
Manipulating Microbiota to Treat Atopic Dermatitis: Functions and Therapies
by
 , , ,
, , ,
Md Jahangir Alam
1,† ,
,
Liang Xie
1,2,†,
Yu-Anne Yap
3,
Francine Z. Marques
2,4 and
Remy Robert
3,* 1
Department of Microbiology, Biomedicine Discovery Institute, Monash University, Clayton, VIC 3800, Australia
2
Hypertension Research Laboratory, School of Biological Sciences, Monash University, Clayton, VIC 3800, Australia
3
Department of Physiology, Biomedicine Discovery Institute, Monash University, Clayton, VIC 3800, Australia
4
Heart Failure Research Laboratory, Baker Heart and Diabetes Institute, Melbourne, VIC 3004, Australia
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Pathogens 2022, 11(6), 642; https://doi.org/10.3390/pathogens11060642
Submission received: 11 May 2022
/
Revised: 27 May 2022
/
Accepted: 29 May 2022
/
Published: 2 June 2022
(This article belongs to the Special Issue Host-Microbe Interaction in Atopic Diseases)
Abstract
:Atopic dermatitis (AD) is a globally prevalent skin inflammation with a particular impact on children. Current therapies for AD are challenged by the limited armamentarium and the high heterogeneity of the disease. A novel promising therapeutic target for AD is the microbiota. Numerous studies have highlighted the involvement of the skin and gut microbiota in the pathogenesis of AD. The resident microbiota at these two epithelial tissues can modulate skin barrier functions and host immune responses, thus regulating AD progression. For example, the pathogenic roles of Staphylococcus aureus in the skin are well-established, making this bacterium an attractive target for AD treatment. Targeting the gut microbiota is another therapeutic strategy for AD. Multiple oral supplements with prebiotics, probiotics, postbiotics, and synbiotics have demonstrated promising efficacy in both AD prevention and treatment. In this review, we summarize the association of microbiota dysbiosis in both the skin and gut with AD, and the current knowledge of the functions of commensal microbiota in AD pathogenesis. Furthermore, we discuss the existing therapies in manipulating both the skin and gut commensal microbiota to prevent or treat AD. We also propose potential novel therapies based on the cutting-edge progress in this area.
1. Introduction
Atopic dermatitis (AD), also known as eczema, is a skin inflammation that exhibits chronic, persistent, pruritic lesions, and is often associated elevated levels of IgE [1,2]. AD affects 10–20% of the population during their lifetime in developed countries, with a particularly high prevalence among children [2,3]. Its prevalence is also rapidly increasing in developing nations [4,5]. The established pathogenesis of AD involves the initiation of barrier disruption, followed by the activation of type 2 (TH2) immune responses [2,6]. Variants in the filaggrin (FLG), the gene encoding an important skin barrier protein, represent a significant risk factor for AD [7,8,9]. AD is often associated with the development of asthma and food allergies, which is known as “atopic march” [10]. Physical therapies that moisturize the skin, preventing water loss, controlling xerosis, and relieving barrier disruptions, are recommended for AD patients [11]. While corticosteroids remain the standard anti-inflammatory treatment against AD, the efficacy of blocking TH2 responses is recognized by various clinical trials [12,13,14,15,16,17,18,19]. Unfortunately, the limited armamentarium [20] and the high heterogeneity of the disease [21] make the management of AD challenging. Therefore, novel therapeutic strategies needed for AD treatment.
The past two decades have highlighted the role of commensal microbiota in health homeostasis and disease [22]. As the largest and outmost organ of the body, the human skin has been estimated to host about 1 billion bacteria per 1 cm2 area [23]. Conversely, the gastrointestinal tract harbors the largest microbiota population in our body, exceeding 1014 bacterial cells [24,25]. AD is probably the most well-characterized disease in which skin dysbiosis plays a causal role [6]. In addition, AD is also associated with gut dysbiosis [26]. The investigations from the pre-clinical animal studies and the emerging human 3D skin models furthered our understanding on the complex interplay between the microbiota and the AD context [27,28]. In this review, we comprehensively summarize the roles of both the skin and gut microbiota in AD and the current approaches aimed at manipulating commensal microbiota to prevent or treat AD. We also provide insights for future investigations to improve the efficacy of current agents and establish novel therapeutic strategies that leverage the microbiota.
2. The Skin Microbiota Alternation in AD Patients: A Particular Focus on Staphylococcus aureus
It is now well established that changes in the normal skin microbiota composition, a condition known as dysbiosis, contribute to the disruption of cutaneous immune homeostasis and promotes the development of skin diseases, including AD [29,30]. Studies that have demonstrated the association of AD with skin dysbiosis are summarized in Table 1. AD is often accompanied with dysbiosis, featured by increased colonization of staphylococcal species and decreased richness and diversity of other bacterial communities [31]. S. aureus, a dominant species among the family of Staphylococcae, can be 100 times more abundant in AD skin compared to normal healthy skin [32]. On the skin surface, S. aureus secretes virulence factors, including phenol-soluble modulins (PSMs) and proteases, thus disrupting normal skin barrier functions, which alter the epidermal environment favouring the development of AD [33,34,35,36,37]. We will discuss the mechanisms involved later in this review.
The increased abundance of S. aureus colonization in AD is associated with a depletion in the coagulase-negative staphylococcal species (CoNS), such as S. epidermidis, S. hominis, and other skin commensal bacterial communities, including Streptococcus salivarius, Propionibacterium, Streptococcus, Acinetobacter, Corynebacterium, Prevotella and Proteobacteria [31,38,39,40]. In contrast, these skin commensal microbiota produce antimicrobial substances that inhibit the growth of pathogenic S. aureus and its biofilm formation [30,41,42]. In the non-lesional skin samples of AD patients, CoNS are the dominant bacterial communities [38]. However, their abundances were lower on the skin of healthy individuals compared with non-lesional AD skin [38,43]. This suggests the existence of complex interactions between the different skin microbial communities, which modulate the host’s susceptibility to AD.

{kind=link}
{kind=link}
Table 1.
Summary of the studies demonstrating the dysbiosis of skin microbiota in AD.
| Year | Subjects, Numbers | Methods | Results (Alternations of Skin Microbiota) | Reference |
|---|---|---|---|---|
| 2012 | 11 Infants with AD and 12 healthy controls | 16S rRNA gene sequencing | AD infants: ↓S. salivarius and ↑S. aureus | [40] |
| 2012 | 12 Children with AD and 11 healthy controls | 16S rRNA gene sequencing | AD lesion: ↑S. aureus and ↑S. epidermidis | [31] |
| 2013 | 13 AD patients and 49 healthy controls | 16S rRNA gene sequencing | AD patients: ↑S. aureus, ↓-diversity | [44] |
| 2015 | 21 AD infants and 17 healthy controls | Real-time PCR analysis of skin scratches | AD infants: ↑S. aureus | [45] |
| 2016 | 128 AD patients (59 young children at 2–12 years, 13 teenagers at 13–17 years, and 56 adults at 18–62 years of age), 68 age-matched healthy controls (13 young children, 10 teenagers, 45 adults) | 16S rRNA gene sequencing | Children with AD: ↑Streptococcus, ↑Granulicatelle, ↑Gemella, ↑Rothia, ↑Haemophilus Adults with AD: ↑Propionibacterium, ↑Corynebacterium, ↑Staphylococcus, ↑Lactobacillus, ↑Finegoldia, ↑Anaerococcus | [46] |
| 2016 | Three male first cousins aged 50–53 years | 16S rRNA gene sequencing | AD patients: ↑S. aureus | [47] |
| 2017 | 10 AD infants, 10 age-matched healthy controls | 16S rRNA gene sequencing | AD infants: ↑Staphylococcus | [29] |
| 2017 | 49 AD patients and 30 non-AD subjects | 16S rRNA gene sequencing | AD patients: ↓S. epidermidis, ↓S. hominis | [30] |
| 2017 | 27 AD patients and 6 healthy controls | High-throughput pyrosequencing | AD patients: ↑Staphylococcus, ↑Pseudomonas, and ↑Streptococcus, ↓Alcaligenaceae (f), ↓Sediminibacterium, and ↓Lactococcus | [48] |
| 2018 | 10 AD patients and 10 healthy controls | 16S rRNA gene sequencing | AD patients: ↑S. aureus, ↓-diversity | [38] |
| 2019 | 91 AD patients, 134 psoriasis patients, and 126 healthy controls | 16S rRNA gene sequencing | AD patients: ↑S. aureus, ↓S. epidermidis and ↓Corynebacterium | [49] |
| 2019 | 172 AD patients and 120 healthy controls | 16S rRNA gene sequencing | AD patients: ↑Staphylococcus | [50] |
| 2020 | 11 AD patients | 16S rRNA gene sequencing | AD skin lesions: ↑S. aureus, ↓C. pseudogenitalium | [51] |
| 2020 | 67 AD patients and 28 healthy controls | 16S rRNA gene sequencing | AD skin lesion: ↑Staphylococcus (S. aureus and S. epidermidis), ↓Corynebacterium, ↓Micrococcus, ↓Cutibacterium and ↓Streptococcus | [52] |
| 2020 | 7 AD patients and 10 healthy controls | 16S rRNA gene sequencing, and Staphylococcus specific SLST sequencing | AD patients: ↑Staphylococcus, ↓Propionibacterium | [53] |
| 2021 | 28 AD patients and 14 healthy controls | 16S rRNA gene sequencing | AD patients: ↑S. aureus, ↓S. capitis and ↓Micrococcus sp. | [54] |
Legend: SLST, single-locus sequencing typing.
3. S. aureus in AD Pathogenesis
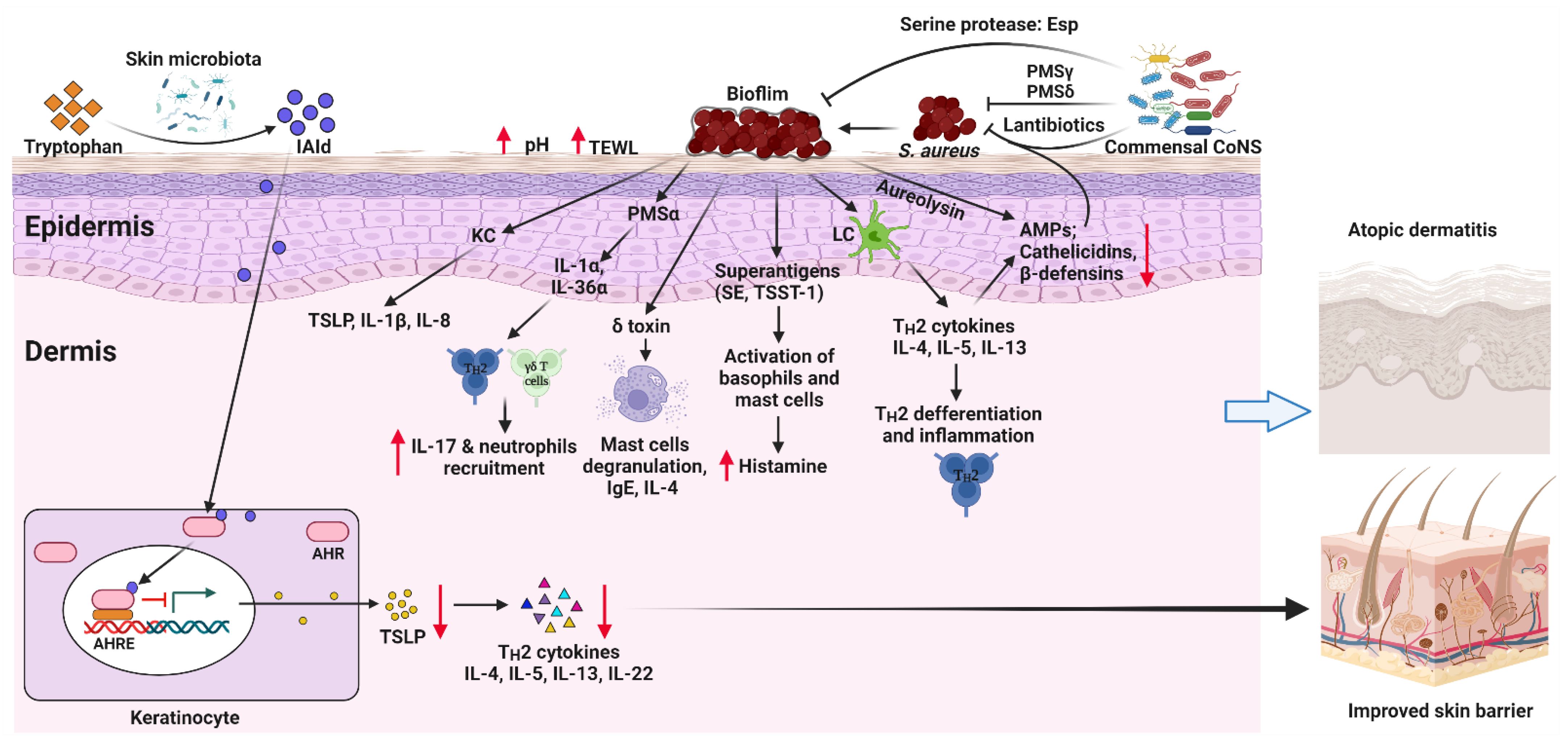
Probably the best understood association between skin microbiota and AD is the involvement of S. aureus. As previously discussed, S. aureus is commonly identified in the skin of AD patients [34,55,56]. S. aureus colonization precedes the onset of AD in infancy [57], and topical application of S. aureus isolates from AD skin suffices to induce AD-like skin inflammation in mice [27]. This evidence supports the causal role of S. aureus in AD pathogenesis. S. aureus adheres to AD skin biopsy specimens more efficiently compared to healthy ones because of the changes to the composition of the stratum corneum and corneocyte morphology [58,59]. The impairment of the skin barrier during AD facilitates the colonization of S. aureus [60]. Filaggrin deficiency is associated with elevated S. aureus in skin microbiota [43]. Reduced filaggrin expression promoted S. aureus colonization in a human 3D epidermal model [61]. TH2 cytokines such as IL-4 and IL-13 can suppress the production of antimicrobial peptides (AMPs) such as cathelicidin and β-defensins that can act against S. aureus, hence promoting its excessive expansion [62]. Additionally, S. aureus proteases cause epidermal disruption [33,63]. S. aureus stimulates keratinocytes to produce endogenous proteases, which exacerbate the barrier dysfunctions [37]. Furthermore, S. aureus produces a cysteine protease staphopain and a metalloprotease aureolysin that cleave and inactivate the AMPs [62,64].
S. aureus expresses a variety of virulence factors inducing inflammation related to AD [65,66,67,68]. For example, S. aureus expresses superantigens (SAgs), such as toxic shock syndrome toxin-1 (TSST-1) and the staphylococcal enterotoxin serotypes [67]. These SAgs bind to major histocompatibility class II (MHC-II) and stimulate the production of cytotoxic cytokines from T cells [69,70,71,72]. In addition, SAgs trigger IgE response, induce mast cell degranulation, and promote skin inflammation [73].
δ-toxin and α-toxin are two other major virulent factors secreted by S. aureus [66,68]. Enhanced by IgE, δ-toxin induces degranulation without lysis of murine mast cells [73]. S. aureus lacking δ-toxin was not able to induce skin inflammation characterized by elevated IL-4 and IgE in mice [73]. Phenol-soluble modulin (PSM) including PSMα and PSMβ are short amphipathic peptides expressed by S. aureus with similar functions as δ-toxin [68]. By opposition to δ-toxin, PSMα2 and PSMα3 cause mast cell death [73]. PSMα also induces proinflammatory cytokines production from keratinocytes, including IL-1α and IL-36α, which stimulates γδ T cells and elicit Th17 responses as well as neutrophil recruitment [36,74,75]. α-toxin is a pore-forming cytolysin, which can induce skin inflammation in mice [66,76]. The keratinocyte death caused by α-toxin directly exacerbates the barrier disruption, thus promoting AD-like inflammation [77]. The keratinocyte toxicity is enhanced by TH2 cytokines during AD. Meanwhile, under a healthy state, the expression of filaggrin and sphingomyelinase reduces keratinocyte susceptibility to α-toxin [78]. Protein A found on the cell wall of S. aureus binds to the tumour necrosis factor receptor 1 (TNFR1) and induces NF-κB and AP-1 activation and their downstream proinflammatory pathways in keratinocytes [79].
Furthermore, S. aureus can enter from the epidermis to the dermis, where it encounters immune cells and triggers the TH2 responses by inducing the production of IL-4, IL-13, IL-22, thymic stromal lymphopoietin (TSLP), and other cytokines associated with AD [63]. The pathogen-associated molecular patterns (PAMPs) from S. aureus predominantly bind to toll-like receptor 2 (TLR2) [80,81]. TLR2 signalling is a double-edged sword in AD pathogenesis. Although TLR2 activation stimulates TSLP production and mast cell degranulation [73,82,83], it can also be protective in AD by rapidly increasing the expression of tight junction protein claudin 1 and AMPs in differentiated epidermal layers from normal skin [84]. AD skin has impaired TLR2 activity, thus disrupting the normal immune response and skewing TH2 immunity [84,85].
4. Other AD Pathogeneses Regulated by the Skin Microbiota
In addition to the pathogenic role of S. aureus, homeostasis of skin microbiota is vital for normal skin barrier function, the cutaneous immune balance, and the elimination of possible pathogens, thus modulating AD pathogenesis [86]. Germ-free mice exhibited a significantly different skin transcriptome [87]. Germ-free mice displayed impaired skin barrier function, a direct consequence of the abnormal epithelial development and differentiation [88]. Treating a 3D skin tissue model with a mix of selected members of normal skin microbiome profoundly improved the barrier integrity of the tissue [89]. In this section, we discuss how the skin microbiota regulates two major aspects: (1) AMPs, and (2) the tryptophan metabolites-aryl hydrocarbon receptor (AHR) axis. Together with the pathogenic roles of S. aureus in AD, these mechanisms are summarized in Figure 1.
4.1. Antimicrobial Peptides
Using antimicrobial mechanisms [90], the skin barrier and its associated microbiota protect against pathogenic microorganisms such as S. aureus in AD. For example, the production of AMPs such as cathelicidin and β-defensin by the host and the microbiota inhibit certain microorganisms. It appears that normal commensal bacteria are more resistant to host AMPs [91], and host production of AMPs is regulated by commensal bacteria in poorly understood mechanisms [92,93,94]. Not only can commensal bacteria control the production of AMPs by the host, they also produce some antimicrobial agents able to suppress pathogenic competitors, thereby providing an additional antimicrobial barrier on the skin. For instance, S. epidermidis produces the peptides PSMγ and PSMδ, which inhibit the growth of pathogenic bacteria on the skin [95,96]. Similarly, S. hominis is another CoNS that produces lantibiotics, a class of cyclic AMPs containing lanthionine and methyllanthionine [30]. Skin dysbiosis likely leads to the dysregulation of the antimicrobial function against pathogenic bacteria. Impaired production of cathelicidin and β-defensins is responsible for the dominance of S. aureus in AD skin [62]. Therefore, the skin microbiota has an interaction with the host AMPs production and contributes to the control of AD.
4.2. Tryptophan Metabolites-AHR Axis
AHR is a ligand-activated transcription factor that modulates tissue homeostasis and immune responses [97,98,99]. AHR agonists range from xenobiotic chemicals to endogenous indole by-products derived from tryptophan metabolism [100,101,102]. The activation of AHR is mostly anti-inflammatory through the regulation of cytokine production and other immune-related transcription factors, such as NF-κB [97,98,99]. AHR is expressed by keratinocytes, epidermal Langerhans cells, dermal and epidermal innate and adaptive immune cells [103]. Although TH2 cells exhibited negligible AHR expression [104], activation of AHR in dendritic cells (DCs) suppressed TH2 differentiation [105,106]. The topical application of coal tar, which is a traditional therapy against AD, works by activating AHR [107]. In the skin epidermis, dead keratinocytes and broken keratin are used as substrates in the tryptophan metabolism of skin microbiota which leads to AHR agonists production [108]. The barrier dysfunction observed in germ-free mice skin is attributed to the attenuated AHR pathway [88]. Activation of AHR upregulates the expression of barrier-related molecules like filaggrin, loricrin and involucrin, by keratinocytes, thus maintaining the healthy epidermal barrier [109,110,111]. This can be achieved by the tryptophan metabolites derived from the skin microbiota [111,112,113,114]. For example, Indole-3-aldehyde (IAld), a skin microbiota-derived AHR agonist, suppresses TSLP expression and protects mice against MC903-induced AD. In humans, the skin of AD patients displays a lower level of IAld compared to that of their healthy counterparts [113]. AHR signalling is essential for AMPs production and can shape the skin microbiome [53], as demonstrated with AHR-deficient mice, which display a more variable and complex skin microbiota compared to WT controls [115].
5. Therapeutic Manipulation of the Skin Microbiota in the Management of AD
Reducing the abundance of pathogenic bacteria and restoring the normal microbial balance in the skin may be of clinical benefit for AD management. In animal models, antibiotic treatment established the causal role of skin dysbiosis in AD. Although a randomized clinical trial (RCT) has demonstrated that antibiotics suppressing the growth of S. aureus significantly attenuated AD [116], the clinical efficacy of antibiotics in this pathology remains questionable [117]. Skin bacteria abundantly reside in hair follicles, eccrine glands or beneath the epidermal barrier, where antibiotics are difficult to reach [63,118]. In addition, most antibiotics can (1) disrupt normal microbiota due to their lack of specificity and (2) favour antibiotic-resistant strains of S. aureus. Nevertheless, a phase III trial (NCT02840955) investigates the therapeutic potential of a bacteriophage endolysin (Staphefekt) in AD adult patients. Staphefekt is a specific bactericidal agent against S. aureus with unlikeliness of resistance and is effective against the methicillin-resistant S. aureus (MRSA) strains that have already acquired resistance to conventional antibiotics. Because bacterial killing by this endolysin is independent of the involvement of the bacterial metabolism [119,120,121].
The reintroduction of normal commensal bacteria represents a promising strategy for AD treatment. Topical application of antimicrobial CoNS strains able to produce specific AMPs against S. aureus, decreased colonization of this bacteria and effectively attenuated AD severity [30,122]. Another angle in manipulating skin microbiota is by reducing the skin surface pH. The acidic epidermal surface is essential for maintaining the skin barrier [123]. Acidity favours commensal bacteria and suppresses pathogenic bacteria, including S. aureus [124]. In healthy individuals, it has been observed that higher skin pH was associated with increased S. aureus skin colonization [125]. It is well known that AD lesions display higher pH values than healthy skin [126,127], and topical application of acids improved AD in murine models [128,129].
