
Japanese Encephalitis Virus Interaction with Mosquitoes: A Review of Vector Competence, Vector Capacity and Mosquito Immunity


Abstract
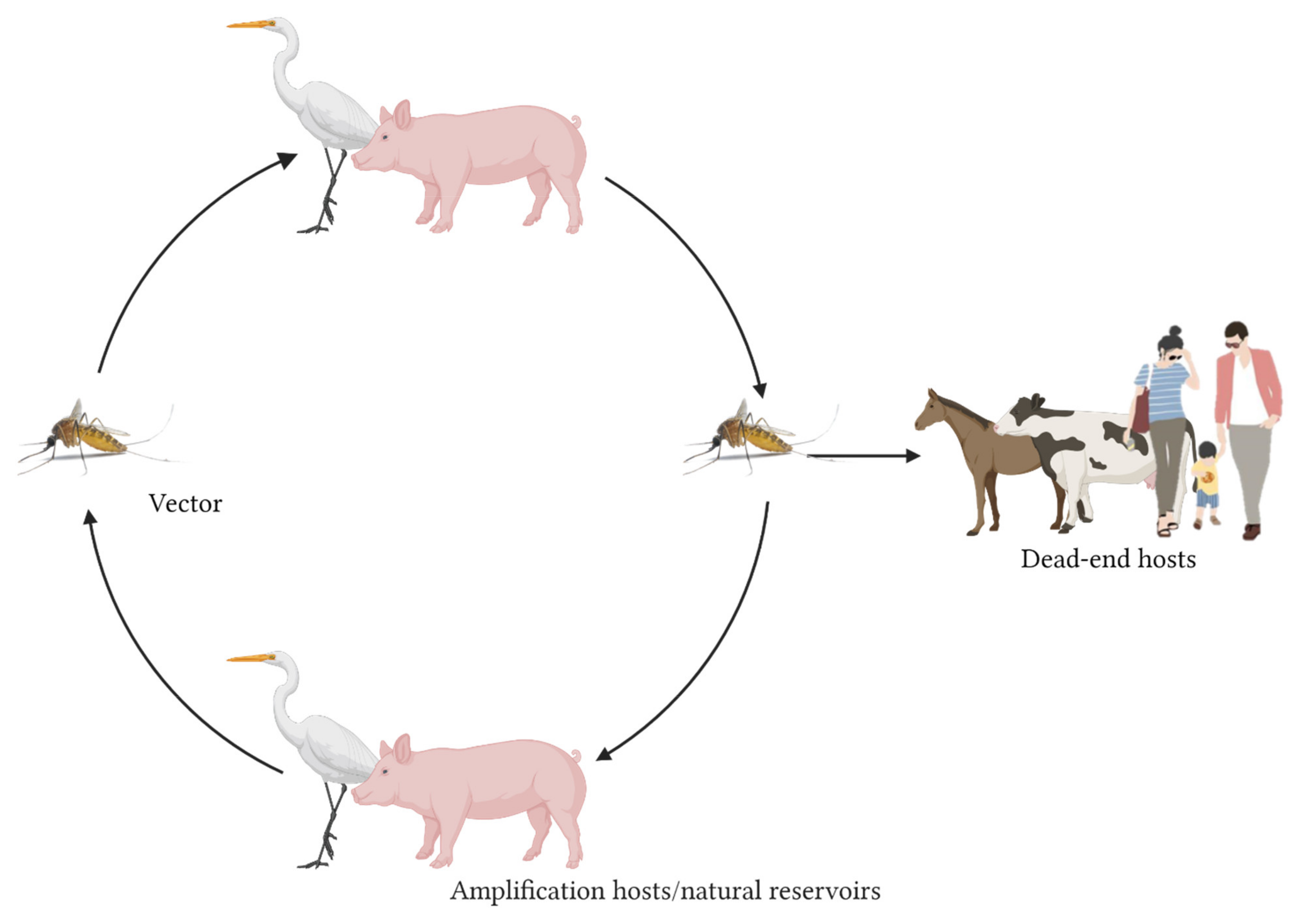
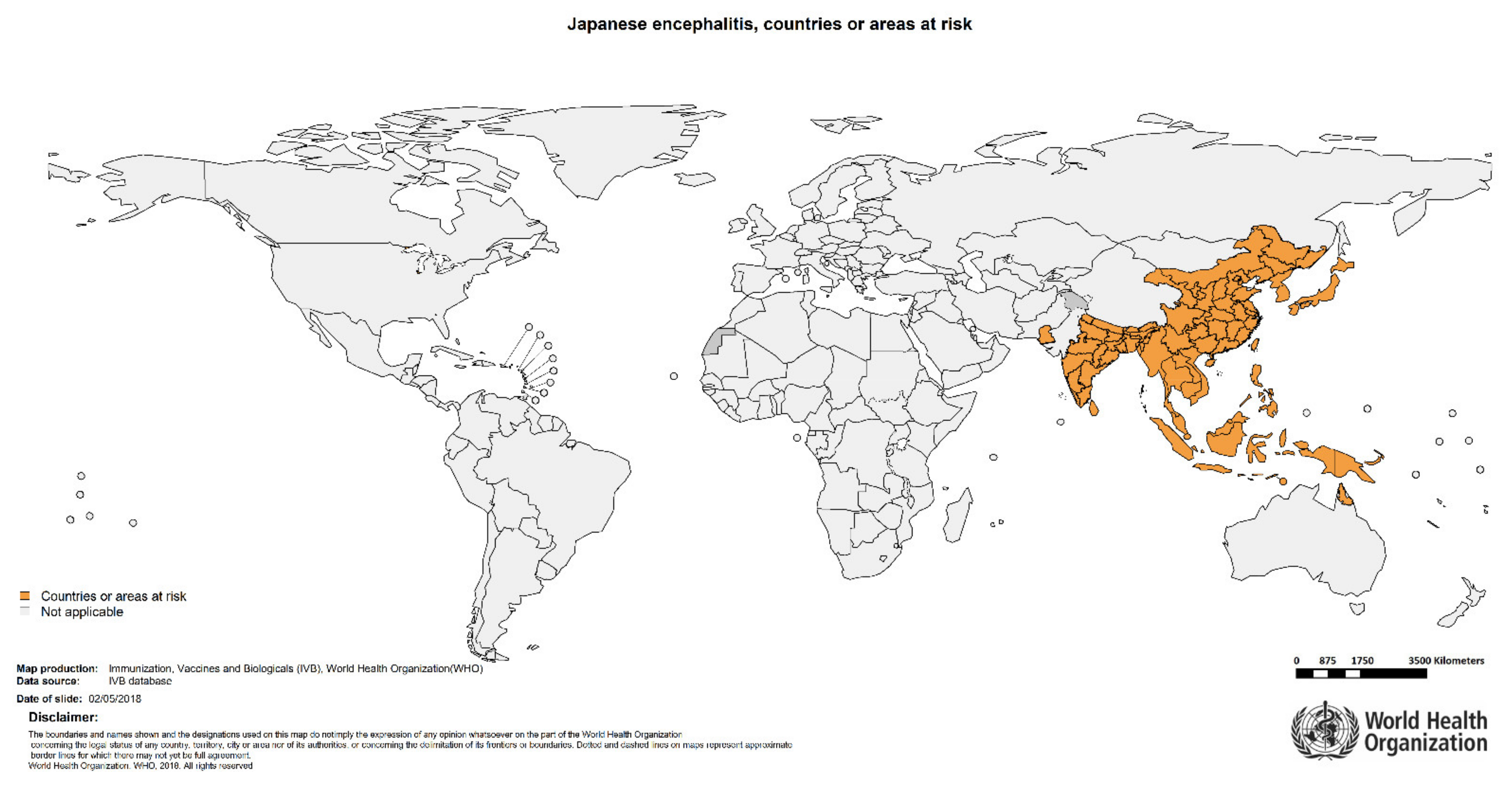
:1. Introduction
2. Results
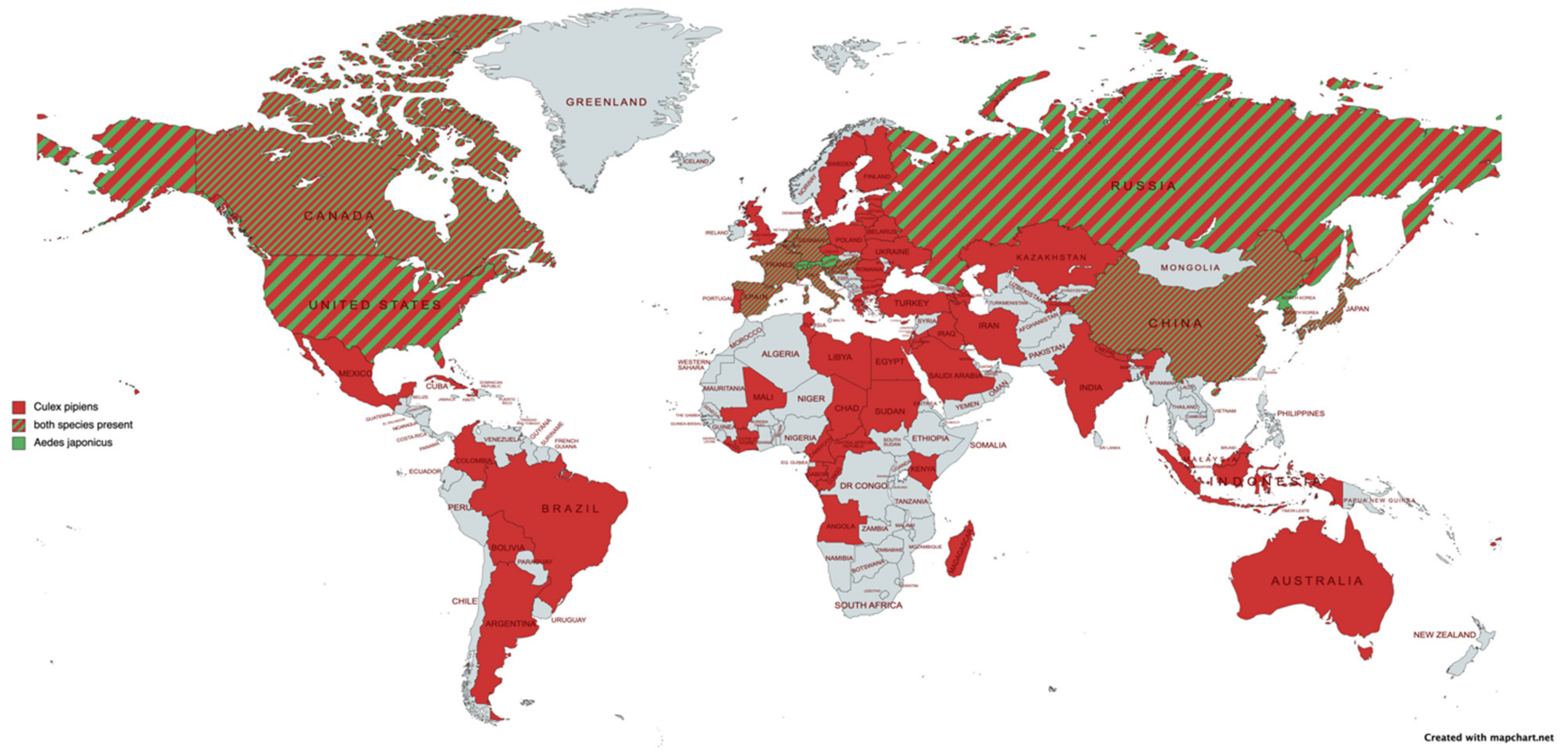
2.1. Mosquito Vectors of JEV: Vector Competence and Capacity
2.1.1. JEV Detection in Field-Collected Mosquitoes
2.1.2. JEV Vector Competence Studies
2.1.3. Vectorial Capacity
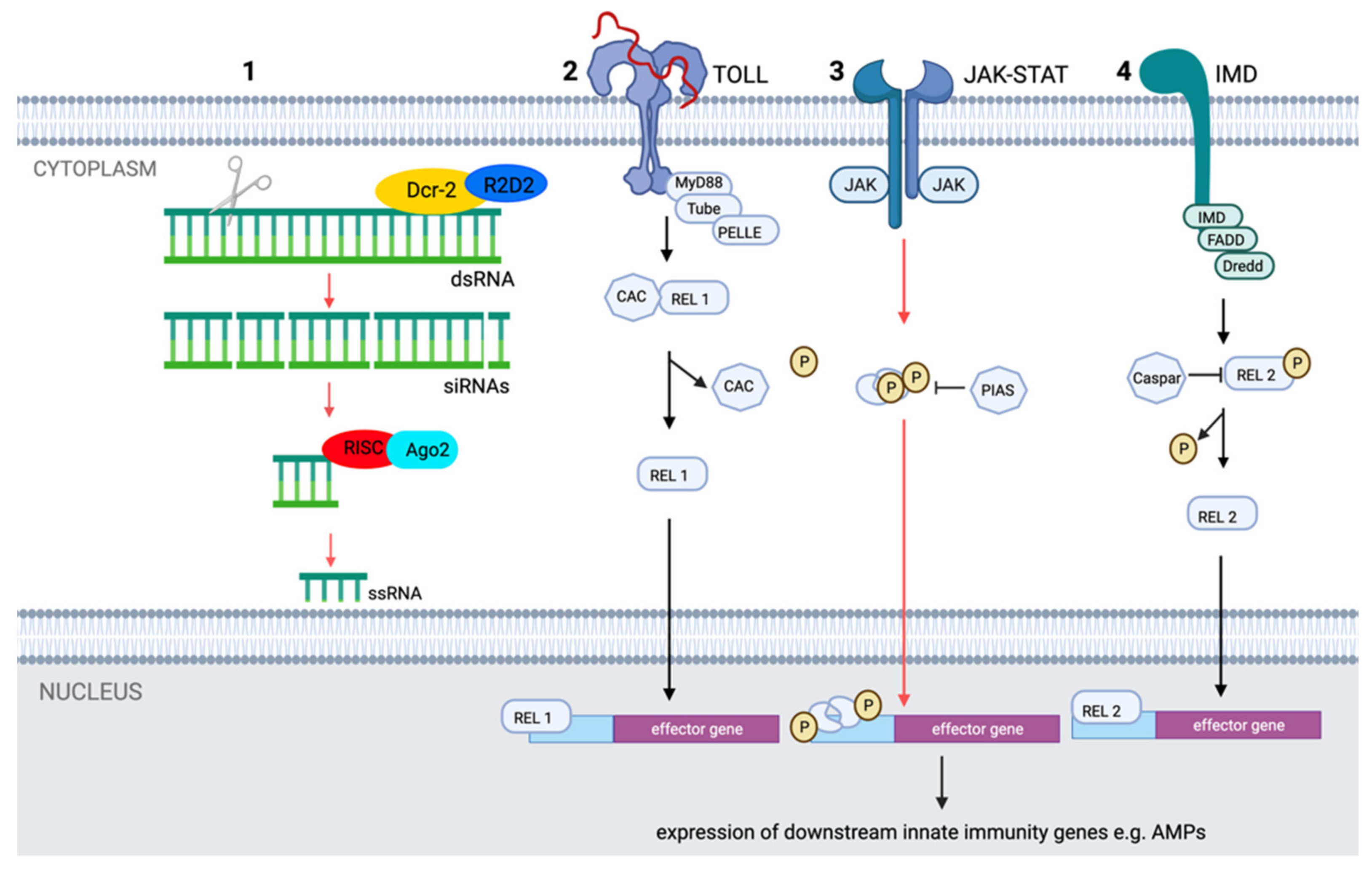
2.2. Mosquito Immunity Controlling JEV Replication and Dissemination
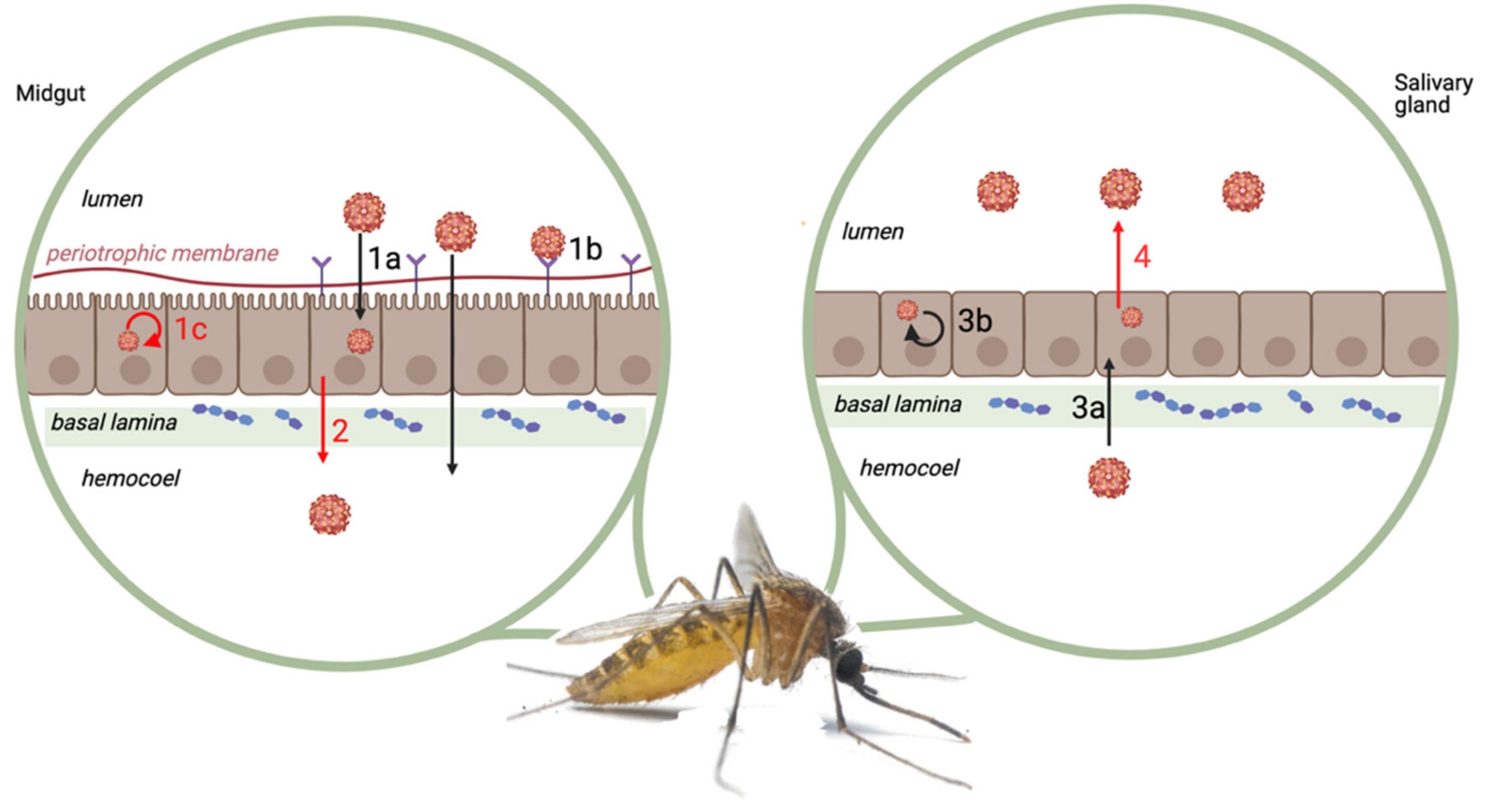
2.2.1. Physical and Physiological Barriers
2.2.2. Molecular Pathways
2.2.3. Antimicrobial Peptides
2.2.4. Vector Microbiome
3. Conclusions
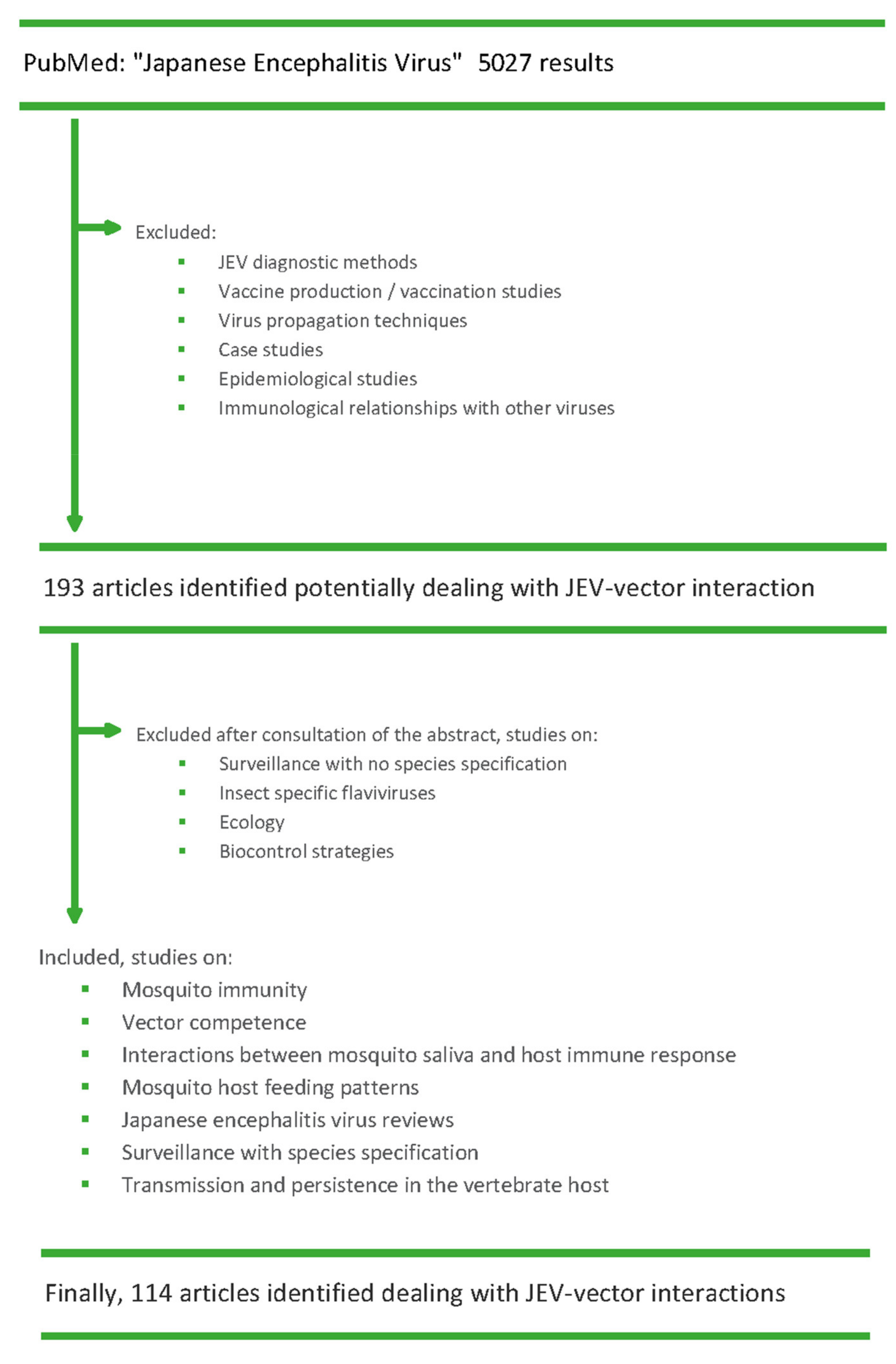
4. Methods
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Gould, E.; Pettersson, J.; Higgs, S.; Charrel, R.; de Lamballerie, X. Emerging arboviruses: Why today? One Health 2017, 4, 1–13. [Google Scholar] [CrossRef]
- Ricklin, M.E.; García-Nicolás, O.; Brechbühl, D.; Python, S.; Zumkehr, B.; Nougairede, A.; Charrel, R.N.; Posthaus, H.; Oevermann, A.; Summerfield, A. Vector-free transmission and persistence of Japanese encephalitis virus in pigs. Nat. Commun. 2016, 7, 10832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The World Organisation for Animal Health; World Health Organization. Japanese Encephalitis. OIE Technical Disease Cards. 2019. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/JAPANESE_ENCEPHALITIS.pdf (accessed on 24 March 2021).
- Hubálek, Z.; Rudolf, I.; Nowotny, N. Arboviruses pathogenic for domestic and wild animals. Adv. Virus Res. 2014, 89, 201–275. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.V.; Kruse, B.; The World Organisation for Animal Health; World Health Organization. Japanese Encephalitis; StatPearls Publishing: Treasure Island, FL, USA, 2019. [Google Scholar]
- Spickler, A.R. Japanese Encephalitis; Center for Food Security and Public Health Established at Iowa State: Ames, IA, USA, 2016. [Google Scholar]
- Solomon, T.; Ni, H.; Beasley, D.W.C.; Ekkelenkamp, M.; Cardosa, M.J.; Barrett, A.D.T. Origin and evolution of Japanese encephalitis virus in southeast Asia. J. Virol. 2003, 77, 3091–3098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Center of Disease Control. Facts about Japanese Encephalitis. Factsheet. 2017. Available online: https://www.ecdc.europa.eu/en/japanese-encephalitis/facts (accessed on 9 February 2021).
