
Emergence of IncHI2 Plasmid-Harboring blaNDM-5 from Porcine Escherichia coli Isolates in Guangdong, China
by
Zhenbao Ma
1,2, 
Zhenling Zeng
2,3,
Jiao Liu
2,3,
Chang Liu
2,3,
Yu Pan
2,3,
Yanan Zhang
4 and
Yafei Li
1,* 1
Institute of Quality Standard and Monitoring Technology for Agro-Products, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
2
Guangdong Provincial Key Laboratory of Veterinary Pharmaceutics Development and Safety Evaluation, South China Agricultural University, Guangzhou 510642, China
3
National Risk Assessment Laboratory for Antimicrobial Resistance of Animal Original Bacteria, South China Agricultural University, Guangzhou 510642, China
4
Institute of Animal Husbandry and Veterinary Medicine, Guizhou Academy of Agricultural Sciences, Guiyang 550005, China
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(8), 954; https://doi.org/10.3390/pathogens10080954
Submission received: 17 June 2021
/
Revised: 19 July 2021
/
Accepted: 23 July 2021
/
Published: 29 July 2021
Abstract
:Carbapenem resistance has posed potential harmful risks to human and animals. The objectives of this study were to understand the prevalence of blaNDM-5 in pigs and investigate the molecular characteristics of NDM-5-producing Escherichia coli isolates in Guangdong province in China. Carbapenem-resistant E. coli isolates were isolated from pigs and obtained using MacConkey plates containing 0.5 mg/L meropenem. Conjugation assay and antimicrobial susceptibility testing were conducted for the isolates and their transconjugants. Whole-genome sequence (WGS) was used to analyze the plasmid genetic feature. A total of five blaNDM-5-carrying E. coli isolates were obtained in the present investigations. They belonged to five ST types. The blaNDM-5 genes were found to be in IncX3 and IncHI2 plasmid. The IncX3 plasmid was 46,161 bp in size and identical to other reports. IncHI2 plasmid was 246,593 bp in size and similar to other IncHI2-ST3 plasmids. It consisted of a typical IncHI2 plasmid backbone region and a multiresistance region (MRR). The blaNDM-5 was closely associated with the IS3000-ISAba125-blaNDM-5-bleMBL-trpF-tat-IS26 unit. We first reported the blaNDM-5-carrying IncHI2 in E. coli isolates recovered from pigs and revealed the molecular characterization. Continued surveillance for the dissemination of blaNDM-5 among food-producing animals is required.
1. Introduction
Carbapenems are considered to be the most active and potent agents in the treatment of severe infections caused by multidrug-resistant (MDR) Gram-negative pathogens [1,2]. In the past few years, the emergence and spread of carbapenem-resistant Enterobacteriaceae (CRE) has led to a variety of infectious diseases with high mortality rates, which has posed a critical threat to clinical treatment and a public health concern [3,4]. New Delhi metallo-β-lactamase (NDM) is a type of metallo-β-lactamase (MBL) that hydrolyzes most carbapenems except for monobactams [5]. Since the blaNDM-5 gene was first reported, it has been the most prevalent carbapenem resistance gene in Enterobacteriaceae from humans and animals as demonstrated by several epidemiological investigations published in recent years [6,7].
Plasmids considered to be an important vehicle, play a vital role in the dissemination of the resistance genes. The blaNDM-5 gene is reported to be located on a variety of plasmid replicon types, such as IncFII-IncFIB [8], with IncX3 being the major carrier [9,10,11]. Recently, blaNDM-5 gene was found from the surrounding via IncX3 plasmid. These findings warn the wide-dissemination possibility of carbapenem resistance gene and emphasize the importance of monitoring the prevalence. Herein, the aim of our study was to investigate the prevalence of blaNDM-5-positive Escherichia coli strains from pigs in Guangdong province, China, and revealed the blaNDM-5-carrying plasmid in E. coli isolates recovered from pigs.
2. Results
In this study, five isolates positive for blaNDM-5 gene from one swine farm were obtained. The susceptibility test indicated that five isolates were phenotypically resistant to β-lactams and displayed patterns of MDR, such as ampicillin, ceftazidime, meropenem, apramycin, florfenicol, sulfamethoxazole/trimethoprim, but sensitive to gentamycin, amikacin, colistin, and tigecycline, as shown in Table 1. According to PFGE analysis, five isolates exhibited five different PFGE patterns, designated A to E (Figure 1). Considering the source of isolates, we presumed that the dissemination of the blaNDM-5 gene may be due to horizontally transfer.
Based on the whole-genome sequence analysis, the MLST of five isolates were divided into five distinct sequence types, namely ST10, ST48, ST155, ST2937, and ST4063. The isolates harbored numerous antimicrobial resistance genes such as blaTEM-1B, blaOXA-10, blaCTX-M-14, aadA2, cmlA1 and floR, as seen in Figure 1. These resistance genes were associated with antibiotic-resistant phenotype. The five blaNDM-5-positive isolates were successfully transferred the blaNDM-5-5 gene to E. coli J53 at frequencies of 0.15 × 10−6 to 5.98 × 10−6 transconjugant/recipient (Table 1). The transconjugants were resistant to ampicillin, cefoxitin, ceftazidime, meropenem, florfenicol, and sulfamethoxazole/trimethoprim (Table 1).