Current attempts to manipulate skin microbiota to control AD are limited and lack RCTs. However, as more causal roles of the skin microbiota in AD are being revealed, therapeutic manipulation of skin microbiota emerges as a promising way to improve the management of AD.
6. The Gut Microbiota Profiles in AD Patients
Aside from the skin microbiota, the gut microbiota, known to have a systemic impact on the host immune responses, is also closely associated with atopic diseases such as AD. A hypothesis of the “gut-skin” axis has been proposed and has opened new paths related to the prevention and treatment of AD [26]. Various studies demonstrated that AD is associated with gut dysbiosis, especially during early life (Table 2). AD patients display poor gut microbial diversity in several clinical trials [130,131,132,133,134,135,136,137,138], while contradictory results exist [139,140,141,142]. Similarly, to the skin, AD patients exhibit abundant S. aureus in their gut microbiota [143,144,145,146]. Other microbes associated with inflammation and epithelial damage, such as Clostridiodes difficile, and coliforms, including pathogenic Escherichia coli, are increased in the gut microbiota of AD patients [136,140,146,147,148,149,150,151,152]. Metagenomic analysis has shown that the gut microbes in AD patients carry extra genes associated with inflammatory responses and the breakdown of gut epithelial layers [139,141]. Breastfed infants have a lower risk of atopic diseases [153,154,155,156,157,158], likely because of the abundance of Bifidobacteria in their gut [159,160,161,162], a genus that is reduced in the gut microbiota of AD patients [151,163,164,165,166]. However, it has been reported that AD development is also associated with the higher abundance of certain species of Bifidobacteria, such as Bifidobacterium catenulatum, B. bifidum and B. pseudocatenulatum [167,168]. Short-chain fatty acids (SCFAs) are microbial metabolites well known for their anti-inflammatory effects. High fecal levels of SCFAs are significantly associated with a lower risk of AD [139,169]. The SCFA-producing bacteria Coprococcus eutactus is less abundant in the gut microbiota of AD patients [135]. Subspecies of Faecalibacterium prausnitzii are enriched in AD fecal samples. Although F. prausnitzii is a major SCFA producer in healthy subjects, this species is an inefficient SCFA producer in AD patients [139]. Furthermore, genes encoding carbohydrate active enzymes (CAZymes), which break down resistant starch into SCFAs, are deficient in the gut microbiota of AD patients [142]. While infantile gut dysbiosis may predict the development of AD [131,132,134,147,148,149,152], childhood AD history had prolonged imprints in gut microbiota that neonates with childhood AD history had a lower abundance of Bifidobacteria, Akkermansia, and Faecalibacteria compared to their healthy counterparts [170].
Despite these reports linking gut dysbiosis with AD, the actual roles of gut microbiota in AD development are still largely unknown. Establishing a causative role for the gut microbiota in the development of diseases remains challenging. An invaluable tool for this type of investigation is the use of germ-free animals. Germ-free mice showed similar disease severity in the oxazolone-induced ear atopic dermatitis model as conventional mice [171]. However, germ-free mice have higher levels of serum IgE and inflammatory cytokines, such as TNF and IL-6 in ear tissues, suggesting exacerbated skin inflammation [171]. Furthermore, the sensitivity to oxazolone is transferable to germ-free mice with fecal microbiota transplant (FMT) [171]. These findings suggest a causative role of gut microbiota in the development of AD. However, the direct link between gut microbiota and type 2 inflammatory responses in AD is still unknown.
7. The Gut Microbiota Regulates AD-Related Immune Responses and the Underlying Mechanisms
There is firm evidence supporting the role of gut microbiota-regulated immune responses in atopic diseases and AD [172]. For example, early colonization of Bifidobacteria subtly regulates the TH1/TH2 immune balance, reducing the risk of AD [173]. Most of the knowledge regarding this arises from the use of probiotics in different clinical trials, as discussed below. In this section, we focus on three aspects: (1) SCFAs-anti-inflammation axis, (2) tryptophan metabolites-AHR axis, and (3) toll-like receptor signalling (Figure 2). These pathways that can be modulated by gut microbiota are well characterized and highly relevant to AD and other allergic immune responses.
7.1. Short-Chain Fatty Acids and Their Anti-Inflammatory Effects
As briefly discussed above, the gut microbiota of AD patients contains less SCFA-producers or is defective in generating SCFAs [135,139,142]. SCFAs are microbial metabolites derived from dietary fibre [174]. The most abundant SCFAs found in the host are acetate (two carbons), propionate (three carbons), and butyrate (four carbons) [175]. Recent studies established a potent anti-inflammatory role of SCFAs against numerous inflammatory diseases, including atopic diseases [169,172,176,177,178]. In atopic models, the anti-inflammatory effects of SCFAs are promoted through the regulation of DCs leading to the inhibition of TH2 [179] and the activation of regulatory T cell (Treg) differentiation [180]. Subcutaneous injection and/or topical application of sodium butyrate attenuate the hapten-induced AD in mice by recruiting Tregs and inducing the production of the anti-inflammatory cytokine, IL-10 [181]. Still, the mechanisms underlying the protective effect of SCFAs in AD are poorly understood and insights from other atopic diseases would be invaluable.
The immune regulations by SCFAs are largely mediated by two key mechanisms: (1) the activation of certain G-protein coupled receptors (GPCRs) and (2) the inhibition of histone deacetylases (HDACs) [176].
7.1.1. SCFA-Sensing GPCRs
There are three GPCRs activated by SCFAs. GPR41 and GPR43 are exclusively activated by all three abundant SCFAs, while GPR109A can only be activated by butyrate [182,183]. The role of GPR41 and GPR109A in the immune cells remains elusive, while more information is available about GPR43, which is more widely expressed by the immune cells compared to the other two receptors [182,184]. Activation of GPR41 and GPR43 inhibits the TNF-induced or LPS-induced production of the pro-inflammatory cytokines IL-6 and IL-8 in human endothelial cells [185]. GPR43 was the first SCFA-sensing receptor to be deorphanized. It plays an anti-inflammatory role in various disease models, including atopic diseases [180,184,186,187,188,189]. A report demonstrated that T cells recruited to the skin after butyrate administration showed higher expression of GPR43 [181]. GPR43 is essential for the expansion of Tregs and the regulatory effects of Tregs induced by SCFAs [190,191]. Activation of GPR43 promotes the production of the anti-inflammatory cytokine IL-10 by CD4+ T cells [192]. Regulation of neutrophil biology, which is involved in AD [193,194,195,196,197], by SCFAs is also dependent on GPR43 activation [184,198].
GPR109A can regulate macrophages and DCs to promote colonic Treg development [199]. Moreover, GPR109A signalling inhibits the production of the pro-inflammatory cytokines, TNF, IL-6, CCL2, and IL-1β from monocytes, macrophages, adipocytes, and epithelial cells [200,201,202,203]. However, the role of GPR109A in regulating TH2 cytokine production, particularly in the AD context, remains unclear.
Conversely, SCFAs significantly reduce the luminal pH in the colon [204,205], thus inhibiting the survival of potential pathogens [206] and activating proton-sensing GPCRs of the host [172,182]. We recently reported that GPR65, a proton sensor highly expressed in various leukocyte subsets, plays a protective role in AD by inhibiting inflammatory cytokines production and immune cell migrations [207]. The polymorphisms resident in or near the human gene GPR65, rs3742704, and rs8005161 were significantly associated with the risk of AD and asthma [207,208].
7.1.2. HDAC Inhibition
HDACs are enzymes that remove acetyl groups from an ε-N-acetyl lysine amino acid on histones [209]. The histone deacetylation facilitates high-affinity binding between histone proteins and DNA, leading to DNA compaction [209], results in lower chromatin accessibility for transcription factors and prevents gene expression [209]. HDACs play a critical role in many pathologies, including atopic diseases, making HDAC inhibitors promising drug candidates against atopic diseases and AD [210]. High HDAC activity impairs tight junction function [211]. HDAC inhibitors such as trichostatin A (TSA) inhibit 2,4-dinitrofluorobenzene-induced dermatitis in mice [212,213]. A recent study demonstrates that belinostat, a pan HDAC inhibitor, can rescue the defective skin barrier in AD through restoring epidermal miR-335 expression [214]. miR-335 directly represses SOX6, which impairs epidermal differentiation [214]. However, a report also demonstrated that HDAC inhibited IL-4 production by human T cells, and TSA promoted IL-4 production [215].
SCFAs, particularly butyrate, are potent inhibitors of HDACs [216,217,218,219,220,221]. SCFAs are able to inhibit HDACs directly by entering the cells [222] and this inhibition is closely associated with GPCR signalling. The activation of GPR41 can suppress histone acetylation in Chinese hamster ovary cell lines, possibly through inhibiting HDACs [223]. HDAC inhibition in colonic Tregs by SCFAs may be dependent on GPR43 [190]. SCFAs restrict pro-inflammatory cytokine production through inhibiting HDAC activity. Acetate reduced HDAC activity in human monocytes in vitro, which correlated with lower production of IL-6, IL-8, and TNF [224]. SCFAs showed a similar effect as TSA in suppressing NF-κB activation and TNF production in human peripheral mononuclear cells, but HDAC activity was not tested [225]. Similarly, HDAC inhibition can partially mediate SCFAs’ inhibition of pro-inflammatory cytokine production by human endothelial cells [185]. Whether SCFAs regulate TH2 cytokine production through HDAC inhibition remains unknown.
Conversely, HDAC inhibition may underlie Treg promotion by SCFAs. HDAC inhibitors induce Foxp3 expression and promote Treg differentiation [226,227,228]. The most well-known HDAC regulating Treg differentiation and function is HDAC9 [228], while inhibition or depletion of HDAC10 and HDAC11 also enhances the immune-suppressive function of Tregs [229,230]. Hdac9−/− mice had ~50% more Foxp3+ Tregs in the lymphoid tissues, and Hdac9−/− Tregs are also more potent in suppressing effector T cells compared to WT controls [228]. Overall, SCFAs promote immunity and suppress inflammatory responses associated with AD.
7.2. Tryptophan, Its Metabolites and Aryl Hydrocarbon Receptor Signalling
Tryptophan is an essential amino acid in human. While L-tryptophan represents the proteinogenic enantiomer, its D-enantiomer, D-tryptophan, is a bacterial metabolite from Bifidobacterium, Lactobacillus and Lactococcus, which are known probiotics [231]. A recent study demonstrated that D-tryptophan expands the Treg population in the colon [231], and suppresses the expression of TH2-associated CCL17 by the human Hodgkin lymphoma T-cell line KM-H2 and the inflammatory activity of human DCs [231].
Meanwhile, by breaking down dietary L-tryptophan into indole, indole-3-acetic acid (IAA), indole-3-propionic acid (IPA), tryptamine (TA), and 3-methyl indole (skatole) [232,233], the gut microbiota also serves as a vital source of AHR agonists. A landmark study demonstrated that the AHR agonist IAld produced from the tryptophan metabolism of Lactobacillus reuteri activates AhR, induces IL-22 secretion, and promotes gut homeostasis [234]. This suggests that AHR serves as the bridge in the crosstalk between commensal bacteria and host health. Knowing that AHR agonists, such as IAld, have potent protection against AD, we hypothesize that AHR agonists derived from gut microbiota have distal protection for AD in the skin.
7.3. Toll-like Receptor Signalling
In addition to skin microbiota, gut microbiota also produces PAMPs recognized by TLRs, thus contributing to systemic immune homeostasis [235], including AD-related immune responses. As mentioned earlier, TLR2 and TLR4 play essential roles in maintaining the balance between TH1 and TH2 immune responses, thus regulating AD symptoms [236,237,238,239]. Gut microbiota composition is associated with TLR signalling in atopic diseases. Infants with eczema have lower fecal Ruminococcaceae, which is negatively associated with TLR2-induced IL-6 and TNF production [240]. Moreover, TLR4 SNP rs10759932 and fecal E. coli had a significant multiplicative interaction regarding allergic sensitization [241]. To the best of our knowledge, few studies have looked at the crosstalk between gut microbiota and TLRs in AD development.
8. Therapeutic Manipulation of Gut Microbiota in the Prevention and Treatment of AD
Since gut microbiota is likely to regulate AD pathogenesis, both gut microbiota and microbial metabolites are promising tools in controlling AD prevalence. The major therapeutic methods relevant to gut microbiota include FMT, the use of prebiotics, probiotics, synbiotics, and postbiotics. These methods all aim to restore the balance of gut ecology in order to modulate allergic immune responses. While abundant trials attempted to use these as therapies to prevent and treat AD, at this stage, their efficacies remain inconclusive due to inconsistent results and potential risks.
8.1. Fecal Microbiota Transplantation
The most direct method to re-establish the balance of the gut microbiota is FMT. FMT has been successfully used to treat various diseases, including Clostridium difficile infection [242]. A recent study reported that FMT using feces from healthy BALB/c donors successfully attenuated ovalbumin-induced AD in BALB/c mice [243], hinting at the promise of FMT in AD management. However, caution should be taken when applying FMT. FMT has resulted in several deaths in the recent decade through the spread of multidrug-resistant bacteria, aspiration, and toxic megacolon [244,245,246,247] and has elicited an important safety alert from United States Food and Drug Administration in 2019 [244]. Due to the limited knowledge on the gut microbiota, the actual operative agents in FMT treatments are mostly unknown, and the uncertain risks are not well known. Since patients with gut dysbiosis tend to have compromised gut barrier, they are more susceptible to the risks associated with FMT. Furthermore, regulatory benchmarks and standardized protocols for FMT are still absent or at least limited in many jurisdictions [248]. The alternative microbiota-based therapies with rationally selected microorganisms or microbial metabolites may be preferable for future AD treatments.
8.2. Prebiotics in Preventing and Treating AD
Prebiotics are non-viable substances that are selectively utilized by host microorganisms conferring health benefits [249]. Prebiotics are naturally rich in human milk and vegetarian food, including cereal, fruits, and vegetables. Modern industry also produces some prebiotics. Therefore, prebiotics can be supplemented either directly or by modifying food intake.
Investigations into the health effects of breastfeeding on infants led to the first applications of prebiotics to benefit health. Breastfed infants have different fecal bacterial composition compared to non-breastfed infants, with enrichment of Bifidobacteria, and such difference correlates with the health status of the infants [159,160,161,162]. Recent studies confirmed that intake of breast milk is the most significant factor shaping the early life gut microbiota [250,251]. Breastfed infants have a reduced risk of childhood AD compared to non-breastfed infants [153,154,155,156,157,158]. The beneficial effect of breast milk has been attributed to the prebiotics, human milk oligosaccharides (HMOs) [252,253,254]. HMOs selectively expand Bifidobacteria, particularly B. bifidum and B. longus, which, together with B. breve are the most abundant bacteria in the gut microbiota of breastfed infants [255]. Supplementing HMO effectively modulates the infant gut microbiota [256]. However, whether supplementing breast milk-derived prebiotics prevents AD is still questionable. Supplementing HMOs, mostly galacto-oligosaccharide (GOS) and fructo-oligosaccharide (FOS), effectively reduced the incidence of AD during infancy but exhibited poor long-term protection in several RCTs [257,258,259,260,261,262,263,264]. The results from other trials were not sufficiently in favour of supplementing HMOs in treating AD [265,266,267,268,269,270,271]. Of note, no adverse side effects were noted in all these trials and prebiotic HMOs have been added to infant milk formula to mimic the bifidogenic effect of breast milk [258,272,273].
Fermentable dietary fibres are probably the most popular prebiotics in recent biomedical studies [249]. They can be fermented by the commensal gut microbiota into SCFAs, whose anti-inflammatory functions have been discussed above. However, whether supplementing fermentable dietary fibre would benefit AD still requires more investigations in both pre-clinical and clinical studies.
Due to the lack of solid scientific evidence, the use of prebiotics is not recommended by World Allergy Organization (WAO) as a preventive or therapeutic measure against AD [270]. Future studies may demonstrate the efficacy of using non-digestible carbohydrates in preventing or even treating AD.
8.3. Probiotics in Preventing and Treating AD
Probiotics are living microorganisms that confer a health benefit on the host when administered in an adequate amount [274]. The benefits of probiotics in AD are studied more extendedly and thus have more solid conclusions. The most commonly used probiotics in RCTs against AD belong to Lactobacilli and Bifidobacteria genus, which are enriched in dairy products. These bacteria are able to modulate immune cells to restore the TH1/TH2 immune balance, enhance the production of the regulatory cytokine IL-10 and expand the population of Tregs [275], all of which benefit AD management. In addition, they compete with pathogenic bacteria, including S. aureus associated with AD, for nutrition and binding mucin [276]. Several RCTs attempted to test the efficacy of probiotics in preventing the development of AD (Table 3). Application of a single strain of probiotic demonstrates potent and enduring prevention against AD. Supplementation with L. rhamnosus GG (LGG) successfully reduced the incidence of AD in the first year of life by ~50% compared to placebo treatment in an RCT performed in Finland [277], and such protection extended to 4 years of age [278]. Another large-scale RCT in New Zealand suggested that early exposure to L. rhamnosus (HN001) had enduring protection against AD, at least in the first decade of life [279,280,281,282]. The further investigation suggested that the prevention against AD provided by HN001 may overcome the risk of AD caused by certain genetic deficiencies, particularly in genes encoding TLRs [283,284]. Nevertheless, supplements of HN001 did not have significant impacts on the infant gut microbiota [285]. A combination of various probiotics appears to be effective in preventing AD as well. An RCT performed in Norway demonstrated that maternal uptake of a combination of three probiotics, LGG, L. acidophilus La-5 and B. lactis Bb-12, had a potent preventive effect against the development of AD up to 6 years, which is accompanied with a reduction in TH22 [286,287,288]. A probiotic mix comprised of B. bifidum BGN4, B. animalis subsp. lactis AD011 and L. acidophilus AD031 also significantly reduced the incidence of AD in the first year of life in a Korean RCT [289]. Supplement of B. breve and B. longum together exhibited similar benefits and reduced fecal Proteobacteria [290]. However, probiotic treatments, particularly prenatal treatments alone, failed to prevent the development of AD in early life in multiple other RCTs without comprehensive follow-ups [291,292,293,294,295]. Such variations may be due to the differences in the participants enrolled, experimental designs, cultures of the probiotics and the dose administered. A follow-up study from the Norwegian RCT [286,287,288] suggested that the efficacy of probiotics may depend on the intrinsic microbiota, that high levels of fecal B. dentium is associated with weaker protection against AD by the probiotics treatment [296]. Overall, systematic reviews and meta-analyses based on the literature above suggest that probiotics have a long-term preventive effect against AD [297,298,299,300,301,302,303,304,305,306,307,308,309,310,311]. Therefore, although contradictory results exist, WAO recommends using probiotics during pregnancy, lactation and infancy to prevent the development of allergy [312].
Supplementation with probiotics has also been used to treat AD in RCTs (Table 3). Many RCTs selected L. rhamnosus, particularly the strain LGG [277,278]. The first RCT using probiotics to manage AD achieved improvement in AD infants by orally applying LGG [313]. LGG alleviated infantile AD with no major impact on the gut microbiota in another RCT [314]. Nevertheless, LGG supplement failed to rescue infantile AD in multiple more recent RCTs [315,316,317]. Another strain of L. rhamnosus, MP108, showed a therapeutic effect on AD [318]. More recently, a mixture of three strains of L. rhamnosus (ŁOCK 0900, ŁOCK 0908, ŁOCK 0918) significantly benefitted infants with AD [319]. Supplement of other Lactobacilli appeared effective in alleviating AD symptoms as well, including L. salivarius [320,321], L. fermentum [322,323], L. sakei [324], L. plantarum [325,326,327], and L. paracasei [323], although contradictory results exist for L. paracasei [328]. Combination treatment of different Lactobacilli also potently attenuate AD [323,329,330]. Bifidobacteria are also used in RCTs against AD, particularly B. lactis. B. lactis Bb-12 was effective in improving AD in the first trial using probiotics to manage AD [313]. Supplementing B. lactis LKM512 expanded Lactobacilli in gut microbiota and alleviated AD, likely through the increased production of kynurenic acids, a product of tryptophan metabolism [331]. However, B. lactis CNCM I-3446 demonstrated no benefit in the treatment of AD [328]. Using the mixtures of Lactobacilli and Bifidobacteria appropriately may be a promising therapy against AD. Supplements of L. salivarius and B. breve combination conferred a significant improvement in adult AD patients and a potent regulation on the TH1/TH2 immune balance [332]. A mixture of B. lactis, B. longum and L. casei accelerated the recovery of moderate AD with steroid treatment [333]. Another combination of probiotics, L. salivarius and S. thermophilus is also beneficial for AD management [334]. Probiotic treatments selectively attenuated IgE-sensitized AD [335,336] and food-sensitized AD infants [337], suggesting probiotics may not be a wide-spectral therapy against AD. Aligned with the mixed results, systematic reviews and meta-analyses cannot conclude the efficacy of probiotics in treating AD [299,311,338,339,340,341,342,343].
While future RCTs may continue to identify novel species or strains of probiotics that may be effective in controlling and treating AD, more effort is needed to standardize the current probiotics applications in the battle against AD and to set up regulations in the use of probiotics.
Table 3.