- Le Flohic, G.; Porphyre, V.; Barbazan, P.; Gonzalez, J.-P. Review of climate, landscape, and viral genetics as drivers of the Japanese encephalitis virus ecology. PLoS Negl. Trop. Dis. 2013, 7, e2208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuh, A.J.; Li, L.; Tesh, R.B.; Innis, B.L.; Barrett, A.D.T. Genetic characterization of early isolates of Japanese encephalitis virus: Genotype II has been circulating since at least 1951. J. Gen. Virol. 2010, 91, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Nga, P.T.; Parquet, M.D.C.; Cuong, V.D.; Ma, S.-P.; Hasebe, F.; Inoue, S.; Makino, Y.; Takagi, M.; Nam, V.S.; Morita, K. Shift in Japanese encephalitis virus (JEV) genotype circulating in northern Vietnam: Implications for frequent introductions of JEV from southeast Asia to East Asia. J. Gen. Virol. 2004, 85, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Faizah, A.N.; Kobayashi, D.; Amoa-Bosompem, M.; Higa, Y.; Tsuda, Y.; Itokawa, K.; Miura, K.; Hirayama, K.; Sawabe, K.; Isawa, H. Evaluating the competence of the primary vector, Culex tritaeniorhynchus, and the invasive mosquito species, Aedes japonicus japonicus, in transmitting three Japanese encephalitis virus genotypes. PLoS Negl. Trop. Dis. 2020, 14, e0008986. [Google Scholar] [CrossRef]
- Oliveira, A.R.S.; Piaggio, J.; Cohnstaedt, L.W.; McVey, D.S.; Cernicchiaro, N. Introduction of the Japanese encephalitis virus (JEV) in the United States—A qualitative risk assessment. Transbound. Emerg. Dis. 2019, 66, 1558–1574. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Ebel, G.D. How do virus–mosquito interactions lead to viral emergence? Trends Parasitol. 2018, 34, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Folly, A.J.; Dorey-Robinson, D.; Hernández-Triana, L.M.; Ackroyd, S.; Vidana, B.; Lean, F.Z.X.; Hicks, D.; Nuñez, A.; Johnson, N. Temperate conditions restrict Japanese encephalitis virus infection to the mid-gut and prevents systemic dissemination in Culex pipiens mosquitoes. Sci. Rep. 2021, 11, 6133. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Keyel, A.C. The role of temperature in transmission of zoonotic arboviruses. Viruses 2019, 11, 1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, W.; Lampman, R.; Novak, R.J. Problems in estimating mosquito infection rates using minimum infection rate. J. Med. Entomol. 2003, 40, 595–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, W.; Unnasch, T.R.; Katholi, C.R.; Lampman, R.; Novak, R.J. Fundamental issues in mosquito surveillance for arboviral transmission. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-L.; Yang, C.-F.; Teng, H.-J.; Lu, L.-C.; Lin, C.; Tsai, K.-H.; Chen, Y.-Y.; Chen, L.-Y.; Chang, S.-F.; Shu, P.-Y. Molecular epidemiology of Japanese encephalitis virus in mosquitoes in Taiwan during 2005–2012. PLoS Negl. Trop. Dis. 2014, 8, 3122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, M.H.; Lien, J.C.; Wang, Y.M.; Lin, C.C.; Lin, H.C.; Chin, C. Isolation of Japanese encephalitis virus from mosquitoes collected in northern Taiwan between 1995 and 1996. J. Microbiol. Immunol. Infect. 1999, 32, 9–13. [Google Scholar]
- Vythilingam, I.; Oda, K.; Chew, T.K.; Mahadevan, S.; Vijayamalar, B.; Morita, K.; Tsuchie, H.; Igarashi, A. Isolation of Japanese encephalitis virus from mosquitoes collected in Sabak Bernam, Selangor, Malaysia in 1992. J. Am. Mosq. Control Assoc. 1995, 11, 94–98. [Google Scholar]
- Vythilingam, I.; Oda, K.; Mahadevan, S.; Abdullah, G.; Thim, C.S.; Hong, C.C.; Vijayamalar, B.; Sinniah, M.; Igarashi, A. Abundance, parity, and Japanese encephalitis virus infection of mosquitoes (Diptera: Culicidae) in Sepang District, Malaysia. J. Med. Entomol. 1997, 34, 257–262. [Google Scholar] [CrossRef]
- Simpson, D.I.H.; Bowen, E.T.W.; Way, H.J.; Platt, G.S.; Hill, M.N.; Kamath, S.; Wah, L.T.; Bendell, P.J.E.; Heathcote, O.H.U. Arbovirus infections in Sarawak, October 1968–February 1970, Japanese encephalitis virus isolations from mosquitoes. Ann. Trop. Med. Parasitol. 1974, 68, 393–404. [Google Scholar] [CrossRef]
- Weng, M.-H.; Lien, J.-C.; Ji, D.-D. Monitoring of Japanese encephalitis virus infection in mosquitoes (Diptera: Culicidae) at Guandu Nature Park, Taipei, 2002–2004. J. Med. Entomol. 2005, 42, 1085–1088. [Google Scholar] [CrossRef]
- Hanna, J.N.; Ritchie, S.A.; Hills, S.L.; Hurk, A.F.V.D.; Phillips, D.A.; Pyke, A.T.; Lee, J.M.; Johansen, C.A.; MacKenzie, J.S. Japanese encephalitis in north Queensland, Australia, 1998. Med. J. Aust. 1999, 170, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Johansen, C.A.; Hurk, A.F.V.D.; Pyke, A.T.; Zborowski, P.; Phillips, D.A.; MacKenzie, J.S.; Ritchie, S.A. Entomological investigations of an outbreak of Japanese encephalitis virus in the Torres Strait, Australia, in 1998. J. Med. Entomol. 2001, 38, 581–588. [Google Scholar] [CrossRef] [Green Version]
- Olson, J.G.; Ksiazek, T.G.; Lee, V.H.; Tan, R.; Shope, R.E. Isolation of Japanese encephalitis virus from Anopheles annularis and Anopheles vagus in Lombok, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1985, 79, 845–847. [Google Scholar] [CrossRef] [Green Version]
- Thenmozhi, V.; Balaji, T.; Selvam, A.; Venkatasubramani, K.; Dhananjeyan, K.J. A longitudinal study on abundance and infection frequency of Japanese encephalitis vectors in Tirunelveli District, Tamil Nadu, India. Int. J. Mosq. Res. 2015, 2, 166–169. [Google Scholar]
- Chakravarty, S.K.; Sarkar, J.K.; Chakravarty, M.S.; Mukherjee, M.K.; Mukherjee, K.K.; Das, B.C.; Hati, A.K. The first epidemic of Japanese encephalitis studied in India–Virological studies. Indian J. Med. Res. 1975, 63, 77–82. [Google Scholar]
- Banerjee, K.; Mahadev, P.V.; Ilkal, M.A.; Mishra, A.C.; Dhanda, V.; Modi, G.B.; Geevarghese, G.; Kaul, H.N.; Shetty, P.S.; George, P.J. Isolation of Japanese encephalitis virus from mosquitoes collected in Bankura District (West Bengal) during October 1974 to December 1975. Indian J. Med. Res. 1979, 69, 201–205. [Google Scholar] [PubMed]
- Mourya, D.T.; Ilkal, M.A.; Mishra, A.C.; Jacob, P.G.; Pant, U.; Ramanujam, S.; Mavale, M.S.; Bhat, H.R.; Dhanda, V. Isolation of Japanese encephalitis virus from mosquitoes collected in Karnataka state, India from 1985 to 1987. Trans. R. Soc. Trop. Med. Hyg. 1989, 83, 550–552. [Google Scholar] [CrossRef]
- Liu, H.; Lu, H.-J.; Liu, Z.-J.; Jing, J.; Ren, J.-Q.; Liu, Y.-Y.; Lu, F.; Jin, N.-Y. Japanese encephalitis virus in mosquitoes and swine in Yunnan province, China 2009–2010. Vector-Borne Zoonotic Dis. 2013, 13, 41–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Guo, X.; Zhao, Q.; Tong, Y.; Fan, H.; Sun, Q.; Xing, S.; Zhou, H.; Zhang, J. Investigation on mosquito-borne biruses at Lancang river and Nu river watersheds in southwestern China. Vector-Borne Zoonotic Dis. 2017, 17, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Fu, S.; Zhang, H.; Li, M.; Zhou, T.; Wang, J.; Zhang, Y.; Wang, H.; Tang, Q.; Liang, G. Distribution of mosquitoes and mosquito-borne viruses along the China-Myanmar border in Yunnan province. Jpn. J. Infect. Dis. 2012, 65, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhanda, V.; Thenmozhi, V.; Kumar, N.P.; Hiriyan, J.; Arunachalam, N.; Balasubramanian, A.; Ilango, A.; Gajanana, A. Virus isolation from wild-caught mosquitoes during a Japanese encephalitis outbreak in Kerala in 1996. Indian J. Med. Res. 1997, 106, 4–6. [Google Scholar]
- Thenmozhi, V.; Rajendran, R.; Ayanar, K.; Manavalan, R.; Tyagi, B.K. Long-term study of Japanese encephalitis virus infection in Anopheles subpictus in Cuddalore District, Tamil Nadu, south India. Trop. Med. Int. Health 2006, 11, 288–293. [Google Scholar] [CrossRef]
- George, S.; Jacob, P.G.; Rao, J.A. Isolation of Japanese encephalitis & West Nile viruses from mosquitoes collected in Kolar District of Karnataka state during 1977–1979. Indian J. Med. Res. 1987, 85, 235–238. [Google Scholar] [PubMed]
- Shi, Q.; Song, X.; Lv, Y.; Huang, X.; Kou, J.; Wang, H.W.; Zhang, H.; Cheng, P.; Gong, M. Potential risks associated with Japanese encephalitis prevalence in Shandong province, China. Vector-Borne Zoonotic Dis. 2019, 19, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Phillips, D.; Poidinger, M.; Mackenzie, J.; Van Den Hurk, A.; Ritchie, S.A.; Broom, A. Isolation of Japanese encephalitis virus from Culex annulirostris in Australia. Am. J. Trop. Med. Hyg. 1997, 56, 80–84. [Google Scholar] [CrossRef]
- Cates, M.D.; Detels, R. Japanese encephalitis virus in Taiwan: Preliminary evidence for Culex annulus Theob. as a vector. J. Med. Entomol. 1969, 6, 327–328. [Google Scholar] [CrossRef] [PubMed]
- Cross, J.H.; Lien, J.C.; Huang, W.C.; Lien, S.C.; Chiu, S.F. Japanese encephalitis virus surveillance in Taiwan. II. Isolations from mosquitoes and bats in Taipei area 1969–1970. Taiwan Yi Xue Hui Za Zhi 1971, 70, 681–686. [Google Scholar]
- Okuno, T.; Tseng, P.T.; Liu, S.Y.; Hsu, S.T.; Huang, C.T. Rates of infection with Japanese encephalitis virus of two culicine species of mosquito in Taiwan. Bull. World Health Organ. 1971, 44, 599–604. [Google Scholar] [PubMed]
- Seo, H.-J.; Kim, H.C.; Klein, T.A.; Ramey, A.M.; Lee, J.-H.; Kyung, S.-G.; Park, J.-Y.; Cho, Y.S.; Cho, I.-S.; Yeh, J.-Y. Molecular detection and genotyping of Japanese encephalitis virus in mosquitoes during a 2010 outbreak in the Republic of Korea. PLoS ONE 2013, 8, e55165. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.C.; Takhampunya, R.; Tippayachai, B.; Chong, S.-T.; Park, J.-Y.; Kim, M.-S.; Seo, H.-J.; Yeh, J.-Y.; Lee, W.-J.; Lee, D.-K.; et al. Japanese encephalitis virus in culicine mosquitoes (Diptera: Culicidae) of the Republic of Korea, 2008–2010. Mil. Med. 2015, 180, 158–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.C.; Klein, T.A.; Takhampunya, R.; Evans, B.P.; Mingmongkolchai, S.; Kengluecha, A.; Grieco, J.; Masuoka, P.; Kim, M.-S.; Chong, S.-T.; et al. Japanese encephalitis virus in culicine mosquitoes (Diptera: Culicidae) collected at Daeseongdong, a village in the demilitarized zone of the Republic of Korea. J. Med. Entomol. 2011, 48, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Gajanana, A.; Rajendran, R.; Samuel, P.P.; Thenmozhi, V.; Tsai, T.F.; Kimura-Kuroda, J.; Reuben, R. Japanese encephalitis in south Arcot District, Tamil Nadu, India: A three-year longitudinal study of vector abundance and infection frequency. J. Med. Entomol. 1997, 34, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.G.; Ksiazek, T.G.; Tan, R.; Atmosoedjono, S.; Lee, V.H.; Converse, J.D. Correlation of population indices of female Culex tritaeniorhynchus with Japanese encephalitis viral activity in Kapuk, Indonesia. Southeast Asian J. Trop. Med. Public Health 1985, 16, 337–342. [Google Scholar]
- Chen, Y.-C.; Wang, C.-Y.; Teng, H.-J.; Chen, C.-F.; Chang, M.-C.; Lu, L.-C.; Lin, C.; Jian, S.-W.; Wu, H.-S. Comparison of the efficacy of CO2-baited and unbaited light traps, gravid traps, backpack aspirators, and sweep net collections for sampling mosquitoes infected with Japanese encephalitis virus. J. Vector Ecol. 2011, 36, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.H.; Huang, W.C.; Cross, J.H. The isolation of Japanese encephalitis virus from Taiwan mosquitoes by mosquito cell cultures and mouse inoculation. J. Med. Entomol. 1978, 14, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Gould, D.J.; Edelman, R.; Grossman, R.A.; Nisalak, A.; Sullivan, M.F. Study of Japanese encephalitis virus in Chiangmai Valley, Thailand. IV. Vector studies. Am. J. Epidemiol. 1974, 100, 49–56. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Hurk, A.F.V.D.; Zborowski, P.; Kerlin, T.J.; Banks, D.; Walker, J.A.; Lee, J.M.; Montgomery, B.L.; Smith, G.A.; Pyke, A.T.; et al. Operational trials of remote mosquito trap systems for Japanese encephalitis virus surveillance in the Torres Strait, Australia. Vector-Borne Zoonotic Dis. 2007, 7, 497–506. [Google Scholar] [CrossRef] [PubMed]