It is noteworthy that blaNDM-5 genes were in two replicon types of plasmids. After we analyzed the genomic features of these five strains by the next-generation sequencing, we found two types of plasmids in the blaNDM-5-positive strains. PCR-based replicon typing indicated that transconjugants GDB8P64J and GDB8P70J carried the IncHI2 plasmid replicon and exhibited the same MDR phenotype, while the remaining transconjugants carried the IncX3 plasmid replicon and only displayed resistance to β-lactams. A complete IncX3 plasmid carrying the blaNDM-5 gene was detected in three isolates (GDB8P65M, GDB8P75M and GDB8P77M) according to PCR mapping and the primers listed in Table 2. Sequence analysis indicated that the IncX3 plasmid pHNGD75-NDM (accession numbers MT296100) was 46,161 bp in size and was almost identical to the IncX3 plasmids found in Enterobacteriaceae such as pCNUH-14 (E. coli, MK986791) and pNDM5_IncX3 (K. pneumoniae, KU761328).
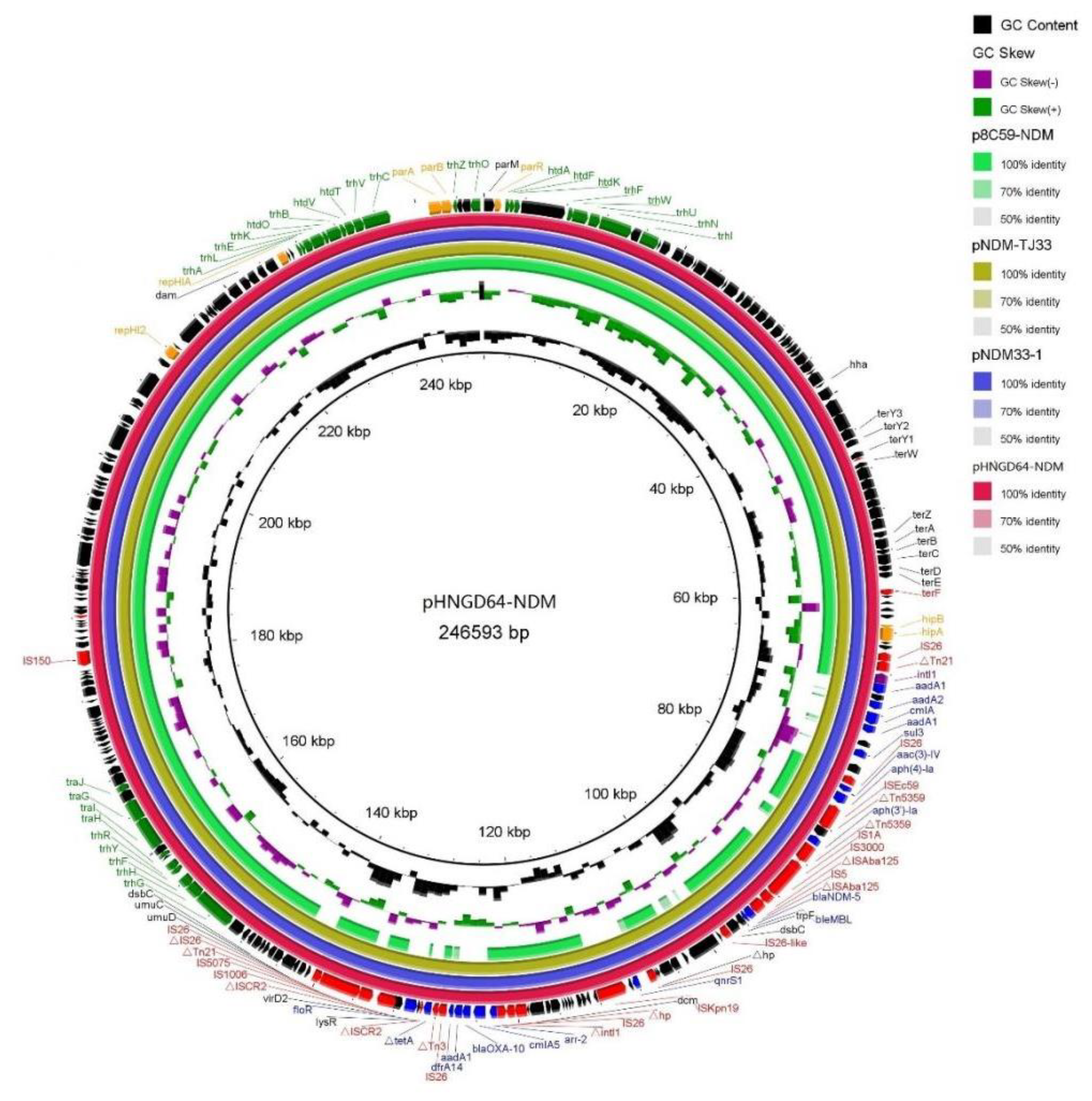
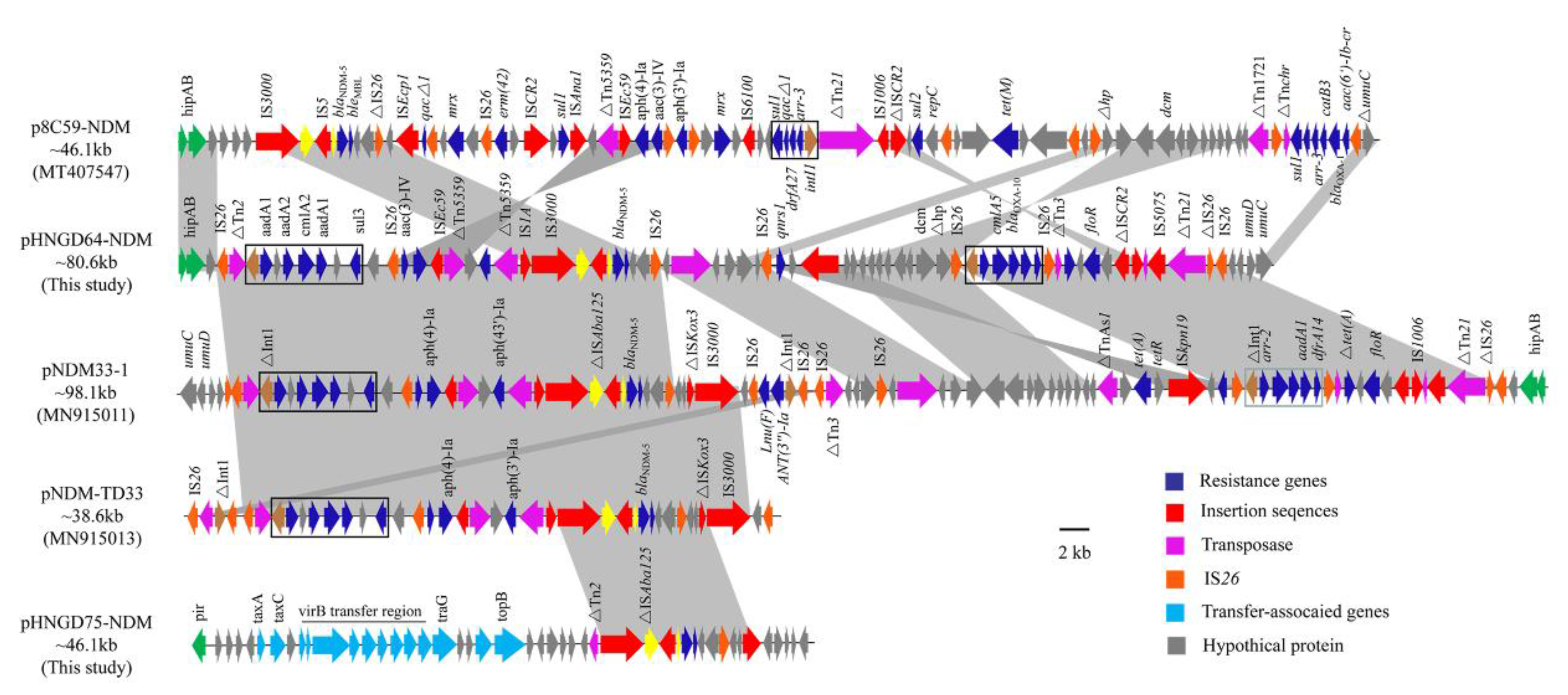
Sequence analysis also demonstrated that the blaNDM-5 gene was in the IncHI2 plasmid, named pHNGD64-NDM (accession numbers MW296099). Plasmid pHNGD64-NDM belonged to IncHI2-ST3. It was 246,593 bp in size and similar to other IncHI2-ST3 plasmids, it consisted of a typical IncHI2 plasmid backbone region and a multiresistance region (MRR). The backbone region of plasmid pHNGD64-NDM contained a typical IncHI2 plasmid backbone region associating with plasmid replication, conjugal transfer, and maintenance