Summary of the randomized clinical trials using probiotics to prevent or treat AD.
| Year | Completed Participants | Probiotics Used | Intervention Route and Duration | Results | Reference |
|---|---|---|---|---|---|
| Prevention | |||||
| 2001 | 132 children with high risk of allergy and their mothers; 64 in probiotic group vs. 68 in placebo group | L. rhamnosus GG (ATCC 53103) | Oral/2–4 weeks prenatally + 6 months postnatally (by either mothers or infants) | Probiotic group: ↓incidence of AD in the first year of life and at 4 years of age | [277,278] |
| 2007 | 178 children with atopic mothers; 89 in probiotic group vs. 89 in placebo group | L. acidophilus (LAVRI-A1) | Oral/the first 6 months of life | Probiotic group: no effect on AD in the first year of life, ↑incidence of allergen sensitization | [291] |
| 2008 | 94 children with high risk of allergy and their mothers; 50 in probiotic group vs. 44 in placebo group | LGG | Oral/2–4 weeks prenatally + 6 months postnatally (by either mothers or infants) | Probiotic group: no effect on AD at 2 years of age, ↑incidence of recurrent wheezing bronchitis | [292] |
| 2008 | 474 children with high risk of allergy and their mothers; 157 in HN001 group vs. 158 in HN019 group vs. 159 in placebo group | L. rhamnosus HN001 or B. animalis HN019 | Oral/From 35 weeks gestation until 2 years postnatally (by mothers and children) | HN001 group: ↓incidence of AD at 2 years, 4 years, 6 years, and 11 years of age; HN019 group: no effect | [279,280,281,282] |
| 2009 | 245 Asian infants with high risk of allergy; 124 in probiotics group vs. 121 in placebo group | Bifidobacterium longum BL999, L. rhamnosus | Oral/The first 6 months of life | No effect at 1 year of age | [293] |
| 2010 | 112 children with high risk of allergy and their mothers; 57 in probiotics group vs. 55 in placebo group | B. bifidum BGN4, B. animalis subsp. lactis (B. lactis) AD011, L. acidophilus AD031 | Oral/4–8 weeks prenatally + 6 months postnatally (by either mothers or infants) | Probiotics group: ↓incidence of AD in the first year of life | [289] |
| 2010 | 278 children and their mother; 138 in probiotics group vs. 140 in placebo group | Combination of LGG, L. acidophilus La-5, B. lactis Bb-12 | Oral/From 36 weeks of gestation until 3 months postnatally during breastfeeding by mothers | Probiotics group: ↓incidence of AD at 2 years and 6 years of age, ↓TH22, No adverse effect | [286,287,288] |
| 2011 | 250 children with high risk of allergy and their mothers; 125 in probiotic group vs. 125 in placebo group | LGG | Oral/From 36 weeks of gestation until delivery | Probiotic group: no effect on AD in the first year of life, ↓CD14 and IgA in maternal breast milk | [294] |
| 2014 | 158 children and their mothers; 122 in probiotics group vs. 36 in placebo group | Combination of B. breve M-16V, B. longum BB536 | Oral/1 month prenatally + 6 months postnatally by infants | Probiotics group: ↓incidence of AD at 10 and 18 months of age, ↓fecal Proteobacteria, no adverse effect | [290] |
| 2018 | 423 children with high risk of allergy and their mothers; 212 in HN001 group vs. 211 in placebo group | L. rhamnosus HN001 | Oral/From 14–16 weeks of gestation until 6 years postnatally during breastfeeding by mothers | No effect on infantile AD | [295] |
| Treatment | |||||
| 2000 | 27 infants with AD; 9 in LGG group vs. 9 in Bb-12 group vs. 9 in placebo group | LGG, B. lactis Bb-12 (Bb-12) | Oral/3 months | Probiotic groups: ↓AD symptoms, ↓serum soluble CD4, ↓urine eosinophilic protein X | [313] |
| 2003 | 35 infants with AD; 14 in LGG group vs. 13 in heat-inactivated LGG group vs. 8 in placebo group | LGG | Oral/7.5 weeks | LGG group: ↓AD symptoms | [314] |
| 2003 | 43 children with AD; 20 in placebo→probiotics, 23 in probiotics→placebo | Mixture of L. rhamnosus 19070-2 and L. reuteri DSM 122460 | Oral/First intervention (6 weeks)→Washout (6 weeks)→Second intervention (6 weeks) | Probiotics treatment: ↓AD symptoms, ↓serum eosinophil cationic proteins | [329] |
| 2004 | 80 in LGG group, 76 in mix group, 74 in placebo group | LGG, Mixture of 4 probiotics (Mix, LGG, L. rhamnosus LC705, B. breve Bbi99, Propionibacterium freudenreichii ssp. Shermanii JS) | Oral/4 weeks | Generally, no obvious effect, Probiotics group: ↓IgE sensitized AD | [336] |
| 2005 | 53 children with moderate-severe AD (Topical corticosteroids were permitted); 26 in probiotic group vs. 27 in placebo group | L. fermentum VRI-003 PCC | Oral/8 weeks | Probiotics treatment: ↓AD symptoms | [322] |
| 2006 | 59 children with AD; 29 in probiotics group vs. 30 in placebo group | Mixture of LGG and B. lactis Bb-12 (Bb-12) | Oral/18 weeks | All participants probiotics: ↓AD symptoms (non-significant); within food sensitized participants, probiotics group: ↓AD symptoms (significant) | [337] |
| 2006 | 50 infants with AD; 17 in Lrh group, 16 in LGG group, 17 in placebo group | L. rhamnosus (Lrh), LGG | Oral/3 months | No therapeutic effect and no immune difference | [315] |
| 2006 | 53 infants with moderate-severe AD (Emollients, class I–II topical corticosteroids and antihistamines were permitted); 26 in probiotic group, 27 in placebo group | LGG | Oral/8 weeks | No therapeutic effect | [316] |
| 2007 | 102 infants with mild-moderate AD; 54 in probiotic group, 48 in placebo group | LGG | Oral/12 weeks | No therapeutic effect | [317] |
| 2010 | 88 children with AD; 45 in probiotic group vs. 43 in placebo group | L. sakei KCTC 10755BP | Oral/12 weeks | Probiotic group: ↓AD symptoms, ↓serum CCL17 and CCL27 | [324] |
| 2011 | 141 children with AD; 45 in LP group, 47 in BL group, 47 in placebo group | L. paracasei CNCM I-2116(LP), B. lactis CNCM I-3446 (BL) | Oral/3 months | No therapeutic effect | [328] |
| 2011 | 38 adult AD patients; 19 in probiotic group vs. 19 in placebo group | L. salivarius LS01 (DSM 22775) | Oral/16 weeks | Probiotic group: ↓AD symptoms, ↓fecal load of Staphylococci, ↓plasma LPS, enduring restoration of TH1/TH2 immune balance | [320,321] |
| 2012 | 46 adult AD patients; 31 in probiotics group vs. 15 in placebo group | The combination of L. salivarius LS01, B. breve BR3 | Oral/12 weeks | Probiotics group: ↓AD symptoms, ↓plasma LPS, ↓activated T cells, ↑TH1, ↓TH2, ↓TH17, ↑ Treg cells, ↓fecal Staphylococci | [332] |
| 2012 | 118 children with AD (Emollients were permitted); 58 in probiotic group vs. 60 in placebo group | L. plantarum CJLP133 | Oral/12 weeks | Probiotic group: ↓AD symptoms, ↓total eosinophil count, ↓IL-4 and IFNγ in blood | [326] |
| 2014 | 25 adult AD patients; 13 in probiotics group vs. 12 in placebo group | L. salivarius LS01 (DSM 22775), S. thermophilus ST10 (DSM25246) | Oral/1 month | Probiotic group: ↓AD symptoms | [334] |
| 2014 | 44 adult AD patients (Medications without probiotic effect and corticosteroid application were permitted); 22 in probiotic group vs. 22 in placebo group | B. lactis LKM512 | Oral/8 weeks | Probiotic group: ↓AD symptoms, ↑fecal Lactobacilli, ↑fecal kynurenic acid | [331] |
| 2015 | 212 children with moderate-severe AD; 55 in LP group vs. 53 in LF group vs. 51 in LP + LF group vs. 53 in placebo group | L. paracasei (LP), L. fermentum (LF) and the combination of LP and LF | Oral/3 months | LP group, LF group and LP + LF group: ↓AD symptoms, ↓serum IL-4, IgE, TNF, ↑serum IFN, TGF, ↓urine eosinophilic protein X, 8-OHdG | [323] |
| 2017 | 62 children with AD; 30 in probiotic group vs. 32 in placebo group | L. rhamnosus (MP108) | Oral/8 weeks | Probiotic group: ↓AD symptoms | [318] |
| 2017 | 22 children with AD; 12 in probiotic group vs. 10 in placebo group | L. plantarun IS-10506 | Oral/12 weeks | Probiotic group: ↓AD symptoms, ↓serum IL-4, IFNγ, IL-17, ↑serum IL-10↑, ↑Treg cells in blood | [325] |
| 2018 | 50 children with moderate AD who were prescribed topical steroids; 26 in probiotics group vs. 24 in placebo group | Mixture of B. lactis CECT 8145, B. longum CECT 7347, L. casei CECT 9104 | Oral/12 weeks | Probiotics group: ↓AD symptoms, ↓steroids treatment | [333] |
| 2020 | 109 adult AD patients; 29 in CCFM16 group vs. 43 in CCFM8610 group vs. 11 in oligosaccharide group vs. 26 in placebo group | B. bifidum CCFM16, L. plantarum CCFM8610 | Oral/8 weeks | L. plantarum CCFM8610 group: ↓AD symptoms, ↑serum IL-10, ↓microbial functional genes involving S. aureus infection AD symptoms, ↑steroid hormone biosynthesis | [327] |
| 2020 | 82 children with mild-moderate AD; 41 in probiotic group vs. 41 in placebo group | L. pentosus | Oral/12 weeks | Generally, no obvious effect, probiotic group: ↓IgE sensitized AD, (no difference in cytokine levels and microbial diversities) | [335] |
| 2021 | 134 children with AD; 66 in probiotics group vs. 68 in placebo group | L. rhamnosus ŁOCK 0900, ŁOCK 0908, ŁOCK 0918 | Oral/3 months | Probiotics group: ↓AD symptoms | [319] |
| 2021 | 80 adult AD patients; 40 in Probiotics group vs. 40 in placebo group | L. plantarum PBS067, L. reuteri PBS072 and L. rhamnosus LRH020 | Oral/56 days | Probiotics group: ↓AD symptoms, ↓skin TNF and TSLP | [330] |
8.4. Synbiotics and Postbiotics in Treating AD
There are attempts to combat AD by combining the administration of prebiotics and probiotics. A new term “synbiotic” was defined in 2019 as “a mixture comprising live microorganisms and substrate(s) selectively utilized by host microorganisms that confers a health benefit on the host” [344]. RCTs against AD using synbiotics with limited scales demonstrated mixed results [264,345,346,347,348,349,350,351]. Nevertheless, well-designed synbiotics are likely to lead to novel, effective therapeutics against AD.
Postbiotics are inanimate microorganisms and/or their components with beneficial effects on host health [352], including the lysates of microorganisms, heat-inactivated microorganisms, and microbial metabolites. The trials using lysates and heat-inactivated microorganisms are highly contradictory. Early attempts using heat-inactivated LGG failed to benefit AD and even resulted in adverse gastrointestinal symptoms, including diarrhea [314]. Similarly, heat-treated L. paracei GM-080 did not accelerate the recovery of infantile AD with topical corticosteroid treatment [353]. On the contrary, trials using the postbiotics from the different strains of the same species were potent in improving AD. Tyndallized L. rhamnosus IDCC3201 (RHT3201) had therapeutic improvement in children with moderate AD and decreased eosinophil cationic protein and IL-31 in blood [354]. Heat-killed L. paracasei K71 alleviated adult AD and reduced the use of topical corticosteroids [355]. Heat-killed L. acidophilus strain L-92 attenuated AD symptoms in both children and adults [356,357], decreased eosinophil count, and increased serum TGF-β [357]. In addition, lysate of Vitreoscilla filiformis significantly improved AD and reduced fecal S. aureus [358]. The application of lysates and heat-inactivated microorganisms faces similar challenges as the application of probiotics in lacking standard and consistency.
Microbial metabolites have advantages in standardization, but current investigations are still limited to the pre-clinical stage [359]. With sufficient knowledge from pre-clinical studies, more trials should utilize microbial metabolites, including SCFAs, D-tryptophan, and IAld to test their clinical efficacy against AD.
9. Concluding Remarks
The recent discoveries on how the skin and gut microbiota modulate AD pathogenesis point out a bright potential in manipulating commensal bacteria to prevent and manage AD. Current therapies that leverage the skin microbiota, however, are still limited. Further investigations are needed to improve the specificity of the therapies against pathogenic bacteria, such as S. aureus, while encouraging the growth of normal bacteria at the same time. Clinical trials have been conducted to manipulate the gut microbiota for the prevention and treatment against AD. Although the efficacies of certain treatments are still controversial, the oral application of probiotics during pregnancy, lactation, and infancy is recommended for the prevention of allergic diseases, including AD [312]. More efforts are needed to standardize gut microbiota-based interventions against AD. In addition, microbial metabolites may aid in AD management, but they need to be tested in future trials. Furthermore, using these microbiota-based treatments in conjunction with the traditional treatments may achieve improved clinical outcomes. The role of commensal bacteria in AD is still a relatively new area. Increased understanding in this area would improve the management of AD, an increasingly prevalent atopic disease.
Author Contributions
M.J.A. and L.X. wrote the original manuscript. Y.-A.Y. and F.Z.M. provided intellectual inputs and reviewed the manuscript. R.R. integrated and reviewed the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
F.Z.M. is supported by a National Heart Foundation Future Leader Fellowship (105663) and a Senior Medical Research Fellowship from the Sylvia and Charles Viertel Charitable Foundation. M.J.A and L.X. are supported by Monash Graduate Scholarship from Monash University, Australia.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Eichenfield, L.F.; Tom, W.L.; Chamlin, S.L.; Feldman, S.R.; Hanifin, J.M.; Simpson, E.L.; Berger, T.G.; Bergman, J.N.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 1. Diagnosis and assessment of atopic dermatitis. J. Am. Acad. Dermatol. 2014, 70, 338–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidinger, S.; Novak, N. Atopic dermatitis. Lancet 2016, 387, 1109–1122. [Google Scholar] [CrossRef]
- Bieber, T. Atopic Dermatitis. Ann. Derm. 2010, 22, 125–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deckers, I.A.G.; McLean, S.; Linssen, S.; Mommers, M.; van Schayck, C.P.; Sheikh, A. Investigating International Time Trends in the Incidence and Prevalence of Atopic Eczema 1990–2010: A Systematic Review of Epidemiological Studies. PLoS ONE 2012, 7, e39803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, H.; Stewart, A.; von Mutius, E.; Cookson, W.; Anderson, H.R. Is eczema really on the increase worldwide? J. Allergy Clin. Immunol. 2008, 121, 947–954.e15. [Google Scholar] [CrossRef]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef]
- Bisgaard, H.; Simpson, A.; Palmer, C.N.; Bonnelykke, K.; McLean, I.; Mukhopadhyay, S.; Pipper, C.B.; Halkjaer, L.B.; Lipworth, B.; Hankinson, J.; et al. Gene-environment interaction in the onset of eczema in infancy: Filaggrin loss-of-function mutations enhanced by neonatal cat exposure. PLoS Med. 2008, 5, e131. [Google Scholar] [CrossRef] [Green Version]
- Palmer, C.N.; Irvine, A.D.; Terron-Kwiatkowski, A.; Zhao, Y.; Liao, H.; Lee, S.P.; Goudie, D.R.; Sandilands, A.; Campbell, L.E.; Smith, F.J.; et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat. Genet. 2006, 38, 441–446. [Google Scholar] [CrossRef]
- Sandilands, A.; Terron-Kwiatkowski, A.; Hull, P.R.; O’Regan, G.M.; Clayton, T.H.; Watson, R.M.; Carrick, T.; Evans, A.T.; Liao, H.; Zhao, Y.; et al. Comprehensive analysis of the gene encoding filaggrin uncovers prevalent and rare mutations in ichthyosis vulgaris and atopic eczema. Nat. Genet. 2007, 39, 650–654. [Google Scholar] [CrossRef]
- Egawa, G.; Kabashima, K. Multifactorial skin barrier deficiency and atopic dermatitis: Essential topics to prevent the atopic march. J. Allergy Clin. Immunol. 2016, 138, 350–358.e1. [Google Scholar] [CrossRef] [Green Version]
- Eichenfield, L.F.; Tom, W.L.; Berger, T.G.; Krol, A.; Paller, A.S.; Schwarzenberger, K.; Bergman, J.N.; Chamlin, S.L.; Cohen, D.E.; Cooper, K.D.; et al. Guidelines of care for the management of atopic dermatitis: Section 2. Management and treatment of atopic dermatitis with topical therapies. J. Am. Acad. Dermatol. 2014, 71, 116–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faiz, S.; Giovannelli, J.; Podevin, C.; Jachiet, M.; Bouaziz, J.-D.; Reguiai, Z.; Nosbaum, A.; Lasek, A.; le Bouedec, M.-C.F.; Du Thanh, A.; et al. Effectiveness and safety of dupilumab for the treatment of atopic dermatitis in a real-life French multicenter adult cohort. J. Am. Acad. Dermatol. 2019, 81, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Abraham, S.; Haufe, E.; Harder, I.; Heratizadeh, A.; Kleinheinz, A.; Wollenberg, A.; Weisshaar, E.; Augustin, M.; Wiemers, F.; Zink, A.; et al. Implementation of dupilumab in routine care of atopic eczema: Results from the German national registry TREATgermany. Br. J. Derm. 2020, 183, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Deleuran, M.; Thaçi, D.; Beck, L.A.; de Bruin-Weller, M.; Blauvelt, A.; Forman, S.; Bissonnette, R.; Reich, K.; Soong, W.; Hussain, I.; et al. Dupilumab shows long-term safety and efficacy in patients with moderate to severe atopic dermatitis enrolled in a phase 3 open-label extension study. J. Am. Acad. Dermatol. 2020, 82, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Kraus, C.N.; Patel, K.G.; Ganesan, A.K.; Grando, S.A. Real-world experience of dupilumab treatment for atopic dermatitis in adults: A retrospective analysis of patients’ records. Int. J. Dermatol. 2020, 59, 253–256. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Blauvelt, A.; Guttman-Yassky, E.; Worm, M.; Lynde, C.; Lacour, J.P.; Spelman, L.; Katoh, N.; Saeki, H.; Poulin, Y.; et al. Tralokinumab for moderate-to-severe atopic dermatitis: Results from two 52-week, randomized, double-blind, multicentre, placebo-controlled phase III trials (ECZTRA 1 and ECZTRA 2). Br. J. Derm. 2021, 184, 437–449. [Google Scholar] [CrossRef]
- Popovic, B.; Breed, J.; Rees, D.G.; Gardener, M.J.; Vinall, L.M.; Kemp, B.; Spooner, J.; Keen, J.; Minter, R.; Uddin, F.; et al. Structural Characterisation Reveals Mechanism of IL-13-Neutralising Monoclonal Antibody Tralokinumab as Inhibition of Binding to IL-13Rα1 and IL-13Rα2. J. Mol. Biol. 2017, 429, 208–219. [Google Scholar] [CrossRef]
- He, H.; Guttman-Yassky, E. JAK Inhibitors for Atopic Dermatitis: An Update. Am. J. Clin. Dermatol. 2019, 20, 181–192. [Google Scholar] [CrossRef]
- Cotter, D.G.; Schairer, D.; Eichenfield, L. Emerging therapies for atopic dermatitis: JAK inhibitors. J. Am. Acad. Dermatol. 2018, 78, S53–S62. [Google Scholar] [CrossRef]
- Bieber, T. Atopic dermatitis: An expanding therapeutic pipeline for a complex disease. Nat. Rev. Drug Discov. 2021, 21, 21–40. [Google Scholar] [CrossRef]
- Yew, Y.W.; Thyssen, J.P.; Silverberg, J.I. A systematic review and meta-analysis of the regional and age-related differences in atopic dermatitis clinical characteristics. J. Am. Acad. Derm. 2019, 80, 390–401. [Google Scholar] [CrossRef] [PubMed]
