
High Prevalence and New Genotype of Coxiella burnetii in Ticks Infesting Camels in Somalia

 ,
,  ,
, 
Abstract
:1. Introduction
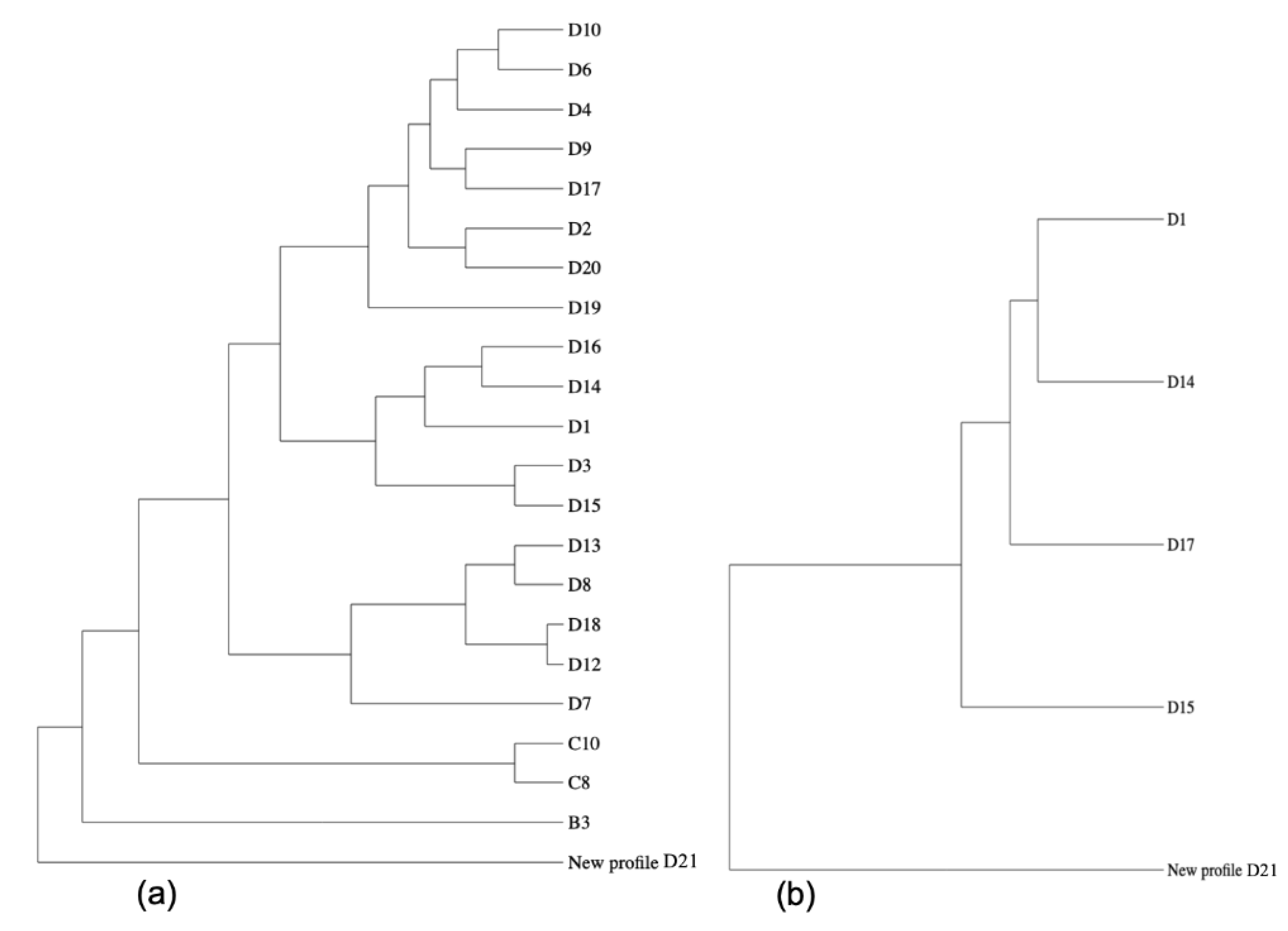
2. Results
3. Discussion
4. Materials and Methods
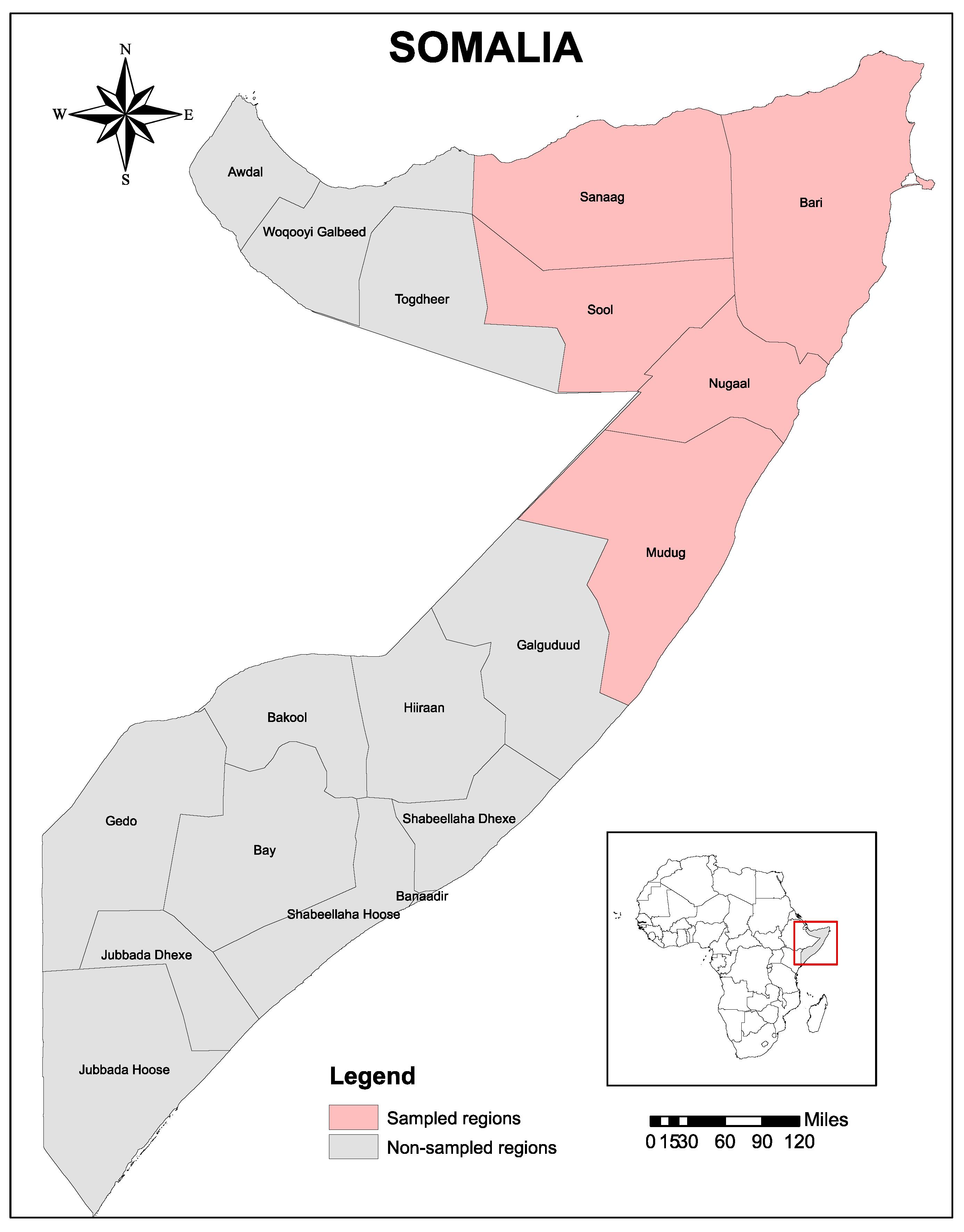
4.1. Study Area
4.2. Study Design and Tick Samples Collection
4.3. Laboratory Procedures
4.3.1. Tick Genera and Species Identification
4.3.2. Detection of Coxiella burnetii
4.3.3. Molecular Characterization of Coxiella burnetii
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maurin, M.; Raoult, D. Q Fever. Clin. Microbiol. Rev. 1999, 12, 518–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazar, J. Coxiella burnetii Infection. Ann. N. Y. Acad. Sci. 2005, 1063, 105–114. [Google Scholar] [CrossRef]
- Angelakis, E.; Raoult, D. Q Fever. Vet. Microbiol. 2010, 140, 297–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, C.A.; Osman, I.O.; Million, M.; Raoult, D. Coxiella burnetii in Dromedary Camels (Camelus Dromedarius): A Possible Threat for Humans and Livestock in North Africa and the Near and Middle East? Front. Vet. Sci. 2020, 7, 558481. [Google Scholar] [CrossRef]
- Duron, O.; Sidi-Boumedine, K.; Rousset, E.; Moutailler, S.; Jourdain, E. The Importance of Ticks in Q Fever Transmission: What Has (and Has Not) Been Demonstrated? Trends Parasitol. 2015, 31, 536–552. [Google Scholar] [CrossRef] [PubMed]
- Klemmer, J.; Njeru, J.; Emam, A.; El-Sayed, A.; Moawad, A.A.; Henning, K.; Elbeskawy, M.A.; Sauter-Louis, C.; Straubinger, R.K.; Neubauer, H.; et al. Q Fever in Egypt: Epidemiological Survey of Coxiella burnetii Specific Antibodies in Cattle, Buffaloes, Sheep, Goats and Camels. PLoS ONE 2018, 13, e0192188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahu, R.; Rawool, D.B.; Vinod, V.K.; Malik, S.V.S.; Barbuddhe, S.B. Current Approaches for the Detection of Coxiella burnetii Infection in Humans and Animals. J. Microbiol. Methods 2020, 179, 106087. [Google Scholar] [CrossRef]