and stability, as well as a set of tellurite resistance determinants. Its similarity to those blaNDM-5-carrying IncHI2-ST3 plasmids such as plasmids p8C59-NDM (E. coli, chicken, MT407547), pNDM33-1 (E. coli, duck, MN915011) and pNDM-TJ33 (E. coli, duck, MN915010) (Figure 2). Furthermore, the MRR of pHNGD64-NDM contained several antibiotic resistance genes (e.g., addA1, cmlA5, aph(4)-Ia, blaOXA-10, blaNDM-5, arr-2, drfA14 and floR) and insertion sequences (e.g., IS26, Tn2, ISEc59, IS3000, IS5, Tn5359, ISKpn19, IS1006, IS5075, and Tn21). The MRR of pHNGD64-NDM was inserted into the downstream region of hipB-A in accordance with plasmids p8C59-NDM and pNDM33-1. However, it was distinct from IncHI2 plasmid p8C59-NDM and displayed high similarity to that of pNDM33-1, which differed in its loss of the segments containing the ~11.8-kb genetic fragment (IS3000-hp-IS26-lnu(F)-∆intI-IS26-IS26-∆Tn3) and a 5489-bp tetA module (∆TnAs1-tetA-tetR) (Figure 1). Moreover, a fragment with a size of 32,661-bp including a class I integron cassette and blaNDM-5 module in pHNGD64-NDM showed >99% homology with plasmids pNDM33-1 and pNDM-TD33, in addition, a fragment was observed with the size of 41,837-bp containing a floR module (∆ISCR2-floR-∆ISCR2), qnrS module (IS26-qnrS-ISKpn19) and an incomplete class I integron harboring the arr-2| cmlA5| blaOXA-10| aadA1| drfA14| cassette array, which was truncated by two copies of IS26 with the same direction, but the direction of the qnrS module in pHNGD64-NDM was the opposite (Figure 3).