- Ruff, W.E.; Greiling, T.M.; Kriegel, M.A. Host–microbiota interactions in immune-mediated diseases. Nat. Rev. Microbiol. 2020, 18, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Grice, E.A.; Kong, H.H.; Renaud, G.; Young, A.C.; Program, N.C.S.; Bouffard, G.G.; Blakesley, R.W.; Wolfsberg, T.G.; Turner, M.L.; Segre, J.A. A diversity profile of the human skin microbiota. Genome Res. 2008, 18, 1043–1050. [Google Scholar] [CrossRef] [Green Version]
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.Y.; Lee, E.; Park, Y.M.; Hong, S.J. Microbiome in the Gut-Skin Axis in Atopic Dermatitis. Allergy Asthma. Immunol. Res. 2018, 10, 354–362. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.L.; Deming, C.; Cassidy, S.K.B.; Harrison, O.J.; Ng, W.I.; Conlan, S.; Belkaid, Y.; Segre, J.A.; Kong, H.H. Staphylococcus aureus and Staphylococcus epidermidis strain diversity underlying pediatric atopic dermatitis. Sci. Transl. Med. 2017, 9, eaal4651. [Google Scholar] [CrossRef] [Green Version]
- Emmert, H.; Rademacher, F.; Gläser, R.; Harder, J. Skin microbiota analysis in human 3D skin models—“Free your mice”. Exp. Dermatol. 2020, 29, 1133–1139. [Google Scholar] [CrossRef]
- Kennedy, E.A.; Connolly, J.; Hourihane, J.O.; Fallon, P.G.; McLean, W.H.I.; Murray, D.; Jo, J.H.; Segre, J.A.; Kong, H.H.; Irvine, A.D. Skin microbiome before development of atopic dermatitis: Early colonization with commensal staphylococci at 2 months is associated with a lower risk of atopic dermatitis at 1 year. J. Allergy Clin. Immunol. 2017, 139, 166–172. [Google Scholar] [CrossRef] [Green Version]
- Nakatsuji, T.; Chen, T.H.; Narala, S.; Chun, K.A.; Two, A.M.; Yun, T.; Shafiq, F.; Kotol, P.F.; Bouslimani, A.; Melnik, A.V.; et al. Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci. Transl. Med. 2017, 9, eaah4680. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.H.; Oh, J.; Deming, C.; Conlan, S.; Grice, E.A.; Beatson, M.A.; Nomicos, E.; Polley, E.C.; Komarow, H.D.; Murray, P.R.; et al. Temporal shifts in the skin microbiome associated with disease flares and treatment in children with atopic dermatitis. Genome Res. 2012, 22, 850–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, M.R.; Nakatsuji, T.; Gallo, R.L. Staphylococcus aureus: Master Manipulator of the Skin. Cell Host Microbe 2017, 22, 579–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirasawa, Y.; Takai, T.; Nakamura, T.; Mitsuishi, K.; Gunawan, H.; Suto, H.; Ogawa, T.; Wang, X.-L.; Ikeda, S.; Okumura, K.; et al. Staphylococcus aureus Extracellular Protease Causes Epidermal Barrier Dysfunction. J. Investig. Dermatol. 2010, 130, 614–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, D.Y. Infection in atopic dermatitis. Curr. Opin. Pediatr. 2003, 15, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Maintz, L.; Novak, N. Modifications of the innate immune system in atopic dermatitis. J. Innate Immun. 2011, 3, 131–141. [Google Scholar] [CrossRef]
- Nakagawa, S.; Matsumoto, M.; Katayama, Y.; Oguma, R.; Wakabayashi, S.; Nygaard, T.; Saijo, S.; Inohara, N.; Otto, M.; Matsue, H.; et al. Staphylococcus aureus Virulent PSMalpha Peptides Induce Keratinocyte Alarmin Release to Orchestrate IL-17-Dependent Skin Inflammation. Cell Host Microbe 2017, 22, 667–677.e5. [Google Scholar] [CrossRef] [Green Version]
- Williams, M.R.; Nakatsuji, T.; Sanford, J.A.; Vrbanac, A.F.; Gallo, R.L. Staphylococcus aureus Induces Increased Serine Protease Activity in Keratinocytes. J. Investig. Dermatol. 2017, 137, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Baurecht, H.; Ruhlemann, M.C.; Rodriguez, E.; Thielking, F.; Harder, I.; Erkens, A.S.; Stolzl, D.; Ellinghaus, E.; Hotze, M.; Lieb, W.; et al. Epidermal lipid composition, barrier integrity, and eczematous inflammation are associated with skin microbiome configuration. J. Allergy Clin. Immunol. 2018, 141, 1668–1676.e16. [Google Scholar] [CrossRef] [Green Version]
- Lunjani, N.; Hlela, C.; O’Mahony, L. Microbiome and skin biology. Curr. Opin. Allergy Clin. Immunol. 2019, 19, 328–333. [Google Scholar] [CrossRef]
- Glatz, M.; Jo, J.H.; Kennedy, E.A.; Polley, E.C.; Segre, J.A.; Simpson, E.L.; Kong, H.H. Emollient use alters skin barrier and microbes in infants at risk for developing atopic dermatitis. PLoS ONE 2018, 13, e0192443. [Google Scholar] [CrossRef] [Green Version]
- Iwase, T.; Uehara, Y.; Shinji, H.; Tajima, A.; Seo, H.; Takada, K.; Agata, T.; Mizunoe, Y. Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 2010, 465, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Kaci, G.; Goudercourt, D.; Dennin, V.; Pot, B.; Doré, J.; Ehrlich, S.D.; Renault, P.; Blottière, H.M.; Daniel, C.; Delorme, C. Anti-inflammatory properties of Streptococcus salivarius, a commensal bacterium of the oral cavity and digestive tract. Appl. Environ. Microbiol. 2014, 80, 928–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clausen, M.L.; Agner, T.; Lilje, B.; Edslev, S.M.; Johannesen, T.B.; Andersen, P.S. Association of Disease Severity with Skin Microbiome and Filaggrin Gene Mutations in Adult Atopic Dermatitis. JAMA Derm. 2018, 154, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.; Freeman, A.F.; Program, N.C.S.; Park, M.; Sokolic, R.; Candotti, F.; Holland, S.M.; Segre, J.A.; Kong, H.H. The altered landscape of the human skin microbiome in patients with primary immunodeficiencies. Genome Res. 2013, 23, 2103–2114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laborel-Preneron, E.; Bianchi, P.; Boralevi, F.; Lehours, P.; Fraysse, F.; Morice-Picard, F.; Sugai, M.; Sato’o, Y.; Badiou, C.; Lina, G.; et al. Effects of the Staphylococcus aureus and Staphylococcus epidermidis Secretomes Isolated from the Skin Microbiota of Atopic Children on CD4+ T Cell Activation. PLoS ONE 2015, 10, e0141067. [Google Scholar] [CrossRef] [Green Version]
- Shi, B.; Bangayan, N.J.; Curd, E.; Taylor, P.A.; Gallo, R.L.; Leung, D.Y.M.; Li, H. The skin microbiome is different in pediatric versus adult atopic dermatitis. J. Allergy Clin. Immunol. 2016, 138, 1233–1236. [Google Scholar] [CrossRef] [Green Version]
- Drago, L.; De Grandi, R.; Altomare, G.; Pigatto, P.; Rossi, O.; Toscano, M. Skin microbiota of first cousins affected by psoriasis and atopic dermatitis. Clin. Mol. Allergy 2016, 14, 2. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.H.; Rho, M.; Choi, J.P.; Choi, H.I.; Park, H.K.; Song, W.J.; Min, T.K.; Cho, S.H.; Cho, Y.J.; Kim, Y.K.; et al. A Metagenomic Analysis Provides a Culture-Independent Pathogen Detection for Atopic Dermatitis. Allergy Asthma Immunol. Res. 2017, 9, 453–461. [Google Scholar] [CrossRef]
- Fyhrquist, N.; Muirhead, G.; Prast-Nielsen, S.; Jeanmougin, M.; Olah, P.; Skoog, T.; Jules-Clement, G.; Feld, M.; Barrientos-Somarribas, M.; Sinkko, H.; et al. Microbe-host interplay in atopic dermatitis and psoriasis. Nat. Commun. 2019, 10, 4703. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Xu, X.; Wen, H.; Wang, Z.; Ding, C.; Liu, X.; Gao, Y.; Su, H.; Zhang, J.; Han, Y.; et al. Inverse Association Between the Skin and Oral Microbiota in Atopic Dermatitis. J. Investig. Derm. 2019, 139, 1779–1787.e12. [Google Scholar] [CrossRef]
- Capone, K.; Kirchner, F.; Klein, S.L.; Tierney, N.K. Effects of Colloidal Oatmeal Topical Atopic Dermatitis Cream on Skin Microbiome and Skin Barrier Properties. J. Drugs Derm. 2020, 19, 524–531. [Google Scholar]
- Xu, Z.; Liu, X.; Niu, Y.; Shen, C.; Heminger, K.; Moulton, L.; Yu, A.; Allen, T.; Zhang, L.; Yue, F.; et al. Skin benefits of moisturising body wash formulas for children with atopic dermatitis: A randomised controlled clinical study in China. Australas J. Derm. 2020, 61, e54–e59. [Google Scholar] [CrossRef] [PubMed]
- Smits, J.P.H.; Ederveen, T.H.A.; Rikken, G.; van den Brink, N.J.M.; van Vlijmen-Willems, I.; Boekhorst, J.; Kamsteeg, M.; Schalkwijk, J.; van Hijum, S.; Zeeuwen, P.; et al. Targeting the Cutaneous Microbiota in Atopic Dermatitis by Coal Tar via AHR-Dependent Induction of Antimicrobial Peptides. J. Investig. Derm. 2020, 140, 415–424.e10. [Google Scholar] [CrossRef] [PubMed]
- Khadka, V.D.; Key, F.M.; Romo-Gonzalez, C.; Martinez-Gayosso, A.; Campos-Cabrera, B.L.; Geronimo-Gallegos, A.; Lynn, T.C.; Duran-McKinster, C.; Coria-Jimenez, R.; Lieberman, T.D.; et al. The Skin Microbiome of Patients With Atopic Dermatitis Normalizes Gradually During Treatment. Front. Cell. Infect. Microbiol. 2021, 11, 720674. [Google Scholar] [CrossRef]
- Leyden, J.J.; Marples, R.R.; Kligman, A.M. Staphylococcus aureus in the lesions of atopic dermatitis. Br. J. Dermatol. 1974, 90, 525–530. [Google Scholar] [CrossRef]
- Gong, J.Q.; Lin, L.; Lin, T.; Hao, F.; Zeng, F.Q.; Bi, Z.G.; Yi, D.; Zhao, B. Skin colonization by Staphylococcus aureus in patients with eczema and atopic dermatitis and relevant combined topical therapy: A double-blind multicentre randomized controlled trial. Br. J. Dermatol. 2006, 155, 680–687. [Google Scholar] [CrossRef]
- Meylan, P.; Lang, C.; Mermoud, S.; Johannsen, A.; Norrenberg, S.; Hohl, D.; Vial, Y.; Prod’hom, G.; Greub, G.; Kypriotou, M.; et al. Skin Colonization by Staphylococcus aureus Precedes the Clinical Diagnosis of Atopic Dermatitis in Infancy. J. Investig. Dermatol. 2017, 137, 2497–2504. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.-H.; Strickland, I.; Boguniewicz, M.; Leung, D.Y.M. Fibronectin and fibrinogen contribute to the enhanced binding of Staphylococcus aureus to atopic skin. J. Allergy Clin. Immunol. 2001, 108, 269–274. [Google Scholar] [CrossRef]
- Geoghegan, J.A.; Irvine, A.D.; Foster, T.J. Staphylococcus aureus and Atopic Dermatitis: A Complex and Evolving Relationship. Trends Microbiol. 2018, 26, 484–497. [Google Scholar] [CrossRef]
- Alexander, H.; Paller, A.S.; Traidl-Hoffmann, C.; Beck, L.A.; De Benedetto, A.; Dhar, S.; Girolomoni, G.; Irvine, A.D.; Spuls, P.; Su, J.; et al. The role of bacterial skin infections in atopic dermatitis: Expert statement and review from the International Eczema Council Skin Infection Group. Br. J. Derm. 2020, 182, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- van Drongelen, V.; Haisma, E.M.; Out-Luiting, J.J.; Nibbering, P.H.; El Ghalbzouri, A. Reduced filaggrin expression is accompanied by increased Staphylococcus aureus colonization of epidermal skin models. Clin. Exp. Allergy 2014, 44, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Sieprawska-Lupa, M.; Mydel, P.; Krawczyk, K.; Wojcik, K.; Puklo, M.; Lupa, B.; Suder, P.; Silberring, J.; Reed, M.; Pohl, J.; et al. Degradation of human antimicrobial peptide LL-37 by Staphylococcus aureus-derived proteinases. Antimicrob. Agents Chemother. 2004, 48, 4673–4679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakatsuji, T.; Chen, T.H.; Two, A.M.; Chun, K.A.; Narala, S.; Geha, R.S.; Hata, T.R.; Gallo, R.L. Staphylococcus aureus Exploits Epidermal Barrier Defects in Atopic Dermatitis to Trigger Cytokine Expression. J. Investig. Derm. 2016, 136, 2192–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonesson, A.; Przybyszewska, K.; Eriksson, S.; Mörgelin, M.; Kjellström, S.; Davies, J.; Potempa, J.; Schmidtchen, A. Identification of bacterial biofilm and the Staphylococcus aureus derived protease, staphopain, on the skin surface of patients with atopic dermatitis. Sci. Rep. 2017, 7, 8689. [Google Scholar] [CrossRef]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [Green Version]
- Berube, B.J.; Wardenburg, J.B. Staphylococcus aureus α-Toxin: Nearly a Century of Intrigue. Toxins 2013, 5, 1140. [Google Scholar] [CrossRef] [Green Version]
- Adam, R.S.; Salgado-Pabón, W.; Petra, L.K.; Alexander, R.H.; Donald, Y.M.L.; Patrick, M.S. Staphylococcal and Streptococcal Superantigen Exotoxins. Clin. Microbiol. Rev. 2013, 26, 422–447. [Google Scholar] [CrossRef] [Green Version]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Microbiol. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Bhardwaj, N.; Friedman, S.M.; Cole, B.C.; Nisanian, A.J. Dendritic cells are potent antigen-presenting cells for microbial superantigens. J. Exp. Med. 1992, 175, 267–273. [Google Scholar] [CrossRef]
- Hirose, A.; Ikejima, T.; Gill, D.M. Established macrophagelike cell lines synthesize interleukin-1 in response to toxic shock syndrome toxin. Infect. Immun. 1985, 50, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Han, J.H.; Chung, J.H.; Cho, K.H.; Eun, H.C. Role of staphylococcal superantigen in atopic dermatitis: Influence on keratinocytes. J. Korean Med. Sci. 2006, 21, 315–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proft, T.; Fraser, J.D. Bacterial superantigens. Clin. Exp. Immunol. 2003, 133, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Munoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Archer, N.K.; Dillen, C.A.; Wang, Y.; Ashbaugh, A.G.; Ortines, R.V.; Kao, T.; Lee, S.K.; Cai, S.S.; Miller, R.J. Staphylococcus aureus Epicutaneous Exposure Drives Skin Inflammation via IL-36-Mediated T Cell Responses. Cell Host Microbe 2017, 22, 653–666.e5. [Google Scholar] [CrossRef] [PubMed]
- Syed, A.K.; Reed, T.J.; Clark, K.L.; Boles, B.R.; Kahlenberg, J.M. Staphlyococcus aureus phenol-soluble modulins stimulate the release of proinflammatory cytokines from keratinocytes and are required for induction of skin inflammation. Infect. Immun. 2015, 83, 3428–3437. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.W.; Choi, E.B.; Min, T.K.; Kim, J.H.; Kim, M.H.; Jeon, S.G.; Lee, B.J.; Gho, Y.S.; Jee, Y.K.; Pyun, B.Y.; et al. An important role of alpha-hemolysin in extracellular vesicles on the development of atopic dermatitis induced by Staphylococcus aureus. PLoS ONE 2014, 9, e100499. [Google Scholar] [CrossRef]
- Brauweiler, A.M.; Goleva, E.; Leung, D.Y.M. Th2 cytokines increase Staphylococcus aureus alpha toxin-induced keratinocyte death through the signal transducer and activator of transcription 6 (STAT6). J. Investig. Derm. 2014, 134, 2114–2121. [Google Scholar] [CrossRef] [Green Version]
- Brauweiler, A.M.; Bin, L.; Kim, B.E.; Oyoshi, M.K.; Geha, R.S.; Goleva, E.; Leung, D.Y.M. Filaggrin-dependent secretion of sphingomyelinase protects against staphylococcal α-toxin–induced keratinocyte death. J. Allergy Clin. Immunol. 2013, 131, 421–427.e2. [Google Scholar] [CrossRef] [Green Version]
- Claßen, A.; Kalali, B.N.; Schnopp, C.; Andres, C.; Aguilar-Pimentel, J.A.; Ring, J.; Ollert, M.; Mempel, M. TNF receptor I on human keratinocytes is a binding partner for staphylococcal protein A resulting in the activation of NF kappa B, AP-1, and downstream gene transcription. Exp. Dermatol. 2011, 20, 48–52. [Google Scholar] [CrossRef]
- Boguniewicz, M.; Leung, D.Y. Atopic dermatitis: A disease of altered skin barrier and immune dysregulation. Immunol. Rev. 2011, 242, 233–246. [Google Scholar] [CrossRef]
- Fournier, B. The function of TLR2 during staphylococcal diseases. Front. Cell. Infect. Microbiol. 2012, 2, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bekeredjian-Ding, I.; Inamura, S.; Giese, T.; Moll, H.; Endres, S.; Sing, A.; Zähringer, U.; Hartmann, G. Staphylococcus aureus protein A triggers T cell-independent B cell proliferation by sensitizing B cells for TLR2 ligands. J. Immunol. 2007, 178, 2803–2812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vu, A.T.; Baba, T.; Chen, X.; Le, T.A.; Kinoshita, H.; Xie, Y.; Kamijo, S.; Hiramatsu, K.; Ikeda, S.; Ogawa, H. Staphylococcus aureus membrane and diacylated lipopeptide induce thymic stromal lymphopoietin in keratinocytes through the Toll-like receptor 2-Toll-like receptor 6 pathway. J. Allergy Clin. Immunol. 2010, 126, 985–993.e3. [Google Scholar] [CrossRef]
- Kuo, I.H.; Carpenter-Mendini, A.; Yoshida, T.; McGirt, L.Y.; Ivanov, A.I.; Barnes, K.C.; Gallo, R.L.; Borkowski, A.W.; Yamasaki, K.; Leung, D.Y.; et al. Activation of epidermal toll-like receptor 2 enhances tight junction function: Implications for atopic dermatitis and skin barrier repair. J. Investig. Derm. 2013, 133, 988–998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, P.Y.; Ohtake, T.; Brandt, C.; Strickland, I.; Boguniewicz, M.; Ganz, T.; Gallo, R.L.; Leung, D.Y. Endogenous antimicrobial peptides and skin infections in atopic dermatitis. N. Engl. J. Med. 2002, 347, 1151–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naik, S.; Bouladoux, N.; Wilhelm, C.; Molloy, M.J.; Salcedo, R.; Kastenmuller, W.; Deming, C.; Quinones, M.; Koo, L.; Conlan, S.; et al. Compartmentalized control of skin immunity by resident commensals. Science 2012, 337, 1115–1119. [Google Scholar] [CrossRef] [Green Version]
- Meisel, J.S.; Sfyroera, G.; Bartow-McKenney, C.; Gimblet, C.; Bugayev, J.; Horwinski, J.; Kim, B.; Brestoff, J.R.; Tyldsley, A.S.; Zheng, Q.; et al. Commensal microbiota modulate gene expression in the skin. Microbiome 2018, 6, 20. [Google Scholar] [CrossRef]
- Uberoi, A.; Bartow-McKenney, C.; Zheng, Q.; Flowers, L.; Campbell, A.; Knight, S.A.B.; Chan, N.; Wei, M.; Lovins, V.; Bugayev, J.; et al. Commensal microbiota regulates skin barrier function and repair via signaling through the aryl hydrocarbon receptor. Cell Host Microbe 2021, 29, 1235–1248.e8. [Google Scholar] [CrossRef]
- Loomis, K.H.; Wu, S.K.; Ernlund, A.; Zudock, K.; Reno, A.; Blount, K.; Karig, D.K. A mixed community of skin microbiome representatives influences cutaneous processes more than individual members. Microbiome 2021, 9, 22. [Google Scholar] [CrossRef]
- Parlet, C.P.; Brown, M.M.; Horswill, A.R. Commensal Staphylococci Influence Staphylococcus aureus Skin Colonization and Disease. Trends Microbiol. 2019, 27, 497–507. [Google Scholar] [CrossRef]
- Schröder, J.M.; Harder, J. Antimicrobial skin peptides and proteins. Cell. Mol. Life Sci. CMLS 2006, 63, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Gallo, R.L.; Hooper, L.V. Epithelial antimicrobial defence of the skin and intestine. Nat. Rev. Immunol. 2012, 12, 503–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagy, I.; Pivarcsi, A.; Kis, K.; Koreck, A.; Bodai, L.; McDowell, A.; Seltmann, H.; Patrick, S.; Zouboulis, C.C.; Kemény, L. Propionibacterium acnes and lipopolysaccharide induce the expression of antimicrobial peptides and proinflammatory cytokines/chemokines in human sebocytes. Microbes Infect. 2006, 8, 2195–2205. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-Y.; Yamasaki, K.; Rudsil, J.; Zouboulis, C.C.; Park, G.T.; Yang, J.-M.; Gallo, R.L. Sebocytes Express Functional Cathelicidin Antimicrobial Peptides and Can Act to Kill Propionibacterium acnes. J. Investig. Dermatol. 2008, 128, 1863–1866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogen, A.L.; Yamasaki, K.; Muto, J.; Sanchez, K.M.; Alexander, L.C.; Tanios, J.; Lai, Y.; Kim, J.E.; Nizet, V.; Gallo, R.L. Staphylococcus epidermidis antimicrobial δ-toxin (phenol-soluble modulin-gamma) cooperates with host antimicrobial peptides to kill group A Streptococcus. PLoS ONE 2010, 5, e8557. [Google Scholar] [CrossRef]
