
Trypanosoma rangeli Genetic, Mammalian Hosts, and Geographical Diversity from Five Brazilian Biomes

 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
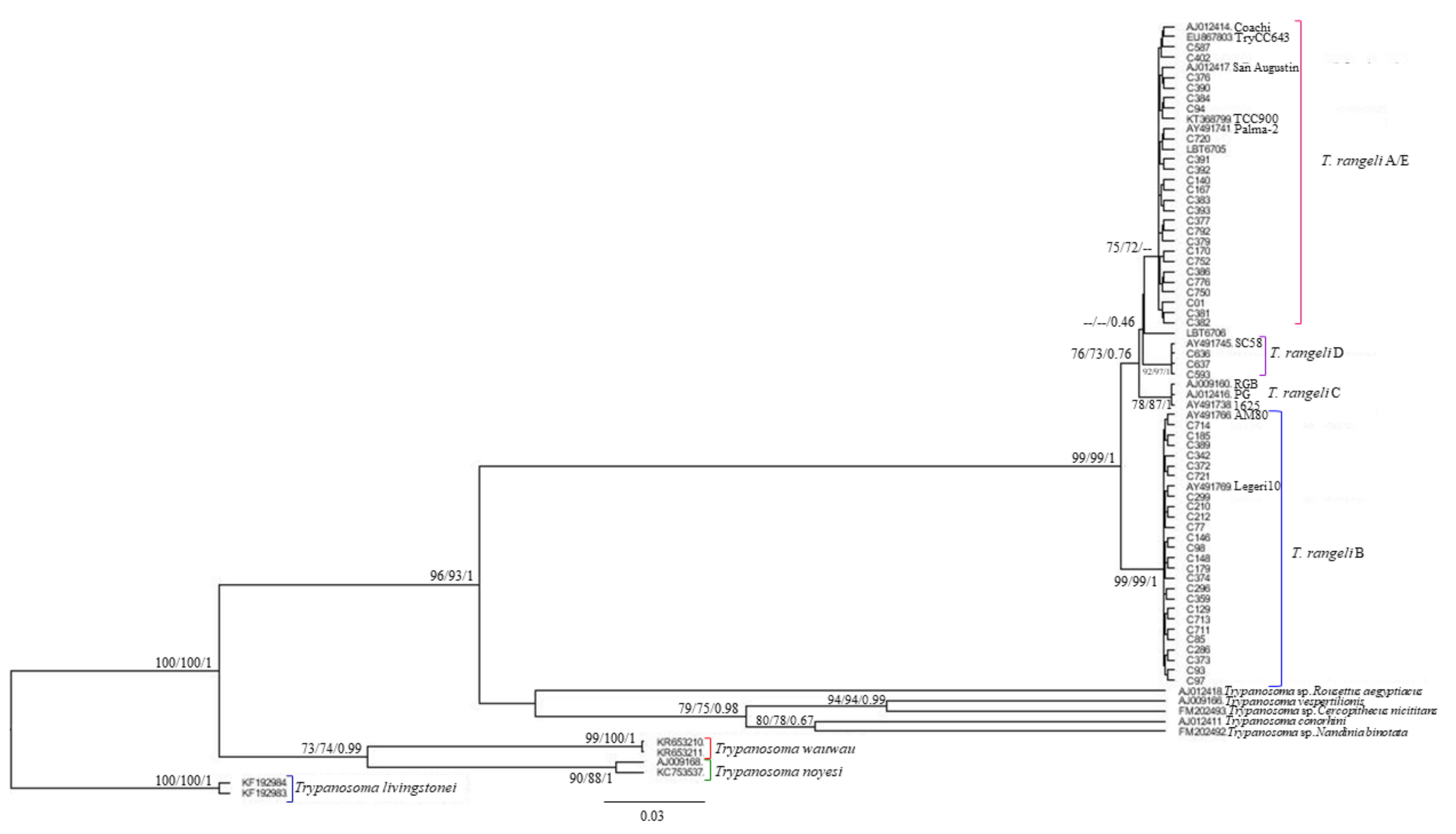
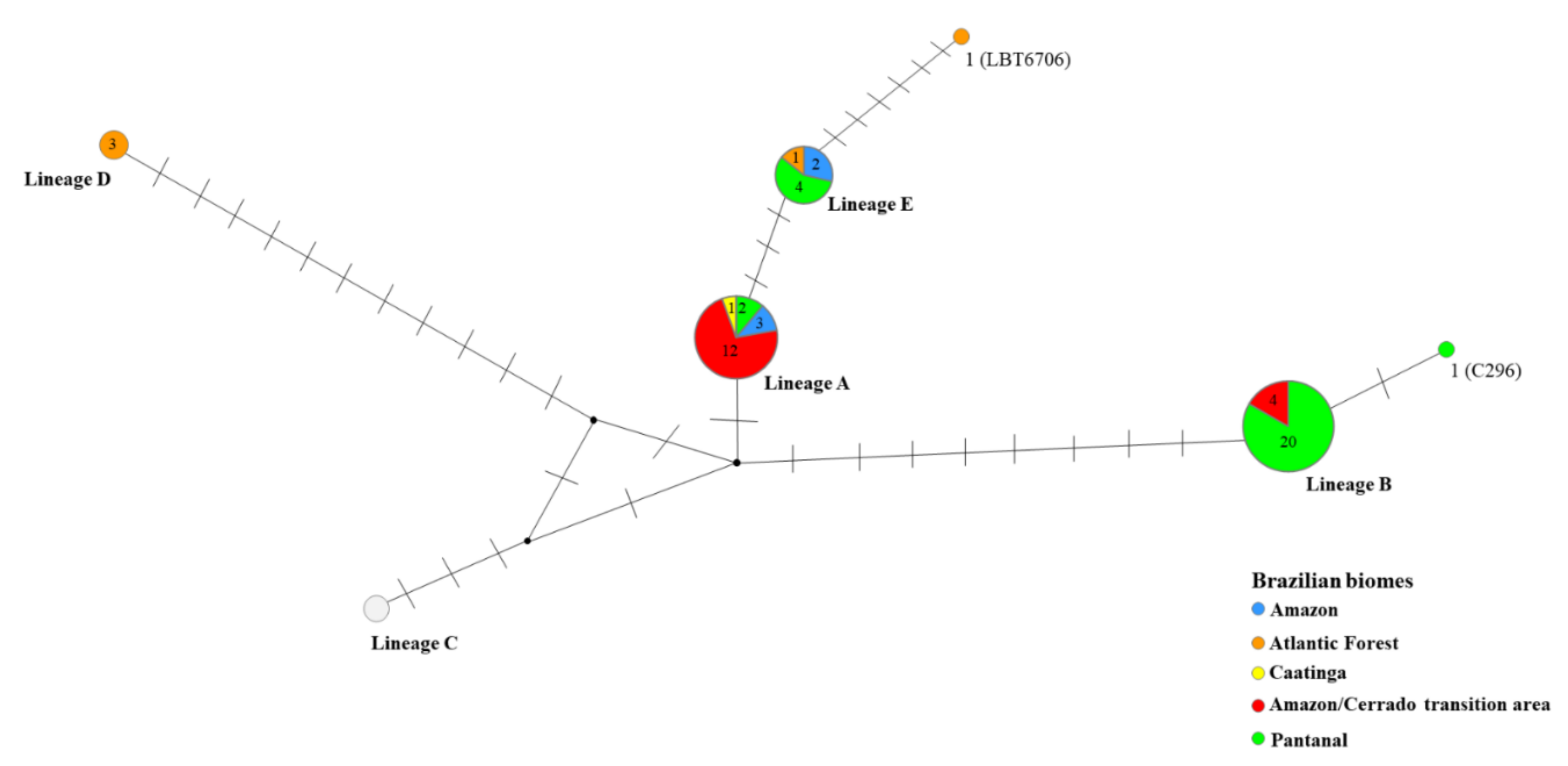
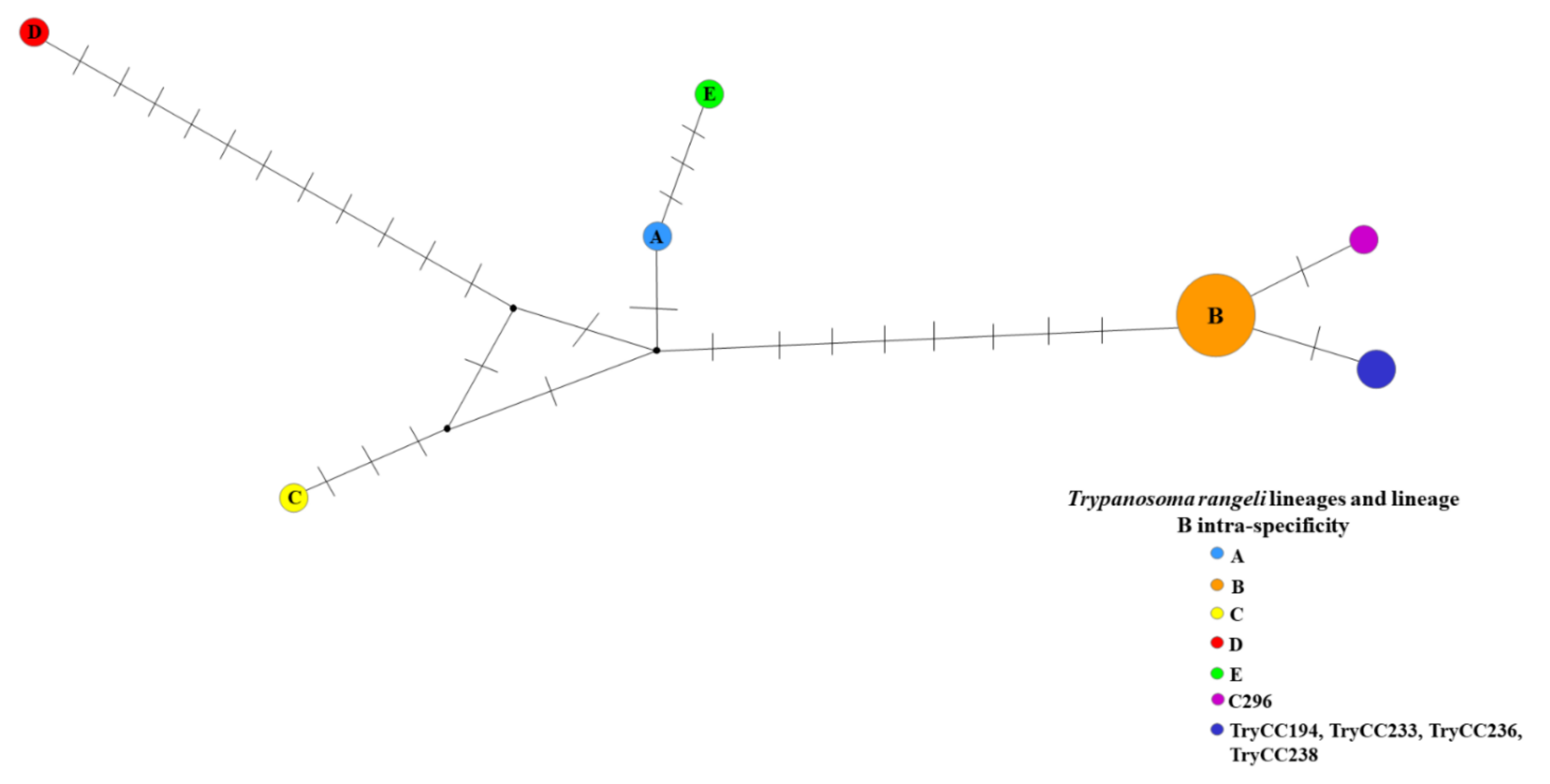
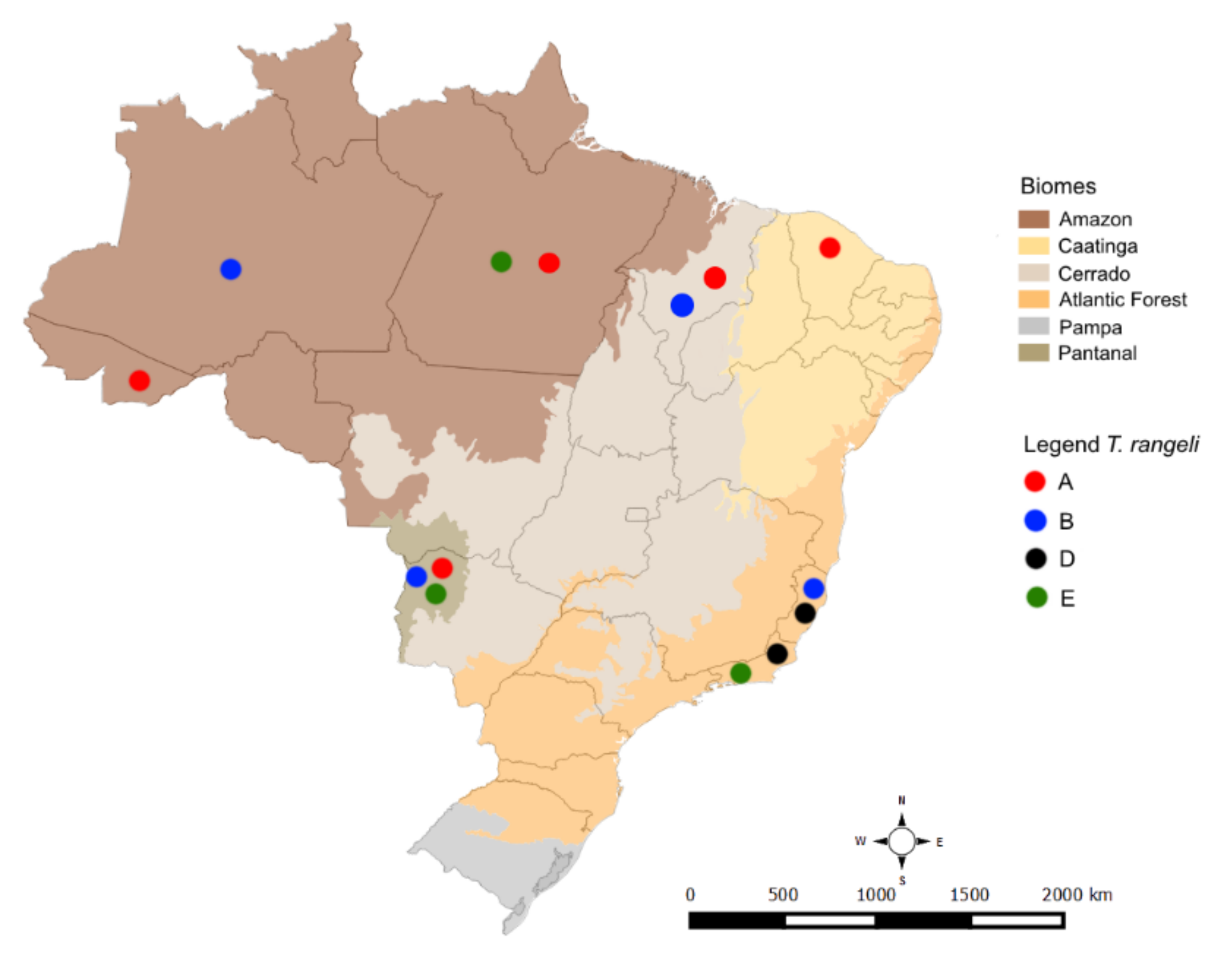
2. Results
3. Discussion
4. Material and Methods
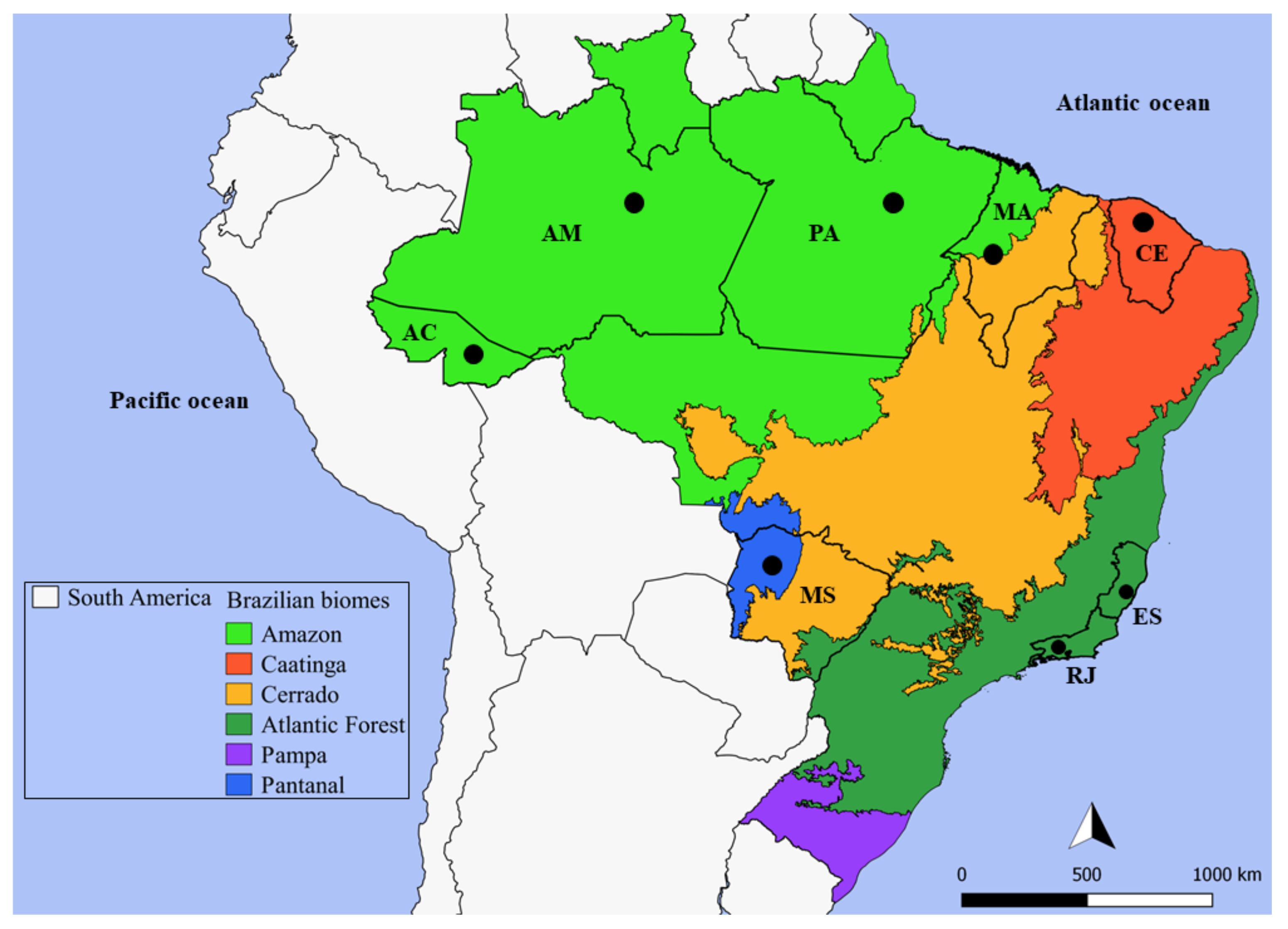
4.1. Trypanosoma Rangeli Study Areas
4.2. Trypanosoma Rangeli DNA Samples Origin
4.3. Trypanosoma Rangeli Molecular Characterization
4.4. Phylogenetic Analysis of Trypanosoma rangeli Lineages
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tereja, E. Um nouveau flagela de Rhodnius prolixus. Trypanosoma (ou Crithidia) rangeli n.sp. Bull. Soc. Path. Exot. 1920, 13, 527–530. [Google Scholar]
- D’Alessandro, A.; Saravia, N.G. Trypanosoma rangeli. In Parasitic Protozoa, 2nd ed.; Academic Press: San Diego, CA, USA, 1992; Volume 2, pp. 1–54. [Google Scholar]
- Coura, J.R.; Fernandes, O.; Arboleda, M.; Barret, T.V.; Carrara, N.; Degrave, W.; Campbell, D.A. Human infection by Trypanosoma rangeli in the Brazilian Amazon. Trans. R. Soc. Trop. Med. Hyg. 1996, 90, 278–279. [Google Scholar] [CrossRef]
- Grisard, E.C.; Steindel, M.; Guarneri, A.A.; Eger-Mangrich, I.; Campbell, D.A.; Romanha, A.J. Characterization of Trypanosoma rangeli strains isolated in Central and South America: An Overview. Mem. Inst. Oswaldo Cruz 1999, 94, 203–209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maia da Silva, F.; Rodrigues, A.C.; Campaner, M.; Takata, C.S.A.; Brigido, M.C.; Junqueira, A.C.V.; Coura, J.R.; Takeda, G.F.; Shaw, J.J.; Teixeira, M.M.G. Randomly amplified polymorphic DNA analysis of Trypanosoma rangeli and allied species from human, monkeys and other sylvatic mammals of the Brazilian Amazon disclosed a new group and a species-specific marker. Parasitology 2004, 128, 283–294. [Google Scholar] [CrossRef]
- Maia da Silva, F.; Marcili, A.; Lima, L.; Cavazzana, M., Jr.; Ortiz, P.A.; Campaner, M.; Takeda, G.F.; Paiva, F.; Nunes, V.L.B.; Camargo, E.P.; et al. Trypanosoma rangeli isolates of bats from Central Brazil: Genotyping and phylogenetic analysis enable description of a new lineage using spliced-leader gene sequences. Acta Trop. 2009, 109, 199–207. [Google Scholar] [CrossRef]
- Vallejo, G.A.; Guhl, F.; Schaub, G.A. Triatominae-Trypanosoma cruzi/T. rangeli: Vector-parasite interactions. Acta Trop. 2009, 110, 137–147. [Google Scholar] [CrossRef]
- Guhl, F.; Vallejo, G.A. Trypanosoma (Herpetosoma) rangeli Tejera, 1920: An updated review. Mem. Inst. Oswaldo Cruz 2003, 98, 435–442. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.A. Morphobiological characterization of Trypanosoma cruzi Chagas, 1909 and its distinction from other trypanosomes. Mem. Inst. Oswaldo Cruz 1999, 94, 205–210. [Google Scholar] [CrossRef] [Green Version]