3. Discussion
Plasmid-mediated horizontal transmission of drug-resistant genes has been recognized as an important pathway for the rapid transmission of drug-resistant genes in Enterobacteriaceae, of which IncX3, IncFII and IncHI2 plasmids play a key role in the transmission of carbapenem-resistant blaNDM-5 genes in Gram-negative bacteria [12]. Until now, blaNDM-5 gene has been reported to be prevalent in various members of Enterobacteriaceae. Recently, plasmid-encoded blaNDM-5 gene was found in Salmonella typhimurium of pork origin [13,14] and Klebsiella pneumonia [4,15] via IncX3 plasmids. IncX3 is a group of narrow-host-range plasmid, with the size of 46,161 bp, and an important transmission vector for blaNDM-5 gene, with a variety of blaNDM-5 subtypes, such as blaNDM-1, blaNDM-4, blaNDM-5, bla NDM-7 and blaNDM-21 [16,17]. IncX3 plasmid has been reported in many regions of the world, and has been widely reported in clinical human medicine, food animals and the environment [9]. In this study, three strains of blaNDM-5-positive E. coli were detected in IncX3 plasmid. Genomic sequence analysis showed that the IncX3 plasmid was highly similar to that reported in the Enterobacteriaceae such as E. coli and Klebsiella pneumoniae from human clinic, environment and food animals published in the database [18,19], suggesting that IncX3 plasmid has been widely prevalent.
In this study, for the first time, we detected and revealed the blaNDM-5-carrying IncHI2 in E. coli isolates recovered from pigs. Although the sample number is small and only two strains carry the IncHI2 plasmid, the presence of this plasmid should not be ignored. IncHI2 is a group of low-copy number, wide-host plasmid, which was widely distributed in Enterobacteriaceae. IncHI2 plasmid were described to carry a variety of drug-resistant genes, such as oqxAB, blaCTX-M, blaNDM, blaKPC, fosA3 and mcr-1 gene [13,20,21,22]. Reports on the blaNDM-5-carrying IncHI2 plasmid are scarce. Previous reports have also suggested that the IncHI2 plasmid was considered to be an important vehicle in the spread of carbapenem resistance genes such as blaNDM-1, blaNDM-4, blaNDM-9 and blaIMP-4 among Enterobacteriaceae from human and animals [13,23,24]. Since the discovery of mcr-1 gene in 2015 that threatens the effectiveness of polymyxins, the IncHI2 plasmid that carried mcr-1 has been found in Enterobacteria from a variety of sources, including humans [25], animals (cattle, pigs, chickens, ducks and turkeys) [26,27], food (chicken, beef, pork, milk and vegetables) [28,29] and the environment (farm soil and wastewater) [30] in several countries around the world. In this study, IncHI2 plasmid was co-transmitted with many drug resistance genes such as floR, cmlA, sul1, acc(3)-IV, aph(4) -Ia and arr-2, which undoubtedly exacerbated the resistance crisis of animal borne bacteria. In addition, after carefully looking up relevant information in the NCBI database, IncHI2 plasmids carrying blaNDM-5 gene were found in E. coli from chicken, ducks, and pigs in Guangdong province, but there were few reports in other regions. The present findings suggest that IncHI2 plasmid has become an important transmission vector for blaNDM-5 gene in food animals in Guangdong province. This type of plasmid may have spread in different areas of Guangdong province, and adaptive evolution occurred in the variable region of the plasmid during transmission. In this study, the sequence of porcine E. coli plasmid pNGD64-NDM showed high homotypes with other IncHI2 plasmid such as pNDM33-1 obtained in the duck farm that uploaded by our teaching and research department. We speculate that this phenomenon may be related to the large number of copies of insertion sequence IS26 in the variable region, which lead to the variation of the variable region and then the inversion, deletion, and insertion of some sequences. It is noteworthy that the 9525-bp blaNDM-5 module was found in various replicon plasmids such as IncHI2 plasmids p8C59-NDM and pNDM33-1, IncI-γ plasmid pNDM-TD33 and IncX3 plasmid pHNGD75-NDM, which suggests that the dissemination of blaNDM-5 was closely associated with the IS3000-ISAba125-blaNDM-5-bleMBL-trpF-tat-IS26 unit.
4. Materials and Methods
4.1. Bacterial Collection, Species Identification, and Molecular Detection of Carbapenemase Genes
A total of 357 fecal samples were collected from three swine farms in two cities (Maoming and Qingyuan) in Guangdong province from May to November in 2018, with the sample of 180, 81, and 96 in each farm, respectively. Carbapenem-resistant E. coli isolates were screened using MacConkey plates containing 0.5 mg/L meropenem and only one red colony was selected from each fecal sample. Meropenem-resistant isolates from these swine farms were identified as E. coli by MALDI-TOF-MS. PCR and sanger sequencing were further done to confirm whether the resistant strains harbored the blaNDM-5 gene. The primer for the blaNDM-5 was conducted to confirm the presence of blaNDM-5 gene as previous report [31].
4.2. Conjugation and Antimicrobial Susceptibility Testing
To determine the transferability of blaNDM-5-bearing isolates, the transfer frequency for each strain was determined using the broth mating method, with sodium-azide-resistant E. coli J53 used as the recipient strain. The donor strains and the recipient strain were mixed at the ratio of 1:4 in Luria-Bertani (LB) broth, and then incubated at 37 °C for 4 h, the transconjugants were selected on MacConkey agar plates supplemented with 0.5 mg/L meropenem and 200 mg/L sodium azide and then confirmed by PCR method.