- Cogen, A.L.; Yamasaki, K.; Sanchez, K.M.; Dorschner, R.A.; Lai, Y.; MacLeod, D.T.; Torpey, J.W.; Otto, M.; Nizet, V.; Kim, J.E.; et al. Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J. Investig. Derm. 2010, 130, 192–200. [Google Scholar] [CrossRef] [Green Version]
- Rothhammer, V.; Quintana, F.J. The aryl hydrocarbon receptor: An environmental sensor integrating immune responses in health and disease. Nat. Rev. Immunol. 2019, 19, 184–197. [Google Scholar] [CrossRef]
- Gutiérrez-Vázquez, C.; Quintana, F.J. Regulation of the Immune Response by the Aryl Hydrocarbon Receptor. Immunity 2018, 48, 19–33. [Google Scholar] [CrossRef] [Green Version]
- Stockinger, B.; Meglio, P.D.; Gialitakis, M.; Duarte, J.H. The Aryl Hydrocarbon Receptor: Multitasking in the Immune System. Annu. Rev. Immunol. 2014, 32, 403–432. [Google Scholar] [CrossRef]
- Poland, A.; Glover, E.; Kende, A.S. Stereospecific, high affinity binding of 2,3,7,8-tetrachlorodibenzo-p-dioxin by hepatic cytosol. Evidence that the binding species is receptor for induction of aryl hydrocarbon hydroxylase. J. Biol. Chem. 1976, 251, 4936–4946. [Google Scholar] [CrossRef]
- Denison, M.S.; Nagy, S.R. Activation of the Aryl Hydrocarbon Receptor by Structurally Diverse Exogenous and Endogenous Chemicals. Annu. Rev. Pharmacol. Toxicol. 2003, 43, 309–334. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.P.; Bradfield, C.A. The Search for Endogenous Activators of the Aryl Hydrocarbon Receptor. Chem. Res. Toxicol. 2008, 21, 102–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser, C.; Bargen, I.; Weighardt, H.; Haarmann-Stemmann, T.; Krutmann, J. Functions of the aryl hydrocarbon receptor in the skin. Semin. Immunopathol. 2013, 35, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hirota, K.; Westendorf, A.M.; Buer, J.; Dumoutier, L.; Renauld, J.-C.; Stockinger, B. The aryl hydrocarbon receptor links TH17-cell-mediated autoimmunity to environmental toxins. Nature 2008, 453, 106–109. [Google Scholar] [CrossRef] [PubMed]
- Thatcher, T.H.; Williams, M.A.; Pollock, S.J.; McCarthy, C.E.; Lacy, S.H.; Phipps, R.P.; Sime, P.J. Endogenous ligands of the aryl hydrocarbon receptor regulate lung dendritic cell function. Immunology 2016, 147, 41–54. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.; Jiang, Y.; Yang, Y.; Shao, J.; Sun, X.; Chen, J.; Dong, L.; Zhang, J. 3,3′-Diindolylmethane alleviates oxazolone-induced colitis through Th2/Th17 suppression and Treg induction. Mol. Immunol. 2013, 53, 335–344. [Google Scholar] [CrossRef]
- van den Bogaard, E.H.; Bergboer, J.G.M.; Vonk-Bergers, M.; van Vlijmen-Willems, I.M.J.J.; Hato, S.V.; van der Valk, P.G.M.; Schröder, J.M.; Joosten, I.; Zeeuwen, P.L.J.M.; Schalkwijk, J. Coal tar induces AHR-dependent skin barrier repair in atopic dermatitis. J. Clin. Investig. 2013, 123, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Colonna, M. AHR: Making the keratinocytes thick skinned. Immunity 2014, 40, 863–864. [Google Scholar] [CrossRef] [Green Version]
- Loertscher, J.A.; Sattler, C.A.; Allen-Hoffmann, B.L. 2,3,7,8-Tetrachlorodibenzo-p-dioxin alters the differentiation pattern of human keratinocytes in organotypic culture. Toxicol. Appl. Pharm. 2001, 175, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Sutter, C.H.; Yin, H.; Li, Y.; Mammen, J.S.; Bodreddigari, S.; Stevens, G.; Cole, J.A.; Sutter, T.R. EGF receptor signaling blocks aryl hydrocarbon receptor-mediated transcription and cell differentiation in human epidermal keratinocytes. Proc. Natl. Acad. Sci. USA 2009, 106, 4266–4271. [Google Scholar] [CrossRef] [Green Version]
- Furue, M.; Tsuji, G.; Mitoma, C.; Nakahara, T.; Chiba, T.; Morino-Koga, S.; Uchi, H. Gene regulation of filaggrin and other skin barrier proteins via aryl hydrocarbon receptor. J. Derm. Sci. 2015, 80, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Buommino, E.; Baroni, A.; Papulino, C.; Nocera, F.P.; Coretti, L.; Donnarumma, G.; De Filippis, A.; De Martino, L. Malassezia pachydermatis up-regulates AhR related CYP1A1 gene and epidermal barrier markers in human keratinocytes. Med. Mycol. 2018, 56, 987–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Luo, Y.; Zhu, Z.; Zhou, Y.; Sun, L.; Gao, J.; Sun, J.; Wang, G.; Yao, X.; Li, W. A tryptophan metabolite of the skin microbiota attenuates inflammation in patients with atopic dermatitis through the aryl hydrocarbon receptor. J. Allergy Clin. Immunol. 2019, 143, 2108–2119.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiatis, P.; Pappas, P.; Gaitanis, G.; Mexia, N.; Melliou, E.; Galanou, M.; Vlachos, C.; Stathopoulou, K.; Skaltsounis, A.L.; Marselos, M.; et al. Malassezia Yeasts Produce a Collection of Exceptionally Potent Activators of the Ah (Dioxin) Receptor Detected in Diseased Human Skin. J. Investig. Dermatol. 2013, 133, 2023–2030. [Google Scholar] [CrossRef] [Green Version]
- Haas, K.; Weighardt, H.; Deenen, R.; Kohrer, K.; Clausen, B.; Zahner, S.; Boukamp, P.; Bloch, W.; Krutmann, J.; Esser, C. Aryl Hydrocarbon Receptor in Keratinocytes Is Essential for Murine Skin Barrier Integrity. J. Investig. Derm. 2016, 136, 2260–2269. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.T.; Abrams, M.; Tlougan, B.; Rademaker, A.; Paller, A.S. Treatment of Staphylococcus aureus Colonization in Atopic Dermatitis Decreases Disease Severity. Pediatrics 2009, 123, e808–e814. [Google Scholar] [CrossRef] [Green Version]
- Bath-Hextall, F.J.; Birnie, A.J.; Ravenscroft, J.C.; Williams, H.C. Interventions to reduce Staphylococcus aureus in the management of atopic eczema: An updated Cochrane review. Br. J. Derm. 2010, 163, 12–26. [Google Scholar] [CrossRef]
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 1431. [Google Scholar] [CrossRef] [Green Version]
- Pastagia, M.; Schuch, R.; Fischetti, V.A.; Huang, D.B. Lysins: The arrival of pathogen-directed anti-infectives. J. Med. Microbiol. 2013, 62, 1506–1516. [Google Scholar] [CrossRef]
- Herpers, B. Endolysins: Redefining antibacterial therapy. Future Microbiol. 2015, 10, 309–311. [Google Scholar] [CrossRef] [Green Version]
- Totté, J.E.; van Doorn, M.B.; Pasmans, S.G. Successful treatment of chronic Staphylococcus aureus-related dermatoses with the topical endolysin Staphefekt SA. 100: A report of 3 cases. Case Rep. Dermatol. 2017, 9, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Scharschmidt, T.C.; Vasquez, K.S.; Truong, H.A.; Gearty, S.V.; Pauli, M.L.; Nosbaum, A.; Gratz, I.K.; Otto, M.; Moon, J.J.; Liese, J.; et al. A Wave of Regulatory T Cells into Neonatal Skin Mediates Tolerance to Commensal Microbes. Immunity 2015, 43, 1011–1021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmid-Wendtner, M.H.; Korting, H.C. The pH of the skin surface and its impact on the barrier function. Ski. Pharm. Physiol 2006, 19, 296–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matousek, J.L.; Campbell, K.L. A comparative review of cutaneous pH. Vet. Derm. 2002, 13, 293–300. [Google Scholar] [CrossRef]
- Korting, H.C.; Hubner, K.; Greiner, K.; Hamm, G.; Braun-Falco, O. Differences in the skin surface pH and bacterial microflora due to the long-term application of synthetic detergent preparations of pH 5.5 and pH 7.0. Results of a crossover trial in healthy volunteers. Acta Derm. Venereol. 1990, 70, 429–431. [Google Scholar] [PubMed]
- Sparavigna, A.; Setaro, M.; Gualandri, V. Cutaneous pH in children affected by atopic dermatitis and in healthy children: A multicenter study. Ski. Res. Technol. 1999, 5, 221–227. [Google Scholar] [CrossRef]
- Eberlein-Konig, B.; Schafer, T.; Huss-Marp, J.; Darsow, U.; Mohrenschlager, M.; Herbert, O.; Abeck, D.; Kramer, U.; Behrendt, H.; Ring, J. Skin surface pH, stratum corneum hydration, trans-epidermal water loss and skin roughness related to atopic eczema and skin dryness in a population of primary school children. Acta Derm. Venereol. 2000, 80, 188–191. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.J.; Yoon, N.Y.; Lee, N.R.; Jung, M.; Kim, D.H.; Choi, E.H. Topical acidic cream prevents the development of atopic dermatitis- and asthma-like lesions in murine model. Exp. Derm. 2014, 23, 736–741. [Google Scholar] [CrossRef]
- Lee, N.R.; Lee, H.J.; Yoon, N.Y.; Kim, D.; Jung, M.; Choi, E.H. Application of Topical Acids Improves Atopic Dermatitis in Murine Model by Enhancement of Skin Barrier Functions Regardless of the Origin of Acids. Ann. Derm. 2016, 28, 690–696. [Google Scholar] [CrossRef] [Green Version]
- Forno, E.; Onderdonk, A.B.; McCracken, J.; Litonjua, A.A.; Laskey, D.; Delaney, M.L.; DuBois, A.M.; Gold, D.R.; Ryan, L.M.; Weiss, S.T.; et al. Diversity of the gut microbiota and eczema in early life. Clin. Mol. Allergy 2008, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Karlsson, C.; Olsson, C.; Adlerberth, I.; Wold, A.E.; Strachan, D.P.; Martricardi, P.M.; Åberg, N.; Perkin, M.R.; Tripodi, S.; et al. Reduced diversity in the early fecal microbiota of infants with atopic eczema. J. Allergy Clin. Immunol. 2008, 121, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, H.; Li, N.; Bonnelykke, K.; Chawes, B.L.K.; Skov, T.; Paludan-Müller, G.; Stokholm, J.; Smith, B.; Krogfelt, K.A. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J. Allergy Clin. Immunol. 2011, 128, 646–652.e5. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, H.E.; Andersson, A.F.; Björkstén, B.; Engstrand, L.; Jenmalm, M.C. Low diversity of the gut microbiota in infants with atopic eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, I.H.; Oppedisano, F.; Joseph, S.J.; Boyle, R.J.; Licciardi, P.V.; Robins-Browne, R.M.; Tang, M.L.K. Reduced gut microbial diversity in early life is associated with later development of eczema but not atopy in high-risk infants. Pediatr. Allergy Immunol. 2012, 23, 674–681. [Google Scholar] [CrossRef]
- Nylund, L.; Nermes, M.; Isolauri, E.; Salminen, S.; de Vos, W.M.; Satokari, R. Severity of atopic disease inversely correlates with intestinal microbiota diversity and butyrate-producing bacteria. Allergy 2015, 70, 241–244. [Google Scholar] [CrossRef]
- Zimmermann, P.; Messina, N.; Mohn, W.W.; Finlay, B.B.; Curtis, N. Association between the intestinal microbiota and allergic sensitization, eczema, and asthma: A systematic review. J. Allergy Clin. Immunol. 2019, 143, 467–485. [Google Scholar] [CrossRef]
- Ye, S.; Yan, F.; Wang, H.; Mo, X.; Liu, J.; Zhang, Y.; Li, H.; Chen, D. Diversity analysis of gut microbiota between healthy controls and those with atopic dermatitis in a Chinese population. J. Dermatol. 2021, 48, 158–167. [Google Scholar] [CrossRef]
- Hu, C.; van Meel, E.R.; Medina-Gomez, C.; Kraaij, R.; Barroso, M.; Kiefte-de Jong, J.; Radjabzadeh, D.; Pasmans, S.G.M.A.; de Jong, N.W.; de Jongste, J.C.; et al. A population-based study on associations of stool microbiota with atopic diseases in school-age children. J. Allergy Clin. Immunol. 2021, 148, 612–620. [Google Scholar] [CrossRef]
- Song, H.; Yoo, Y.; Hwang, J.; Na, Y.-C.; Kim, H.S. Faecalibacterium prausnitzii subspecies-level dysbiosis in the human gut microbiome underlying atopic dermatitis. J. Allergy Clin. Immunol. 2016, 137, 852–860. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Li, Y.; Feng, X.; Li, Y.; Wang, W.; Qiu, C.; Xu, J.; Yang, Z.; Li, Z.; Zhou, Q.; et al. Dysfunctional gut microbiota and relative co-abundance network in infantile eczema. Gut Pathog. 2016, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.-J.; Kang, M.-J.; Lee, S.-Y.; Lee, E.; Kim, K.; Won, S.; Suh, D.I.; Kim, K.W.; Sheen, Y.H.; Ahn, K.; et al. Perturbations of gut microbiome genes in infants with atopic dermatitis according to feeding type. J. Allergy Clin. Immunol. 2018, 141, 1310–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cait, A.; Cardenas, E.; Dimitriu, P.A.; Amenyogbe, N.; Dai, D.; Cait, J.; Sbihi, H.; Stiemsma, L.; Subbarao, P.; Mandhane, P.J.; et al. Reduced genetic potential for butyrate fermentation in the gut microbiome of infants who develop allergic sensitization. J. Allergy Clin. Immunol. 2019, 144, 1638–1647.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkstén, B.; Naaber, P.; Sepp, E.; Mikelsaar, M. The intestinal microflora in allergic Estonian and Swedish 2-year-old children. Clin. Exp. Allergy 1999, 29, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Björkstén, B.; Sepp, E.; Julge, K.; Voor, T.; Mikelsaar, M. Allergy development and the intestinal microflora during the first year of life. J. Allergy Clin. Immunol. 2001, 108, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Adlerberth, I.; Strachan, D.P.; Matricardi, P.M.; Ahrné, S.; Orfei, L.; Åberg, N.; Perkin, M.R.; Tripodi, S.; Hesselmar, B.; Saalman, R.; et al. Gut microbiota and development of atopic eczema in 3 European birth cohorts. J. Allergy Clin. Immunol. 2007, 120, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Melli, L.C.F.L.; do Carmo-Rodrigues, M.S.; Araújo-Filho, H.B.; Mello, C.S.; Tahan, S.; Pignatari, A.C.C.; Solé, D.; de Morais, M.B. Gut microbiota of children with atopic dermatitis: Controlled study in the metropolitan region of São Paulo, Brazil. Allergol. Immunopathol. 2020, 48, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Penders, J.; Stobberingh, E.E.; Thijs, C.; Adams, H.; Vink, C.; Van Ree, R.; Van Den Brandt, P.A. Molecular fingerprinting of the intestinal microbiota of infants in whom atopic eczema was or was not developing. Clin. Exp. Allergy 2006, 36, 1602–1608. [Google Scholar] [CrossRef] [Green Version]
- Penders, J.; Thijs, C.; van den Brandt, P.A.; Kummeling, I.; Snijders, B.; Stelma, F.; Adams, H.; van Ree, R.; Stobberingh, E.E. Gut microbiota composition and development of atopic manifestations in infancy: The KOALA Birth Cohort Study. Gut 2007, 56, 661. [Google Scholar] [CrossRef] [Green Version]
- van Nimwegen, F.A.; Penders, J.; Stobberingh, E.E.; Postma, D.S.; Koppelman, G.H.; Kerkhof, M.; Reijmerink, N.E.; Dompeling, E.; van den Brandt, P.A.; Ferreira, I.; et al. Mode and place of delivery, gastrointestinal microbiota, and their influence on asthma and atopy. J. Allergy Clin. Immunol. 2011, 128, 948–955.e3. [Google Scholar] [CrossRef]
- Penders, J.; Gerhold, K.; Stobberingh, E.E.; Thijs, C.; Zimmermann, K.; Lau, S.; Hamelmann, E. Establishment of the intestinal microbiota and its role for atopic dermatitis in early childhood. J. Allergy Clin. Immunol. 2013, 132, 601–607.e8. [Google Scholar] [CrossRef]
- Zheng, H.; Liang, H.; Wang, Y.; Miao, M.; Shi, T.; Yang, F.; Liu, E.; Yuan, W.; Ji, Z.-S.; Li, D.-K. Altered Gut Microbiota Composition Associated with Eczema in Infants. PLoS ONE 2016, 11, e0166026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, E.; Lee, S.-Y.; Kang, M.-J.; Kim, K.; Won, S.; Kim, B.-J.; Choi, K.Y.; Kim, B.-S.; Cho, H.-J.; Kim, Y.; et al. Clostridia in the gut and onset of atopic dermatitis via eosinophilic inflammation. Ann. Allergy Asthma Immunol. 2016, 117, 91–92.e1. [Google Scholar] [CrossRef] [PubMed]
- Kull, I.; Böhme, M.; Wahlgren, C.F.; Nordvall, L.; Pershagen, G.; Wickman, M. Breast-feeding reduces the risk for childhood eczema. J. Allergy Clin. Immunol. 2005, 116, 657–661. [Google Scholar] [CrossRef] [PubMed]
- Ehlayel, M.S.; Bener, A. Duration of breast-feeding and the risk of childhood allergic diseases in a developing country. Allergy Asthma Proc. 2008, 29, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.Y.; Liao, S.L.; Su, K.W.; Tsai, M.H.; Hua, M.C.; Lai, S.H.; Chen, L.C.; Yao, T.C.; Yeh, K.W.; Huang, J.L. Exclusive or Partial Breastfeeding for 6 Months Is Associated With Reduced Milk Sensitization and Risk of Eczema in Early Childhood: The PATCH Birth Cohort Study. Medicine 2016, 95, e3391. [Google Scholar] [CrossRef] [PubMed]
- Gdalevich, M.; Mimouni, D.; David, M.; Mimouni, M. Breast-feeding and the onset of atopic dermatitis in childhood: A systematic review and meta-analysis of prospective studies. J. Am. Acad. Derm. 2001, 45, 520–527. [Google Scholar] [CrossRef]
- Schoetzau, A.; Filipiak-Pittroff, B.; Franke, K.; Koletzko, S.; Von Berg, A.; Gruebl, A.; Bauer, C.P.; Berdel, D.; Reinhardt, D.; Wichmann, H.E. Effect of exclusive breast-feeding and early solid food avoidance on the incidence of atopic dermatitis in high-risk infants at 1 year of age. Pediatr. Allergy Immunol. 2002, 13, 234–242. [Google Scholar] [CrossRef]
- Flohr, C.; Nagel, G.; Weinmayr, G.; Kleiner, A.; Strachan, D.P.; Williams, H.C. Lack of evidence for a protective effect of prolonged breastfeeding on childhood eczema: Lessons from the International Study of Asthma and Allergies in Childhood (ISAAC) Phase Two. Br. J. Derm. 2011, 165, 1280–1289. [Google Scholar] [CrossRef]
- Escherich, T. Die Darmbakterien des Säuglings und Ihre Beziehungen zur Physiologie der Verdauung; F. Enke: Stuttgart, Germany, 1886. [Google Scholar]
- Moro, E. Morphologie und bakteriologische Untersuchungen über die Darmbakterien des Säuglings: Die Bakterien-flora des normalen Frauenmilchstuhls. Jahrb. Kinderh 1900, 61, 686–734. [Google Scholar]
- Tissier, H. Recherches sur la Flore Intestinale des Nourrissons: (état Normal et Pathologique). Ph.D. Thesis, University of Paris, Paris, France, 1900. [Google Scholar]
- Harmsen, H.J.; Wildeboer-Veloo, A.C.; Raangs, G.C.; Wagendorp, A.A.; Klijn, N.; Bindels, J.G.; Welling, G.W. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 61–67. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Kirjavainen, P.; Eerola, E.; Kero, P.; Salminen, S.; Isolauri, E. Distinct patterns of neonatal gut microflora in infants in whom atopy was and was not developing. J. Allergy Clin. Immunol. 2001, 107, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Narisawa, Y.; Arase, S.; Okamatsu, H.; Ikenaga, T.; Tajiri, Y.; Kumemura, M. Differences in fecal microflora between patients with atopic dermatitis and healthy control subjects. J. Allergy Clin. Immunol. 2003, 111, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Mah, K.W.; Björkstén, B.; Lee, B.W.; van Bever, H.P.; Shek, L.P.; Tan, T.N.; Lee, Y.K.; Chua, K.Y. Distinct Pattern of Commensal Gut Microbiota in Toddlers with Eczema. Int. Arch. Allergy Immunol. 2006, 140, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.-Y.; Lee, B.W.; Aw, M.; Shek, L.P.C.; Yap, G.C.; Chua, K.Y.; Liu, W.-T. Comparative Analysis of Fecal Microbiota in Infants with and without Eczema. PLoS ONE 2010, 5, e9964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Shimojo, N.; Tajiri, Y.; Kumemura, M.; Kohno, Y. Differences in the composition of intestinal Bifidobacterium species and the development of allergic diseases in infants in rural Japan. Clin. Exp. Allergy 2007, 37, 506–511. [Google Scholar] [CrossRef]