- Tewari, S.C.; Thenmozhi, V.; Rajendran, R.; Appavoo, N.C.; Gajanana, A. Detection of Japanese encephalitis virus antigen in desiccated mosquitoes: An improved surveillance system. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 525–526. [Google Scholar] [CrossRef]
- Tewari, S.C.; Thenmozhi, V.; Arunachalam, N.; Philip Samuel, P.; Tyagi, B.K. Desiccated vector mosquitoes used for the surveillance of Japanese encephalitis virus activity in endemic southern India. Trop. Med. Int. Health 2008, 13, 286–290. [Google Scholar] [CrossRef]
- Rajendran, R.; Thenmozhi, V.; Tewari, S.C.; Balasubramanian, A.; Ayanar, K.; Manavalan, R.; Gajanana, A.; Kabilan, L.; Thakare, J.P.; Satyanarayana, K. Longitudinal studies in south Indian villages on Japanese encephalitis virus infection in mosquitoes and seroconversion in goats. Trop. Med. Int. Health 2003, 8, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Samuel, P.P.; Ramesh, D.; Muniaraj, M.; Thenmozhi, V.; Venkatesh, A.; Nagaraj, J.; Tyagi, B. Seasonal abundance role of predominant Japanese encephalitis vectors Culex tritaeniorhynchus and Cx. gelidus Theobald in Cuddalore District, Tamil Nadu. Indian J. Med. Res. 2015, 142, 23. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, N.; Samuel, P.P.; Hiriyan, J.; Thenmozhi, V.; Gajanana, A. Japanese encephalitis in Kerala, south India: Can Mansonia (Diptera: Culicidae) play a supplemental role in transmission? J. Med. Entomol. 2004, 41, 456–461. [Google Scholar] [CrossRef] [PubMed]
- Van Peenen, P.F.D.; Joseph, P.L.; Atmosoedjono, S.; Irsiana, R.; Saroso, J.S. Japanese encephalitis virus from pigs and mosquitoes in Jakarta, Indonesia. Trans. R. Soc. Trop. Med. Hyg. 1975, 69, 477–479. [Google Scholar] [CrossRef]
- Simpson, D.; Bowen, E.; Platt, G.; Way, H.; Smith, C.; Peto, S.; Kamath, S.; Lim, B.L.; Lim, T.W. Japanese encephalitis in Sarawak: Virus isolation and serology in a Land Dyak village. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 503–510. [Google Scholar] [CrossRef]
- Peiris, J.S.M.; Amerasinghe, F.P.; Amerasinghe, P.H.; Ratnayake, C.B.; Karunaratne, S.H.P.P.; Tsai, T.F. Japanese encephalitis in Sri Lanka—The study of an epidemic: Vector incrimination, porcine infection and human disease. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 307–313. [Google Scholar] [CrossRef]
- Simasathien, P.; Rohitayodhin, S.; Nisalak, A.; Singharaj, P.; Halstead, S.B.; Russell, P.K. Recovery of Japanese encephalitis virus from wild caught mosquitoes in Thailand. Southeast Asian J. Trop. Med. Public Health 1972, 3, 52–54. [Google Scholar]
- Thi-Kim-Thoa, N.; Ngo-Thi-Vien Tran-Tuyet-Mai Thi-Ngoc-Xuan, N. Japanese encephalitis vectors: Isolation of virus from culicine mosquitoes in the Saigon area. Southeast Asian J. Trop. Med. Public Health 1974, 5, 408–412. [Google Scholar] [PubMed]
- Kim, H.; Cha, G.-W.; Jeong, Y.E.; Lee, W.-G.; Chang, K.S.; Roh, J.Y.; Yang, S.C.; Park, M.Y.; Park, C.; Shin, E.-H. Detection of Japanese encephalitis virus genotype V in Culex orientalis and Culex pipiens (Diptera: Culicidae) in Korea. PLoS ONE 2015, 10, e0116547. [Google Scholar] [CrossRef] [Green Version]
- Ravanini, P.; Huhtamo, E.; Ilaria, V.; Crobu, M.G.; Nicosia, A.M.; Servino, L.; Rivasi, F.; Allegrini, S.; Miglio, U.; Magri, A.; et al. Japanese encephalitis virus RNA detected in Culex pipiens mosquitoes in Italy. Eurosurveillance 2012, 17, 20221. [Google Scholar] [CrossRef]
- Fang, Y.; Zhang, Y.; Zhou, Z.-B.; Xia, S.; Shi, W.-Q.; Xue, J.-B.; Li, Y.-Y.; Wu, J.-T. New strains of Japanese encephalitis virus circulating in Shanghai, China after a ten-year hiatus in local mosquito surveillance. Parasites Vectors 2019, 12, 22. [Google Scholar] [CrossRef]
- Dhanda, V.; Mourya, D.T.; Mishra, A.C.; Ilkal, M.A.; Pant, U.; Jacob, P.G.; Bhat, H.R. Japanese encephalitis virus infection in mosquitoes reared from field-collected immatures and in wild-caught males. Am. J. Trop. Med. Hyg. 1989, 41, 732–736. [Google Scholar] [CrossRef] [PubMed]
- Borah, J.; Dutta, P.; Khan, S.A.; Mahanta, J. Epidemiological concordance of Japanese encephalitis virus infection among mosquito vectors, amplifying hosts and humans in India. Epidemiol. Infect. 2013, 141, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Nyari, N.; Singh, D.; Kakkar, K.; Sharma, S.; Pandey, S.N.; Dhole, T.N. Entomological and serological investigation of Japanese encephalitis in endemic area of eastern Uttar Pradesh, India. J. Vector Borne Dis. 2015, 52, 321–328. [Google Scholar] [PubMed]
- Naik, P.S.; Ilkal, M.A.; Pant, U.; Kulkarni, S.M.; Dhanda, V. Isolation of Japanese encephalitis virus from Culex pseudovishnui Colless, 1957 (Diptera: Culicidae) in Goa. Indian J. Med. Res. 1990, 91, 331–333. [Google Scholar]
- Nitatpattana, N.; Apiwathnasorn, C.; Barbazan, P.; Leemingsawat, S.; Yoksan, S.; Gonzalez, J.-P. First isolation of Japanese encephalitis from Culex quinquefasciatus in Thailand. Southeast Asian J. Trop. Med. Public Health 2005, 36, 875–878. [Google Scholar]
- Lindahl, J.F.; Ståhl, K.; Chirico, J.; Boqvist, S.; Thu, H.T.V.; Magnusson, U. Circulation of Japanese encephalitis virus in pigs and mosquito vectors within Can Tho City. PLoS Negl. Trop. Dis. 2013, 7, e2153. [Google Scholar] [CrossRef]
- Hurk, A.F.V.D.; MacKenzie, J.S.; Smith, I.L.; Ritchie, S.A.; Smith, G.A.; Northill, J.A.; Montgomery, B.L.; Zborowski, P. Short report: The first isolation of Japanese encephalitis virus from mosquitoes collected from mainland Australia. Am. J. Trop. Med. Hyg. 2006, 75, 21–25. [Google Scholar] [CrossRef] [Green Version]
- Nisbet, D.J.; Foley, P.N.; Van Den Hurk, A.F.; Mackenzie, J.S. Flavivirus isolations from mosquitoes collected from Saibai Island in the Torres Strait, Australia, during an incursion of Japanese encephalitis virus. Med. Veter. Entomol. 2004, 18, 281–287. [Google Scholar]
- Johansen, C.A.; van den Hurk, A.F.; Ritchie, S.A.; Zborowski, P.; Nisbet, D.J.; Paru, R.; Bockarie, M.J.; MacDonald, J.; Drew, A.C.; Khromykh, T.I.; et al. Isolation of Japanese encephalitis virus from mosquitoes (Diptera: Culicidae) collected in the western province of Papua New Guinea, 1997–1998. Am. J. Trop. Med. Hyg. 2000, 62, 631–638. [Google Scholar] [CrossRef]
- Duong, V.; Choeung, R.; Gorman, C.; Laurent, D.; Crabol, Y.; Mey, C.; Peng, B.; Di Francesco, J.; Hul, V.; Sothy, H.; et al. Isolation and full-genome sequences of Japanese encephalitis virus genotype I strains from Cambodian human patients, mosquitoes and pigs. J. Gen. Virol. 2017, 98, 2287–2296. [Google Scholar] [CrossRef]
- Fang, Y.; Li, X.-S.; Zhang, W.; Xue, J.-B.; Wang, J.-Z.; Yin, S.-Q.; Li, S.-G.; Li, X.-H.; Zhang, Y. Molecular epidemiology of mosquito-borne viruses at the China–Myanmar border: Discovery of a potential epidemic focus of Japanese encephalitis. Infect. Dis. Poverty 2021, 10, 57. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Zhang, W.; Xue, J.-B.; Zhang, Y. Monitoring mosquito-borne arbovirus in various insect regions in China in 2018. Front. Cell Infect. Microbiol. 2021, 11, 640993. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, R.; Sugiyama, H.; Yonemitsu, K.; Van Dung, N.; Terada, Y.; Taniguchi, M.; Shimoda, H.; Takano, A.; Maeda, K. Isolation of Japanese encephalitis virus and a novel insect-specific flavivirus from mosquitoes collected in a cowshed in Japan. Arch. Virol. 2015, 160, 2151–2159. [Google Scholar] [CrossRef]
- Yap, G.; Mailepessov, D.; Lim, X.F.; Chan, S.; How, C.B.; Humaidi, M.; Yeo, G.; Chong, C.S.; Lam-Phua, S.G.; Lee, R.; et al. Detection of Japanese encephalitis virus in Culex mosquitoes in Singapore. Am. J. Trop. Med. Hyg. 2020, 103, 1234–1240. [Google Scholar] [CrossRef]
- Leake, C.; Ussery, M.; Nisalak, A.; Hoke, C.H.; Andre, R.G.; Burke, D.S. Virus isolations from mosquitoes collected during the 1982 Japanese encephalitis epidemic in northern Thailand. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 831–837. [Google Scholar] [CrossRef]
- Kuwata, R.; Trang, B.M.; Tsuda, Y.; Sawabe, K.; Nga, P.T.; Sasaki, T.; Yen, N.T.; Kobayashi, M.; Takagi, M.; Loan, D.P.; et al. Surveillance of Japanese encephalitis virus infection in mosquitoes in Vietnam from 2006 to 2008. Am. J. Trop. Med. Hyg. 2013, 88, 681–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearce, J.C.; Learoyd, T.P.; Langendorf, B.J.; Logan, J.G. Japanese encephalitis: The vectors, ecology and potential for expansion. J. Travel Med. 2018, 25, S16–S26. [Google Scholar] [CrossRef] [Green Version]
- Souza-Neto, J.A.; Powell, J.R.; Bonizzoni, M. Aedes aegypti vector competence studies: A review. Infect. Genet. Evol. 2019, 67, 191–209. [Google Scholar] [CrossRef]
- Schulz, C.; Becker, S.C. Mosquitoes as arbovirus vectors: From species identification to vector competence. Mosquito-Borne Dis. 2018, 10, 163–212. [Google Scholar] [CrossRef]
- Kramer, L.D.; Chin, P.; Cane, R.P.; Kauffman, E.B.; Mackereth, G. Vector competence of New Zealand mosquitoes for selected arboviruses. Am. J. Trop. Med. Hyg. 2011, 85, 182–189. [Google Scholar] [CrossRef]
- Mackenzie-Impoinvil, L.; Impoinvil, D.E.; Galbraith, S.E.; Dillon, R.J.; Ranson, H.; Johnson, N.; Fooks, A.R.; Solomon, T.; Baylis, M. Evaluation of a temperate climate mosquito, Ochlerotatus detritus (=Aedes detritus), as a potential vector of Japanese encephalitis virus. Med. Vet. Entomol. 2015, 29, 1–9. [Google Scholar] [CrossRef]
- Conn, J.E.; Lainhart, W.; Rios, C.T.; Vinetz, J.M.; Bickersmith, S.A.; Moreno, M. Changes in genetic diversity from field to laboratory during colonization of Anopheles darlingi Root (Diptera: Culicidae). Am. J. Trop. Med. Hyg. 2015, 93, 998–1001. [Google Scholar] [CrossRef] [Green Version]
- De Wispelaere, M.; Desprès, P.; Choumet, V. European Aedes albopictus and Culex pipiens are competent vectors for Japanese encephalitis virus. PLoS Negl. Trop. Dis. 2017, 11, e0005294. [Google Scholar] [CrossRef]
- Huang, Y.-J.S.; Hettenbach, S.M.; Park, S.L.; Higgs, S.; Barrett, A.D.T.; Hsu, W.-W.; Harbin, J.N.; Cohnstaedt, L.W.; VanLandingham, D.L. Differential infectivities among different Japanese encephalitis virus genotypes in Culex quinquefasciatus mosquitoes. PLoS Negl. Trop. Dis. 2016, 10, e0005038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, G.E.; Sherlock, K.; Hesson, J.C.; Blagrove, M.S.C.; Lycett, G.J.; Archer, D.; Solomon, T.; Baylis, M. Laboratory transmission potential of British mosquitoes for equine arboviruses. Parasites Vectors 2020, 13, 413. [Google Scholar] [CrossRef] [PubMed]