The minimum inhibitory concentrations (MICs) of 15 antimicrobial drugs against the blaNDM-5 positive isolates and their transconjugants were determined using the agar dilution method or broth microdilution method (colistin) and values were then interpreted according to the document of CLSI-2020. Breakpoints of neomycin (>8 mg/L) and florfenicol (>16 mg/L) were interpreted according to EUCAST (http://mic.eucast.org/Eucast2/, accessed on 24 September 2020). The following antimicrobial agents were used in the present research: ampicillin, cefoxitin, ceftazidime, meropenem, gentamycin, amikacin, neomycin, apramycin, doxycycline, tigecycline, florfenicol, colistin, ciprofloxacin, enrofloxacin and sulfamethoxazole/trimethoprim. E. coli ATCC25922 served as the quality control strain.
4.3. Molecular Analysis of blaNDM-5-Positive Isolates
The genetic typing of blaNDM-5-positive E. coli isolates was digested with restriction endonuclease Xbal and conducted by pulsed-field gel electrophoresis (PFGE) according to our previous study [32]. The band patterns were analyzed with BioNumerics software version 5.10 (Applied Math s, Austin, TX, USA). Cluster analysis of fingerprinting similarity was conducted on software MasterScripts v4.0. The bacterial DNA of blaNDM-5-positive E. coli isolates were extracted by the DNA Extraction Kit, and then were sent to laboratory by next-generation sequencing. The multi-locus sequence type (MLST) of E. coli was performed by Escherichia typing database (https://pubmlst.org/bigsdb?db=pubmlst_escherichia_seqdef, accessed on 10 September 2020).
4.4. Whole-Genome Sequence and Plasmid Analysis
The blaNDM-5-carrying plasmid pHNGD64-NDM was sequenced using PacBio platform and assembled using HGAP version 4.0. to analyze the genetic feature. BRIG software was used to comparative analysis with other plasmid sequences published in NCBI (Genbank accession number MT407547, MN915011, MN915010). The complete genome of plasmid was annotated and analysis using RAST (https://rast.nmpdr.org/rast.cgi, accessed on 21 September 2020), ISfinder (https://www-is.biotoul.fr/, accessed on 12 September 2020), Resfinder (https://cge.cbs.dtu.dk//services/ResFinder/, accessed on 12 September 2020) and Vector NTI program (Invitrogen).
5. Conclusions
To the best of our knowledge, this study was the first to identify the blaNDM-5-carrying IncHI2 recovered from porcine E. coli isolates. Although carbapenems have not been approved for use in food-producing animals, multiple resistance genes were observed in the IncHI2 plasmid pHNGD64-NDM in the present research, which suggests that the spread and evolution of the blaNDM-5 gene may be selected and accelerated under the selective pressure of other antibacterial drugs. Thus, the dissemination of blaNDM-5 among food-producing animals requires continued surveillance.
Author Contributions
Z.M., Z.Z. and Y.L. designed the study. Z.M. and Y.L. wrote the manuscript. J.L., C.L. and Y.P. carried out the experiment. Y.Z. helped to analyze the data. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the National Key R&D Program of China (Grant Number 2018YFD0500300) and the National Natural Science Foundation of China (Grant Number 31902323).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Breilh, D.; Texier-Maugein, J.; Allaouchiche, B.; Saux, M.C.; Boselli, E. Carbapenems. J. Chemother. 2013, 25, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guducuoglu, H.; Gursoy, N.C.; Yakupogullari, Y.; Parlak, M.; Karasin, G.; Sunnetcioglu, M.; Otlu, B. Hospital outbreak of a colistin-resistant, NDM-1- and OXA-48-producing Klebsiella Pneumoniae: High mortality from pandrug resistance. Microb. Drug Resist. 2018, 24, 966–972. [Google Scholar] [CrossRef]
- Doi, Y. Treatment options for carbapenem-resistant gram-negative bacterial infections. Clin. Infect. Dis. 2019, 69, S565–S575. [Google Scholar] [CrossRef] [Green Version]
- Cao, T.; Liu, Y.; Li, Y.; Wang, Y.; Shen, Z.; Shao, B.; Walsh, T.R.; Shen, J.; Wang, S. A public health concern: Emergence of carbapenem-resistant Klebsiella pneumoniae in a public transportation environment. J. Antimicrob. Chemother. 2020, 75, 2769–2772. [Google Scholar] [CrossRef]
- Nordmann, P.; Poirel, L.; Walsh, T.R.; Livermore, D.M. The emerging NDM carbapenemases. Trends Microbiol. 2011, 19, 588–595. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Kong, Z.; Qian, H.; Jiang, F.; Kang, H.; Gu, B.; Ma, P. High prevalence of blaNDM variants among carbapenem-resistant Escherichia coli in northern Jiangsu Province, China. Front. Microbiol. 2018, 9, 2704. [Google Scholar] [CrossRef] [Green Version]