- Gürtler, L.; Bauerfeind, U.; Blümel, J.; Burger, R.; Drosten, C.; Gröner, A.; Heiden, M.; Hildebrandt, M.; Jansen, B.; Offergeld, R.; et al. Coxiella Burnetii—Pathogenic Agent of Q (Query) Fever. Transfus. Med. Hemother. 2014, 41, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Gale, P.; Kelly, L.; Mearns, R.; Duggan, J.; Snary, E.L. Q Fever through Consumption of Unpasteurised Milk and Milk Products—A Risk Profile and Exposure Assessment. J. Appl. Microbiol. 2015, 118, 1083–1095. [Google Scholar] [CrossRef] [Green Version]
- Hogema, B.M.; Slot, E.; Molier, M.; Schneeberger, P.M.; Hermans, M.H.; van Hannen, E.J.; van der Hoek, W.; Cuijpers, H.T.; Zaaijer, H.L. Coxiella burnetii Infection among Blood Donors during the 2009 Q-Fever Outbreak in the Netherlands. Transfusion 2012, 52, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Million, M.; Raoult, D. Recent Advances in the Study of Q Fever Epidemiology, Diagnosis and Management. J. Infect. 2015, 71, S2–S9. [Google Scholar] [CrossRef] [PubMed]
- Wegdam-Blans, M.C.A.; Vainas, T.; van Sambeek, M.R.; Cuypers, P.W.; Tjhie, H.T.J.; van Straten, A.H.M.; Teijink, J.A. Vascular Complications of Q-Fever Infections. Eur. J. Vasc. Endovasc. Surg. 2011, 42, 384–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broos, P.P.H.L.; Hagenaars, J.C.J.P.; Kampschreur, L.M.; Wever, P.C.; Bleeker-Rovers, C.P.; Koning, O.H.J.; Teijink, J.A.W.; Wegdam-Blans, M.C.A. Vascular Complications and Surgical Interventions after World’s Largest Q Fever Outbreak. J. Vasc. Surg. 2015, 62, 1273–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wegdam-Blans, M.C.A.; Kampschreur, L.M.; Delsing, C.E.; Bleeker-Rovers, C.P.; Sprong, T.; van Kasteren, M.E.E.; Notermans, D.W.; Renders, N.H.M.; Bijlmer, H.A.; Lestrade, P.J.; et al. Chronic Q Fever: Review of the Literature and a Proposal of New Diagnostic Criteria. J. Infect. 2012, 64, 247–259. [Google Scholar] [CrossRef]