- Gore, C.; Munro, K.; Lay, C.; Bibiloni, R.; Morris, J.; Woodcock, A.; Custovic, A.; Tannock, G.W. Bifidobacterium pseudocatenulatum is associated with atopic eczema: A nested case-control study investigating the fecal microbiota of infants. J. Allergy Clin. Immunol. 2008, 121, 135–140. [Google Scholar] [CrossRef]
- Roduit, C.; Frei, R.; Ferstl, R.; Loeliger, S.; Westermann, P.; Rhyner, C.; Schiavi, E.; Barcik, W.; Rodriguez-Perez, N.; Wawrzyniak, M.; et al. High levels of butyrate and propionate in early life are associated with protection against atopy. Allergy 2019, 74, 799–809. [Google Scholar] [CrossRef]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef] [Green Version]
- Zachariassen, L.F.; Krych, L.; Engkilde, K.; Nielsen, D.S.; Kot, W.; Hansen, C.H.F.; Hansen, A.K. Sensitivity to oxazolone induced dermatitis is transferable with gut microbiota in mice. Sci. Rep. 2017, 7, 44385. [Google Scholar] [CrossRef] [Green Version]
- McKenzie, C.; Tan, J.; Macia, L.; Mackay, C.R. The nutrition-gut microbiome-physiology axis and allergic diseases. Immunol. Rev. 2017, 278, 277–295. [Google Scholar] [CrossRef]
- Sestito, S.; D’Auria, E.; Baldassarre, M.E.; Salvatore, S.; Tallarico, V.; Stefanelli, E.; Tarsitano, F.; Concolino, D.; Pensabene, L. The Role of Prebiotics and Probiotics in Prevention of Allergic Diseases in Infants. Front. Pediatr. 2020, 8, 583946. [Google Scholar] [CrossRef]
- Cummings, J.H.; Macfarlane, G.T. The control and consequences of bacterial fermentation in the human colon. J. Appl. Bacteriol. 1991, 70, 443–459. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Macfarlane, G.T. Role of intestinal bacteria in nutrient metabolism. JPEN J. Parenter Enter. Nutr. 1997, 21, 357–365. [Google Scholar] [CrossRef]
- Tan, J.; McKenzie, C.; Potamitis, M.; Thorburn, A.N.; Mackay, C.R.; Macia, L. The role of short-chain fatty acids in health and disease. Adv. Immunol. 2014, 121, 91–119. [Google Scholar] [CrossRef]
- Thorburn, A.N.; Macia, L.; Mackay, C.R. Diet, metabolites, and “western-lifestyle” inflammatory diseases. Immunity 2014, 40, 833–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; McKenzie, C.; Vuillermin, P.J.; Goverse, G.; Vinuesa, C.G.; Mebius, R.E.; Macia, L.; Mackay, C.R. Dietary fiber and bacterial SCFA enhance oral tolerance and protect against food allergy through diverse cellular pathways. Cell Rep. 2016, 15, 2809–2824. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, A.; Bruhs, A.; Schwarz, T. The Short-Chain Fatty Acid Sodium Butyrate Functions as a Regulator of the Skin Immune System. J. Investig. Dermatol. 2017, 137, 855–864. [Google Scholar] [CrossRef] [Green Version]
- Tan, J.K.; McKenzie, C.; Marino, E.; Macia, L.; Mackay, C.R. Metabolite-Sensing G Protein-Coupled Receptors-Facilitators of Diet-Related Immune Regulation. Annu. Rev. Immunol. 2017, 35, 371–402. [Google Scholar] [CrossRef]
- Husted, A.S.; Trauelsen, M.; Rudenko, O.; Hjorth, S.A.; Schwartz, T.W. GPCR-Mediated Signaling of Metabolites. Cell Metab. 2017, 25, 777–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; van Esch, B.C.A.M.; Henricks, P.A.J.; Folkerts, G.; Garssen, J. The Anti-inflammatory Effects of Short Chain Fatty Acids on Lipopolysaccharide- or Tumor Necrosis Factor α-Stimulated Endothelial Cells via Activation of GPR41/43 and Inhibition of HDACs. Front. Pharmacol. 2018, 9, 533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [Green Version]
- Yap, Y.A.; McLeod, K.H.; McKenzie, C.I.; Gavin, P.G.; Davalos-Salas, M.; Richards, J.L.; Moore, R.J.; Lockett, T.J.; Clarke, J.M.; Eng, V.V.; et al. An acetate-yielding diet imprints an immune and anti-microbial programme against enteric infection. Clin. Transl. Immunol. 2021, 10, e1233. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of Prebiotic Fiber and Insufficient Signaling Through Gut Metabolite-Sensing Receptors Leads to Cardiovascular Disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Marino, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef]
- Patrick, M.S.; Michael, R.H.; Panikov, N.; Michaud, M.; Carey, A.G.; Bohlooly, Y.M.; Jonathan, N.G.; Wendy, S.G. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Mamontov, P.; Neiman, E.; Cao, T.; Perrigoue, J.; Friedman, J.; Das, A.; Mora, J. Effects of short chain fatty acids and GPR43 stimulation on human Treg function (IRC5P.631). J. Immunol. 2015, 194, 58.14. [Google Scholar]
- Sun, M.; Wu, W.; Chen, L.; Yang, W.; Huang, X.; Ma, C.; Chen, F.; Xiao, Y.; Zhao, Y.; Ma, C.; et al. Microbiota-derived short-chain fatty acids promote Th1 cell IL-10 production to maintain intestinal homeostasis. Nat. Commun. 2018, 9, 3555. [Google Scholar] [CrossRef] [Green Version]
- Alam, M.J.; Xie, L.; Ang, C.; Fahimi, F.; Willingham, S.B.; Kueh, A.J.; Herold, M.J.; Mackay, C.R.; Robert, R. Therapeutic blockade of CXCR2 rapidly clears inflammation in arthritis and atopic dermatitis models: Demonstration with surrogate and humanized antibodies. MAbs 2020, 12, 1856460. [Google Scholar] [CrossRef] [PubMed]
- Oyoshi, M.K.; He, R.; Li, Y.; Mondal, S.; Yoon, J.; Afshar, R.; Chen, M.; Lee, D.M.; Luo, H.R.; Luster, A.D.; et al. Leukotriene B4-driven neutrophil recruitment to the skin is essential for allergic skin inflammation. Immunity 2012, 37, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Walsh, C.M.; Hill, R.Z.; Schwendinger-Schreck, J.; Deguine, J.; Brock, E.C.; Kucirek, N.; Rifi, Z.; Wei, J.; Gronert, K.; Brem, R.B.; et al. Neutrophils promote CXCR3-dependent itch in the development of atopic dermatitis. elife 2019, 8, e48448. [Google Scholar] [CrossRef] [PubMed]
- Radermecker, C.; Louis, R.; Bureau, F.; Marichal, T. Role of neutrophils in allergic asthma. Curr. Opin. Immunol. 2018, 54, 28–34. [Google Scholar] [CrossRef]
- Johnston, L.K.; Chien, K.B.; Bryce, P.J. The Immunology of Food Allergy. J. Immunol. 2014, 192, 2529. [Google Scholar] [CrossRef] [Green Version]
- Vinolo, M.A.; Ferguson, G.J.; Kulkarni, S.; Damoulakis, G.; Anderson, K.; Bohlooly, Y.M.; Stephens, L.; Hawkins, P.T.; Curi, R. SCFAs induce mouse neutrophil chemotaxis through the GPR43 receptor. PLoS ONE 2011, 6, e21205. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [Green Version]
- Digby, J.E.; Martinez, F.; Jefferson, A.; Ruparelia, N.; Chai, J.; Wamil, M.; Greaves, D.R.; Choudhury, R.P. Anti-Inflammatory Effects of Nicotinic Acid in Human Monocytes Are Mediated by GPR109A Dependent Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 669–676. [Google Scholar] [CrossRef] [Green Version]
- Zandi-Nejad, K.; Takakura, A.; Jurewicz, M.; Chandraker, A.K.; Offermanns, S.; Mount, D.; Abdi, R. The role of HCA2 (GPR109A) in regulating macrophage function. FASEB J. 2013, 27, 4366–4374. [Google Scholar] [CrossRef] [Green Version]
- Digby, J.E.; McNeill, E.; Dyar, O.J.; Lam, V.; Greaves, D.R.; Choudhury, R.P. Anti-inflammatory effects of nicotinic acid in adipocytes demonstrated by suppression of fractalkine, RANTES, and MCP-1 and upregulation of adiponectin. Atherosclerosis 2010, 209, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Gambhir, D.; Ananth, S.; Veeranan-Karmegam, R.; Elangovan, S.; Hester, S.; Jennings, E.; Offermanns, S.; Nussbaum, J.J.; Smith, S.B.; Thangaraju, M.; et al. GPR109A as an anti-inflammatory receptor in retinal pigment epithelial cells and its relevance to diabetic retinopathy. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2208–2217. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Campbell, J.M.; Fahey, G.C., Jr.; Wolf, B.W. Selected indigestible oligosaccharides affect large bowel mass, cecal and fecal short-chain fatty acids, pH and microflora in rats. J. Nutr. 1997, 127, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Thomson, J.M.; Flint, H.J. The role of pH in determining the species composition of the human colonic microbiota. Environ. Microbiol. 2009, 11, 2112–2122. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; McKenzie, C.I.; Qu, X.; Mu, Y.; Wang, Q.; Bing, N.; Naidoo, K.; Alam, M.J.; Yu, D.; Gong, F.; et al. pH and Proton Sensor GPR65 Determine Susceptibility to Atopic Dermatitis. J. Immunol. 2021, 207, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Diogo, D.; Tian, C.; Franklin, C.S.; Alanne-Kinnunen, M.; March, M.; Spencer, C.C.A.; Vangjeli, C.; Weale, M.E.; Mattsson, H.; Kilpeläinen, E.; et al. Phenome-wide association studies across large population cohorts support drug target validation. Nat. Commun. 2018, 9, 4285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milazzo, G.; Mercatelli, D.; Di Muzio, G.; Triboli, L.; De Rosa, P.; Perini, G.; Giorgi, F.M. Histone Deacetylases (HDACs): Evolution, Specificity, Role in Transcriptional Complexes, and Pharmacological Actionability. Genes 2020, 11, 556. [Google Scholar] [CrossRef] [PubMed]
- Tost, J. Strengthening epithelial barriers through modulation of the histone code in allergic diseases—A novel approach for preventing the atopic march? J. Allergy Clin. Immunol. 2020, 146, 515–517. [Google Scholar] [CrossRef]
- Wawrzyniak, P.; Wawrzyniak, M.; Wanke, K.; Sokolowska, M.; Bendelja, K.; Rückert, B.; Globinska, A.; Jakiela, B.; Kast, J.I.; Idzko, M.; et al. Regulation of bronchial epithelial barrier integrity by type 2 cytokines and histone deacetylases in asthmatic patients. J. Allergy Clin. Immunol. 2017, 139, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-H.; Jung, J.-A.; Kim, G.-D.; Jang, A.-H.; Cho, J.-J.; Park, Y.S.; Park, C.-S. The histone deacetylase inhibitor, trichostatin A, inhibits the development of 2,4-dinitrofluorobenzene-induced dermatitis in NC/Nga mice. Int. Immunopharmacol. 2010, 10, 1310–1315. [Google Scholar] [CrossRef]
- Shi, Y.-L.; Gu, J.; Park, J.-Y.; Xu, Y.-P.; Yu, F.-S.; Zhou, L.; Mi, Q.-S. Histone deacetylases inhibitor Trichostatin A ameliorates DNFB-induced allergic contact dermatitis and reduces epidermal Langerhans cells in mice. J. Dermatol. Sci. 2012, 68, 99–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liew, W.C.; Sundaram, G.M.; Quah, S.; Lum, G.G.; Tan, J.S.L.; Ramalingam, R.; Common, J.E.A.; Tang, M.B.Y.; Lane, E.B.; Thng, S.T.G.; et al. Belinostat resolves skin barrier defects in atopic dermatitis by targeting the dysregulated miR-335:SOX6 axis. J. Allergy Clin. Immunol. 2020, 146, 606–620.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valapour, M.; Guo, J.; Schroeder, J.T.; Keen, J.; Cianferoni, A.; Casolaro, V.; Georas, S.N. Histone deacetylation inhibits IL4 gene expression in T cells. J. Allergy Clin. Immunol. 2002, 109, 238–245. [Google Scholar] [CrossRef] [PubMed]
- Boffa, L.C.; Vidali, G.; Mann, R.S.; Allfrey, V.G. Suppression of histone deacetylation in vivo and in vitro by sodium butyrate. J. Biol. Chem. 1978, 253, 3364–3366. [Google Scholar] [CrossRef]
- Sealy, L.; Chalkley, R. The effect of sodium butyrate on histone modification. Cell 1978, 14, 115–121. [Google Scholar] [CrossRef]
- Soliman, M.L.; Rosenberger, T.A. Acetate supplementation increases brain histone acetylation and inhibits histone deacetylase activity and expression. Mol. Cell. Biochem. 2011, 352, 173–180. [Google Scholar] [CrossRef]
- Hinnebusch, B.F.; Meng, S.; Wu, J.T.; Archer, S.Y.; Hodin, R.A. The Effects of Short-Chain Fatty Acids on Human Colon Cancer Cell Phenotype Are Associated with Histone Hyperacetylation. J. Nutr. 2002, 132, 1012–1017. [Google Scholar] [CrossRef] [Green Version]
- Waldecker, M.; Kautenburger, T.; Daumann, H.; Busch, C.; Schrenk, D. Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. J. Nutr. Biochem. 2008, 19, 587–593. [Google Scholar] [CrossRef]
- Kiefer, J.; Beyer-Sehlmeyer, G.; Pool-Zobel, B.L. Mixtures of SCFA, composed according to physiologically available concentrations in the gut lumen, modulate histone acetylation in human HT29 colon cancer cells. Br. J. Nutr. 2006, 96, 803–810. [Google Scholar] [CrossRef] [Green Version]
- Cousens, L.S.; Gallwitz, D.; Alberts, B.M. Different accessibilities in chromatin to histone acetylase. J. Biol. Chem. 1979, 254, 1716–1723. [Google Scholar] [CrossRef]
- Wu, J.; Zhou, Z.; Hu, Y.; Dong, S. Butyrate-induced GPR41 Activation Inhibits Histone Acetylation and Cell Growth. J. Genet. Genom. 2012, 39, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, S.F.W.; O’Boyle, G.; Mann, J.; Zeybel, M.; Palmer, J.; Jones, D.E.J.; Day, C.P. Acetate, the key modulator of inflammatory responses in acute alcoholic hepatitis. Hepatology 2010, 51, 1988–1997. [Google Scholar] [CrossRef] [PubMed]
- Usami, M.; Kishimoto, K.; Ohata, A.; Miyoshi, M.; Aoyama, M.; Fueda, Y.; Kotani, J. Butyrate and trichostatin A attenuate nuclear factor κB activation and tumor necrosis factor α secretion and increase prostaglandin E2 secretion in human peripheral blood mononuclear cells. Nutr. Res. 2008, 28, 321–328. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.L.; Mirshahpanah, P.; Haas-Stapleton, E.; Asadullah, K.; Zollner, T.M.; Numerof, R.P. Induction of Foxp3+ regulatory T cells with histone deacetylase inhibitors. Cell. Immunol. 2009, 257, 97–104. [Google Scholar] [CrossRef]
- Wang, L.; Tao, R.; Hancock, W.W. Using histone deacetylase inhibitors to enhance Foxp3+ regulatory T-cell function and induce allograft tolerance. Immunol. Cell Biol. 2009, 87, 195–202. [Google Scholar] [CrossRef]
- Tao, R.; de Zoeten, E.F.; Özkaynak, E.; Chen, C.; Wang, L.; Porrett, P.M.; Li, B.; Turka, L.A.; Olson, E.N.; Greene, M.I.; et al. Deacetylase inhibition promotes the generation and function of regulatory T cells. Nat. Med. 2007, 13, 1299–1307. [Google Scholar] [CrossRef]
- Huang, J.; Wang, L.; Dahiya, S.; Beier, U.H.; Han, R.; Samanta, A.; Bergman, J.; Sotomayor, E.M.; Seto, E.; Kozikowski, A.P.; et al. Histone/protein deacetylase 11 targeting promotes Foxp3+ Treg function. Sci. Rep. 2017, 7, 8626. [Google Scholar] [CrossRef] [Green Version]
- Dahiya, S.; Beier, U.H.; Wang, L.; Han, R.; Jiao, J.; Akimova, T.; Angelin, A.; Wallace, D.C.; Hancock, W.W. HDAC10 deletion promotes Foxp3+ T-regulatory cell function. Sci. Rep. 2020, 10, 424. [Google Scholar] [CrossRef] [Green Version]
- Kepert, I.; Fonseca, J.; Müller, C.; Milger, K.; Hochwind, K.; Kostric, M.; Fedoseeva, M.; Ohnmacht, C.; Dehmel, S.; Nathan, P.; et al. D-tryptophan from probiotic bacteria influences the gut microbiome and allergic airway disease. J. Allergy Clin. Immunol. 2017, 139, 1525–1535. [Google Scholar] [CrossRef] [Green Version]
- Jin, U.-H.; Lee, S.-O.; Sridharan, G.; Lee, K.; Davidson, L.A.; Jayaraman, A.; Chapkin, R.S.; Alaniz, R.; Safe, S. Microbiome-Derived Tryptophan Metabolites and Their Aryl Hydrocarbon Receptor-Dependent Agonist and Antagonist Activities. Mol. Pharmacol. 2014, 85, 777. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, M.T.; Carlson, J.R. Microbial metabolites of tryptophan in the intestinal tract with special reference to skatole. Am. J. Clin. Nutr. 1979, 32, 173–178. [Google Scholar] [CrossRef] [PubMed]
- Zelante, T.; Iannitti, R.G.; Cunha, C.; De Luca, A.; Giovannini, G.; Pieraccini, G.; Zecchi, R.; D’Angelo, C.; Massi-Benedetti, C.; Fallarino, F.; et al. Tryptophan Catabolites from Microbiota Engage Aryl Hydrocarbon Receptor and Balance Mucosal Reactivity via Interleukin-22. Immunity 2013, 39, 372–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yiu, J.H.; Dorweiler, B.; Woo, C.W. Interaction between gut microbiota and toll-like receptor: From immunity to metabolism. J. Mol. Med. 2017, 95, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, H.-C.; Feng, C.; Yan, M. Analysis of the Association of Polymorphisms rs5743708 in TLR2 and rs4986790 in TLR4 with Atopic Dermatitis Risk. Immunol. Investig. 2019, 48, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Belderbos, M.E.; Knol, E.F.; Houben, M.L.; van Bleek, G.M.; Wilbrink, B.; Kimpen, J.L.L.; Rovers, M.; Bont, L. Low neonatal Toll-like receptor 4-mediated interleukin-10 production is associated with subsequent atopic dermatitis. Clin. Exp. Allergy 2012, 42, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Hüls, A.; Klümper, C.; MacIntyre, E.A.; Brauer, M.; Melén, E.; Bauer, M.; Berdel, D.; Bergström, A.; Brunekreef, B.; Chan-Yeung, M.; et al. Atopic dermatitis: Interaction between genetic variants of GSTP1, TNF, TLR2, and TLR4 and air pollution in early life. Pediatr. Allergy Immunol. 2018, 29, 596–605. [Google Scholar] [CrossRef]
- Lin, L.; Xie, M.; Chen, X.; Yu, Y.; Liu, Y.; Lei, K.; Wang, D.; Zeng, J.; Zhou, J.; Zhang, L.; et al. Toll-like receptor 4 attenuates a murine model of atopic dermatitis through inhibition of langerin-positive DCs migration. Exp. Dermatol. 2018, 27, 1015–1022. [Google Scholar] [CrossRef]
- West, C.E.; Rydén, P.; Lundin, D.; Engstrand, L.; Tulic, M.K.; Prescott, S.L. Gut microbiome and innate immune response patterns in IgE-associated eczema. Clin. Exp. Allergy 2015, 45, 1419–1429. [Google Scholar] [CrossRef] [Green Version]
- Penders, J.; Thijs, C.; Mommers, M.; Stobberingh, E.E.; Dompeling, E.; Reijmerink, N.E.; van den Brandt, P.A.; Kerkhof, M.; Koppelman, G.H.; Postma, D.S. Host-microbial interactions in childhood atopy: Toll-like receptor 4 (TLR4), CD14, and fecal Escherichia coli. J. Allergy Clin. Immunol. 2010, 125, 231–236.e5. [Google Scholar] [CrossRef]
- van Nood, E.; Vrieze, A.; Nieuwdorp, M.; Fuentes, S.; Zoetendal, E.G.; de Vos, W.M.; Visser, C.E.; Kuijper, E.J.; Bartelsman, J.F.W.M.; Tijssen, J.G.P.; et al. Duodenal Infusion of Donor Feces for Recurrent Clostridium difficile. N. Engl. J. Med. 2013, 368, 407–415. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Kim, K.; Kim, W. Gut microbiota restoration through fecal microbiota transplantation: A new atopic dermatitis therapy. Exp. Mol. Med. 2021, 53, 907–916. [Google Scholar] [CrossRef] [PubMed]
- FDA. Important Safety Alert Regarding Use of Fecal Microbiota for Transplantation and Risk of Serious Adverse Reactions Due to Transmission of Multi-Drug Resistant Organisms. 2019. Available online: https://www.fda.gov/vaccines-blood-biologics/safety-availability-biologics/important-safety-alert-regarding-use-fecal-microbiota-transplantation-and-risk-serious-adverse (accessed on 13 June 2019).