- Xiang, R.; Zhang, A.Y.; Ye, X.L.; Kang, Z.Z.; Lei, C.W.; Wang, H.N. Various sequence types of Enterobacteriaceae isolated from commercial chicken farms in China and carrying the blaNDM-5 gene. Antimicrob. Agents Chemother. 2018, 62, e0077918. [Google Scholar] [CrossRef] [Green Version]
- Takayama, Y.; Sekizuka, T.; Matsui, H.; Adachi, Y.; Eda, R.; Nihonyanagi, S.; Wada, T.; Matsui, M.; Suzuki, S.; Takaso, M.; et al. Characterization of the IncFII-IncFIB (pB171) plasmid carrying blaNDM-5 in Escherichia coli ST405 clinical isolate in Japan. Infect. Drug Resist. 2020, 13, 561–566. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Feng, Y.; Tang, G.; Qiao, F.; McNally, A.; Zong, Z. NDM Metallo-β-Lactamases and their bacterial producers in health care settings. Clin. Microbiol. Rev. 2019, 32, e00115-18. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Berglund, B.; Zou, H.; Zhou, Z.; Xia, H.; Zhao, L.; Nilsson, L.E.; Li, X. Dissemination of blaNDM-5 via IncX3 plasmids in carbapenem-resistant Enterobacteriaceae among humans and in the environment in an intensive vegetable vultivation area in eastern China. Environ. Pollut. 2021, 273, 116370. [Google Scholar] [CrossRef]
- Ma, T.; Fu, J.; Xie, N.; Ma, S.; Lei, L.; Zhai, W.; Shen, Y.; Sun, C.; Wang, S.; Shen, Z.; et al. Fitness cost of blaNDM-5-carrying p3R-IncX3 plasmids in wild-type NDM-free Enterobacteriaceae. Microorganisms 2020, 8, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Liu, Y.; Zhang, Q.; Jin, L.; Wang, Q.; Zhang, Y.; Wang, X.; Hu, M.; Li, L.; Qi, J.; et al. The prevalence of colistin resistance in Escherichia coli and Klebsiella pneumoniae isolated from food animals in China: Coexistence of mcr-1 and blaNDM with low fitness cost. Int. J. Antimicrob. Agents 2018, 51, 739–744. [Google Scholar] [CrossRef]
- Gao, Y.; Wen, J.; Wang, S.; Xu, X.; Zhan, Z.; Chen, Z.; Bai, J.; Qu, X.; Zhang, H.; Zhang, J.; et al. Plasmid-encoded blaNDM-5 gene that confers high-level carbapenem resistance in Salmonella Typhimurium of pork origin. Infect. Drug Resist. 2020, 13, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; He, J.; Li, Q.; Tang, Y.; Wang, J.; Pan, Z.; Chen, X.; Jiao, X. First detection of blaNDM-5-positive Salmonella Enterica serovar typhimurium isolated from retail pork in China. Microb. Drug Resist. 2020, 26, 434–437. [Google Scholar] [CrossRef]
- Sherchan, J.B.; Tada, T.; Shrestha, S.; Uchida, H.; Hishinuma, T.; Morioka, S.; Shahi, R.K.; Bhandari, S.; Twi, R.T.; Kirikae, T.; et al. Emergence of clinical isolates of highly carbapenem-resistant Klebsiella pneumoniae co-harboring blaNDM-5 and blaOXA-181 or -232 in Nepal. Int. J. Infect. Dis. 2020, 92, 247–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mouftah, S.F.; Pál, T.; Darwish, D.; Ghazawi, A.; Villa, L.; Carattoli, A.; Sonnevend, Á. Epidemic IncX3 plasmids spreading carbapenemase genes in the United Arab Emirates and Worldwide. Infect Drug Resist. 2019, 12, 1729–1742. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Feng, Y.; McNally, A.; Zong, Z. blaNDM-21, a New Variant of blaNDM in an Escherichia coli clinical isolate carrying blaCTX-M-55 and rmtb. J. Antimicrob. Chemother. 2018, 73, 2336–2339. [Google Scholar] [CrossRef]
- Liu, B.; Shui, L.; Zhou, K.; Jiang, Y.; Li, X.; Guan, J.; Li, Q.; Zhuo, C. Impact of plasmid-encoded H-NS-like protein on blaNDM-1-bearing IncX3 plasmid in Escherichia coli. J. Infect. Dis. 2020, 221, s229–s236. [Google Scholar] [CrossRef]
- Zhu, W.; Wang, X.; Qin, J.; Liang, W.; Shen, Z. Dissemination and stability of the blaNDM-5-carrying IncX3-Type plasmid among multiclonal Klebsiella pneumoniae isolates. mSphere 2020, 6, e00917-20. [Google Scholar] [CrossRef]
- Pérez-Vázquez, M.; Sola Campoy, P.J.; Ortega, A.; Bautista, V.; Monzón, S.; Ruiz-Carrascoso, G.; Mingorance, J.; González-Barberá, E.M.; Gimeno, C.; Aracil, B.; et al. Emergence of NDM-producing Klebsiella pneumoniae and Escherichia coli in Spain: Phylogeny, resistome, virulence and plasmids encoding blaNDM-like genes as determined by WGS. J. Antimicrob Chemother. 2019, 74, 3489–3496. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Vazquez, M.; Oteo-Iglesias, J.; Sola-Campoy, P.J.; Carrizo-Manzoni, H.; Bautista, V.; Lara, N.; Aracil, B.; Alhambra, A.; Martínez-Martínez, L.; Campos, J. Characterization of carbapenemase-producing Klebsiella oxytoca in Spain, 2016–2017. Antimicrob. Agents Chemother. 2019, 63, e02529-18. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Huang, X.Y.; Xia, Y.B.; Guo, Z.W.; Ma, Z.B.; Yi, M.Y.; Lv, L.C.; Lu, P.L.; Yan, J.C.; Huang, J.W.; et al. Clonal spread of Escherichia coli ST93 carrying mcr-1-harboring IncN1-IncHI2/ST3 plasmid among companion Animals, China. Front. Microbiol. 2018, 9, 02989. [Google Scholar] [CrossRef]