- Abdullah, H.H.A.M.; Hussein, H.A.; El-Razik, K.A.A.; Barakat, A.M.A.; Soliman, Y.A. Q Fever: A Neglected Disease of Camels in Giza and Cairo Provinces, Egypt. Vet. World 2019, 12, 1945–1950. [Google Scholar] [CrossRef] [Green Version]
- Schmatz, H.D.; Krauss, H.; Viertel, P.; Ismail, A.S.; Hussein, A.A. Seroepidemiological investigations in domestic ruminants from Egypt, Somalia and Jordan for the demonstration of complement fixing antibodies against Rickettsia and Chlamydia (author’s transl). Acta Trop. 1978, 35, 101–111. [Google Scholar] [PubMed]
- Gray, G.C.; Rodier, G.R.; Matras-Maslin, V.C.; Honein, M.A.; Ismail, E.A.; Botros, B.A.M.; Soliman, A.K.; Merrell, B.R.; Wang, S.P.; Grayston, J.T. Serologic Evidence of Respiratory and Rickettsial Infections among Somali Refugees. Am. J. Trop. Med. Hyg. 1995, 52, 349–353. [Google Scholar] [CrossRef]
- Botros, B.A.; Soliman, A.K.; Salib, A.W.; Olson, J.; Hibbs, R.G.; Williams, J.C.; Darwish, M.; el Tigani, A.; Watts, D.M. Coxiella burnetii Antibody Prevalences among Human Populations in North-East Africa Determined by Enzyme Immunoassay. J. Trop. Med. Hyg. 1995, 98, 173–178. [Google Scholar]
- Knap, N.; Žele, D.; Glinšek Biškup, U.; Avšič-Županc, T.; Vengušt, G. The Prevalence of Coxiella burnetii in Ticks and Animals in Slovenia. BMC Vet. Res. 2019, 15, 368. [Google Scholar] [CrossRef] [Green Version]
- Roest, H.I.J.; Bossers, A.; van Zijderveld, F.G.; Rebel, J.M.L. Clinical Microbiology of Coxiella Burnetii and Relevant Aspects for the Diagnosis and Control of the Zoonotic Disease Q Fever. Vet. Q. 2013, 33, 148–160. [Google Scholar] [CrossRef] [PubMed]