- Baxter, M.; Ahmad, T.; Colville, A.; Sheridan, R. Fatal Aspiration Pneumonia as a Complication of Fecal Microbiota Transplant. Clin. Infect. Dis. 2015, 61, 136–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.R.; Ihunnah, C.; Fischer, M.; Khoruts, A.; Surawicz, C.; Afzali, A.; Aroniadis, O.; Barto, A.; Borody, T.; Giovanelli, A.; et al. Fecal Microbiota Transplant for Treatment of Clostridium difficileInfection in Immunocompromised Patients. Off. J. Am. Coll. Gastroenterol. ACG 2014, 109, 1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solari, P.R.; Fairchild, P.G.; Noa, L.J.; Wallace, M.R. Tempered Enthusiasm for Fecal Transplant. Clin. Infect. Dis. 2014, 59, 319. [Google Scholar] [CrossRef] [Green Version]
- Giles, E.M.; D’Adamo, G.L.; Forster, S.C. The future of faecal transplants. Nat. Rev. Microbiol. 2019, 17, 719. [Google Scholar] [CrossRef] [PubMed]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, C.J.; Ajami, N.J.; O’Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [Green Version]
- Chouraqui, J.P. Does the contribution of human milk oligosaccharides to the beneficial effects of breast milk allow us to hope for an improvement in infant formulas? Crit. Rev. Food Sci. Nutr. 2021, 61, 1503–1514. [Google Scholar] [CrossRef]
- Bode, L. Human milk oligosaccharides: Every baby needs a sugar mama. Glycobiology 2012, 22, 1147–1162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Li, T.; Xie, J.; Zhang, D.; Pi, C.; Zhou, L.; Yang, W. Gold standard for nutrition: A review of human milk oligosaccharide and its effects on infant gut microbiota. Microb. Cell Factories 2021, 20, 108. [Google Scholar] [CrossRef]
- Sela, D.A.; Mills, D.A. Nursing our microbiota: Molecular linkages between bifidobacteria and milk oligosaccharides. Trends Microbiol. 2010, 18, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Fonvig, C.E.; Amundsen, I.D.; Vigsnæs, L.K.; Sørensen, N.; Frithioff-Bøjsøe, C.; Christiansen, M.; Hedley, P.L.; Holm, L.A.; McConnell, B.; Holm, J.-C. Human Milk Oligosaccharides Modulate Fecal Microbiota and Are Safe for Use in Children with Overweight: A Randomized Controlled Trial. J. Pediatr. Gastroenterol. Nutr. 2021, 73, 408–414. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.A.; Sinn, J.K. Prebiotics in infants for prevention of allergy. Cochrane Database Syst. Rev. 2013, 3, Cd006474. [Google Scholar] [CrossRef] [PubMed]
- Moro, G.; Arslanoglu, S.; Stahl, B.; Jelinek, J.; Wahn, U.; Boehm, G. A mixture of prebiotic oligosaccharides reduces the incidence of atopic dermatitis during the first six months of age. Arch. Dis. Child. 2006, 91, 814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grüber, C.; van Stuijvenberg, M.; Mosca, F.; Moro, G.; Chirico, G.; Braegger, C.P.; Riedler, J.; Boehm, G.; Wahn, U. Reduced occurrence of early atopic dermatitis because of immunoactive prebiotics among low-atopy-risk infants. J. Allergy Clin. Immunol. 2010, 126, 791–797. [Google Scholar] [CrossRef]
- Grüber, C.; van Stuivenberg, M.; Mosca, F.; Moro, G.; Chirico, G.; Braegger, C.P.; Riedler, J.; Yavuz, Y.; Boehm, G.; Wahn, U. Immunoactive prebiotics transiently prevent occurrence of early atopic dermatitis among low-atopy-risk infants. J. Allergy Clin. Immunol. 2015, 136, 1696–1698.e1. [Google Scholar] [CrossRef]
- Pontes, M.V.; Ribeiro, T.C.M.; Ribeiro, H.; de Mattos, A.P.; Almeida, I.R.; Leal, V.M.; Cabral, G.N.; Stolz, S.; Zhuang, W.; Scalabrin, D.M.F. Cow’s milk-based beverage consumption in 1- to 4-year-olds and allergic manifestations: An RCT. Nutr. J. 2016, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Arslanoglu, S.; Moro, G.E.; Schmitt, J.; Tandoi, L.; Rizzardi, S.; Boehm, G. Early Dietary Intervention with a Mixture of Prebiotic Oligosaccharides Reduces the Incidence of Allergic Manifestations and Infections during the First Two Years of Life. J. Nutr. 2008, 138, 1091–1095. [Google Scholar] [CrossRef]
- Arslanoglu, S.; Moro, G.E.; Boehm, G.; Wienz, F.; Stahl, B.; Bertino, E. Early neutral prebiotic oligosaccharide supplementation reduces the incidence of some allergic manifestations in the first 5 years of life. J. Biol. Regul. Homeost. Agents 2012, 26, 49–59. [Google Scholar]
- Passeron, T.; Lacour, J.P.; Fontas, E.; Ortonne, J.P. Prebiotics and synbiotics: Two promising approaches for the treatment of atopic dermatitis in children above 2 years. Allergy 2006, 61, 431–437. [Google Scholar] [CrossRef]
- Niele, N.; van Zwol, A.; Westerbeek, E.A.M.; Lafeber, H.N.; van Elburg, R.M. Effect of non-human neutral and acidic oligosaccharides on allergic and infectious diseases in preterm infants. Eur. J. Pediatr. 2013, 172, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Ranucci, G.; Buccigrossi, V.; Borgia, E.; Piacentini, D.; Visentin, F.; Cantarutti, L.; Baiardi, P.; Felisi, M.; Spagnuolo, M.I.; Zanconato, S.; et al. Galacto-Oligosaccharide/Polidextrose Enriched Formula Protects against Respiratory Infections in Infants at High Risk of Atopy: A Randomized Clinical Trial. Nutrients 2018, 10, 286. [Google Scholar] [CrossRef] [Green Version]
- Boyle, R.J.; Tang, M.L.K.; Chiang, W.C.; Chua, M.C.; Ismail, I.; Nauta, A.; Hourihane, J.O.B.; Smith, P.; Gold, M.; Ziegler, J.; et al. Prebiotic-supplemented partially hydrolysed cow’s milk formula for the prevention of eczema in high-risk infants: A randomized controlled trial. Allergy 2016, 71, 701–710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, H.C.; Grindlay, D.J.C. What’s new in atopic eczema? An analysis of systematic reviews published in 2007 and 2008. Part 2. Disease prevention and treatment. Clin. Exp. Dermatol. 2010, 35, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Braegger, C.; Chmielewska, A.; Decsi, T.; Kolacek, S.; Mihatsch, W.; Moreno, L.; Piescik, M.; Puntis, J.; Shamir, R.; Szajewska, H.; et al. Supplementation of Infant Formula With Probiotics and/or Prebiotics: A Systematic Review and Comment by the ESPGHAN Committee on Nutrition. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 238–250. [Google Scholar] [CrossRef] [Green Version]
- Cuello-Garcia, C.A.; Fiocchi, A.; Pawankar, R.; Yepes-Nuñez, J.J.; Morgano, G.P.; Zhang, Y.; Ahn, K.; Al-Hammadi, S.; Agarwal, A.; Gandhi, S.; et al. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Prebiotics. World Allergy Organ. J. 2016, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Cuello-Garcia, C.; Fiocchi, A.; Pawankar, R.; Yepes-Nuñez, J.J.; Morgano, G.P.; Zhang, Y.; Agarwal, A.; Gandhi, S.; Terracciano, L.; Schünemann, H.J.; et al. Prebiotics for the prevention of allergies: A systematic review and meta-analysis of randomized controlled trials. Clin. Exp. Allergy 2017, 47, 1468–1477. [Google Scholar] [CrossRef]
- Vandenplas, Y. Prevention and Management of Cow’s Milk Allergy in Non-Exclusively Breastfed Infants. Nutrients 2017, 9, 731. [Google Scholar] [CrossRef]
- D’Auria, E.; Salvatore, S.; Pozzi, E.; Mantegazza, C.; Sartorio, M.U.A.; Pensabene, L.; Baldassarre, M.E.; Agosti, M.; Vandenplas, Y.; Zuccotti, G. Cow’s Milk Allergy: Immunomodulation by Dietary Intervention. Nutrients 2019, 11, 1399. [Google Scholar] [CrossRef] [Green Version]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dargahi, N.; Johnson, J.; Donkor, O.; Vasiljevic, T.; Apostolopoulos, V. Immunomodulatory effects of probiotics: Can they be used to treat allergies and autoimmune diseases? Maturitas 2019, 119, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.-S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef] [PubMed]
- Kalliomäki, M.; Salminen, S.; Arvilommi, H.; Kero, P.; Koskinen, P.; Isolauri, E. Probiotics in primary prevention of atopic disease: A randomised placebo-controlled trial. Lancet 2001, 357, 1076–1079. [Google Scholar] [CrossRef]
- Kalliomäki, M.; Salminen, S.; Poussa, T.; Arvilommi, H.; Isolauri, E. Probiotics and prevention of atopic disease: 4-year follow-up of a randomised placebo-controlled trial. Lancet 2003, 361, 1869–1871. [Google Scholar] [CrossRef]
- Wickens, K.; Black, P.N.; Stanley, T.V.; Mitchell, E.; Fitzharris, P.; Tannock, G.W.; Purdie, G.; Crane, J. A differential effect of 2 probiotics in the prevention of eczema and atopy: A double-blind, randomized, placebo-controlled trial. J. Allergy Clin. Immunol. 2008, 122, 788–794. [Google Scholar] [CrossRef] [PubMed]
- Wickens, K.; Black, P.; Stanley, T.V.; Mitchell, E.; Barthow, C.; Fitzharris, P.; Purdie, G.; Crane, J. A protective effect of Lactobacillus rhamnosus HN001 against eczema in the first 2 years of life persists to age 4 years. Clin. Exp. Allergy 2012, 42, 1071–1079. [Google Scholar] [CrossRef]
- Wickens, K.; Stanley, T.V.; Mitchell, E.A.; Barthow, C.; Fitzharris, P.; Purdie, G.; Siebers, R.; Black, P.N.; Crane, J. Early supplementation with Lactobacillus rhamnosus HN001 reduces eczema prevalence to 6 years: Does it also reduce atopic sensitization? Clin. Exp. Allergy 2013, 43, 1048–1057. [Google Scholar] [CrossRef]
- Wickens, K.; Barthow, C.; Mitchell, E.A.; Kang, J.; van Zyl, N.; Purdie, G.; Stanley, T.; Fitzharris, P.; Murphy, R.; Crane, J. Effects of Lactobacillus rhamnosus HN001 in early life on the cumulative prevalence of allergic disease to 11 years. Pediatr. Allergy Immunol. 2018, 29, 808–814. [Google Scholar] [CrossRef]
- Morgan, A.R.; Han, D.Y.; Wickens, K.; Barthow, C.; Mitchell, E.A.; Stanley, T.V.; Dekker, J.; Crane, J.; Ferguson, L.R. Differential modification of genetic susceptibility to childhood eczema by two probiotics. Clin. Exp. Allergy 2014, 44, 1255–1265. [Google Scholar] [CrossRef]
- Marlow, G.; Han, D.Y.; Wickens, K.; Stanley, T.; Crane, J.; Mitchell, E.A.; Dekker, J.; Barthow, C.; Fitzharris, P.; Ferguson, L.R.; et al. Differential effects of two probiotics on the risks of eczema and atopy associated with single nucleotide polymorphisms to Toll-like receptors. Pediatr. Allergy Immunol. 2015, 26, 262–271. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.; Morgan, X.C.; Wang, X.Y.; Wickens, K.; Purdie, G.; Fitzharris, P.; Otal, A.; Lawley, B.; Stanley, T.; Barthow, C.; et al. Eczema-protective probiotic alters infant gut microbiome functional capacity but not composition: Sub-sample analysis from a RCT. Benef. Microbes 2019, 10, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Dotterud, C.K.; Storrø, O.; Johnsen, R.; Øien, T. Probiotics in pregnant women to prevent allergic disease: A randomized, double-blind trial. Br. J. Dermatol. 2010, 163, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Simpson, M.R.; Dotterud, C.K.; Storrø, O.; Johnsen, R.; Øien, T. Perinatal probiotic supplementation in the prevention of allergy related disease: 6 year follow up of a randomised controlled trial. BMC Dermatol. 2015, 15, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rø, A.D.B.; Simpson, M.R.; Rø, T.B.; Storrø, O.; Johnsen, R.; Videm, V.; Øien, T. Reduced Th22 cell proportion and prevention of atopic dermatitis in infants following maternal probiotic supplementation. Clin. Exp. Allergy 2017, 47, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Kwon, J.H.; Ahn, S.H.; Lee, S.I.; Han, Y.S.; Choi, Y.O.; Lee, S.Y.; Ahn, K.M.; Ji, G.E. Effect of probiotic mix (Bifidobacterium bifidum, Bifidobacterium lactis, Lactobacillus acidophilus) in the primary prevention of eczema: A double-blind, randomized, placebo-controlled trial. Pediatr. Allergy Immunol. 2010, 21, e386–e393. [Google Scholar] [CrossRef]
- Enomoto, T.; Sowa, M.; Nishimori, K.; Shimazu, S.; Yoshida, A.; Yamada, K.; Furukawa, F.; Nakagawa, T.; Yanagisawa, N.; Iwabuchi, N.; et al. Effects of Bifidobacterial Supplementation to Pregnant Women and Infants in the Prevention of Allergy Development in Infants and on Fecal Microbiota. Allergol. Int. 2014, 63, 575–585. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.L.; Dunstan, J.A.; Prescott, S.L. Probiotic supplementation for the first 6 months of life fails to reduce the risk of atopic dermatitis and increases the risk of allergen sensitization in high-risk children: A randomized controlled trial. J. Allergy Clin. Immunol. 2007, 119, 184–191. [Google Scholar] [CrossRef]
- Kopp, M.V.; Hennemuth, I.; Heinzmann, A.; Urbanek, R. Randomized, Double-Blind, Placebo-Controlled Trial of Probiotics for Primary Prevention: No Clinical Effects of Lactobacillus GG Supplementation. Pediatrics 2008, 121, e850. [Google Scholar] [CrossRef]
- Soh, S.E.; Aw, M.; Gerez, I.; Chong, Y.S.; Rauff, M.; Ng, Y.P.M.; Wong, H.B.; Pai, N.; Lee, B.W.; Shek, L.P.C. Probiotic supplementation in the first 6 months of life in at risk Asian infants—Effects on eczema and atopic sensitization at the age of 1 year. Clin. Exp. Allergy 2009, 39, 571–578. [Google Scholar] [CrossRef]
- Boyle, R.J.; Ismail, I.H.; Kivivuori, S.; Licciardi, P.V.; Robins-Browne, R.M.; Mah, L.J.; Axelrad, C.; Moore, S.; Donath, S.; Carlin, J.B.; et al. Lactobacillus GG treatment during pregnancy for the prevention of eczema: A randomized controlled trial. Allergy 2011, 66, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Wickens, K.; Barthow, C.; Mitchell, E.A.; Stanley, T.V.; Purdie, G.; Rowden, J.; Kang, J.; Hood, F.; van den Elsen, L.; Forbes-Blom, E.; et al. Maternal supplementation alone with Lactobacillus rhamnosus HN001 during pregnancy and breastfeeding does not reduce infant eczema. Pediatr. Allergy Immunol. 2018, 29, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Avershina, E.; Rubio, R.C.; Lundgård, K.; Martinez, G.P.; Collado, M.C.; Storrø, O.; Øien, T.; Dotterud, C.K.; Johnsen, R.; Rudi, K. Effect of probiotics in prevention of atopic dermatitis is dependent on the intrinsic microbiota at early infancy. J. Allergy Clin. Immunol. 2017, 139, 1399–1402.e8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foolad, N.; Brezinski, E.A.; Chase, E.P.; Armstrong, A.W. Effect of Nutrient Supplementation on Atopic Dermatitis in Children: A Systematic Review of Probiotics, Prebiotics, Formula, and Fatty Acids. JAMA Dermatol. 2013, 149, 350–355. [Google Scholar] [CrossRef]
- Pelucchi, C.; Chatenoud, L.; Turati, F.; Galeone, C.; Moja, L.; Bach, J.-F.; La Vecchia, C. Probiotics Supplementation During Pregnancy or Infancy for the Prevention of Atopic Dermatitis: A Meta-analysis. Epidemiology 2012, 23, 402–414. [Google Scholar] [CrossRef]
- Lee, J.; Seto, D.; Bielory, L. Meta-analysis of clinical trials of probiotics for prevention and treatment of pediatric atopic dermatitis. J. Allergy Clin. Immunol. 2008, 121, 116–121.e11. [Google Scholar] [CrossRef]
- Mansfield, J.A.; Bergin, S.W.; Cooper, J.R.; Olsen, C.H. Comparative Probiotic Strain Efficacy in the Prevention of Eczema in Infants and Children: A Systematic Review and Meta-Analysis. Mil. Med. 2014, 179, 580–592. [Google Scholar] [CrossRef] [Green Version]
- Elazab, N.; Mendy, A.; Gasana, J.; Vieira, E.R.; Quizon, A.; Forno, E. Probiotic Administration in Early Life, Atopy, and Asthma: A Meta-analysis of Clinical Trials. Pediatrics 2013, 132, e666. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Han, Z.; Niu, X.; Zhang, G.; Jia, Y.; Zhang, S.; He, C. Probiotic Supplementation for Prevention of Atopic Dermatitis in Infants and Children: A Systematic Review and Meta-analysis. Am. J. Clin. Dermatol. 2019, 20, 367–377. [Google Scholar] [CrossRef]
- Tan-Lim, C.S.C.; Esteban-Ipac, N.A.R.; Recto, M.S.T.; Castor, M.A.R.; Casis-Hao, R.J.; Nano, A.L.M. Comparative effectiveness of probiotic strains on the prevention of pediatric atopic dermatitis: A systematic review and network meta-analysis. Pediatr. Allergy Immunol. 2021, 32, 1255–1270. [Google Scholar] [CrossRef]
- Doege, K.; Grajecki, D.; Zyriax, B.-C.; Detinkina, E.; zu Eulenburg, C.; Buhling, K.J. Impact of maternal supplementation with probiotics during pregnancy on atopic eczema in childhood—A meta-analysis. Br. J. Nutr. 2012, 107, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Panduru, M.; Panduru, N.M.; Sălăvăstru, C.M.; Tiplica, G.S. Probiotics and primary prevention of atopic dermatitis: A meta-analysis of randomized controlled studies. J. Eur. Acad. Dermatol. Venereol. 2015, 29, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.L.; Yang, W.X.; Yang, H.M. Meta analysis of lactic acid bacteria as probiotics for the primary prevention of infantile eczema. Zhongguo Dang Dai Er Ke Za Zhi Chin. J. Contemp. Pediatr. 2010, 12, 734–739. [Google Scholar]
- Tang, L.J.; Chen, J.; Shen, Y. Meta-analysis of probiotics preventing allergic diseases in infants. Zhonghua Er Ke Za Zhi Chin. J. Pediatr. 2012, 50, 504–509. [Google Scholar]
- Cao, L.; Wang, L.; Yang, L.; Tao, S.; Xia, R.; Fan, W. Long-term effect of early-life supplementation with probiotics on preventing atopic dermatitis: A meta-analysis. J. Dermatol. Treat. 2015, 26, 537–540. [Google Scholar] [CrossRef]
- Amalia, N.; Orchard, D.; Francis, K.L.; King, E. Systematic review and meta-analysis on the use of probiotic supplementation in pregnant mother, breastfeeding mother and infant for the prevention of atopic dermatitis in children. Australas. J. Dermatol. 2020, 61, e158–e173. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.G.; He, Z.; Duan, X.Y.; Fan, F.X.; Liao, X.B.; Wang, Q.C. Effect of probiotic supplementation during pregnancy and infancy in preventing atopic dermatitis in children: A Meta analysis. Zhongguo Dang Dai Er Ke Za Zhi Chin. J. Contemp. Pediatr. 2019, 21, 82–88. [Google Scholar] [CrossRef]
- Jiang, W.; Ni, B.; Liu, Z.; Liu, X.; Xie, W.; Wu, I.X.Y.; Li, X. The Role of Probiotics in the Prevention and Treatment of Atopic Dermatitis in Children: An Updated Systematic Review and Meta-Analysis of Randomized Controlled Trials. Pediatr. Drugs 2020, 22, 535–549. [Google Scholar] [CrossRef]
- Fiocchi, A.; Pawankar, R.; Cuello-Garcia, C.; Ahn, K.; Al-Hammadi, S.; Agarwal, A.; Beyer, K.; Burks, W.; Canonica, G.W.; Ebisawa, M.; et al. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Probiotics. World Allergy Organ. J. 2015, 8, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Isolauri, E.; Arvola, T.; Sütas, Y.; Moilanen, E.; Salminen, S. Probiotics in the management of atopic eczema. Clin. Exp. Allergy 2000, 30, 1605–1610. [Google Scholar] [CrossRef] [Green Version]
- Kirjavainen, P.V.; Salminen, S.J.; Isolauri, E. Probiotic Bacteria in the Management of Atopic Disease: Underscoring the Importance of Viability. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Brouwer, M.L.; Wolt-Plompen, S.A.A.; Dubois, A.E.J.; Van Der Heide, S.; Jansen, D.F.; Hoijer, M.A.; Kauffman, H.F.; Duiverman, E.J. No effects of probiotics on atopic dermatitis in infancy: A randomized placebo-controlled trial. Clin. Exp. Allergy 2006, 36, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Fölster-Holst, R.; Müller, F.; Schnopp, N.; Abeck, D.; Kreiselmaier, I.; Lenz, T.; Von Rüden, U.; Schrezenmeir, J.; Christophers, E.; Weichenthal, M. Prospective, randomized controlled trial on Lactobacillus rhamnosus in infants with moderate to severe atopic dermatitis. Br. J. Dermatol. 2006, 155, 1256–1261. [Google Scholar] [CrossRef] [PubMed]
- Grüber, C.; Wendt, M.; Sulser, C.; Lau, S.; Kulig, M.; Wahn, U.; Werfel, T.; Niggemann, B. Randomized, placebo-controlled trial of Lactobacillus rhamnosus GG as treatment of atopic dermatitis in infancy. Allergy 2007, 62, 1270–1276. [Google Scholar] [CrossRef]
- Wu, Y.-J.; Wu, W.-F.; Hung, C.-W.; Ku, M.-S.; Liao, P.-F.; Sun, H.-L.; Lu, K.-H.; Sheu, J.-N.; Lue, K.-H. Evaluation of efficacy and safety of Lactobacillus rhamnosus in children aged 4–48 months with atopic dermatitis: An 8-week, double-blind, randomized, placebo-controlled study. J. Microbiol. Immunol. Infect. 2017, 50, 684–692. [Google Scholar] [CrossRef] [Green Version]
- Cukrowska, B.; Ceregra, A.; Maciorkowska, E.; Surowska, B.; Zegadło-Mylik, M.A.; Konopka, E.; Trojanowska, I.; Zakrzewska, M.; Bierła, J.B.; Zakrzewski, M.; et al. The Effectiveness of Probiotic Lactobacillus rhamnosus and Lactobacillus casei Strains in Children with Atopic Dermatitis and Cow’s Milk Protein Allergy: A Multicenter, Randomized, Double Blind, Placebo Controlled Study. Nutrients 2021, 13, 1169. [Google Scholar] [CrossRef]
- Drago, L.; Iemoli, E.; Rodighiero, V.; Nicola, L.; De Vecchi, E.; Piconi, S. Effects of Lactobacillus salivarius LS01 (DSM 22775) Treatment on Adult Atopic Dermatitis: A Randomized Placebo-Controlled Study. Int. J. Immunopathol. Pharmacol. 2011, 24, 1037–1048. [Google Scholar] [CrossRef]
- Drago, L.; Toscano, M.; De Vecchi, E.; Piconi, S.; Iemoli, E. Changing of fecal flora and clinical effect of L. salivarius LS01 in adults with atopic dermatitis. J. Clin. Gastroenterol. 2012, 46, S56–S63. [Google Scholar] [CrossRef]
- Weston, S.; Halbert, A.; Richmond, P.; Prescott, S.L. Effects of probiotics on atopic dermatitis: A randomised controlled trial. Arch. Dis. Child. 2005, 90, 892. [Google Scholar] [CrossRef]
- Wang, I.J.; Wang, J.Y. Children with atopic dermatitis show clinical improvement after Lactobacillus exposure. Clin. Exp. Allergy 2015, 45, 779–787. [Google Scholar] [CrossRef]
- Woo, S.-I.; Kim, J.-Y.; Lee, Y.-J.; Kim, N.-S.; Hahn, Y.-S. Effect of Lactobacillus sakei supplementation in children with atopic eczema–dermatitis syndrome. Ann. Allergy Asthma Immunol. 2010, 104, 343–348. [Google Scholar] [CrossRef]
- Prakoeswa, C.R.S.; Herwanto, N.; Prameswari, R.; Astari, L.; Sawitri, S.; Hidayati, A.N.; Indramaya, D.M.; Kusumowidagdo, E.R.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation reduced SCORAD in children with atopic dermatitis. Benef. Microbes 2017, 8, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Kim, B.; Ban, J.; Lee, J.; Kim, B.J.; Choi, B.S.; Hwang, S.; Ahn, K.; Kim, J. A randomized trial of Lactobacillus plantarum CJLP133 for the treatment of atopic dermatitis. Pediatr. Allergy Immunol. 2012, 23, 667–673. [Google Scholar] [CrossRef]
