element (dtxR) from Corynebacterium diphtheriae. Proc. Natl. Acad. Sci. USA 1990, 87, 5968–5972. [Google Scholar] [CrossRef] [Green Version]
- Lieser, S.A.; Davis, T.C.; Helmann, J.D.; Cohen, S.M. DNA-Binding and Oligomerization Studies of the Manganese(II) Metalloregulatory Protein MntR from Bacillus subtilis. Biochemistry 2003, 42, 12634–12642. [Google Scholar] [CrossRef]
- Banerjee, G.; Basak, S.; Roy, T.; Chattopadhyay, P. Intrinsic role of bacterial secretion systems in phylogenetic niche conservation of Bradyrhizobium spp. FEMS Microbiol. Ecol. 2019, 95, fiz165. [Google Scholar] [CrossRef]
- Fang, F.C.; Frawley, E.R.; Tapscott, T.; Vázquez-Torres, A. Bacterial Stress Responses during Host Infection. Cell Host Microbe 2016, 20, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Lage, C.; Leitao, A. Membrane permeability and sensitivity to lethal heat are affected by lexA and recA mutations in Escherichia coli K12. Braz. J. Med. Biol. Res. Rev. Bras. De Pesqui. Med. E Biol. 1994, 27, 2383–2389. [Google Scholar]
- Mozaheb, N.; Mingeot-Leclercq, M.-P. Membrane vesicle production as a bacterial defense against stress. Front. Microbiol. 2020, 11, 600221. [Google Scholar] [CrossRef]
- Chen, J.; Li, S.; Luo, J.; Zhang, Y.; Ding, W. Graphene oxide induces toxicity and alters energy metabolism and gene expression in Ralstonia solanacearum. J. Nanosci. Nanotechnol. 2017, 17, 186–195. [Google Scholar] [CrossRef]
- Tomlinson, D.L.; Elphinstone, J.G.; Soliman, M.Y.; Hanafy, M.S.; Shoala, T.M.; Abd El-Fatah, H.; Agag, S.H.; Kamal, M.; Abd El-Aliem, M.M.; Fawzi, F.G.; et al. Recovery of Ralstonia solanacearum from canal water in traditional potato-growing areas of Egypt but not from designated Pest-Free Areas (PFAs). Eur. J. Plant Pathol. 2009, 125, 589–601. [Google Scholar] [CrossRef]
- Wood, J.M. Osmotic stress. In Bacterial Stress Responses, 2nd ed.; Storz, G., Hengge, R., Eds.; Wiley: Hoboken, NJ, USA, 2010; pp. 133–156. [Google Scholar] [CrossRef]
- MacIntyre, A.M.; Barth, J.X.; Pellitteri Hahn, M.C.; Scarlett, C.O.; Genin, S.; Allen, C. Trehalose synthesis contributes to osmotic stress tolerance and virulence of the bacterial wilt pathogen Ralstonia solanacearum. Mol. Plant-Microbe Interact. 2020, 33, 462–473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, M.; Shah Jahan, M.; Wang, Y.; Sun, J.; Shu, S.; Guo, S. Compost amendments based on vinegar residue promote tomato growth and suppress bacterial wilt caused by Ralstonia Solanacearum. Pathogens 2020, 9, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, L.; Wei, Z.; Li, S.; Xiao, R.; Xu, Q.; Ran, Y.; Ding, W. Plant secondary metabolite, daphnetin reduces extracellular polysaccharides production and virulence factors of Ralstonia solanacearum. Pestic. Biochem. Physiol. 2021, 179, 104948. [Google Scholar] [CrossRef]
- Poole, R.K. Nitric oxide and nitrosative stress tolerance in bacteria. Biochem. Soc. Trans. 2005, 33, 176–180. [Google Scholar] [CrossRef]
- Truchon, A.N.; Hendrich, C.G.; Bigott, A.F.; Dalsing, B.L.; Allen, C. NorA, HmpX, and NorB cooperate to reduce NO toxicity during denitrification and plant pathogenesis in Ralstonia solanacearum. Microbiol. Spectr. 2022, 10, e0026422. [Google Scholar] [CrossRef]
- Jeong, K.C.; Hung, K.F.; Baumler, D.J.; Byrd, J.J.; Kaspar, C.W. Acid stress damage of DNA is prevented by Dps binding in Escherichia coliO157:H7. BMC Microbiol. 2008, 8, 181. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.E.; Barea, J.-M.; McNeill, A.M.; Prigent-Combaret, C. Acquisition of phosphorus and nitrogen in the rhizosphere and plant growth promotion by microorganisms. Plant Soil 2009, 321, 305–339. [Google Scholar] [CrossRef]
- Henard, C.A.; Tapscott, T.; Crawford, M.A.; Husain, M.; Doulias, P.-T.; Porwollik, S.; Liu, L.; McClelland, M.; Ischiropoulos, H.; Vázquez-Torres, A. The 4-cysteine zinc-finger motif of the RNA polymerase regulator DksA serves as a thiol switch for sensing oxidative and nitrosative stress. Mol. Microbiol. 2014, 91, 790–804. [Google Scholar] [CrossRef] [Green Version]