- Cleton, N.B.; Bosco-Lauth, A.; Page, M.J.; Bowen, R.A. Age-related susceptibility to Japanese encephalitis virus in domestic ducklings and chicks. Am. J. Trop. Med. Hyg. 2014, 90, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Vogels, C.B.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azar, S.R.; Weaver, S.C. Vector competence: What has Zika virus taught us? Viruses 2019, 11, 867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Den Hurk, A.F.; Nisbet, D.J.; Hall, R.A.; Kay, B.H.; Mackenzie, J.S.; Ritchie, S.A. Vector competence of Australian mosquitoes (Diptera: Culicidae) for Japanese encephalitis virus. J. Med. Entomol. 2003, 40, 82–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholson, J.; Ritchie, S.A.; Van Den Hurk, A.F. Aedes albopictus (Diptera: Culicidae) as a potential vector of endemic and exotic arboviruses in Australia. J. Med. Entomol. 2014, 51, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Weng, M.H.; Lien, J.C.; Wang, Y.U.M.; Wu, H.L.; Chin, C. Susceptibility of three laboratory strains of Aedes albopictus (Diptera: Culicidae) to Japanese encephalitis virus from Taiwan. J. Med. Entomol. 1997, 34, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.C.; Hammon, W.M. Laboratory transmission of Japanese B encephalitis virus by seven species (three genera) of North American mosquitoes. J. Exp. Med. 1946, 83, 185–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, K.; Jansen, S.; Leggewie, M.; Badusche, M.; Schmidt-Chanasit, J.; Becker, N.; Tannich, E.; Becker, S.C. Aedes japonicus japonicus (Diptera: Culicidae) from Germany have vector competence for Japanese encephalitis virus but are refractory to infection with West Nile virus. Parasitol. Res. 2014, 113, 3195–3199. [Google Scholar] [CrossRef] [PubMed]
- Takashima, I.; Rosen, L. Horizontal and vertical transmission of Japanese encephalitis virus by Aedes japonicus (Diptera: Culicidae). J. Med. Entomol. 1989, 26, 454–458. [Google Scholar] [CrossRef] [PubMed]
- Hodes, H.L. Experimental transmission of Japanese B. encephalitis by mosquitoes and mosquito larvae. Bull. Johns Hopkins Hosp. 1946, 79, 358. [Google Scholar] [PubMed]
- Banerjee, K.; Deshmukh, P.K.; Ilkal, M.A.; Dhanda, V. Experimental transmission of Japanese encephalitis virus through Anopheles tessellatus and Culex fatigans mosquitoes. Indian J. Med. Res. 1977, 65, 746–752. [Google Scholar] [PubMed]
- Chen, W.-J.; Dong, C.-F.; Chiou, L.-Y.; Chuang, W.-L. Potential role of Armigeres subalbatus (Diptera: Culicidae) in the transmission of Japanese encephalitis virus in the absence of rice culture on Liu-Chiu islet, Taiwan. J. Med. Entomol. 2000, 37, 108–113. [Google Scholar] [CrossRef]
- Dhanda, V.; Banerjee, K.; Deshmukh, P.K.; Ilkal, M.A. Experimental viraemia and transmission of Japanese encephalitis virus by mosquitoes in domestic ducks. Indian J. Med. Res. 1977, 66, 881–888. [Google Scholar] [PubMed]
- Banerjee, K.; Deshmukh, P.K.; Ilkal, M.A.; Dhanda, V. Transmission of Japanese encephalitis virus by Culex bitaeniorhynchus Giles. Indian J. Med. Res. 1978, 67, 889–893. [Google Scholar] [PubMed]
- Banerjee, K.; Deshmukh, P.K.; Ilkal, M.A.; Dhanda, V. Comparative susceptibility of three species of mosquitoes to infection with Japanese encephalitis virus. Indian J. Med. Res. 1983, 78, 603–606. [Google Scholar] [PubMed]
- Okuno, T.; Mitchell, C.J.; Chen, P.S.; Hsu, S.; Ryu, E. Experimental transmission of Japanese encephalitis virus by Culex tritaeniorhynchus and C. fuscocephalus. Ann. Trop. Med. Parasitol. 1975, 69, 203–206. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Edelman, R.; Muangman, D.; Gould, D.J. Experimental transmission of Japanese encephalitis virus by Culex fuscocephala. Am. J. Trop. Med. Hyg. 1972, 21, 482–486. [Google Scholar] [CrossRef]
- Gould, D.J.; Barnett, H.C.; Suyemoto, W. Transmission of Japanese encephalitis virus by Culex gelidus Theobald. Trans. R. Soc. Trop. Med. Hyg. 1962, 56, 429–435. [Google Scholar] [CrossRef] [Green Version]
- Hameed, M.; Liu, K.; Anwar, N.; Wahaab, A.; Safdar, A.; Di, D.; Boruah, P.; Xu, J.; Wang, X.; Li, B.; et al. The emerged genotype I of Japanese encephalitis virus shows an infectivity similar to genotype III in Culex pipiens mosquitoes from China. PLoS Negl. Trop. Dis. 2019, 13, e0007716. [Google Scholar] [CrossRef] [Green Version]
- Weng, M.-H.H.; Lien, J.-C.C.; Lin, C.-C.C.; Yao, C.-W.W. Vector competence of Culex pipiens molestus (Diptera: Culicidae) from Taiwan for a sympatric strain of Japanese encephalitis virus. J. Med. Entomol. 2000, 37, 780–783. [Google Scholar] [CrossRef] [PubMed]
- Turell, M.J.; Mores, C.N.; Dohm, D.J.; Komilov, N.; Paragas, J.; Lee, J.S.; Shermuhemedova, D.; Endy, T.P.; Kodirov, A.; Khodjaev, S. Laboratory transmission of Japanese encephalitis and West Nile viruses by molestus form of Culex pipiens (Diptera: Culicidae) collected in Uzbekistan in 2004. J. Med. Entomol. 2006, 43, 296–300. [Google Scholar] [CrossRef]
- Doi, R.; Oya, A.; Shirasaka, A.; Yabe, S.; Sasa, M. Studies on Japanese encephalitis virus infection of reptiles. II. Role of lizards on hibernation of Japanese encephalitis virus. Jpn. J. Exp. Med. 1983, 53, 125–134. [Google Scholar] [PubMed]
- Turell, M.J.; Mores, C.N.; Dohm, D.J.; Lee, W.-J.; Kim, H.-C.; Klein, T.A. Laboratory transmission of Japanese encephalitis, West Nile, and Getah viruses by mosquitoes (Diptera: Culicidae) collected near Camp Greaves, Gyeonggi province, Republic of Korea 2003. J. Med. Entomol. 2006, 43, 1076–1081. [Google Scholar] [CrossRef] [PubMed]
- Mourya, D.T.; Mishra, A.C.; Soman, R.S. Transmission of Japanese encephalitis virus in Culex pseudovishnui & C. tritaeniorhynchus mosquitoes. Indian J. Med. Res. 1991, 93, 250–252. [Google Scholar] [PubMed]
- Mourya, D.T.; Mishra, A.C. Antigen distribution pattern of Japanese encephalitis virus in Culex tritaeniorhynchus, C. vishnui & C. pseudovishnui. Indian J. Med. Res. 2000, 111, 157–161. [Google Scholar] [PubMed]
- Heathcote, O.H.U. Japanese encephalitis in Sarawak: Studies on juvenile mosquito populations. Trans. R. Soc. Trop. Med. Hyg. 1970, 64, 483–488. [Google Scholar] [CrossRef]
- Beerntsen, B.T.; James, A.A.; Christensen, B.M. Genetics of mosquito vector competence. Microbiol. Mol. Biol. Rev. 2000, 64, 115–137. [Google Scholar] [CrossRef] [Green Version]
- Kauffman, E.B.; Kramer, L.D. Zika virus mosquito vectors: Competence, biology, and vector control. J. Infect. Dis. 2017, 216, S976–S990. [Google Scholar] [CrossRef] [Green Version]
- Rocklöv, J.; Dubrow, R. Author Correction: Climate change: An enduring challenge for vector-borne disease prevention and control. Nat. Immunol. 2020, 21, 695. [Google Scholar] [CrossRef]
- Githeko, A.K.; Lindsay, S.W.; Confalonieri, U.E.; Patz, J.A. Climate change and vector-borne diseases: A regional analysis. Bull. World Health Organ. 2000, 78, 1136–1147. [Google Scholar]
- Rosen, L.; Tesh, R.B.; Lien, J.C.; Cross, J.H. Transovarial transmission of Japanese encephalitis virus by mosquitoes. Science 1978, 199, 909–911. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Wang, Y.; Atoni, E.; Zhang, B.; Yuan, Z. Mosquito-associated viruses in China. Virol. Sin. 2018, 33, 5–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, S.-I.; Lee, Y.-M. Early events in Japanese encephalitis virus infection: Viral entry. Pathogens 2018, 7, 68. [Google Scholar] [CrossRef] [Green Version]
- Larish, L.B.; Savage, H.M. Introduction and establishment of Aedes (Finlaya) Japonicus japonicus (Theobald) on the island of Hawaii: Implications for arbovirus transmission. J. Am. Mosq. Control Assoc. 2005, 21, 318–321. [Google Scholar] [CrossRef]
- Smitz, N.; De Wolf, K.; Deblauwe, I.; Kampen, H.; Schaffner, F.; De Witte, J.; Schneider, A.; Verlé, I.; Vanslembrouck, A.; Dekoninck, W.; et al. Population genetic structure of the Asian bush mosquito, Aedes japonicus (Diptera, Culicidae), in Belgium suggests multiple introductions. Parasites Vectors 2021, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Versteirt, V.; Schaffner, F.; Garros, C.; Dekoninck, W.; Coosemans, M.; Van Bortel, W. Introduction and establishment of the exotic mosquito species Aedes japonicus japonicus (Diptera: Culicidae) in Belgium. J. Med. Entomol. 2009, 46, 1464–1467. [Google Scholar] [CrossRef] [Green Version]
- Kaufman, M.G.; Fonseca, D.M. Invasion biology of Aedes japonicus japonicus (Diptera: Culicidae). Annu. Rev. Entomol. 2014, 59, 31. [Google Scholar] [CrossRef] [Green Version]
- Dussault, C.; Nelder, M.P.; Russell, C.; Johnson, S.; Vrbova, L. Evaluating the impact of Aedes japonicus invasion on the mosquito community in the Greater Golden Horseshoe region (Ontario, Canada). PLoS ONE 2018, 13, e0208911. [Google Scholar] [CrossRef]
- Oliveira, A.R.; Strathe, E.; Etcheverry, L.; Cohnstaedt, L.W.; McVey, D.S.; Piaggio, J.; Cernicchiaro, N. Assessment of data on vector and host competence for Japanese encephalitis virus: A systematic review of the literature. Prev. Vet. Med. 2018, 154, 71–89. [Google Scholar] [CrossRef] [PubMed]
- Peach, D.A.H.; Almond, M.; Pol, J.C. Modeled distributions of Aedes japonicus japonicus and Aedes togoi (Diptera: Culicidae) in the United States, Canada, and northern Latin America. J. Vector Ecol. 2019, 44, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Invasive Species Compendium. Available online: https://www.cabi.org/isc/ (accessed on 21 December 2021).
- European Centre for Disease Prevention and Control and European Food Safety Authority. Aedes japonicus—Current Known Distribution: March 2021. 2021. Available online: https://www.ecdc.europa.eu/en/publications-data/aedes-japonicus-current-known-distribution-march-2021 (accessed on 4 June 2021).
- Hoffmann, J.A. Immune responsiveness in vector insects. Proc. Natl. Acad. Sci. USA 1997, 94, 11152–11153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baxter, R.H.G.; Contet, A.; Krueger, K. Arthropod innate immune systems and vector-borne diseases. Biochemistry 2017, 56, 907–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Srivastava, P.; Sirisena, P.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito innate immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabrieli, P.; Caccia, S.; Varotto-Boccazzi, I.; Arnoldi, I.; Barbieri, G.; Comandatore, F.; Epis, S. Mosquito trilogy: Microbiota, immunity and pathogens, and their implications for the control of disease transmission. Front. Microbiol. 2021, 12, 630438. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito defense strategies against viral infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.S.; Webster, J.A.; Madzokere, E.T.; Stephenson, E.B.; Herrero, L.J. Mosquito antiviral defense mechanisms: A delicate balance between innate immunity and persistent viral infection. Parasites Vectors 2019, 12, 165. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.K.M.; Han, Y.S.; Jo, Y.H. An overview of insect innate immunity. Entomol. Res. 2020, 50, 282–291. [Google Scholar] [CrossRef]
- Flemming, A. Mechanism of adaptive immunity found in the fruitfly. Nat. Rev. Immunol. 2017, 17, 279. [Google Scholar] [CrossRef] [PubMed]
- Mongelli, V.; Saleh, M.C. Bugs are not to be silenced: Small RNA pathways and antiviral responses in insects. Annu. Rev. Virol. 2016, 3, 573–589. [Google Scholar] [CrossRef]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef]
- Romoser, W.S.; Turell, M.J.; Lerdthusnee, K.; Neira, M.; Dohm, D.; Ludwig, G.; Wasieloski, L. Pathogenesis of Rift Valley fever virus in mosquitoes: Tracheal conduits & the basal lamina as an extra-cellular barrier. Infect. Dis. Nat. Mech. Viral Emerg. Persistence 2005, 19, 89–100. [Google Scholar] [CrossRef]