- Eger-Mangrich, I.; de Oliveira, M.A.; Grisard, E.C.; de Souza, W.; Steindel, M. Interaction of Trypanosoma rangeli Tejera, 1920 with different cell lines in vitro. Parasitol. Res. 2001, 87, 505–509. [Google Scholar] [CrossRef]
- Grewal, M.S. Pathogenicity of Trypanosoma rangeli Tejera, in the invertebrate host. Exp. Parasitol. 1957, 6, 123–130. [Google Scholar] [CrossRef]
- Grisard, E.C. Salivaria or stercoraria? The Trypanosoma rangeli dilemma. Kinetoplast. Biol. Dis. 2002, 1, 5. [Google Scholar] [CrossRef]
- Guhl, F.; Hudson, L.; Marinkelle, C.J.; Jaramillo, C.A.; Bridge, D. Clinical Trypanosoma rangeli infection as a complication of Chagas’ disease. Parasitology 1987, 94, 475–484. [Google Scholar] [CrossRef]
- Machado, P.E.; Eger-Mangrich, I.; Rosa, G.; Koerich, L.B.; Grisard, E.C.; Steindel, M. Differential susceptibility of triatominae of the genus Rhodnius to Trypanosoma rangeli strains from different geographical origins. Int. J. Parasitol. 2001, 31, 631–633. [Google Scholar] [CrossRef]
- Ramirez, L.E.; Lages-Silva, E.; Alvarenga-Franco, F.; Matos, A.; Vargas, N.; Fernandes, O.; Zingales, B. High prevalence of Trypanosoma rangeli and Trypanosoma cruzi in opossums and triatomids in a formerly-endemic area of Chagas disease in Southeast Brazil. Acta Trop. 2002, 84, 189–198. [Google Scholar] [CrossRef]
- Peterson, J.K.; Graham, A.L.; Elliott, R.J.; Dobson, A.P.; Chávez, O.T. Trypanosoma cruzi–Trypanosoma rangeli co-infection ameliorates negative effects of single trypanosome infections in experimentally infected Rhodnius prolixus. Parasitology 2016, 143, 1157–1167. [Google Scholar] [CrossRef]
- Hoare, C.A. The Trypanosomes of mammals. In A Zoological Monograph; Blackweel Scientific Publications: Oxford, UK, 1972; pp. 327–400. [Google Scholar]
- Hecker, H.; Schwarzenbach, M.; Rudin, W. Development and interactions of Trypanosoma rangeli in and with the reduviid bug Rhodnius prolixus. Parasitol. Res. 1990, 76, 311–318. [Google Scholar] [CrossRef]
- Azambuja, P.; Garcia, E.S. Trypanosoma rangeli interactions within the vector Rhodnius prolixus—A mini review. Mem. Inst. Oswaldo Cruz 2005, 100, 567–572. [Google Scholar] [CrossRef] [Green Version]
- Macedo, A.M.; Vallejo, G.A.; Chiari, E.; Pena, S.D.J. DNA fingerprinting reveals relationships between strains of Trypanosoma rangeli and Trypanosoma cruzi. In DNA Fingerprinting: State of the Science; Pena, S.D.J., Chakraborty, R., Epplen, J.T., Jeffreys, A.J., Eds.; Birkhauser Verlag: Basel, Switzerland, 1993; pp. 321–329. [Google Scholar]
- Steindel, M.; Dias Neto, E.; Pinto, C.J.; Grisard, E.C.; Menezes, C.L.; Murta, S.M.; Simpson, A.J.; Romanha, A.J. Randomly amplified polymorphic DNA (RAPD) and isoenzyme analysis of Trypanosoma rangeli strains. J. Eukaryot. Microbiol. 1994, 41, 261–267. [Google Scholar] [CrossRef]
- Henríksson, J.; Solarj, A.; Rydaker, M.; Sousa, O.E.; Pettersson, U. Karyotype variability in Trypanosoma rangeli. Parasitology 1996, 112, 385–391. [Google Scholar] [CrossRef]