- Brown, D.G.; Allen, C. Ralstonia solanacearum genes induced during growth in tomato: An inside view of bacterial wilt. Mol. Microbiol. 2004, 53, 1641–1660. [Google Scholar] [CrossRef]
- Castillo, J.A.; Greenberg, J.T. Evolutionary Dynamics of Ralstonia solanacearum. Appl. Environ. Microbiol. 2007, 73, 1225–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feil, E.J.; Spratt, B.G. Recombination and the Population Structures of Bacterial Pathogens. Annu. Rev. Microbiol. 2001, 55, 561–590. [Google Scholar] [CrossRef] [PubMed]
- Puigvert, M.; Guarischi-Sousa, R.; Zuluaga, P.; Coll, N.S.; Macho, A.P.; Setubal, J.C.; Valls, M. Transcriptomes of Ralstonia solanacearum during root colonization of Solanum commersonii. Front. Plant Sci. 2017, 8, 370. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Li, S.; Li, F.; Yang, H.; Yang, F.; Xiang, J. Characterization of two types of vascular endothelial growth factor from Litopenaeus vannamei and their involvements during WSSV infection. Fish Shellfish. Immunol. 2015, 47, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; Kim, H.B. Protein interaction network related to Helicobacter pylori infection response. World J. Gastroenterol. 2009, 15, 4518. [Google Scholar] [CrossRef] [Green Version]
- Marchadier, E.; Carballido-López, R.; Brinster, S.; Fabret, C.; Mervelet, P.; Bessières, P.; Noirot-Gros, M.-F.; Fromion, V.; Noirot, P. An expanded protein-protein interaction network in Bacillus subtilis reveals a group of hubs: Exploration by an integrative approach. Proteomics 2011, 11, 2981–2991. [Google Scholar] [CrossRef]
- Van Gijsegem, F.; Genin, S.; Boucher, C. Conservation of secretion pathways for pathogenicity determinants of plant and animal bacteria. Trends Microbiol. 1993, 1, 175–180. [Google Scholar] [CrossRef]
- Sikdar, R.; Simmons, A.R.; Doerrler, W.T. Multiple envelope stress response pathways are activated in an Escherichia coli strain with mutations in two members of the DedA membrane protein family. J. Bacteriol. 2013, 195, 12–24. [Google Scholar] [CrossRef] [Green Version]
- da Cruz Nizer, W.S.; Inkovskiy, V.; Versey, Z.; Strempel, N.; Cassol, E.; Overhage, J. Oxidative Stress Response in Pseudomonas aeruginosa. Pathogens 2021, 10, 1187. [Google Scholar] [CrossRef]
- González, E.T.; Brown, D.G.; Swanson, J.K.; Allen, C. Using the Ralstonia solanacearum Tat Secretome To Identify Bacterial Wilt Virulence Factors. Appl. Environ. Microbiol. 2007, 73, 3779–3786. [Google Scholar] [CrossRef] [Green Version]
- Sarvas, M. Post-translocational folding of secretory proteins in Gram-positive bacteria. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2004, 1694, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Wickner, S. Posttranslational Quality Control: Folding, Refolding, and Degrading Proteins. Science 1999, 286, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, J.A.; Frost, N.A.; Hoppert, M.; Fernandes, P.J.; Vogt, S.L.; Raivio, T.L.; Blanpied, T.A.; Donnenberg, M.S. Outer membrane targeting, ultrastructure, and single molecule localization of the enteropathogenic Escherichia coli type IV pilus secretin BfpB. J. Bacteriol. 2012, 194, 1646–1658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrich, C.G.; Truchon, A.N.; Dalsing, B.L.; Allen, C. Nitric Oxide Regulates the Ralstonia solanacearum Type 3 Secretion System. Microbiol. Spectr. 2022, 10, e00264-22. [Google Scholar]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2018, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Pramanik, K.; Ghosh, P.K.; Ray, S.; Sarkar, A.; Mitra, S.; Maiti, T.K. An in silico structural, functional and phylogenetic analysis with three dimensional protein modeling of alkaline phosphatase enzyme of Pseudomonas aeruginosa. J. Genet. Eng. Biotechnol. 2017, 15, 527–537. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P. Paleontological statistics software package for education and data analysis. Paleontol. Electron. 2001, 4, 1–9. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No. | Regulator | Stress Response | Reported Candidates | References |