- Takahashi, M. Differential transmission efficiency for Japanese encephalitis virus among colonized strains of Culex tritaeniorhynchus. Jpn. J. Sanit. Zool. 1982, 33, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Sanchez-Vargas, I.; Olson, K.; Black, W. The genetic basis for salivary gland barriers to arboviral transmission. Insects 2021, 12, 73. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Kuwata, R.; Hoshino, K.; Isawa, H.; Sawabe, K.; Kobayashi, M. Argonaute 2 suppresses Japanese encephalitis virus infection in Aedes aegypti. Jpn. J. Infect. Dis. 2017, 70, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Terradas, G.; Joubert, D.A.; McGraw, E.A. The RNAi pathway plays a small part in Wolbachia-mediated blocking of dengue virus in mosquito cells. Sci. Rep. 2017, 7, 43847. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [Green Version]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Théron, E.; Dennis, C.; Brasset, E.; Vaury, C. Distinct features of the piRNA pathway in somatic and germ cells: From piRNA cluster transcription to piRNA processing and amplification. Mob. DNA 2014, 5, 28. [Google Scholar] [CrossRef]
- Lin, C.-C.; Chou, C.-M.; Hsu, Y.-L.; Lien, J.-C.; Wang, Y.-M.; Chen, S.-T.; Tsai, S.-C.; Hsiao, P.-W.; Huang, C.-J. Characterization of two mosquito STATs, AaSTAT and CtSTAT. J. Biol. Chem. 2004, 279, 3308–3317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, C.; Sun, P.; Yu, X.; Wang, P.; Cheng, G. Roles of Symbiotic Microorganisms in Arboviral Infection of Arthropod Vectors; Elsevier: Amsterdam, The Netherlands, 2020; Volume 36. [Google Scholar] [CrossRef]
- Zhag, R.; Zhu, Y.; Pang, X.; Xiao, X.; Zhang, R.; Cheng, G. Regulation of antimicrobial peptides in Aedes aegypti Aag2 cells. Front. Cell. Infect. Microbiol. 2017, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Schmid-Hempel, P. Evolutionary ecology of insect immune defenses. Annu. Rev. Entomol. 2005, 50, 529–551. [Google Scholar] [CrossRef]
- Liu, K.; Xiao, C.; Xi, S.; Hameed, M.; Wahaaab, A.; Shao, D.; Li, Z.; Li, B.; Wei, J.; Qiu, Y.; et al. Mosquito defensins enhance Japanese encephalitis virus infection by facilitating virus adsorption and entry within the mosquito. J. Virol. 2020, 94, e01164-20. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, T.; Oliveira, I.F.; Melo-Santos, M.A.V.; Oliveira, C.M.F.; Lima, C.A.; Ayres, C.F.J. Expression of defensin, cecropin, and transferrin in Aedes aegypti (Diptera: Culicidae) infected with Wuchereria bancrofti (Spirurida: Onchocercidae), and the abnormal development of nematodes in the mosquito. Exp. Parasitol. 2008, 120, 364–371. [Google Scholar] [CrossRef]
- Liu, K.; Hou, F.; Wahaab, A.; Kang, L.; Xie, F.; Ma, X.; Xia, Q.; Xiao, C.; Shao, D.; Li, B.; et al. Mosquito defensin facilitates Japanese encephalitis virus infection by downregulating the C6/36 cell-surface antiviral protein HSC70B. Vet. Microbiol. 2021, 253, 108971. [Google Scholar] [CrossRef] [PubMed]
- Jupatanakul, N.; Sim, S.; Dimopoulos, G. The insect microbiome modulates vector competence for arboviruses. Viruses 2014, 6, 4294–4313. [Google Scholar] [CrossRef]
- Dennison, N.J.; Jupatanakul, N.; Dimopoulos, G. The mosquito microbiota influences vector competence for human pathogens. Curr. Opin. Insect. Sci. 2014, 3, 6–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, F.H.; Gendrin, M.; Wyer, C.A.S.; Christophides, G.K. Microbiota-induced peritrophic matrix regulates midgut homeostasis and prevents systemic infection of malaria vector mosquitoes. PLoS Pathog. 2017, 13, e1006391. [Google Scholar] [CrossRef]
- Rancès, E.; Ye, Y.H.; Woolfit, M.; McGraw, E.A.; O’Neill, S.L. The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS Pathog. 2012, 8, e1002548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, K.-H.; Huang, C.-G.; Wu, W.-J.; Chuang, C.-K.; Lin, C.-C.; Chen, W.-J. Parallel infection of Japanese encephalitis virus and Wolbachia within cells of mosquito salivary glands. J. Med. Entomol. 2006, 43, 752–756. [Google Scholar] [CrossRef]
- Mourya, D.T.; Soman, R.S. Effect of gregarine parasite, Ascogregarina culicis & tetracycline on the susceptibility of Culex bitaeniorhynchus to JE virus. Indian J. Med. Res. 1985, 81, 247–250. [Google Scholar] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.; McMeniman, C.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Johnson, K.N. The impact of Wolbachia on virus infection in mosquitoes. Viruses 2015, 7, 5705–5717. [Google Scholar] [CrossRef] [Green Version]
- Van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.D.; McElroy, K.; Day, A.; Higgs, S.; O’Neill, S.L. Impact of Wolbachia on infection with Chikungunya and Yellow Fever viruses in the mosquito vector Aedes aegypti. PLoS Negl. Trop. Dis. 2012, 6, e1892. [Google Scholar] [CrossRef] [Green Version]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23. [Google Scholar] [CrossRef] [Green Version]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mosquito Species | Country of Sampling | JEV Genotype and/or Strain | Detection Method | Total # Tested | # JEV Positive Pools/Total # of Pools | MIR (# Positive Pools/Total Tested) × 100 | Reference |
|---|---|---|---|---|---|---|---|
| Aedes albopictus (Skuse, 1895) | Taiwan | G-I and III | RT-PCR | 177 | 1/25 | 0.56% | [19] |
| Taiwan | ND | Isolation | ND | 20 | ND | [20] | |
| Aedes butleri (Theobald, 1901) | Malaysia | ND | Isolation and RT-PCR | 3950 | 4/79 | 0.1% | [21,22] |
| Aedes curtipes (Edwards, 1915) | Malaysia | ND | ND | ND | ND | ND | [23] |
| Aedes lineatopennis (Ludlow, 1905) | Malaysia | ND | Isolation and RT-PCR | 300 | 1/6 | 0.33% | [22] |
| Aedes vexans (Meigen, 1830) | Taiwan | G-I and III | RT-PCR | 246 | 3/32 | 1.22% | [19] |
| Taiwan | ND | Isolation | ND | 1 | ND | [20] | |
| Taiwan | ND | RT-PCR | 61 | 1/9 | 1.64% | [24] | |
| Aedes vigilax (Skuse, 1889) | Australia | ND | Isolation | ND | 1 | ND | [25] |
| Australia | G-II | Isolation | 3073 | 1 | 0.03% | [26] | |
| Anopheles annularis (Wulp, 1884) | Indonesia | ND | Isolation | 250 | 1/28 | 0.4% | [27] |
| Anopheles barbirostris (Wulp, 1884) | India | ND | ELISA and Toxo-IFA | 22 | 1/8 | 4.55% | [28] |
| India | ND | ND | ND | ND | ND | [29] | |
| Anopheles minimus (Theobald, 1901) | Taiwan | G-I and III | RT-PCR | 18 | 1/7 | 5.56% | [19] |
| Anopheles hyrcanus (Pallas, 1771) | India | ND | ND | ND | ND | ND | [30] |
| India | ND | ND | ND | ND | ND | [29] | |
| Anopheles pallidus (Theobald, 1901) | India | ND | ELISA and Toxo-IFA | 28 | 1/12 | 3.57% | [28] |
| Anopheles peditaeniatus (Leicester, 1908) | India | ND | Isolation | 6306 | 1/133 | 0.02% | [31] |
| Anopheles sinensis (Wiedemann, 1828) | China | G-III | RT-PCR | ND | 12 | ND | [32] |
| China | ND | RT-PCR | 2802 | 5/55 | 0.18% | [33] | |
| China | G-I | Isolation | 14,170 | 3 | 0.02% | [34] | |
| Taiwan | G-I and III | RT-PCR | 2638 | 6/119 | 0.23% | [19] | |
| Anopheles subpictus (Grassi, 1899) | India | ND | ELISA and Toxo-IFA | 1432 | 7/67 | 0.49% | [28] |
| India | ND | ELISA and Toxo-IFA | ND | ND | ND | [35] | |
| India | ND | ELISA and Toxo-IFA | 6550 | 4/131 | 0.06% | [36] | |
| India | ND | ND | ND | ND | ND | [37] | |
| Anopheles tessellatus (Theobald, 1901) | Taiwan | G-I and III | RT-PCR | 536 | 2/31 | 0.37% | [19] |
| Anopheles vagus (Dönitz, 1902) | Indonesia | ND | Isolation | 2700 | 1/42 | 0.04% | [27] |
| Armigeres subalbatus (Coquillet, 1898) | China | G-I | Isolation | 394 | 2 | 0.51% | [34] |
| China | G-III | RT-PCR | ND | 3 | ND | [32] | |
| India | ND | ELISA and Toxo-IFA | 110 | 1/21 | 0.91% | [28] | |
| Taiwan | G-I and III | RT-PCR | 225 | 3/30 | 1.33% | [19] | |
| Taiwan | ND | Isolation | ND | 8 | ND | [20] | |
| Coquillettidia ochracea (Theobald, 1903) | China | ND | RT-PCR | 155 | 1/6 | 0.65% | [38] |
| Culex annulirostris (Skuse, 1889) | Australia | G-III | RT-PCR | 2871 | 8/134 | 0.28% | [39] |
| Australia | ND | Isolation | 23,890 | 42 | 0.18% | [25] | |
| Culex annulus (Theobald, 1901) | Taiwan | G-I and III | RT-PCR | 991 | 9/79 | 0.91% | [19] |
| Taiwan | ND | Intracerebral inoculation of mice | 1338 | 3 | 0.23% | [40] | |
| Taiwan | ND | Isolation | 54,910 | 31/703 | 0.06% | [41] | |
| Taiwan | ND | Isolation | ND | 7/31 | ND | [42] | |
| Taiwan | ND | Isolation | ND | 1 | ND | [20] | |
| Culex bitaeniorhynchus (Giles, 1901) | India | ND | ELISA and Toxo-IFA | 44 | 1/9 | 2.28% | [28] |
| Korea | G-I | RT-PCR | 344 | 1/26 | 0.29% | [43] | |
| Korea | G-I and V | RT-PCR | 1960 | 2/175 | 0.1% | [44] | |
| Korea | ND | RT-PCR | 1140 | 1/45 | 0.09% | [45] | |
| Malaysia | ND | Isolation and RT-PCR | 550 | 3/11 | 0.55% | [21] | |
| Culex epidesmus (Theobald, 1905) | India | ND | Isolation | ND | 1 | ND | [30] |
| Culex fuscanus (Wiedemann, 1820) | Taiwan | ND | Isolation | ND | 1 | ND | [20] |
| Culex fuscocephala (Theobald, 1907) | India | ND | Isolation | 14,664 | 1/257 | 0.007% | [31] |
| India | ND | Isolation | 15,250 | 6/305 | 0.04% | [46] | |
| Indonesia | ND | Isolation | ND | 1 | ND | [47] | |
| Malaysia | ND | Isolation and RT-PCR | 3800 | 2/76 | 0.05% | [22] | |
| Taiwan | G-I and III | RT-PCR | 394 | 3/19 | 0.76% | [19] | |
| Taiwan | ND | RT-PCR | 1150 | 1/23 | 0.09% | [48] | |
| Taiwan | ND | Isolation | 22,895 | 19/282 | 0.08% | [49] | |
| Thailand | ND | Isolation or HI and CF assays | 142,375 | 2 | 0.001% | [50] | |
| Thailand | ND | ND | ND | 2 | ND | [50] | |
| Culex gelidus (Theobald, 1901) | Australia | ND | RT-PCR | 4872 | 3 | 0.06% | [51] |
| India | ND | Isolation | 6038 | 3/127 | 0.05% | [31] | |
| India | ND | ND | ND | ND | ND | [52] | |
| India | ND | ELISA and Toxo-IFA | 7485 | 4/177 | 0.05% | [53] | |
| India | ND | Isolation | 9700 | 5/194 | 0.05% | [46] | |
| India | ND | ELISA and Toxo-IFA | 8750 | 17/175 | 0.2% | [54] | |
| India | ND | ELISA and Toxo-IFA | 17,678 | 12/403 | 0.07% | [55] | |
| India | ND | ELISA and Toxo-IFA | 16,658 | 10 | 0.06% | [56] | |
| Indonesia | ND | ND | ND | 1 | ND | [57] | |
| Indonesia | ND | Isolation | 7144 | 2/154 | 0.03% | [47] | |
| Malaysia | ND | Isolation and RT-PCR | 11,200 | 12/224 | 0.11% | [22] | |
| Malaysia | ND | ND | ND | 1 | ND | [58] | |
| Malaysia | ND | ND | ND | ND | ND | [23] | |
| Sri Lanka | ND | Isolation | 13,043 | 4 | 0.03% | [59] | |
| Thailand | ND | Isolation or HI and CF assays | 11,495 | 3 | 0.03% | [50] | |
| Thailand | Not given | Inoculation in mice | 3097 | 18 | 0.6% | [60] | |
| Vietnam | Not given | ND | ND | ND | ND | [61] | |
| Culex infula (Theobald, 1901) | India | ND | ELISA and Toxo-IFA | 119 | 2/16 | 1.68% | [28] |
| Culex orientalis (Edwards, 1921) | Korea | G-V | RT-PCR | 498 | 5/83 | 1% | [62] |
| Culex pipiens (Linnaeus, 1758) | Italy | G-III | RT-PCR | ND | 1/57 | ND | [63] |
| Korea | G-I | RT-PCR | 736 | 4/64 | 0.54% | [43] | |
| Korea | G-I | RT-PCR | 11,237 | 4/804 | 0.04% | [44] | |
| Korea | G-V | RT-PCR | 9295 | 1/264 | 0.01% | [62] | |
| China | G-I | RT-PCR | 1540 | 1/256 | 0.06% | [64] | |
| Culex pipiens pallens (Coquillett, 1898) | China | ND | RT-PCR | 6465 | 10/132 | 0.15% | [38] |
| Culex pseudovishnui (Colless, 1957) | India | ND | ND | ND | 1 | ND | [65] |
| India | ND | ELISA and Toxo-IFA or RT-PCR | ND | 3/107 | ND | [66] | |
| India | ND | ELISA and Toxo-IFA or RT-PCR | 1406 | 1 | 0.07% | [67] | |
| India | ND | ND | ND | ND | ND | [68] | |
| India | ND | ND | ND | ND | ND | [37] | |
| Culex quinquefasciatus (Say, 1823) | India | ND | ELISA and Toxo-IFA | 59 | 1/13 | 1.69% | [28] |
| India | ND | Isolation | 304 | 1/18 | 0.33% | [31] | |
| Malaysia | ND | Isolation and RT-PCR | 2400 | 1/48 | 0.4% | [22] | |
| Taiwan | G-I and III | RT-PCR | 1333 | 2/74 | 0.15% | [19] | |
| Taiwan | ND | Isolation | ND | 7 | ND | [20] | |
| Thailand | ND | Isolation | 1023 | 2/25 | 0.2% | [69] | |
| Vietnam | G-III | RT-PCR | ND | 30 | ND | [70] | |