- Vallejo, G.A.; Guhl, F.; Carranza, J.C.; Lozano, L.E.; Sánchez, J.L.; Jaramillo, J.C.; Gualtero, D.; Castañeda, N.; Silva, J.C.; Steindel, M. kDNA markers define two major Trypanosoma rangeli lineages in Latin America. Acta Trop. 2002, 81, 77–82. [Google Scholar] [CrossRef]
- Vallejo, G.A.; Guhl, F.; Carranza, J.C.; Moreno, J.; Triana, O.; Grisard, C.E. Parity between kinetoplast: DNA and miniexon gene sequences supports either clonal evolution or speciation in Trypanosoma rangeli strains isolated from Rhodnius colombiensis, R. pallescens and R. prolixus in Colombia. Infect. Genet. Evol. 2003, 3, 39–45. [Google Scholar] [CrossRef]
- Urrea, D.A.; Carranza, J.C.; Cuba Cuba, C.A.; Gurgel-Gonçalves, R.; Guhl, F.; Schofield, C.J.; Triana, O.; Vallejo, G.A. Molecular characterization of Trypanosoma rangeli strains isolated from Rhodnius ecuadoriensis in Peru, R. colombiensis in Colombia and R. pallescens in Panama, supports a co-evolutionary association between parasites and vectors. Infect. Genet. Evol. 2005, 5, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Cabrine-Santos, M.; Ferreira, K.A.M.; Tosi, L.R.O.; Lages-Silva, E.; Ramírez, L.E.; Pedrosa, A.L. Karyotype variability in KP1(+) and KP1(−) strains of Trypanosoma rangeli isolated in Brazil and Colombia. Acta Trop. 2009, 110, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Maia da Silva, F.; Noyes, H.; Campaner, M.; Junqueira, A.C.V.; Coura, J.R.; Añez, N.; Shaw, J.J.; Stevens, J.R.; Teixeira, M.M.G. Phylogeny, taxonomy and grouping of Trypanosoma rangeli isolates from man, triatomines and sylvatic mammals from widespread geographical origin based on SSU and ITS ribosomal sequences. Parasitology 2004, 129, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Maia da Silva, F.; Junqueira, A.C.; Campaner, M.; Rodrigues, A.C.; Crisante, G.; Ramírez, L.E.; Caballero, Z.C.E.; Monteiro, F.A.; Coura, J.R.; Añez, N.; et al. Comparative phylogeography of Trypanosoma rangeli and Rhodnius (Hemiptera: Reduviidae) supports a long coexistence of parasite lineages and their sympatric vectors. Mol. Ecol. 2007, 16, 3361–3373. [Google Scholar] [CrossRef] [Green Version]
- Maia da Silva, F.; Naiff, R.D.; Marcili, A.; Gordo, M.; D’Affonseca Neto, J.A.; Naiff, M.F.; Franco, A.M.; Campaner, M.; Valente, V.; Valente, S.A.; et al. Infection rates and genotypes of Trypanosoma rangeli and T. cruzi infecting free-ranging Saguinus bicolor (Callitrichidae), a critically endangered primate of the Amazon Rainforest. Acta Trop. 2008, 107, 168–173. [Google Scholar] [CrossRef]
- Ortiz, P.A.; Maia da Silva, F.; Cortez, A.P.; Lima, L.; Campaner, M.; Pral, E.M.; Alfieri, S.C.; Teixeira, M.M.G. Genes of cathepsin L-like proteases in Trypanosoma rangeli isolates: Markers for diagnosis, genotyping and phylogenetic relationships. Acta Trop. 2009, 112, 249–259. [Google Scholar] [CrossRef]