- van Roeden, S.E.; van Houwelingen, F.; Donkers, C.M.J.; Hogewoning, S.J.; de Lange, M.M.A.; van der Hoek, W.; Kampschreur, L.M.; Bonten, M.J.M.; Hoepelman, A.I.M.; Bleeker-Rovers, C.P.; et al. Exposure to Coxiella burnetii and Risk of Non-Hodgkin Lymphoma: A Retrospective Population-Based Analysis in the Netherlands. Lancet Haematol. 2018, 5, e211–e219. [Google Scholar] [CrossRef]
- Salifu, S.P.; Bukari, A.-R.A.; Frangoulidis, D.; Wheelhouse, N. Current Perspectives on the Transmission of Q Fever: Highlighting the Need for a Systematic Molecular Approach for a Neglected Disease in Africa. Acta Trop. 2019, 193, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Njeru, J.; Henning, K.; Pletz, M.W.; Heller, R.; Neubauer, H. Q Fever Is an Old and Neglected Zoonotic Disease in Kenya: A Systematic Review. BMC Public Health 2016, 16, 297. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, M.; Nicholson, W.L.; Roche, A.J.; Kersh, G.J.; Fitzpatrick, K.A.; Oliver, L.D.; Massung, R.F.; Morrissey, A.B.; Bartlett, J.A.; Onyango, J.J.; et al. Q Fever, Spotted Fever Group, and Typhus Group Rickettsioses among Hospitalized Febrile Patients in Northern Tanzania. Clin. Infect. Dis. 2011, 53, e8–e15. [Google Scholar] [CrossRef] [Green Version]
- Pilloux, L.; Baumgartner, A.; Jaton, K.; Lienhard, R.; Ackermann-Gäumann, R.; Beuret, C.; Greub, G. Prevalence of Anaplasma phagocytophilum and Coxiella burnetii in Ixodes ricinus Ticks in Switzerland: An Underestimated Epidemiologic Risk. New Microbes New Infect. 2019, 27, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, R.C.; Echaide, I.E.; Alves, R.N.; Beletti, M.E.; Nava, S.; Labruna, M.B. Coxiella burnetii in Ticks, Argentina. Emerg. Infect. Dis. 2013, 19, 344–346. [Google Scholar] [CrossRef] [PubMed]
- Tokarevich, N.K.; Panferova, Y.A.; Freylikhman, O.A.; Blinova, O.V.; Medvedev, S.G.; Mironov, S.V.; Grigoryeva, L.A.; Tretyakov, K.A.; Dimova, T.; Zaharieva, M.M.; et al. Coxiella burnetii in Ticks and Wild Birds. Ticks Tick-Borne Dis. 2019, 10, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Koka, H.; Sang, R.; Kutima, H.L.; Musila, L. Coxiella burnetii Detected in Tick Samples from Pastoral Communities in Kenya. BioMed Res. Int. 2018, 2018, 8158102. [Google Scholar] [CrossRef] [Green Version]