| Culex rubithoracis (Leicester, 1908) | Taiwan | ND | RT-PCR | 130 | 4/22 | 3.08% | [24] |
| Culex sitiens (Wiedemann, 1828) | Australia | ND | RT-PCR | 18,680 | 5 | 0.03% | [51] |
| Australia | ND | RT-PCR | 22,833 | 1 | 0.004% | [71] | |
| Australia | G-II | Isolation | 25,292 | 42 | 0.16% | [26] | |
| Australia | G-I | Isolation | 44,755 | 1 | 0.002% | [72] | |
| Malaysia | ND | Isolation and RT-PCR | 400 | 2/8 | 0.5% | [21] | |
| Papua New Guinea | G-II | Isolation | 245,483 | 3 | 0.001% | [73] | |
| Taiwan | ND | RT-PCR | 604 | 1/34 | 0.17% | [24] | |
| Taiwan | ND | Isolation | ND | 2 | ND | [20] | |
| Vietnam | G-I and III | RT-PCR | ND | 73 | ND | [70] | |
| Culex tarsalis (Coquillett, 1896) | China | G-III | RT-PCR | ND | 57 | ND | [32] |
| Culex tritaeniorhynchus (Giles, 1901) | Cambodia | G-I | Isolation | 7218 | 1/729 | 0.01% | [74] |
| China | ND | RT-PCR | 6610 | 31/135 | 0.47% | [38] | |
| China | ND | RT-PCR | 15,795 | 24/158 | 0.15% | [33] | |
| China | G-I | Isolation | 37,119 | 15 | 0.04% | [34] | |
| China | G-I | RT-PCR | 3945 | 4/255 | 0.1% | [64] | |
| China | G-I | RT-PCR | 6490 | 15/149 | 0.23% | [75] | |
| China | G-I | RT-PCR | 2927 | 3/152 | 0.1% | [76] | |
| India | ND | ELISA and Toxo-IFA | 9937 | 10/245 | 0.10% | [28] | |
| India | ND | Isolation | 12,161 | 2/272 | 0.02% | [31] | |
| India | ND | Isolation | 206,424 | 58/4128 | 0.03% | [46] | |
| India | ND | ELISA and Toxo-IFA | ND | ND | ND | [35] | |
| India | ND | ELISA and Toxo-IFA | 7485 | 4/177 | 0.05% | [53] | |
| India | ND | ELISA and Toxo-IFA | 45,100 | 62/902 | 0.14% | [54] | |
| India | ND | ELISA and Toxo-IFA | 21,005 | 13/429 | 0.06% | [53] | |
| India | ND | ELISA and Toxo-IFA | 14,358 | 14/309 | 0.1% | [55] | |
| India | ND | ELISA and Toxo-IFA | 100,611 | 64 | 0.06% | [56] | |
| India | ND | ELISA and Toxo-IFA or RT-PCR | 862 | 2 | 0.23% | [67] | |
| Indonesia | ND | Isolation | 112,398 | 1/596 | 0.0009% | [27] | |
| Indonesia | ND | Isolation | 18,486 | 19/359 | 0.1% | [47] | |
| Japan | G-I | Isolation | 3328 | 3/141 | 0.09% | [77] | |
| Korea | G-I | RT-PCR | 2880 | 29/121 | 1.01% | [43] | |
| Korea | G-I and V | RT-PCR | 55,135 | 92/2031 | 0.17% | [44] | |
| Korea | ND | RT-PCR | 5909 | 50/207 | 0.85% | [45] | |
| Malaysia | ND | Isolation and RT-PCR | 1300 | 3/26 | 0.23% | [21] | |
| Malaysia | ND | Isolation and RT-PCR | 36,550 | 24/731 | 0.07% | [22] | |
| Singapore | G-II | RT-PCR | 882 | 5/88 | 0.57% | [78] | |
| Sri Lanka | ND | Isolation | 17,436 | 4 | 0.02% | [59] | |
| Taiwan | ND | Isolation | 16,776 | 18/267 | 0.11% | [41] | |
| Taiwan | ND | RT-PCR | 28,773 | 95/1061 | 0.33% | [24] | |
| Taiwan | ND | RT-PCR | 37,500 | 25/750 | 0.07% | [48] | |
| Taiwan | ND | Isolation | ND | 97 | ND | [20] | |
| Taiwan | G-I and III | RT-PCR | 89,189 | 468/2242 | 0.52% | [19] | |
| Taiwan | ND | Isolation | ND | 2/6 | ND | [42] | |
| Thailand | ND | Isolation or HI and CF assays | 183,140 | 8 | 0.004% | [50] | |
| Thailand | ND | Isolation | 290,126 | 34 | 0.01% | [79] | |
| Vietnam | G-I and III | RT-PCR | ND | 3 | ND | [70] | |
| Vietnam | G-I | Isolation | 4199 | 3/131 | 0.07% | [80] | |
| Culex vishnui (Theobald, 1901) | India | ND | ELISA and Toxo-IFA or RT-PCR | 1512 | 3 | 0.2% | [67] |
| India | ND | ND | ND | ND | ND | [37] | |
| India | ND | ELISA and Toxo-IFA | 2787 | 1/61 | 0.04% | [53] | |
| India | ND | Isolation | 54,007 | 22/1080 | 0.04% | [46] | |
| Indonesia | ND | Isolation | ND | 1 | ND | [47] | |
| Malaysia | ND | Isolation and RT-PCR | 1650 | 4/33 | 0.24% | [21] | |
| Thailand | ND | Isolation | 8408 | 1 | 0.01% | [79] | |
| Vietnam | G-I | Isolation | 1542 | 2/46 | 0.13% | [80] | |
| Culex whitmorei (Giles, 1904) | India | ND | ELISA and Toxo-IFA | 47 | 2/17 | 4.26% | [28] |
| Sri Lanka | ND | Isolation | 167 | 1 | 0.6% | [59] | |
| Mansonia bonneae/dives (Edwards, 1930/Schiner, 1868) | Malaysia | ND | ND | ND | ND | ND | [23] |
| Mansonia annulifera (Theobald, 1901) | India | ND | ELISA and Toxo-IFA | ND | ND | ND | [35] |
| India | ND | ELISA and Toxo-IFA | 4530 | 3 | 0.07% | [56] | |
| Mansonia indiana (Edwards, 1930) | India | ND | ELISA and Toxo-IFA | 12,362 | 12 | 0.1% | [56] |
| India | ND | ELISA and Toxo-IFA | 62 | 2/13 | 3.23% | [28] | |
| Mansonia uniformis (Theobald, 1901) | India | ND | ELISA and Toxo-IFA | ND | ND | ND | [35] |
| India | ND | ELISA and Toxo-IFA | 14,503 | 5 | 0.03% | [56] | |
| Malaysia | ND | ND | ND | ND | ND | [23] | |
| Sri Lanka | ND | ND | ND | ND | ND | [59] | |
| Taiwan | G-I and III | RT-PCR | 75 | 1/19 | 1.33% | [19] |
| Mosquito Species | Country of Sampling | JEV Genotype and/or Strain | Detection Method | Total # Tested | # JEV Positive Pools | Reference |
|---|---|---|---|---|---|---|
| Aedes aegypti (Linnaeus, 1762) | Taiwan | G-I and III | RT-PCR | 3 | 0/2 pools | [19] |
| Aedes albolateralis (Theobald, 1908) | Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] |
| Aedes albopictus * | Korea | G-V | RT-PCR | 564 | 0/64 pools | [62] |
| Korea | G-I | RT-PCR | 66 | 0/15 pools | [43] | |
| Aedes dorsalis (Meigen, 1830) | Korea | G-V | RT-PCR | 6 | 0/6 pools | [62] |
| Aedes koreicus (Edwards, 1917) | Korea | G-I | RT-PCR | 181 | 0/24 pools | [43] |
| Aedes lineatopennis * | Korea | G-I | RT-PCR | 1 | 0/1 pools | [43] |
| Thailand | ND | Isolation or HI and CF assays | 16,230 | 0 pools | [50] | |
| Aedes mediolineatus (Theobald, 1901) | Thailand | ND | Isolation or HI and CF assays | 15,122 | 0 pools | [50] |
| Aedes nipponicus (LaCasse & Yamaguti, 1948) | Korea | G-I | RT-PCR | 1 | 0/1 pools | [43] |
| Aedes penghuensis (Lien, 1968) | Taiwan | G-I and III | RT-PCR | 283 | 0/10 pools | [19] |
| Aedes togoi (Theobald, 1907) | Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] |
| Aedes vexans *[81] | Thailand | ND | Isolation or HI and CF assays | 11,022 | 0 pools | [50] |
| Aedes vexans nipponii (Theobald, 1907) | Korea | G-I | RT-PCR | 2091 | 0/106 pools | [43] |
| Anopheles ludlowae (Theobald, 1903) | Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] |
| Armigeres subalbatus * | Korea | G-V | RT-PCR | 1132 | 0/145 pools | [62] |
| Korea | G-I | RT-PCR | 23 | 0/9 pools | [43] | |
| Coquillettidia crassipes (Van der Wulp, 1881) | Taiwan | G-I and III | RT-PCR | 47 | 0/3 pools | [19] |
| Coquillettidia ochracea * | Korea | G-V | RT-PCR | 115 | 0/14 pools | [62] |
| Culex bitaeniorhynchus * | Korea | G-V | RT-PCR | 50 | 0/16 pools | [62] |
| Taiwan | G-I and III | RT-PCR | 60 | 0/7 pools | [19] | |
| Culex brevipalpis (Giles, 1902) | Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] |
| Culex fuscanus * | Taiwan | G-I and III | RT-PCR | 4 | 0/3 pools | [19] |
| Culex fuscocephalus* | Thailand | ND | Isolation | 9140 | 0 pools | [79] |
| Culex gelidus * | Thailand | ND | Isolation | 17,530 | 0 pools | [79] |
| Culex hayshii (Yamada, 1917) | Korea | G-V | RT-PCR | 4 | 0/2 pools | [62] |
| Culex inatomii (Kaminura & Wada, 1974) | Korea | G-V | RT-PCR | 470 | 0/16 pools | [62] |
| Korea | G-I | RT-PCR | 1 | 0/1 pools | [43] | |
| Culex mimeticus (Noè, 1899) | Korea | G-V | RT-PCR | 1 | 0/1 pools | [62] |
| Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] | |
| Culex murrelli (Lien, 1968) | Taiwan | G-I and III | RT-PCR | 39 | 0/3 pools | [19] |
| Culex nigropunctatus (Edwards, 1926) | Taiwan | G-I and III | RT-PCR | 9 | 0/1 pools | [19] |
| Culex orientalis * | Korea | G-I | RT-PCR | 3 | 0/2 pools | [43] |
| Culex quinquefasciatus * | Thailand | ND | Isolation | 73 | 0 pools | [79] |
| Culex rubensis (Sasa & Takahashi, 1948) | Korea | G-V | RT-PCR | 1 | 0/1 pools | [62] |
| Culex rubithoracis * | Taiwan | G-I and III | RT-PCR | 65 | 0/8 pools | [19] |
| Culex sitiens * | Taiwan | G-I and III | RT-PCR | 6295 | 0/128 pools | [19] |
| Culex tritaeniorhynchus * | Korea | G-V | RT-PCR | 10 | 0/7 pools | [62] |
| Culex vagans (Wiedemann, 1828) | Korea | G-V | RT-PCR | 5 | 0/2 pools | [62] |
| Culex vishnui * | Thailand | ND | Isolation or HI and CF assays | 22,005 | 0 pools | [50] |
| Culex whitmorei * | Thailand | ND | Isolation | 530 | 0 pools | [79] |
| Culiseta bergrothi (Edwards, 1921) | Korea | G-V | RT-PCR | 1 | 0/1 pools | [62] |
| Mansonia uniformis * | Korea | G-V | RT-PCR | 2176 | 0/66 pools | [62] |
| Korea | G-I | RT-PCR | 1 | 0/1 pools | [43] | |
| Tripteroides bambusa (Yamada, 1917) | Korea | G-V | RT-PCR | 30 | 0/9 pools | [62] |
| Uranotaenia macfarlanei (Edwards, 1914) | Taiwan | G-I and III | RT-PCR | 1 | 0/1 pools | [19] |
| Mosquito Species | Origin of Mosquito Colony | JEV Strain Used | Cell Type Used for Virus Production | Virus Titer in Bloodmeal | Blood Origin | Feeding Method | Inc. Temperature | Inc. Period (Days) | # Mosquitoes | % Infected * | % Disseminated ** | % Transmission Competent *** | Detection Method | Reference |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Aedes aegypti | Australia, Townsville | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 14–15 | 60 | 27% | 17% | ND | Porcine stable-equine kidney cells | [93] |
| Aedes albopictus | Australia, Masig Island | G-I (TS00) | Porcine stable equine kidney and C6/36 cells | 3.5 ± 0.3 log10 CCID50/mL | Washed defibrinated sheep | Cotton pledged | 28 °C | 14 | 25 | 20% | 16% | 16% | Vero cells | [94] |
| France, Montpellier and Nice | G-III (RP-9) and G-V (XZ0934) | Chicken fibroblast-derived DF1 cells | 8 × 106 FFU/mL | Washed rabbit erythrocytes | Cotton pledgets | 26 °C | 7–13 | 5–20 | 70–100% | 57–100% ◊ | 20–63% | BHK-21 cells | [87] | |
| Taiwan, Tapei and Taichung County | ND (Sanshia MQ1-2) | C6/36 cells | 5.42 log10 WMICLD50 | NA | Intraperitoneal inoculated mice | 26–28 °C | 14 | 20 | ND | ND | 27–45% | BHK-21 cells | [95] | |
| Aedes detritus (Haliday, 1833) | UK, Northwest England | G-V (Muar) | Vero cells | 4 log10 PFU/mL | Defibrinated horse | Hemotek with Parafilm membrane | 23 and 28 °C | 0–21 | 6–32 | 32–100% | 20–100% | 3–67% | Vero cells | [85] |
| Aedes dorsalis | US | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 16 | 2–10 | ND | ND | 4% # | Development of encephalitis in laboratory-reared mice | [96] |
| Aedes japonicus (Theobald, 1901) | Germany, Stuttgart | ND | ND | ND | Human | Cotton pledgets | 25 °C | 0–14 | 3–4 | 100% | ND | ND | RT-qPCR | [97] |
| Japan, Narita | G-I (17CxIT-I4-D31), 3 (JaGAr 01) and V (Muar) | C6/36 cells | 8.9, 8.6, and 7.1 log10 FFU/mL | Defibrinated rabbit | Hemotek with pig intestine membrane | 27 °C | 7–14 | 3–36 | 2–19% | 2–19% | 2–16% | RT-qPCR or FFA in Vero cells | [12] | |
| Japan, Sapporo | G-III (JANAr-5681) | C6/36 cells | 6.2 PFU/mL (blood) and 3.7 PFU/mL (chicken) | ND | Cotton pledgets or viremic chicken | 20 or 28 °C | 0–20 | 40 | 67.5% | ND | 50% | BHK-21 cells and IFA | [98] | |
| Aedes kochi (Dönitz, 1901) | Australia, Bamaga and Cairns (wild) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 37 | 19% | ND | 6% | Detection of virus in brain aspirates of recipient suckling mice | [93] |
| Aedes nigromaculis (Ludlow, 1906) | US | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 16 | 11–100 | ND | ND | 4% # | Development of encephalitis in laboratory-reared mice | [96] |
| Aedes notoscriptus(Skuse, 1889) | Australia, Closeburn | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 13/14 | 11–48 | 27% | 8% | 27% | Porcine stable-equine kidney cells | [93] |
| Aedes vexans | Guam | ND (Okinawa, human 1945) | ND | ND | NA | Inoculated mice | ND | ND | ND | ND | ND | Successful | Development of encephalitis in laboratory-reared mice | [99] |