|---|---|---|---|---|
| 1 | OxyR | Oxidative stress | Salmonella enterica | [10] |
| 2 | DksA | Oxidative stress | - | [11] |
| 3 | NsrR | NO | S. enterica | [12] |
| 4 | FNR | Oxygen limitation | E. coli | [13] |
| 5 | DNR | Nitrosative stress | - | [14] |
| 6 | Bae | Membrane stress | - | [15] |
| 7 | Cpx | Misfolded OMPs at acid pH | S. enterica | [16] |
| 8 | Rcs | Membrane stress | - | [15] |
| 9 | σE | Membrane stress | S. enterica | [17] |
| 10 | RseA | Membrane stress | - | [18] |
| 11 | DegS | Membrane stress | - | [18] |
| 12 | RecA | DNA damage | - | [19] |
| 13 | LexA | DNA damage | Vibrio cholerae | [20] |
| 14 | FruR | Carbohydrate starvation | E. coli | [21] |
| 15 | SpoT | Amino acid starvation | S. enterica | [22] |
| 16 | FUR | Iron starvation | S. enterica | [23] |
| 17 | ZUR | Zinc starvation | S. enterica | [24] |
| 18 | SrrA/SrrB | Oxygen limitation | S. enterica | [25] |
| 19 | ResD/ResE | Oxygen limitation | S. enterica | [26] |
| 20 | ArcB/ArcA | Oxygen limitation | E. coli | [27] |
| 21 | DosR/DosS | Oxygen limitation | M. tuberculosis | [28] |
| 22 | Psp | Envelop stress | S. enterica | [29] |
| 23 | Rex | Oxygen limitation | S. aureus | [30] |
| 24 | NarQ/NarP | Oxygen limitation | S. enterica | [31] |
| 25 | NarX/NarL | Oxygen limitation | E. coli | [32] |
| 26 | Trg | Oxygen limitation | - | [33] |
| 27 | Tsr | Oxygen limitation | E. coli | [34] |
| 28 | Aer | Oxygen limitation | E. coli | [34] |
| 29 | H-NS | Temperature | S. flexneri | [35] |
| 30 | Hha | Temperature | E. coli | [36] |
| 31 | GmaR | Temperature | L. monocytogenes | [37] |
| 32 | MogR | Temperature | L. monocytogenes | [37] |
| 33 | BvgA/BvgS | Temperature | B. pertussis | [38] |
| 34 | RovA | Temperature | Y. pestis | [39] |
| 35 | SlyA | Temperature | S. enterica | [40] |
| 36 | PhoP/PhoQ | Temperature | Y. enterocolitica | [41] |
| 37 | PmrA/PmrB | Temperature | Y. enterocolitica | [41] |
| 38 | OmpR | Envelope | S. enterica | [42] |
| 39 | Dps | Nutrient limitation | R. solanacearum | [7] |
| 40 | ArsA | Acid pH | H. pylori | [43] |
| 41 | BvgA | Temperature regulates | B. pertussis | [38] |
| 42 | CsrA | Starvation | S. flexneri | [44] |
| 43 | DosR (NosR) | Hypoxia/NO. | M. tuberculosis | [45] |
| 44 | DtxR | Iron deprivation | C. diphtheriae | [46] |
| 45 | MntR | Manganese starvation | Bacillus subtilis | [47] |
| 46 | GmaR | Temperature | Listeria monocytogenes | [37] |
| 47 | σS | Starvation | S. enterica | [18] |
| Sl. No. | SRG | KO No. | Function | Cluster |
|---|---|---|---|---|
| 1 | nlpD | K06194 | Lipoprotein NlpD | (GenBank) putative outer membrane metallopeptidase lipoprotein nlpD/ | MSR |
| 2 | degQ | K04772 | Serine protease DegQ (EC:3.4.21.-) | (GenBank) putative serine protease do-like precursor (degP) | |
| 3 | rseB | K03598 | Sigma-E factor negative regulatory protein RseB | (GenBank) rseB; Sigma-E factor regulatory protein | |
| 4 | rseA | K03597 | Sigma-E factor negative regulatory protein RseA | (GenBank) rseA; Sigma-E factor negative regulatory protein | |
| 5 | rpoE | K03088 | RNA polymerase sigma-70 factor, ECF subfamily (misfolded OMPs at acid pH; required for virulence) | (GenBank) rpoE | |
| 6 | lexA | K01356 | Repressor LexA (EC:3.4.21.88) (Regulates toxin production) | (GenBank) lexA | |
| 7 | recX | K03565 | Regulatory protein (Modulates recA activity) | (GenBank) recX | |
| 8 | recA | K03553 | Catalyzes the hydrolysis of ATP in the presence of single-stranded DNA, RecA | (GenBank) recAn | |
| 9 | baeR | K07664 | Two-component system, OmpR family, response regulator BaeR for mdtABCD and acrD | (GenBank) baeR | OSR |