- Knobel, D.L.; Maina, A.N.; Cutler, S.J.; Ogola, E.; Feikin, D.R.; Junghae, M.; Halliday, J.E.B.; Richards, A.L.; Breiman, R.F.; Cleaveland, S.; et al. Coxiella burnetii in Humans, Domestic Ruminants, and Ticks in Rural Western Kenya. Am. J. Trop. Med. Hyg. 2013, 88, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Mediannikov, O.; Fenollar, F.; Socolovschi, C.; Diatta, G.; Bassene, H.; Molez, J.-F.; Sokhna, C.; Trape, J.-F.; Raoult, D. Coxiella burnetii in Humans and Ticks in Rural Senegal. PLoS Negl. Trop. Dis. 2010, 4, e654. [Google Scholar] [CrossRef] [Green Version]
- Al-Hajjar, S.; Hussain Qadri, S.M.; Al-Sabban, E.; Jäger, C. Coxiella burnetii Endokarditis in a Child. Pediatr. Infect. Dis. J. 1997, 16, 911–913. [Google Scholar] [CrossRef] [PubMed]
- Arricau-Bouvery, N.; Hauck, Y.; Bejaoui, A.; Frangoulidis, D.; Bodier, C.C.; Souriau, A.; Meyer, H.; Neubauer, H.; Rodolakis, A.; Vergnaud, G. Molecular Characterization of Coxiella burnetii Isolates by Infrequent Restriction Site-PCR and MLVA Typing. BMC Microbiol. 2006, 6, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frangoulidis, D.; Walter, M.C.; Antwerpen, M.; Zimmermann, P.; Janowetz, B.; Alex, M.; Böttcher, J.; Henning, K.; Hilbert, A.; Ganter, M.; et al. Molecular Analysis of Coxiella burnetii in Germany Reveals Evolution of Unique Clonal Clusters. Int. J. Med. Microbiol. IJMM 2014, 304, 868–876. [Google Scholar] [CrossRef]
- Shriver, M.D.; Jin, L.; Boerwinkle, E.; Deka, R.; Ferrell, R.E.; Chakraborty, R. A Novel Measure of Genetic Distance for Highly Polymorphic Tandem Repeat Loci. Mol. Biol. Evol. 1995, 12, 914–920. [Google Scholar]