- Caballero, Z.C.; Costa-Martins, A.G.; Ferreira, R.C.; Alves, J.M.P.; Serrano, M.G.; Camargo, E.P.; Buck, G.A.; Minoprio, P.; Teixeira, M.M.G. Phylogenetic and syntenic data support a single horizontal transference to a Trypanosoma ancestor of a prokaryotic proline racemase implicated in parasite evasion from host defences. Parasites Vectors 2015, 8, 222. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Álvarez, O.; Ortiz, P.A.; Lima, L.; Costa-Martins, A.G.; Serrano, M.G.; Herder, S.; Buck, G.A.; Camargo, E.P.; Hamilton, P.B.; Stevens, J.R.; et al. Trypanosoma rangeli is phylogenetically closer to Old World trypanosomes than to Trypanosoma cruzi. Int. J. Parasitol. 2018, 48, 569–584. [Google Scholar] [CrossRef]
- Miles, M.A.; Arias, J.R.; Valente, S.A.; Naiff, R.D.; de Souza, A.A.; Povoa, M.M.; Lima, J.A.; Cedillos, R.A. Vertebrate hosts and vectors of Trypanosoma rangeli in the Amazon Basin of Brazil. Am. J. Trop. Med. Hyg. 1983, 32, 1251–1259. [Google Scholar] [CrossRef]
- De Araújo, V.A.; Boité, M.C.; Cupolillo, E.; Jansen, A.M.; Roque, A.L. Mixed infection in the anteater Tamandua tetradactyla (Mammalia: Pilosa) from Para State, Brazil: Trypanosoma cruzi, T. rangeli and Leishmania infantum. Parasitology 2013, 140, 455–460. [Google Scholar] [CrossRef]
- Alves, F.M.; Saab de Lima, J.; Rocha, F.L.; Herrera, H.M.; Mourão, G.D.M.; Jansen, A.M. Complexity and multi-factorial of Trypanosoma cruzi sylvatic cycle in coatis, Nasua Nasua (Procynidae), and triatomine bugs in Brazilian Pantanal. Parasites Vectors 2016, 9, 378. [Google Scholar] [CrossRef] [Green Version]
- Dario, M.A.; Moratelli, R.; Schwabl, P.; Jansen, A.M.; Llewellyn, M.S. Small subunit ribosomal metabarcoding reveals extraordinary trypanosomatid diversity in Brazilian bats. PLoS Negl. Trop. Dis. 2017, 11, e0005790. [Google Scholar] [CrossRef] [Green Version]
- Dario, M.A.; Lisboa, C.V.; Costa, L.M.; Moratelli, R.; Nascimento, M.P.; Costa, L.P.; Leite, Y.L.R.; Llewellyn, M.S.; Xavier, S.C.D.C.; Roque, A.L.R.; et al. High Trypanosoma spp. diversity is maintained by bats and triatomines in Espírito Santo state, Brazil. PLoS ONE 2017, 12, e0188412. [Google Scholar] [CrossRef]
- Dos Santos, F.C.B.; Lisboa, C.V.; Xavier, S.C.C.; Dario, M.A.; Verde, R.S.; Calouro, A.M.; Roque, A.L.R.; Jansen, A.M. Trypanosoma sp. diversity in Amazonian bats (Chiroptera; Mammalia) from Acre State, Brazil. Parasitology 2018, 145, 828–837. [Google Scholar] [CrossRef]
- Steindel, M.; Carvalho Pinto, J.C.; Toma, H.K.; Mangia, R.H.R.; Ribeiro-Rodrigues, R.; Romanha, A.J. Trypanosoma rangeli (Tejera, 1920) isolated from a sylvatic rodent (Echimys dasythrix) in Santa Catarina Island, Santa Catarina State: First report of this trypanosome in southern Brazil. Mem. Inst. Oswaldo Cruz 1991, 86, 73–79. [Google Scholar] [CrossRef] [Green Version]
- Kluyber, D.; Desbiez, A.; Attias, N.; Massocato, G.; Gennari, S.; Soares, H.; Bagagli, E.; Bosco, S.M.G.; Garcés, H.G.; Ferreira, J.D.S.; et al. Zoonotic parasites infecting free-living armadillos from Brazil. Transbound. Emerg. Dis. 2020. [Google Scholar] [CrossRef]