| 10 | baeS | K07642 | Two-component system, OmpR family, sensor histidine kinase BaeS (EC:2.7.13.3) | (GenBank) baeS | |
| 11 | ompR | K07659 | Two-component system, OmpR family, phosphate regulon response regulator OmpR | (GenBank) ompR | |
| 12 | ompR | K07638 | Two-component system, OmpR family (EC:2.7.13.3) | (GenBank) putative sensory histidine kinase in Two-component regulatory system with OmpR | |
| 13 | envZ | K07638 | Two-component system, OmpR family, osmolarity sensor histidine kinase EnvZ (EC:2.7.13.3) | (GenBank) envZ | |
| 14 | rcsB | K07687 | Two-component system, NarL family, captular synthesis response regulator RcsB | (GenBank) putative response regulator receiver | |
| 15 | Kata | K03781 | Catalase (EC:1.11.1.6) | (GenBank) kata | OxSR |
| 16 | katB | K03781 | Catalase [EC:1.11.1.6] (Paraquat-inducible catalase isozyme B) | (GenBank) katB | |
| 17 | katG | K03782 | Catalase-peroxidase (EC:1.11.1.21) | (GenBank) katG | |
| 18 | oxyR | K04761 | LysR family transcriptional regulator, hydrogen peroxide-inducible genes activator | (GenBank) oxyR | |
| 19 | sodB | K04564 | Superoxide dismutase, Fe-Mn family (EC:1.15.1.1) | (GenBank) sodB | |
| 20 | sodC | K04565 | Superoxide dismutase, Cu-Zn family (EC:1.15.1.1) | (GenBank) sodC | |
| 21 | Dps | K04047 | Starvation-inducible DNA-binding protein | (GenBank) putative DNA-binding protein | |
| 22 | nsrR | K13771 | Rrf2 family transcriptional regulator, nitric oxide-sensitive transcriptional repressor (regulates genes required for virulence) | (GenBank) nsrR | NxSR |
| 23 | norB | K04561 | Nitric oxide reductase subunit B (transmembrane protein) (EC:1.7.2.5) | (GenBank) norB | |
| 24 | aniA | K00368 | Nitrite reductase (NO-forming) (EC:1.7.2.1) | (GenBank) ainA | |
| 25 | Fnr | K01420 | CRP/FNR family transcriptional regulator, anaerobic regulatory protein | (GenBank) fnr | DdSR |
| 26 | Dnr | K21563 | CRP/FNR family transcriptional regulator, dissimilatory nitrate respiration regulator | (GenBank) putative transcription regulator protein | |
| 27 | dskA | K06204 | DnaK suppressor protein | (GenBank) dskA | |
| 28 | rpoZ | K03060 | DNA-directed RNA polymerase subunit omega (EC:2.7.7.6) | (GenBank) rpoZ | |
| 29 | rpoS | K03087 | RNA polymerase nonessential primary-like sigma factor (regulates expression of plasmid virulence genes) | (GenBank) rpoS | |
| 30 | rpoB | K03043 | DNA-directed RNA polymerase subunit beta (EC:2.7.7.6) | (GenBank) RNA polymerase, beta subunit | |
| 31 | rpoA | K03040 | DNA-directed RNA polymerase subunit alpha (EC:2.7.7.6) | (GenBank) RNA polymerase, alpha subunit | |
| 32 | polA | K02335 | DNA polymerase I (EC:2.7.7.7) | (GenBank) polA | |
| 33 | h-ns | K03746 | DNA-binding protein H-NS | (GenBank) Histone-like nucleoid-structuring protein H-NS | |
| 34 | Fur | K03711 | Fur family transcriptional regulator, ferric uptake regulator | (GenBank) fur | |
| 35 | Gmk | K00942 | Guanylate kinase | (GenBank) gmk |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banerjee, G.; Quan, F.-S.; Mondal, A.K.; Sur, S.; Banerjee, P.; Chattopadhyay, P. Interrelation between Stress Management and Secretion Systems of Ralstonia solanacearum: An In Silico Assessment. Pathogens 2022, 11, 730. https://doi.org/10.3390/pathogens11070730
Banerjee G, Quan F-S, Mondal AK, Sur S, Banerjee P, Chattopadhyay P. Interrelation between Stress Management and Secretion Systems of Ralstonia solanacearum: An In Silico Assessment. Pathogens. 2022; 11(7):730. https://doi.org/10.3390/pathogens11070730
Chicago/Turabian StyleBanerjee, Goutam, Fu-Shi Quan, Amit Kumar Mondal, Shantanu Sur, Pratik Banerjee, and Pritam Chattopadhyay. 2022. "Interrelation between Stress Management and Secretion Systems of Ralstonia solanacearum: An In Silico Assessment" Pathogens 11, no. 7: 730. https://doi.org/10.3390/pathogens11070730