- Bellabidi, M.; Benaissa, M.H.; Bissati-Bouafia, S.; Harrat, Z.; Brahmi, K.; Kernif, T. Coxiella Burnetii in Camels (Camelus Dromedarius) from Algeria: Seroprevalence, Molecular Characterization, and Ticks (Acari: Ixodidae) Vectors. Acta Trop. 2020, 206, 105443. [Google Scholar] [CrossRef]
- Al-Deeb, M.A.; Frangoulidis, D.; Walter, M.C.; Kömpf, D.; Fischer, S.F.; Petney, T.; Muzaffar, S.B. Coxiella-like Endosymbiont in Argasid Ticks (Ornithodoros Muesebecki) from a Socotra Cormorant Colony in Umm Al Quwain, United Arab Emirates. Ticks Tick-Borne Dis. 2016, 7, 166–171. [Google Scholar] [CrossRef] [PubMed]
- McKinney, P. Puntland Food Security and Vulnerability Assessment April 2007; 2007. Available online: https://documents.wfp.org/stellent/groups/public/documents/ena/wfp144576.pdf (accessed on 8 April 2021).
- PLHDS. Puntland Report 2020 Somali Health and Demographic Survey; 2020. Available online: http://pl.statistics.so/wp-content/uploads/2020/11/Puntland_SHDS-Report-2020.pdf, (accessed on 8 April 2021).
- Shuaib, Y.A.; Elhag, A.M.-A.W.; Brima, Y.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.-T.; Lemhöfer, G.; Bestehorn, M.; Poppert, S.; Schaper, S.; et al. Ixodid Tick Species and Two Tick-Borne Pathogens in Three Areas in the Sudan. Parasitol. Res. 2020, 119, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Springer, A.; Shuaib, Y.A.; Isaa, M.H.; Ezz-Eldin, M.I.-E.; Osman, A.Y.; Yagoub, I.A.; Abdalla, M.A.; Bakiet, A.O.; Mohmed-Noor, S.E.-T.; Schaper, S.; et al. Tick Fauna and Associated Rickettsia, Theileria, and Babesia spp. in Domestic Animals in Sudan (North Kordofan and Kassala States). Microorganisms 2020, 8, 1969. [Google Scholar] [CrossRef]
- Voltzit, O.V.; Keirans, J.E. A Review of African Amblyomma Species (Acari, Ixodida, Ixodidae). In Acarina 112135-214, 2003. Available online: http://publikationen.ub.uni-frankfurt.de/frontdoor/index/index/docId/11904, (accessed on 5 February 2021).