- Fang, Z.; Lu, W.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; Chen, W. Probiotics modulate the gut microbiota composition and immune responses in patients with atopic dermatitis: A pilot study. Eur. J. Nutr. 2020, 59, 2119–2130. [Google Scholar] [CrossRef]
- Gore, C.; Custovic, A.; Tannock, G.W.; Munro, K.; Kerry, G.; Johnson, K.; Peterson, C.; Morris, J.; Chaloner, C.; Murray, C.S.; et al. Treatment and secondary prevention effects of the probiotics Lactobacillus paracasei or Bifidobacterium lactis on early infant eczema: Randomized controlled trial with follow-up until age 3 years. Clin. Exp. Allergy 2012, 42, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Rosenfeldt, V.; Benfeldt, E.; Nielsen, S.D.; Michaelsen, K.F.; Jeppesen, D.L.; Valerius, N.H.; Paerregaard, A. Effect of probiotic Lactobacillus strains in children with atopic dermatitis. J. Allergy Clin. Immunol. 2003, 111, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Michelotti, A.; Cestone, E.; De Ponti, I.; Giardina, S.; Pisati, M.; Spartà, E.; Tursi, F. Efficacy of a probiotic supplement in patients with atopic dermatitis: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Dermatol. 2021, 31, 225–232. [Google Scholar] [CrossRef]
- Matsumoto, M.; Ebata, T.; Hirooka, J.; Hosoya, R.; Inoue, N.; Itami, S.; Tsuji, K.; Yaginuma, T.; Muramatsu, K.; Nakamura, A.; et al. Antipruritic effects of the probiotic strain LKM512 in adults with atopic dermatitis. Ann. Allergy Asthma Immunol. 2014, 113, 209–216.e7. [Google Scholar] [CrossRef]
- Iemoli, E.; Trabattoni, D.; Parisotto, S.; Borgonovo, L.; Toscano, M.; Rizzardini, G.; Clerici, M.; Ricci, E.; Fusi, A.; De Vecchi, E.; et al. Probiotics reduce gut microbial translocation and improve adult atopic dermatitis. J. Clin. Gastroenterol. 2012, 46, S33–S40. [Google Scholar] [CrossRef]
- Navarro-López, V.; Ramírez-Boscá, A.; Ramón-Vidal, D.; Ruzafa-Costas, B.; Genovés-Martínez, S.; Chenoll-Cuadros, E.; Carrión-Gutiérrez, M.; de la Parte, J.H.; Prieto-Merino, D.; Codoñer-Cortés, F.M. Effect of Oral Administration of a Mixture of Probiotic Strains on SCORAD Index and Use of Topical Steroids in Young Patients with Moderate Atopic Dermatitis: A Randomized Clinical Trial. JAMA Dermatol. 2018, 154, 37–43. [Google Scholar] [CrossRef]
- Drago, L.; De Vecchi, E.; Toscano, M.; Vassena, C.; Altomare, G.; Pigatto, P. Treatment of atopic dermatitis eczema with a high concentration of Lactobacillus salivarius LS01 associated with an innovative gelling complex: A pilot study on adults. J. Clin. Gastroenterol. 2014, 48 (Suppl. 1), S47–S51. [Google Scholar] [CrossRef] [PubMed]
- Ahn, S.H.; Yoon, W.; Lee, S.Y.; Shin, H.S.; Lim, M.Y.; Nam, Y.-D.; Yoo, Y. Effects of Lactobacillus pentosus in Children with Allergen-Sensitized Atopic Dermatitis. J. Korean Med. Sci. 2020, 35, e128. [Google Scholar] [CrossRef] [PubMed]
- Viljanen, M.; Savilahti, E.; Haahtela, T.; Juntunen-Backman, K.; Korpela, R.; Poussa, T.; Tuure, T.; Kuitunen, M. Probiotics in the treatment of atopic eczema/dermatitis syndrome in infants: A double-blind placebo-controlled trial. Allergy 2005, 60, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Sistek, D.; Kelly, R.; Wickens, K.; Stanley, T.; Fitzharris, P.; Crane, J. Is the effect of probiotics on atopic dermatitis confined to food sensitized children? Clin. Exp. Allergy 2006, 36, 629–633. [Google Scholar] [CrossRef] [PubMed]
- da Costa Baptista, I.P.; Accioly, E.; de Carvalho Padilha, P. Effect of the use of probiotics in the treatment of children with atopic dermatitis; a literature review. Nutr. Hosp. 2013, 28, 16–26. [Google Scholar] [CrossRef]
- Huang, R.; Ning, H.; Shen, M.; Li, J.; Zhang, J.; Chen, X. Probiotics for the Treatment of Atopic Dermatitis in Children: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Front. Cell. Infect. Microbiol. 2017, 7, 392. [Google Scholar] [CrossRef]
- Zhao, M.; Shen, C.; Ma, L. Treatment efficacy of probiotics on atopic dermatitis, zooming in on infants: A systematic review and meta-analysis. Int. J. Dermatol. 2018, 57, 635–641. [Google Scholar] [CrossRef]
- Makrgeorgou, A.; Leonardi-Bee, J.; Bath-Hextall, F.J.; Murrell, D.F.; Tang, M.L.K.; Roberts, A.; Boyle, R.J. Probiotics for treating eczema. Cochrane Database Syst. Rev. 2018, 11, CD006135. [Google Scholar] [CrossRef]
- Michail, S.K.; Stolfi, A.; Johnson, T.; Onady, G.M. Efficacy of probiotics in the treatment of pediatric atopic dermatitis: A meta-analysis of randomized controlled trials. Ann. Allergy Asthma Immunol. 2008, 101, 508–516. [Google Scholar] [CrossRef]
- Kim, S.-O.; Ah, Y.-M.; Yu, Y.M.; Choi, K.H.; Shin, W.-G.; Lee, J.-Y. Effects of probiotics for the treatment of atopic dermatitis: A meta-analysis of randomized controlled trials. Ann. Allergy Asthma Immunol. 2014, 113, 217–226. [Google Scholar] [CrossRef]
- Swanson, K.S.; Gibson, G.R.; Hutkins, R.; Reimer, R.A.; Reid, G.; Verbeke, K.; Scott, K.P.; Holscher, H.D.; Azad, M.B.; Delzenne, N.M.; et al. The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 687–701. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, A.; Moin, M.; Pourpak, Z.; Gharagozlou, M.; Aghamohammadi, A.; Sajedi, V.; Soheili, H.; Sotoodeh, S.; Movahedi, M. Synbiotics could not reduce the scoring of childhood atopic dermatitis (SCORAD): A randomized double blind placebo-controlled trial. Iran. J. Allergy Asthma Immunol. 2011, 10, 21–28. [Google Scholar] [PubMed]
- Chang, Y.-S.; Trivedi, M.K.; Jha, A.; Lin, Y.-F.; Dimaano, L.; García-Romero, M.T. Synbiotics for Prevention and Treatment of Atopic Dermatitis: A Meta-analysis of Randomized Clinical Trials. JAMA Pediatr. 2016, 170, 236–242. [Google Scholar] [CrossRef] [PubMed]
- Gerasimov, S.V.; Vasjuta, V.V.; Myhovych, O.O.; Bondarchuk, L.I. Probiotic Supplement Reduces Atopic Dermatitis in Preschool Children. Am. J. Clin. Dermatol. 2010, 11, 351–361. [Google Scholar] [CrossRef]
- Van Der Aa, L.B.; Heymans, H.S.; Van Aalderen, W.M.; Sillevis Smitt, J.H.; Knol, J.; Ben Amor, K.; Goossens, D.A.; Sprikkelman, A.B.; The Synbad Study Group. Effect of a new synbiotic mixture on atopic dermatitis in infants: A randomized-controlled trial. Clin. Exp. Allergy 2010, 40, 795–804. [Google Scholar] [CrossRef]
- Ogawa, T.; Hashikawa, S.; Asai, Y.; Sakamoto, H.; Yasuda, K.; Makimura, Y. A new synbiotic, Lactobacillus casei subsp. casei together with dextran, reduces murine and human allergic reaction. FEMS Immunol. Med. Microbiol. 2006, 46, 400–409. [Google Scholar] [CrossRef] [Green Version]
- van der Aa, L.B.; Lutter, R.; Heymans, H.S.A.; Smids, B.S.; Dekker, T.; van Aalderen, W.M.C.; Smitt, J.H.S.; Knippels, L.M.J.; Garssen, J.; Nauta, A.J.; et al. No detectable beneficial systemic immunomodulatory effects of a specific synbiotic mixture in infants with atopic dermatitis. Clin. Exp. Allergy 2012, 42, 531–539. [Google Scholar] [CrossRef]
- Wu, K.G.; Li, T.H.; Peng, H.J. Lactobacillus salivarius plus fructo-oligosaccharide is superior to fructo-oligosaccharide alone for treating children with moderate to severe atopic dermatitis: A double-blind, randomized, clinical trial of efficacy and safety. Br. J. Dermatol. 2012, 166, 129–136. [Google Scholar] [CrossRef]
- Salminen, S.; Collado, M.C.; Endo, A.; Hill, C.; Lebeer, S.; Quigley, E.M.M.; Sanders, M.E.; Shamir, R.; Swann, J.R.; Szajewska, H.; et al. The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 649–667. [Google Scholar] [CrossRef]
- Yan, D.C.; Hung, C.H.; Sy, L.B.; Lue, K.H.; Shih, I.H.; Yang, C.Y.; Chen, L.C.; Sun, H.L.; Lee, M.S.; Chambard, J.; et al. A Randomized, Double-Blind, Placebo-Controlled Trial Assessing the Oral Administration of a Heat-Treated Lactobacillus paracasei Supplement in Infants with Atopic Dermatitis Receiving Topical Corticosteroid Therapy. Ski. Pharmacol. Physiol. 2019, 32, 201–211. [Google Scholar] [CrossRef]
- Jeong, K.; Kim, M.; Jeon, S.A.; Kim, Y.-H.; Lee, S. A randomized trial of Lactobacillus rhamnosus IDCC 3201 tyndallizate (RHT3201) for treating atopic dermatitis. Pediatr. Allergy Immunol. 2020, 31, 783–792. [Google Scholar] [CrossRef] [PubMed]
- Moroi, M.; Uchi, S.; Nakamura, K.; Sato, S.; Shimizu, N.; Fujii, M.; Kumagai, T.; Saito, M.; Uchiyama, K.; Watanabe, T.; et al. Beneficial effect of a diet containing heat-killed Lactobacillus paracasei K71 on adult type atopic dermatitis. J. Dermatol. 2011, 38, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Torii, S.; Torii, A.; Itoh, K.; Urisu, A.; Terada, A.; Fujisawa, T.; Yamada, K.; Suzuki, H.; Ishida, Y.; Nakamura, F.; et al. Effects of Oral Administration of Lactobacillus acidophilus L-92 on the Symptoms and Serum Markers of Atopic Dermatitis in Children. Int. Arch. Allergy Immunol. 2011, 154, 236–245. [Google Scholar] [CrossRef]
- Inoue, Y.; Kambara, T.; Murata, N.; Komori-Yamaguchi, J.; Matsukura, S.; Takahashi, Y.; Ikezawa, Z.; Aihara, M. Effects of Oral Administration of Lactobacillus acidophilus L-92 on the Symptoms and Serum Cytokines of Atopic Dermatitis in Japanese Adults: A Double-Blind, Randomized, Clinical Trial. Int. Arch. Allergy Immunol. 2014, 165, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Gueniche, A.; Knaudt, B.; Schuck, E.; Volz, T.; Bastien, P.; Martin, R.; Röcken, M.; Breton, L.; Biedermann, T. Effects of nonpathogenic gram-negative bacterium Vitreoscilla filiformis lysate on atopic dermatitis: A prospective, randomized, double-blind, placebo-controlled clinical study. Br. J. Dermatol. 2008, 159, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yosipovitch, G. The Role of the Microbiome and Microbiome-Derived Metabolites in Atopic Dermatitis and Non-Histaminergic Itch. Am. J. Clin. Dermatol. 2020, 21, 44–50. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mechanisms of how skin microbiota regulates AD pathogenesis. Excessive S. aureus colonization on the skin leads to the formation of biofilms and the secretion of virulence factors. These virulence factors facilitate mast cell degranulation, enhance inflammatory cytokine productions and the release of histamine, and increase IgE levels. In addition, S. aureus directly stimulates keratinocytes (KC) and Langerhans cells (LC) to release proinflammatory cytokines, including TH2 cytokines, TSLP, IL-8, and IL-1β, thus inducing TH2 differentiation and inflammation. Together, excessive cutaneous S. aureus colonization promotes AD pathogenesis. Conversely, commensal Coagulase-negative staphylococci (CoNS) inhibit the colonization of S. aureus by producing lantibiotics, PSMγ, and PSMδ. They also inhibit S. aureus biofilm formation by producing the serine protease glutamyl endopeptidase (Esp). Tryptophan metabolites derived from skin microbiota can activate AHR, thus inhibiting TSLP production by KCs and improving the epidermal barrier of the skin. AHR, aryl hydrocarbon receptor; AHRE, AHR element; AMPs, antimicrobial peptides; CoNS, Coagulase-negative staphylococci; IAId, indole-3-aldehyde; IL, interleukin; ILC3, lymphoid cells type 3; KC, keratinocyte; LC, Langerhans cell; PSM, phenol-soluble modulin, TSLP, thymic stromal lymphopoietin; SE, Staphylococcal enterotoxin; TSST-1, toxic shock syndrome toxin-1. Created with BioRender.com (accessed on 4 May 2022).
Figure 1.
Mechanisms of how skin microbiota regulates AD pathogenesis. Excessive S. aureus colonization on the skin leads to the formation of biofilms and the secretion of virulence factors. These virulence factors facilitate mast cell degranulation, enhance inflammatory cytokine productions and the release of histamine, and increase IgE levels. In addition, S. aureus directly stimulates keratinocytes (KC) and Langerhans cells (LC) to release proinflammatory cytokines, including TH2 cytokines, TSLP, IL-8, and IL-1β, thus inducing TH2 differentiation and inflammation. Together, excessive cutaneous S. aureus colonization promotes AD pathogenesis. Conversely, commensal Coagulase-negative staphylococci (CoNS) inhibit the colonization of S. aureus by producing lantibiotics, PSMγ, and PSMδ. They also inhibit S. aureus biofilm formation by producing the serine protease glutamyl endopeptidase (Esp). Tryptophan metabolites derived from skin microbiota can activate AHR, thus inhibiting TSLP production by KCs and improving the epidermal barrier of the skin. AHR, aryl hydrocarbon receptor; AHRE, AHR element; AMPs, antimicrobial peptides; CoNS, Coagulase-negative staphylococci; IAId, indole-3-aldehyde; IL, interleukin; ILC3, lymphoid cells type 3; KC, keratinocyte; LC, Langerhans cell; PSM, phenol-soluble modulin, TSLP, thymic stromal lymphopoietin; SE, Staphylococcal enterotoxin; TSST-1, toxic shock syndrome toxin-1. Created with BioRender.com (accessed on 4 May 2022).

Figure 2.
Mechanisms of how gut microbiota regulate AD pathogenesis. Short-chain fatty acids (SCFAs) produced by the gut microbiota are able to activate SCFA-sensing G-protein coupled receptors (GPCRs) and/or inhibit histone deacetylases (HDACs), thus activating downstream signalling cascades that suppress inflammatory responses and restoring TH1/TH2 balance. Microbial metabolite, D-tryptophan can also restore TH1/TH2 balance. Bifidobacteria, a genus of bacteria to which many probiotics belong, are an important source of these metabolites. Microbial tryptophan metabolites can activate the aryl hydrocarbon receptor (AHR), which inhibits inflammatory responses and improve the epidermal barrier of skin. Pathogen-associated molecular patterns produced by the gut microbiota can activate toll-like receptors (TLRs) thus restoring TH1/TH2 balance. All these mechanisms benefit AD. IAld, indole-3-aldehyde; IAA, indole-3-acetic acid; IPA, indole-3-propionic acid; TA, tryptamine; ARNT, aryl hydrocarbon receptor nuclear translocator. Created with BioRender.com (accessed on 4 May 2022).
Figure 2.
Mechanisms of how gut microbiota regulate AD pathogenesis. Short-chain fatty acids (SCFAs) produced by the gut microbiota are able to activate SCFA-sensing G-protein coupled receptors (GPCRs) and/or inhibit histone deacetylases (HDACs), thus activating downstream signalling cascades that suppress inflammatory responses and restoring TH1/TH2 balance. Microbial metabolite, D-tryptophan can also restore TH1/TH2 balance. Bifidobacteria, a genus of bacteria to which many probiotics belong, are an important source of these metabolites. Microbial tryptophan metabolites can activate the aryl hydrocarbon receptor (AHR), which inhibits inflammatory responses and improve the epidermal barrier of skin. Pathogen-associated molecular patterns produced by the gut microbiota can activate toll-like receptors (TLRs) thus restoring TH1/TH2 balance. All these mechanisms benefit AD. IAld, indole-3-aldehyde; IAA, indole-3-acetic acid; IPA, indole-3-propionic acid; TA, tryptamine; ARNT, aryl hydrocarbon receptor nuclear translocator. Created with BioRender.com (accessed on 4 May 2022).

Table 2.
Summary of clinical studies demonstrating the alternation of gut microbiota in patients with AD.
Table 2.
Summary of clinical studies demonstrating the alternation of gut microbiota in patients with AD.
| Year | Subjects, Numbers | Methods | Results (Alternations of Gut Microbiota) | Reference |
|---|---|---|---|---|
| 1999 | Two-year aged children; 13 Estonian and 14 Swedish allergic subjects; 16 Estonian and 19 Swedish nonallergic subjects | Culture | Allergic subjects: ↑Aerobic bacteria, ↑Coliforms, ↑S. aureus | [143] |
| 2001 | 22 atopic infants and 54 nonatopic infants | Culture, FISH | Atopic infants: ↑Clostridia | [163] |
| 2001 | Two-year aged children; 9 Estonian and 9 Swedish AD/allergic subjects; 13 Estonian and 11 Swedish healthy subjects | Culture | AD/allergic subjects: ↓Enterococci, ↓Bifidobacteria, ↓Bacteroides, ↑Clostridia, ↑S. aureus | [144] |
| 2003 | 30 AD patients and 68 healthy individuals | Culture | AD patients: ↓Bifidobacteria, ↑Staphylococci | [164] |
| 2006 | 21 AD toddlers and 28 healthy toddlers | Culture, FISH, quantitative flow cytometry, 16S rRNA gene sequencing | AD toddlers: ↓Bifidobacteria, ↑Clostridia, ↑Lactic producing bacteria, ↑Enterococci | [165] |
| 2006 | 26 AD infants and 52 healthy infants | PCR, 16S rRNA DGGE profile | Infants who developed AD within the first year of life: ↑E. coli | [147] |
| 2007 | 10 allergic infants and 16 healthy infants | 16S rRNA gene sequencing | Allergic infants: ↑Bifidobacterium catenulatum, ↑Bifidobacterium bifidum | [167] |
| 2007 | 324 infants | Culture | AD infants: ↑S. aureus, ↑Bacteroides | [145] |
| 2007 | 957 infants with high risk of allergic diseases | Real-time PCR | Subjects with higher risk of AD: ↑E. coli, ↑Clostridioides difficile | [148] |
| 2008 | 37 AD infants and 24 heathy infants | TTGE, FISH | AD infants: ↑Bifidobacterium pseudocatenulatum | [168] |
| 2008 | 9 AD infants and 12 healthy infants | DGGE | AD infants: ↓-diversity | [130] |
| 2008 | 15 AD infants and 20 healthy infants | T-RFLP, TTGE | AD infants: ↓-diversity | [131] |
| 2010 | 19 AD infants and 22 healthy infants | 16S rRNA gene sequencing | AD infants: ↓Bifidobacteria, ↓Enterobacteriaceae | [166] |
| 2011 | 3303 children | Real-time PCR | AD children: ↑Clostridioides difficile | [149] |
| 2011 | 411 infants with high risk of allergic diseases | PCR, 16S rRNA DGGE profile | ↓-diversity | [132] |
| 2012 | 20 AD infants and 20 healthy infants | 16S rRNA gene sequencing | AD infants: ↓-diversity, ↑Enterococcus spp., ↑Peptostreptococcaceae Incertae Sedis, ↑Eggerthella spp., ↑Coprobacillus spp., ↑Peptoniphilus spp., ↓Sutterela spp., ↓Fusobacterium spp. | [133] |
| 2012 | 33 AD infants and 65 healthy infants | T-RFLP | AD infants: ↓-diversity | [134] |
| 2013 | 1402 infants | Real-time PCR | AD infants: ↑Clostridia | [150] |
| 2015 | 28 AD infants and 11 healthy infants | HITChip | AD infants: ↓-diversity, ↓Butyrate-producing bacteria, ↓Coprococcus eutactus | [135] |
| 2016 | 298 neonates | 16S rRNA gene sequencing | Childhood AD history: ↓Bifidobacteria, ↓Akkermansias, ↓Faecalibacteria | [170] |
| 2016 | 19 AD infants and 14 healthy infants | 16S rRNA gene sequencing | AD infants: ↑Bacteroidaceae, ↑Deinococcaceae, ↓Clostridiaceae, ↓Veillonellaceae, ↓Lactobacillaceae | [140] |
| 2016 | 50 AD infants and 51 healthy infants | 16S rRNA gene sequencing | AD infants: ↑Escherichia/Shigella, ↑Veillonella, ↑Faecalibacterium, ↑Lachnospiraceae incertae sedis, ↑Clostridium XIVa, ↑Faecalibacterium prausnitzii, ↑Ruminococcus gnavus, ↑Akkermansia muciniphila, ↓Bifidobacterium, Megasphaera↓, ↓Haemophilus, ↓Streptococcus, ↓Bacteroides fragilis, ↓Streptococcus salivarius | [151] |
| 2016 | 90 AD patients and 42 healthy subjects | 16S rRNA gene sequencing and metagenomic sequencing | AD patients: A subspecies with of ↑Faecalibacterium prausnitzii, which is deficient in producing SCFAs, ↑Metabolic pathways responsive to oxidative stress↑, ↑Microbial genes encoding various transition metal transporters, ↑Microbial genes damaging mucin | [139] |
| 2016 | 12 AD infants and 12 healthy infants | 16S rRNA gene sequencing | AD infants: ↑Clostridia | [152] |
| 2018 | 63 AD infants and 66 healthy infants | Metagenomic sequencing and real-time PCR | AD infants: Microbial genes associated with PI3K-Akt signalling, estrogen signalling, NOD-like receptor signalling, ↑antigen processing and presentation | [141] |
| 2020 | 105 children | Metagenomic sequencing | AD children: ↓Microbial genes encoding carbohydrate active enzymes (CAZymes) from SCFA producers | [142] |
| 2020 | 81 AD children and 58 healthy children | Real-time PCR | AD children: ↑Clostridioides difficile, ↑Bifidobacteria, ↓Lactobacilli, ↓Eubacteria, ↓Lactobacillus spp., ↓B. fragilis, ↓E. coli, ↓Methanobrevibacter smithii | [146] |
| 2021 | 44 AD patients and 49 healthy subjects | 16S rRNA gene sequencing | AD patients: ↓α-diversity, ↑Porphyromonadaceae↑, Blautia, ↑Parabacteroides, ↑Bacteroides ovatus, ↑Bacteroides uniformis, ↑Prevotella stercorea, ↓Clostridium, ↓P. stercorea | [137] |
| 2021 | 1440 children | 16S rRNA gene sequencing | AD children: ↓α-diversity, ↓Lachnospiraceae, ↓Ruminococcaceae_UCG-005, ↓Christensenellaceae_R-7_group spp. | [138] |
Legend: FISH, fluorescence in situ hybridization; AD, atopic dermatitis; DGGE, denaturing gradient gel electrophoresis; TTGE, temporal temperature gel electrophoresis; T-RFLP, terminal restriction fragment length polymorphism; HITChip, the phylogenetic Human Intestinal Tract chip; PI3K, phosphatidylinositol 3-kinase; NOD, nucleotide-binding domain.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Alam, M.J.; Xie, L.; Yap, Y.-A.; Marques, F.Z.; Robert, R. Manipulating Microbiota to Treat Atopic Dermatitis: Functions and Therapies. Pathogens 2022, 11, 642. https://doi.org/10.3390/pathogens11060642
AMA Style
Alam MJ, Xie L, Yap Y-A, Marques FZ, Robert R. Manipulating Microbiota to Treat Atopic Dermatitis: Functions and Therapies. Pathogens. 2022; 11(6):642. https://doi.org/10.3390/pathogens11060642
Chicago/Turabian StyleAlam, Md Jahangir, Liang Xie, Yu-Anne Yap, Francine Z. Marques, and Remy Robert. 2022. "Manipulating Microbiota to Treat Atopic Dermatitis: Functions and Therapies" Pathogens 11, no. 6: 642. https://doi.org/10.3390/pathogens11060642
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.