- Crespo, J.A. Ecologia de la comunidad de mamiferos del Parque Nacional Iguazu, Misiones. Rev. MACN Ecol. 1982, 3, 45–162. [Google Scholar]
- Schaller, G.B. Mammals and their biomass on a Brazilian ranch. Arq. Zool. São Paulo 1983, 31, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Emmons, L.H. Neotropical Rainforest Mammals: A Field Guide; University of Chicago Press: Chicago, IL, USA, 1990; p. 281. [Google Scholar]
- Gompper, M.E.; Decker, D.M. Nasua nasua. Mamm. Species 1998, 580, 1–9. [Google Scholar] [CrossRef] [Green Version]
- McClearn, D. Locomotion, posture, and feeding behavior of kinkajous, coatis, and raccoons. J. Mammal. 1992, 73, 245–261. [Google Scholar] [CrossRef]
- Bianchi, R.C.; Campos, R.C.; Xavier-Filho, N.L.; Olifiers, N.; Gompper, M.E.; Mourão, G.M. Intraspecific, interspecific, and seasonal differences in the diet of three mid-sized carnivores in a large neotropical wetland. Acta Theriol. 2014, 59, 13–23. [Google Scholar] [CrossRef]
- Sussman, R.W.; Garber, P.A. A new interpretation of the social organization and mating system of the Callitrichidae. Int. J. Primatol. 1987, 8, 73–92. [Google Scholar] [CrossRef]
- Snowdon, C.T.; Soini, P. The tamarins, genus Saguinus. In Ecology and Behavior of Neotropical Primates; MitterMeier, A., Rylands, A.B., Coimbra-Filho, A.F., da Fonseca, G.A., Eds.; World Wildlife Fund: Washington, DC, USA, 1988; Volume 2, pp. 223–298. [Google Scholar]
- Tardif, S.D.; Harrison, M.L.; Simek, M.A. Communal infant care in marmosets and tamarins: Relation to energetics, ecology, and social organization. In Marmosets and Tamarins: Systematics, Behaviour and Ecology; Rylands, A.B., Ed.; Oxford University Press: Oxford, UK, 1993; pp. 220–234. [Google Scholar]
- Auricchio, P. Primatas do Brasil; Terra Brasilis Editora: São Paulo, Brazil, 1995. [Google Scholar]
- Garber, P.A.; Paciulli, I.M. Experimental field study of spatial memory and learning in wild capuchin monkeys (Cebus capucinus). Folia Primatol. 1997, 68, 236–253. [Google Scholar] [CrossRef]
- Porfirio, G.; Santos, F.M.; Foster, V.; Nascimento, L.F.; Macedo, G.C.; Barreto, W.T.G.; Fonseca, C.; Herrera, H.M. Terrestriality of wild Sapajus cay (Illiger, 1815) as revealed by camera traps. Folia Primatol. 2017, 88, 1–8. [Google Scholar] [CrossRef]
- Visalberghi, E. Responsiveness to objects in two social groups of tufted capuchin monkeys (Cebus apella). Am. J. Primatol. 1988, 15, 349–360. [Google Scholar] [CrossRef]
- Deane, L. Finding of Trypanosoma of the rangeli type in opossums of the species Didelphis marsupialis marsupialis in the state of Pará. Rev. Bras. Malariol. Doencas. Trop. 1958, 10, 451–458. [Google Scholar]
- Brandão, E.M.V.; Xavier, S.C.C.; Carvalhaes, J.G.; D’Andrea, P.S.; Lemos, F.G.; Azevedo, F.C.; Cássia-Pires, R.; Jansen, A.M.; Roque, A.L.R. Trypanosomatids in small mammals of an agroecosystem in central Brazil: Another piece in the puzzle of parasite transmission in an anthropogenic landscape. Pathogens 2019, 8, 190. [Google Scholar] [CrossRef] [Green Version]