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma. VI. Systematics of H. (Euhyalomma) truncatum and the Closely Related Species, H. (E.) albiparmatum and H. (E.) nitidum (Acari: Ixodidae). Exp. Appl. Acarol. 2008, 44, 115–136. [Google Scholar] [CrossRef] [Green Version]
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma Koch, 1844: V. Re-Evaluation of the Taxonomic Rank of Taxa Comprising the H. (Euhyalomma) marginatum Koch Complex of Species (Acari: Ixodidae) with Redescription of All Parasitic Stages and Notes on Biology. Int. J. Acarol. 2008, 34, 13–42. [Google Scholar] [CrossRef]
- Apanaskevich, D.A.; Horak, I.G. The Genus Hyalomma Koch, 1844. IX. Redescription of All Parasitic Stages of H. (Euhyalomma) impeltatum Schulze & Schlottke, 1930 and H. (E.) somalicum Tonelli Rondelli, 1935 (Acari: Ixodidae). Syst. Parasitol. 2009, 73, 199–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apanaskevich, D.A.; Schuster, A.L.; Horak, I.G. The Genus Hyalomma: VII. Redescription of All Parasitic Stages of H. (Euhyalomma) dromedarii and H. (E.) schulzei (Acari: Ixodidae). J. Med. Entomol. 2008, 45, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA Sequences and Phylogenetic Relationships of Species of Rhipicephalus and Other Tick Genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef] [PubMed]
- Duron, O. The IS1111 Insertion Sequence Used for Detection of Coxiella burnetii Is Widespread in Coxiella-like Endosymbionts of Ticks. FEMS Microbiol. Lett. 2015, 362, fnv132. [Google Scholar] [CrossRef] [Green Version]
- Jourdain, E.; Duron, O.; Barry, S.; González-Acuña, D.; Sidi-Boumedine, K. Molecular Methods Routinely Used to Detect Coxiella burnetii in Ticks Cross-React with Coxiella-like Bacteria. Infect. Ecol. Epidemiol. 2015, 5, 29230. [Google Scholar] [CrossRef] [Green Version]
- Frangoulidis, D.; Splettstoesser, W.D.; Landt, O.; Dehnhardt, J.; Henning, K.; Hilbert, A.; Bauer, T.; Antwerpen, M.; Meyer, H.; Walter, M.C.; et al. Microevolution of the Chromosomal Region of Acute Disease Antigen A (AdaA) in the Query (Q) Fever Agent Coxiella burnetii. PLoS ONE 2013, 8, e53440. [Google Scholar] [CrossRef] [Green Version]
- Fasemore, A.M.; Helbich, A.; Walter, M.C.; Dandekar, T.; Vergnaud, G.; Förstner, K.U.; Frangoulidis, D. CoxBase: An Online Platform for Epidemiological Surveillance, Visualization, Analysis and Typing of Coxiella burnetii Genomic Sequence. bioRxiv 2020. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| 4 | 6 | nr * | 7 | 6 | nr | nr | 3 | 3 | 5 | 5.5 | 3 | 7 | 2 | D21 (New) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ms01 | ms03 | ms01 | ms20 | ms22 | ms23 | ms24 | ms26 | ms27 | ms28 | ms30 | ms31 | ms33 | ms34 | Genotype |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 14.0 | 3.0 | 3.0 | 6.0 | 5.5 | 3.0 | 7.0 | 2.0 | D14 |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | −1.0 | 17.0 | −1.0 | 3.0 | 8.0 | 5.5 | 3.0 | 7.0 | 2.0 | D15 |