| Aedes vexans nipponii | Japan, Sapporo | G-III (JANAr-5681) | C6/36 cells | 6.2 PFU/mL (blood) and 3.7 PFU/mL (chicken) | ND | Cotton pledgets or viremic chicken | 20 or 28 °C | 0–20 | 12 | 25% | ND | ND | BHK-21 cells and IFA | [98] |
| Aedes vigilax | Australia, Cairns (wild) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 75 | 57% | ND | 17% | Detection of virus in brain aspirates of recipient suckling mice | [93] |
| Australia, Redlands Shire | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 107.1±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 9–13 | 4–62 | 19–39% | 18–39% | 0% | Porcine stable-equine kidney cells | [93] | |
| Anopheles tessellatus | India | G-I (733913) | NA | ND | Viremic chickens | NA | ND | 11 | 13 | ND | ND | 31% | Transmission to chickens | [100] |
| Armigeres subalbatus | Taiwan, Liu-Chiu | G-III (T1P1) | C6/36 | 1.25 × 107 PFU/mL | Rabbit | Drop of blood | ND | 1–20 | 8–14 | ND | ND | 0–79% | IFAT | [101] |
| Culex annulirostris | Guam | ND (Okinawa, human 1945) | ND | ND | NA | Inoculated mice | ND | ND | ND | ND | ND | Successful | encephalitis in laboratory-reared mice | [99] |
| Australia, Bamaga and Cairns (wild) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 25–57 | 93% | ND | 56% | Detection of virus in brain aspirates of recipient suckling mice | [93] | |
| Australia, Brisbane | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 5–14 | 18–36 | 78–100% | 6–64% | 24–81% | Porcine stable-equine kidney cells | [93] | |
|
Culex bitaeniorhynchus | India | G-I (733913) | NA | ND | Viremic ducklings | NA | ND | 9–12 | 1 | 9–100% | ND | 100% | Transmission to ducklings | [102] |
| India | G-I (733913) | NA | ND | Viremic chickens | NA | ND | 10–12 | 24 | 47–62% | ND | 64–89% | Transmission to chickens | [103] | |
| India | G-I (733913) | NA | ND | Viremic chickens | NA | ND | ND | ND | ND | ND | Successful | Transmission to chickens | [104] | |
|
Culex fuscocephala | Taiwan | ND (TaiAn 171) | NA | 10−0.89–10−1.91 mouse LD50 | NA | Viremic pigs | ND | 12–21 | ND | ND | ND | 0–68% | Transmission to chickens | [105] |
| Thailand, Chiengmai valley | ND (BKM-984-70) | NA | 8 PFU per mosquito | NA | Viremic chicken | ND | 10–27 | ND | 95–100% | ND | 10–20% | Transmission to chickens | [106] | |
| Culex gelidus | Australia, Cairns (wild) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 4 | 100% | ND | 100% | Detection of virus in brain aspirates of recipient suckling mice | [93] |
| US, Malayan strain | ND (FM380) | ND | ND | NA | Viraemic chicken | 27 °C | 6–21 | 4–43 | ND | ND | 8–63% | Development of encephalitis in laboratory-reared mice | [107] | |
| Culex pipiens | China, Shangai | G-I (SH7), G-III (SH15) | C6/36 cells | 4.9–8.3 log TCID50/mL | Defibrinated mice | Hemotek membrane feeding and cotton pledgets | ND | 7–14 | 11–52 | 45% | 30% ◊ | 23% | TCID50 assay on BHK-21 cells | [108] |
| Pennsylvania, US | G-III (Nakayama) | C6/36 cells | 8.1 log10 PFU/mL | Goose | Cotton pledgets | 26 °C | 14 | 5–50 | 10% | 40% | 0% | Vero cells | [84] | |
| UK, Liverpool | G-II (CNS138-11) | Vero cells | 106 PFU/mL | Heparinized human | Hemotek with collagen membrane | 18 °C | 21 | 18 | 100% | ND | 72% | Semi-quantitative qPCR | [89] | |
| Culex pipiens molestus (Forsskål, 1775) | Taiwan, Taipei | ND (SH) | C6/36 cells and suckling mice brains | 5.54 log10 PFU/mL | Defibrinated rabbit | Hanging drop method | 28–32 °C | 14 | 3–5 | ND | ND | 91% | Inoculation of brain tissue aspirates from recipient mice on to C6/36 cells | [109] |
| US, Oakland | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 7–20 | 1 | ND | ND | 22% # | Development of encephalitis in laboratory-reared mice | [96] | |
| Tashkent, Uzbekistan | ND (ROK-2.0028) | Vero cells | 104 PFU/mL | NA | Viremic chicken | 26 °C | 16–27 | 13–53 | 47–56% | 25–26% | 8% | Vero cells | [110] | |
| Culex pipiens pallens | Japan | G-III (JaGAr 01) | ND | ND | NA | Infected lizards | ND | ND | ND | ND | ND | Successful | Transmission from infected mosquitoes to uninfected lizards and from infected lizards to mice via mosquito | [111] |
| Japan, Sapporo | G-III (JANAr-5681) | C6/36 cells | 6.2 PFU/mL (blood) and 3.7 PFU/mL (chicken) | ND | Cotton pledgets or viremic chicken | 20 or 28 °C | 0–20 | 10 | 30% | ND | ND | BHK-21 cells and IFA | [98] | |
| Korea, Gyeonggi Province | ND (ROK-2.0028) | Vero cells | 105.2 PFU/mL | NA | Viremic chicken | 26 °C | 13–34 | 32 | 6% | 0% | ND | Vero cells | [112] | |
| Culex pipiens pipiens | France, Montpellier and Nice | G-III (RP-9) and G-V (XZ0934) | Chicken fibroblast-derived DF1 cells | 8 × 106 FFU/mL | Washed rabbit erythrocytes | Cotton pledgets | 26 °C | 7–13 | 5–20 | 70–92% | 26–80% ◊ | 12–41% | BHK-21 cells | [87] |
| UK, Caldbeck | G-III (SA14) | Vero cells | 1.8 × 106 PFU/mL | Defibrinated horse | Hemotek with parafilm membrane | 20 and 25 °C | 14 | 20–56 | 69–90% | 12–70% | 0–70% | RT-PCR and isolation in Vero cells | [15] | |
| US, Yakima | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 14–20 | 1–4 | ND | ND | 12% # | Development of encephalitis in laboratory-reared mice | [96] | |
|
Culex pseudovishnui | India | G-III (P20778) | NA | ND | Viremic chicks | NA | ND | 8 | ND | ND | 60% | 75% | Transmission to chickens | [113] |
| India | G-III (P20778) | NA | ND | ND | ND | ND | 1–10 | ND | ND | 49% | 51% | Antigen detection is mosquito heads resp. salivary glands | [114] | |
| Culex quinquefasciatus | Australia, Mareeba (wild) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit blood | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 27 | 56% | ND | 0% | Detection of virus in brain aspirates of recipient suckling mice | [93] |
| Australia, Gold coast | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 17/19 | 8–51 | 98% | 28% | 50% | Porcine stable-equine kidney cells | [93] | |
| New-Zealand, Wellington | G-III (Nakayama) | C6/36 cells | 8.1 log10 PFU/mL | Goose | Cotton pledgets | 24 °C | 14 | 6–36 | 17% | 0% | ND | Vero cells | [84] | |
| US, Rutgers | G-III (Nakayama) | C6/36 cells | 8.1 log10 PFU/mL | Goose | Cotton pledgets | 26 °C | 14 | 43–50 | 86% | 0% | 0% | Vero cells | [84] | |
| Brazil | G-V (Muar) | Vero cells | 4 log10 PFU/mL | Defibrinated horse | Hemotek with Parafilm membrane | 23 and 28 °C | 0–21 | 3–32 | 25–100% | 21–70% | 3–70% | Vero cells | [85] | |
| US | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 11–25 | 1–9 | ND | ND | 3% # | Development of encephalitis in laboratory-reared mice | [96] | |
| Culex sitiens | Australia, Coomera Islands | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine | 28 °C | 5–14 | 15–36 | 83–92% | 6–33% | 7–67% | Porcine stable-equine kidney cells | [93] |
| Culex tarsalis | US | G-II (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 6–10 | 1–12 | ND | ND | 1% # | Development of encephalitis in laboratory-reared mice | [96] |
|
Culex tritaeniorhynchus | Japan, Sapporo | G-III (JANAr-5681) | C6/36 cells | 6.2 PFU/mL (blood) and 3.7 PFU/mL (chicken) | ND | Cotton pledgets or viremic chicken | 20 or 28 °C | 0–20 | 15 | 100% | ND | 100% | BHK-21 cells and IFA | [98] |
| Japan, Narita | G-I (17CxIT-I4-D31), 3 (JaGAr 01) and 5 (Muar) | C6/36 cells | 8.9, 8.6, and 7.1 log10 FFU/mL | Defibrinated rabbit | Hemotek with pig intestine membrane | 27° C | 7–14 | 27–51 | 85–99% | 81–96% | 76–89% | RT-qPCR or Vero cells | [12] | |
| Korea, Gyeonggi Province | ND (ROK-2.0028) | Vero cells | 104.3 or 105.2 | NA | Viremic chicken | 26 °C | 13–34 | 10–18 | 100% | 80–93% | 50% | Vero cells | [112] | |
| Taiwan, Taipei | ND (SH) | C6/36 cells and suckling mice brains | 5.48 log10 PFU/mL | Defibrinated rabbit | Hanging drop method | 28–32 °C | 14 | 6–8 | ND | ND | 100% | Inoculation of brain tissue aspirates from recipient mice on to C6/36 cells | [109] | |
| Culex vishnui | India | G-III (P20778) | ND | ND | ND | Oral infection | ND | 1–10 | 100 | ND | 34% | 48% | Antigen detection in mosquito heads resp. salivary glands | [115] |
| Culiseta annulata (Schrank, 1776) | UK, Little Neston | G-II (CNS138-11) | Vero cells | 106 PFU/mL | Heparinized human | Hemotek with collagen membrane | 21 and 24 °C | 14–28 | 5–35 | 0–57% | ND | 0–30% | Semi-quantitative qPCR | [89] |
| Culiseta incidens (Thomson, 1869) | US | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 8–14 | 1–22 | ND | ND | 5% # | Development of encephalitis in laboratory-reared mice | [96] |
| Culiseta inornata (Williston, 1893) | US | G-III (Nakayama) | ND | ND | Defibrinated rabbit | Cotton pledgets | 27 °C | 10–20 | 2–12 | ND | ND | 4% # | Development of encephalitis in laboratory-reared mice | [96] |
| Opifex fuscus (Hutton, 1902) | New-Zealand, Wellington | G-III (Nakayama) | C6/36 cells | 108.1 | Goose | Cotton pledgets | 24 °C | 14 | 37–50 | 74% | 70% | 0% | Vero cells | [84] |
| Verrallina funerea | Australia, Cairns (wild s) | G-II (TS3306) | C6/36 and porcine stable-equine kidney cells | 104.5±0.1 CCID50/mL | Heparinized rabbit | Glass membrane feeder with pig intestine membrane | 28 °C | 14–15 | 36 | 11% | ND | 7% | Detection of virus in brain aspirates of recipient suckling mice | [93] |
| Mosquito Species | Potential Vectors |
Confirmed Vectors | References |
|---|---|---|---|
| Aedes albopictus | X | [19,20,88,95,116] | |
| Aedes detritus | X | [85] | |
| Aedes dorsalis | X | [96] | |
| Aedes japonicus | X | [12,97,98] | |
| Aedes kochi | X | [93] | |
| Aedes nigromaculis | X | [96] | |
| Aedes notoscriptus | X | [93] | |
| Aedes vexans | X | [19,20,99] | |
| Aedes vigilax | X | [25,26,93] | |
| Anopheles tessellatus | X | [19,100] | |
| Armigeres subalbatus | X | [19,20,28,32,34,101] | |
| Culex annulirostris | X | [39,93,99] | |
| Culex bitaeniorhynchus | X | [21,28,43,44,45,102,103,104] | |
| Culex fuscocephala | X | [19,22,27,31,46,48,49,50,105,106] | |
| Culex gelidus | X | [22,23,27,31,46,50,51,52,53,54,55,56,57,58,59,60,61,93,107,115] | |
| Culex pipiens | X | [15,43,44,62,63,64,84,87,89,96,108,109,110] | |
| Culex pipiens pallens | X | [38,98,111,112] | |
| Culex pseudovishnui | X | [37,65,66,67,68,113,114] | |
| Culex quinquefasciatus | X | [19,20,22,28,31,69,70,84,85,93,96] | |
| Culex sitiens | X | [20,21,24,26,51,70,71,72,73,93] | |
| Culex tarsalis | X | [32,96] | |
| Culex tritaeniorhynchus | X | [12,19,21,24,27,28,31,33,34,35,38,41,42,43,44,46,47,50,53,54,55,56,59,64,67,70,74,75,76,77,78,79,80,98,109,112] | |
| Culex vishnui | X | [21,37,46,47,53,67,79,80,114] | |
| Culiseta annulata | X | [89] | |
| Culiseta incidens | X | [96] | |
| Culiseta inornata | X | [96] | |
| Verrallina funerea | X | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Van den Eynde, C.; Sohier, C.; Matthijs, S.; De Regge, N. Japanese Encephalitis Virus Interaction with Mosquitoes: A Review of Vector Competence, Vector Capacity and Mosquito Immunity. Pathogens 2022, 11, 317. https://doi.org/10.3390/pathogens11030317
Van den Eynde C, Sohier C, Matthijs S, De Regge N. Japanese Encephalitis Virus Interaction with Mosquitoes: A Review of Vector Competence, Vector Capacity and Mosquito Immunity. Pathogens. 2022; 11(3):317. https://doi.org/10.3390/pathogens11030317
Chicago/Turabian StyleVan den Eynde, Claudia, Charlotte Sohier, Severine Matthijs, and Nick De Regge. 2022. "Japanese Encephalitis Virus Interaction with Mosquitoes: A Review of Vector Competence, Vector Capacity and Mosquito Immunity" Pathogens 11, no. 3: 317. https://doi.org/10.3390/pathogens11030317