- Roberts, L.W.; Catchpoole, E.; Jennison, A.V.; Bergh, H.; Hume, A.; Heney, C.; George, N.; Paterson, D.L.; Schembri, M.A.; Beatson, S.A.; et al. Genomic analysis of carbapenemase-producing Enterobacteriaceae in Queensland reveals widespread transmission of blaIMP-4 on an IncHI2 plasmid. Microb. Genom. 2020, 6, e000321. [Google Scholar]
- Liu, B.T.; Song, F.J.; Zou, M.; Zhang, Q.D.; Shan, H. High incidence of Escherichia coli strains coharboring mcr-1 and blaNDM from chickens. Antimicrob. Agents Chemother. 2017, 61, e02347-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Xu, H.; Tang, Y.; Li, Q.; Jiao, X. A multidrug-resistant monophasic Salmonella Typhimurium co-harboring mcr-1, fosA3, blaCTX-M-14 in a transferable IncHI2 plasmid from a healthy catering worker in China. Infect. Drug Resist. 2020, 13, 3569–3574. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Sun, R.Y.; Song, J.Q.; Fang, L.X.; Zhang, R.M.; Lian, X.L.; Liao, X.P.; Liu, Y.H.; Lin, J.; Sun, J. Within-host heterogeneity and flexibility of mcr-1 transmission in chicken gut. Int. J. Antimicrob. Agents 2020, 55, 105806. [Google Scholar] [CrossRef] [PubMed]
- Zając, M.; Sztromwasser, P.; Bortolaia, V.; Leekitcharoenphon, P.; Cavaco, L.M.; Ziȩtek-Barszcz, A.; Hendriksen, R.S.; Wasyl, D. Occurrence and characterization of mcr-1-positive Escherichia coli isolated from food-producing animals in Poland, 2011–2016. Front. Microbiol. 2019, 10, 1753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Yan, Y.; Chen, J.; Sun, R.; Wang, Y.; Wang, T.; Feng, Z.; Peng, K.; Wang, J.; Chen, S.; et al. Genomic characterization of conjugative plasmids carrying the mcr-1 gene in foodborne and clinical strains of Salmonella and Escherichia coli. Food Control. 2021, 125, 108032. [Google Scholar] [CrossRef]
- Hammad, A.M.; Hoffmann, M.; Gonzalez-Escalona, N.; Abbas, N.H.; Yao, K.; Koenig, S.; Allué-Guardia, A.; Eppinger, M. Genomic features of colistin resistant Escherichia coli ST69 strain harboring mcr-1 on IncHI2 plasmid from raw milk cheese in Egypt. Infect. Genet. Evol. 2019, 73, 126–131. [Google Scholar] [CrossRef]
- Al-Tawfiq, J.A.; Laxminarayan, R.; Mendelson, M. How should we respond to the emergence of plasmid-mediated colistin resistance in humans and animals? Int. J. Infect Dis. 2017, 54, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Campos, J.C.; Silva, M.J.F.; Santos, P.R.V.; Barros, E.M.; Pereira, M.O.; Seco, B.M.S.; Magagnin, C.M.; Leiroz, L.K.; Oliveira, T.G.M.; Faria-Júnior, C.; et al. Characterization of Tn3000, a transposon responsible for blaNDM-1 dissemination among Enterobacteriaceae in Brazil, Nepal, Morocco, and India. Antimicrob. Agents Chemother. 2015, 59, 7387–7395. [Google Scholar] [CrossRef] [Green Version]
- Ma, Z.; Liu, J.; Chen, L.; Liu, X.; Xiong, W.; Liu, J.H.; Zeng, Z. Rapid increase in the IS26-mediated cfr gene in E. coli isolates with IncP and IncX4 plasmids and Co-existing cfr and mcr-1 genes in a swine farm. Pathogens 2021, 10, 33. [Google Scholar] [CrossRef]
Figure 1.
PFGE, antibiotic resistance genes of five porcine blaNDM-5-producing E. coli isolates.

Figure 2.
Ring comparison of IncHI2 plasmid pHNGD64-NDM using BRIG. Arrows indicate the positions and directions of gene transcription. ∆Indicates a truncated gene. Blue, red, green and black arrows indicate resistance genes, insertion sequences and transposes, transfer-associated protein, and hypothetical protein.
Figure 2.
Ring comparison of IncHI2 plasmid pHNGD64-NDM using BRIG. Arrows indicate the positions and directions of gene transcription. ∆Indicates a truncated gene. Blue, red, green and black arrows indicate resistance genes, insertion sequences and transposes, transfer-associated protein, and hypothetical protein.

Figure 3.
Liner comparison of the MRR region of plasmid pHNGD64-NDM. Arrows indicate the positions of the genes and the direction. Regions with >99% homology are shaded in gray. ∆indicates a truncated gene or mobile element.
Figure 3.