| 4.0 | 6.0 | 8.5 | 7.0 | 6.0 | −1.0 | 10.0 | 3.0 | 4.0 | 5.0 | 5.5 | 3.0 | 6.0 | 2.0 | D17 |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | −1.0 | 0.0 | −1.0 | 3.0 | 7.0 | 5.5 | 3.0 | 6.0 | 2.0 | D8 |
| 4.0 | 6.0 | 13.0 | 7.0 | 6.0 | 9.0 | 8.0 | 3.0 | 3.0 | 6.0 | 5.5 | 3.0 | 6.0 | 2.0 | D1 |
| 3.0 | 6.0 | 10.5 | 6.0 | 6.0 | 9.0 | 29.0 | 3.0 | 3.0 | 5.0 | 5.5 | 3.0 | 5.0 | 2.0 | B3 |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 14.0 | 3.0 | 3.0 | 6.0 | 5.5 | 3.0 | 7.0 | 2.0 | D14 |
| 4.0 | 6.0 | 8.5 | 7.0 | 7.0 | −1.0 | 11.0 | 2.0 | 4.0 | 5.0 | 5.5 | 3.0 | 8.0 | 2.0 | D10 |
| 3.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 3.0 | −1.0 | 3.0 | 7.0 | 5.5 | 3.0 | 7.0 | 2.0 | D12 |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 0.0 | −1.0 | 3.0 | 7.0 | 5.5 | 3.0 | 5.0 | 2.0 | D13 |
| 4.0 | 6.0 | 13.0 | 7.0 | 6.0 | 9.0 | 15.0 | 4.0 | 3.0 | 6.0 | 5.5 | 3.0 | 6.0 | 2.0 | D16 |
| 4.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 17.0 | −1.0 | 3.0 | 8.0 | 5.5 | 3.0 | 6.0 | 2.0 | D3 |
| 3.0 | 6.0 | 8.5 | 6.0 | 6.0 | 9.0 | 11.0 | 3.0 | 5.0 | 5.0 | 5.5 | 3.0 | 6.0 | 2.0 | D9 |
| 4.0 | 6.0 | 8.5 | 6.0 | 7.0 | 4.0 | 11.0 | 2.0 | 4.0 | 5.0 | 5.5 | 3.0 | 7.0 | 2.0 | D6 |
| 4.0 | 6.0 | 8.5 | 6.0 | 7.0 | 7.0 | 8.0 | 2.0 | 4.0 | 5.0 | 5.5 | 3.0 | 7.0 | 2.0 | D4 |
| 4.0 | 6.0 | 9.0 | 6.0 | 6.0 | 7.0 | 11.0 | 2.0 | 3.0 | 4.0 | 5.5 | 4.0 | 7.0 | 2.0 | D20 |
| 4.0 | 6.0 | 9.0 | 6.0 | 6.0 | 2.0 | 10.0 | 2.0 | 3.0 | 4.0 | 5.5 | 4.0 | 7.0 | 2.0 | D2 |
| 3.0 | 6.0 | 11.0 | 7.0 | 6.0 | 9.0 | 3.0 | −1.0 | 3.0 | 7.0 | 5.5 | 3.0 | 6.0 | 2.0 | D18 |
| 4.0 | 6.0 | 9.0 | 6.0 | 6.0 | 10.0 | 11.0 | 2.0 | 3.0 | 4.0 | 5.5 | 4.0 | 3.0 | 2.0 | D19 |
| 3.0 | 6.0 | 8.5 | 7.0 | 7.0 | 9.0 | 0.0 | 2.0 | 4.0 | 5.0 | 5.5 | 3.0 | 8.0 | 2.0 | D7 |
| 4.0 | 6.0 | 9.0 | 6.0 | 6.0 | 5.0 | 15.0 | 4.0 | 2.0 | 7.0 | 5.5 | 3.0 | 9.0 | 8.0 | C8 |
| 3.0 | 6.0 | 9.0 | 6.0 | 6.0 | 5.0 | 15.0 | 3.0 | 2.0 | 7.0 | 5.5 | 3.0 | 9.0 | 9.0 | C10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frangoulidis, D.; Kahlhofer, C.; Said, A.S.; Osman, A.Y.; Chitimia-Dobler, L.; Shuaib, Y.A. High Prevalence and New Genotype of Coxiella burnetii in Ticks Infesting Camels in Somalia. Pathogens 2021, 10, 741. https://doi.org/10.3390/pathogens10060741
Frangoulidis D, Kahlhofer C, Said AS, Osman AY, Chitimia-Dobler L, Shuaib YA. High Prevalence and New Genotype of Coxiella burnetii in Ticks Infesting Camels in Somalia. Pathogens. 2021; 10(6):741. https://doi.org/10.3390/pathogens10060741
Chicago/Turabian StyleFrangoulidis, Dimitrios, Claudia Kahlhofer, Ahmed Shire Said, Abdinasir Yusuf Osman, Lidia Chitimia-Dobler, and Yassir Adam Shuaib. 2021. "High Prevalence and New Genotype of Coxiella burnetii in Ticks Infesting Camels in Somalia" Pathogens 10, no. 6: 741. https://doi.org/10.3390/pathogens10060741