- Ocaña-Mayorga, S.; Aguirre-Villacis, F.; Pinto, C.M.; Vallejo, G.A.; Grijalva, M.J. Prevalence, genetic characterization, and 18S small subunit ribosomal RNA diversity of Trypanosoma rangeli in triatomine and mammal hosts in endemic areas for Chagas disease in Ecuador. Vector Borne Zoonotic Dis. 2015, 15, 732–742. [Google Scholar] [CrossRef] [Green Version]
- Jansen, A.M.; Xavier, S.C.C.; Roque, A.L.R. Trypanosoma cruzi transmission in the wild and its most important reservoir hosts in Brazil. Parasites Vectors 2018, 11, 502. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.S.; Lima, L.; Xavier, S.C.D.C.; Herrera, H.M.; Rocha, F.L.; Roque, A.L.R.; Teixeira, M.M.G.; Jansen, A.M. Uncovering Trypanosoma spp. diversity of wild mammals by the use of DNA from blood clots. Int. J. Parasitol. Parasites Wildl. 2019, 8, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Miguel Pinto, C.; Ocaña-Mayorga, S.; Lascano, M.S.; Grijalva, M.J. Infection by trypanosomes in marsupials and rodents associated with human dwellings in Ecuador. J. Parasitol. 2006, 92, 1251–1255. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, J.A.; Bonvicino, C.R. Ordem rodentia. In Mamíferos do Brasil; Reis, N.R.D., Peracchi, A.L., Pedro, W.A., de Lima, I.P.d., Eds.; Suprema: Londrina, Brazil, 2006; pp. 347–400. [Google Scholar]
- Charles-Dominique, P.; Atramentowicz, M.; Charles-Dominique, M.; Gerard, H.; Hladik, A.; Hladik, C.M.; Prévost, M. Les mammiferes frugivores arboricoles nocturnes d’une foret Guyanaise: Inter-relations plants-animaux. Terre Vie 1981, 35, 341–435. [Google Scholar]
- Eisenberg, J.F. The evolution of arboreal herbivores in the Class Mammalia. In The Ecology of Arboreal Folivores; Montgomery, G., Ed.; Smithsonian Inst. Press: Washington, DC, USA, 1978; pp. 135–152. [Google Scholar]
- Attias, N.; Raíces, D.S.L.; Pessoa, F.S.; Albuquerque, H.; Jordão-Nogueira, T.; Modesto, T.C.; Bergallo, H.D.G. Potential distribution and new records of Trinomys species (Rodentia: Echimyidae) in the state of Rio de Janeiro. Zoologia 2009, 26, 305–315. [Google Scholar] [CrossRef] [Green Version]
- Desbiez, A.L.J.; Kluyber, D. The role of giant armadillos (Priodontes maximus) as physical ecosystem engineers. Biotropica 2013, 45, 537–540. [Google Scholar] [CrossRef]
- Anacleto, T.C.S.; Marinho-Filho, J. Hábito alimentar do tatu-canastras (Xenarthra, Dasypodidae) em uma área do cerrado do Brasil central. Rev. Bras. Zool. 2001, 18, 681–688. [Google Scholar] [CrossRef] [Green Version]
- Vaz, V.C.; Santori, R.T.; Jansen, A.M.; Delciellos, A.C.; D’Andrea, P.S. Notes on food habits of armadillos (Cingulata, Dasypodidae) and anteaters (Pilosa, Myrmecophagidae) at Serra da Capivara National Park (Piauí State, Brazil). Edentata 2012, 13, 84–89. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, A.; Barreto, P.; Saravia, N.; Barreto, S. Epidemiology of Trypanosoma cruzi in the oriental plains of Colombia. Am. J. Trop. Med. Hyg. 1984, 33, 1084–1095. [Google Scholar] [CrossRef]