Liner comparison of the MRR region of plasmid pHNGD64-NDM. Arrows indicate the positions of the genes and the direction. Regions with >99% homology are shaded in gray. ∆indicates a truncated gene or mobile element.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Characterization of blaNDM-5-producing E. coli and their transconjugants.
| Strain | GDB8P64 | GDB8P64J | GDB8P65 | GDB8P65J | GDB8P70 | GDB8P70J | GDB8P75 | GDB8P75J | GDB8P77 | GDB8P77J | E. coli J53 | E. coli 25922 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MLST a | ST4063 | - | ST10 | - | ST2937 | - | ST48 | - | ST155 | - | - | - |
| Plasmid type b | IncHI2 IncFIB IncQ1 p0111 | IncHI2 | IncX3 IncY | IncX3 | IncHI2 IncFIB IncFII IncY | IncHI2 | IncFIA IncR IncFII IncX3 | IncX3 | IncFIB IncN IncX3 p0111 | IncX3 | - | - |
| Transfer frequencies c | 5.70 × 10−6 | - | 0.15 × 10−6 | - | 5.98 × 10−6 | - | 2.32 × 10−6 | - | 0.68 × 10−6 | - | - | - |
| MIC d | ||||||||||||
| AMP | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 | >128 | 4 | 4 |
| CTX | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | 0.06 | 0.03 |
| CAZ | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | >64 | 0.06 | 0.06 |
| MEM | 8 | 2 | 16 | 4 | 8 | 2 | 16 | 4 | 16 | 4 | 0.016 | 0.016 |
| GEN | 4 | 0.5 | 0.5 | 0.5 | 4 | 0.5 | 0.5 | 0.25 | 0.5 | 0.25 | 0.25 | 0.25 |
| AMI | 1 | 0.5 | 4 | 0.25 | 2 | 0.5 | 2 | 0.5 | 2 | 0.5 | 0.5 | 0.5 |
| NEO | 64 | 0.5 | 1 | 0.5 | 64 | 0.5 | 1 | 0.5 | 32 | 1 | 0.5 | 1 |
| APR | >128 | 1 | 4 | 1 | >128 | 1 | 4 | 1 | 4 | 1 | 1 | 1 |
| DOX | 32 | 0.25 | 64 | 0.5 | 32 | 0.5 | 64 | 0.5 | 32 | 0.5 | 0.5 | 1 |
| TIG | 1 | 0.06 | 1 | 0.06 | 1 | 0.06 | 1 | 0.06 | 0.5 | 0.06 | 0.06 | 0.06 |
| FLR | 128 | 64 | >128 | 2 | >128 | 128 | >128 | 2 | >128 | 4 | 2 | 2 |
| CL | 0.25 | 0.125 | 0.25 | 0.125 | 0.25 | 0.125 | 0.25 | 0.125 | 0.25 | 0.125 | 0.125 | 0.125 |
| ENR | 32 | 0.5 | 1 | 0.016 | 16 | 0.5 | 2 | 0.016 | 16 | 0.016 | 0.016 | 0.008 |
| CIP | 8 | 0.25 | 0.5 | 0.008 | 8 | 0.25 | 2 | 0.008 | 8 | 0.004 | 0.008 | 0.008 |
| SXT | >64/1216 | 4/76 | >64/1216 | <0.25/4.75 | >64/1216 | 8/152 | >64/1216 | <0.25 | >64/1216 | <0.25/4.75 | <0.25/4.75 | <0.25/4.75 |
Note: a: letter “J” represents transconjugants. b: The multi-locus sequence type (MLST) of E. coli was performed by Escherichia typing database (https://pubmlst.org/bigsdb?db=pubmlst_escherichia_seqdef, accessed on 10 September 2020). c: Plasmid type of Enterobacteriaceae was determined using CGE online website (https://cge.cbs.dtu.dk/services/PlasmidFinder/, accessed on 12 September 2020). d: Transfer frequencies were calculated as the ratio of transconjugants over recipient cells, value was average mumble at least three times. Abbr. AMP: ampicillin, CTX: cefoxitin, CAZ: ceftazidime, MEM: meropenem, GEN: gentamycin, AMI: amikacin, NEO: neomycin; APR: apramycin, DOX: doxycycline, TIG: tigecycline, FLR: florfenicol, CL: colistin, CIP: ciprofloxacin, ENR: enrofloxacin and STX: sulfamethoxazole/trimethoprim.
Table 2.
Primer sequences of PCR mapping for blaNDM gene.
| Name | Prime Sequence (5′ to 3′) | Target Fragment | Reference |
|---|---|---|---|
| umuC-tat | F:GCGTAGCGTTTCCATAGCGG | 1912 bp | This study |
| R:GTTGACGGGTCTTTGGTGCT | |||
| ∆Tn2-hp | F:TGAAATGGCATGGGAATGAG | 1294 bp | This study |
| R:TTTCTGCGACAGTGATAGCG | |||
| R:GCTTTTGAAACTGTCGCACCT |
Note: F: forward primer; R: reverse primer.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ma, Z.; Zeng, Z.; Liu, J.; Liu, C.; Pan, Y.; Zhang, Y.; Li, Y. Emergence of IncHI2 Plasmid-Harboring blaNDM-5 from Porcine Escherichia coli Isolates in Guangdong, China. Pathogens 2021, 10, 954. https://doi.org/10.3390/pathogens10080954
AMA Style
Ma Z, Zeng Z, Liu J, Liu C, Pan Y, Zhang Y, Li Y. Emergence of IncHI2 Plasmid-Harboring blaNDM-5 from Porcine Escherichia coli Isolates in Guangdong, China. Pathogens. 2021; 10(8):954. https://doi.org/10.3390/pathogens10080954
Chicago/Turabian StyleMa, Zhenbao, Zhenling Zeng, Jiao Liu, Chang Liu, Yu Pan, Yanan Zhang, and Yafei Li. 2021. "Emergence of IncHI2 Plasmid-Harboring blaNDM-5 from Porcine Escherichia coli Isolates in Guangdong, China" Pathogens 10, no. 8: 954. https://doi.org/10.3390/pathogens10080954
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.