- Scorza, C.; Urdaneta-Morales, S.; Tejero, F. Trypanosoma (Herpetosoma) rangeli Tejera, 1920. Preliminary report on histopathology in experimentally infected mice. Rev. Inst. Med. Trop. Sao Paulo 1986, 28, 371–378. [Google Scholar] [CrossRef] [Green Version]
- Osório, Y.; Travi, B.L.; Palma, G.; Saravia, N.G. Infectivity of Trypanosoma rangeli in a promonocytic mammalian cell line. J. Parasitol. 1995, 81, 687–693. [Google Scholar] [CrossRef]
- Ferreira, L.D.L.; Pereira, M.H.; Guarneri, A.A. Revisiting Trypanosoma rangeli transmission involving susceptible and non-susceptible Hosts. PLoS ONE 2015, 10, e0140575. [Google Scholar] [CrossRef] [Green Version]
- Lima, L.; Espinosa-Alvarez, O.; Hamilton, P.B.; Neves, L.; Takata, C.S.; Campaner, M.; Attias, M.; de Souza, W.; Camargo, E.P.; Teixeira, M.M.G. Trypanosoma livingstonei: A new species from African bats supports the bat seeding hypothesis for the Trypanosoma cruzi clade. Parasites Vectors 2013, 6, 221. [Google Scholar] [CrossRef] [Green Version]
- Ramirez, J.D.; Tapia-Calle, G.; Munoz-Cruz, G.; Poveda, C.; Lina, M.R.; Hincapié, E.; Guhl, F. Trypanosome species in neo-tropical bats: Biological, evolutionary and epidemiological implications. Infect. Genet. Evol. 2014, 22, 250–256. [Google Scholar] [CrossRef]
- Lima, L.; Espinosa-Alvarez, O.; Pinto, C.M.; Cavazzana, M., Jr.; Pavan, A.C.; Carranza, J.C.; Lim, B.; Campaner, M.; Takata, C.S.A.; Camargo, E.P.; et al. New insights into the evolution of the Trypanosoma cruzi clade provided by a new trypanosome species tightly linked to Neotropical Pteronotus bats and related to an Australian lineage of trypanosomes. Parasites Vectors 2015, 8, 657. [Google Scholar] [CrossRef]
- Rangel, D.A.; Lisboa, C.V.; Novaes, R.L.M.; Silva, B.A.; Souza, R.D.F.; Jansen, A.M.; Moratelli, R.; Roque, A.L.R. Isolation and characterization of trypanosomatids, including Crithidia mellificae, in bats from the Atlantic Forest of Rio de Janeiro, Brazil. PLoS Negl. Trop. Dis. 2019, 13, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Cavazzana, M.; Marcili, A.; Lima, L.; da Silva, F.M.; Junqueira, A.C.V.; Veludo, H.H.; Viola, L.B.; Campaner, M.; Nunes, V.L.B.; Paiva, F.; et al. Phylogeographic, ecological and biological patterns shown by nuclear (ssrRNA and gGAPDH) and mitochondrial (Cyt b) genes of trypanosomes of the subgenus Schizotypal parasitic in Brazilian bats. Int. J. Parasitol. 2010, 40, 345–355. [Google Scholar] [CrossRef]
- Wilkinson, G.S. Social grooming in the common 855 vampire bat, Desmodus rotundus. Anim. Behav. 1986, 34, 1880–1889. [Google Scholar] [CrossRef] [Green Version]
- Findley, J.S. Bats: A Community Perspective, 1st ed.; Cambridge University Press: New York, NY, USA, 1993. [Google Scholar]
- Kalko, E.K.V. Neotropical leaf-nosed bats (Phyllostomidae): “Whispering” bats or candidates for acoustic survey? In Proceedings of a Workshop on Identification and Acoustic Monitoring of Bats; Brigham, M., Jones, G., Kalko, E.K.V., Eds.; Bat Conservation International: Austin, TX, USA, 2002; pp. 63–69. [Google Scholar]
- Thomas, M.E.; Rasweiler, J.J.; D’Alessandro, A. Experimental transmission of the parasitic flagellates Trypanosoma cruzi and Trypanosoma rangeli between triatomine bugs or mice and captive neotropical bats. Mem. Inst. Oswaldo Cruz 2007, 102, 559–565. [Google Scholar] [CrossRef]
- Pífano, F.; Peñalver, L.M.; Medina, R.; Dominguez, E. La infección natural del perro por el Trypanosoma rangeli. Primera comprobación de un reservorio vertebrado extra-humano de protozoario de la natureza. Gac. Med. Car. 1948, 56, 7. [Google Scholar]
- D’alessandro, A. Biology of Trypanosoma (Herpetosoma) rangeli, Tejera, 1920. In The Biology of the Kinetoplastida; Lumsden, W.H.R., Evans, D.A., Eds.; Academic Press: New York, NY, USA, 1976; pp. 327–403. [Google Scholar]
- Xavier, S.C.; Roque, A.L.; Bilac, D.; de Araújo, V.A.; da Costa Neto, S.F.; Lorosa, E.S.; da Silva, L.F.; Jansen, A.M. Distantiae transmission of Trypanosoma cruzi: A new epidemiological feature of acute Chagas disease in Brazil. PLoS Negl. Trop. Dis. 2014, 8, e2878. [Google Scholar] [CrossRef] [Green Version]
- D’Alessandro, A.; de Hincapie, O. Rhodnius neivai: A new experimental vector of Trypanosoma rangeli. Am. J. Trop. Med. Hyg. 1986, 35, 512–514. [Google Scholar] [CrossRef]
- De Stefani Marquez, D.; Rodrigues-Ottaiano, C.; Monica Oliveira, R.; Pedrosa, A.L.; Cabrine-Santos, M.; Lages-Silva, E.; Ramírez, L.E. Susceptibility of different triatomine species to Trypanosoma rangeli experimental infection. Vector Borne Zoonotic Dis. 2006, 6, 50–56. [Google Scholar] [CrossRef]
- Gurgel-Gonçalves, R.; Galvão, C.; Costa, J.; Peterson, A.T. Geographic distribution of Chagas disease vectors in Brazil based on ecological niche modelling. J. Trop. Med. 2012, 2012, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Santos, F.M.; Jansen, A.M.; Mourão, G.M.; Jurberg, J.; Nunes, A.P.; Herrera, H.M. Triatominae (Hemiptera, Reduviidae) in the Pantanal region: Association with Trypanosoma cruzi, different habitats and vertebrate hosts. Rev. Soc. Bras. Med. Trop. 2015, 48, 532–538. [Google Scholar] [CrossRef] [Green Version]
- Lima-Oliveira, T.M.; Fontes, F.V.H.M.; Lilioso, M.; Pires-Silva, D.; Teixeira, M.M.G.; Meza, J.G.V.; Harry, M.; Fileé, J.; Costa, J.; Valença-Barbosa, C.; et al. Molecular eco-epidemiology on the sympatric Chagas disease vectors Triatoma braziliensis and Triatoma petrocchiae: Ectopes, genetic variation, natural infection prevalence by trypanosomatids and parasite genotyping. Acta Trop. 2020, 201, 105188. [Google Scholar] [CrossRef]
- Ingales, B.; Miles, M.A.; Campbell, D.A.; Tibayrenc, M.; Macedo, A.M.; Teixeira, M.M.; Schijman, A.G.; Llewellyn, M.S.; Lages-Silva, E.; Machado, C.R.; et al. The revised Trypanosoma cruzi subspecific nomenclature: Rationale, epidemiological relevance and research applications. Infect. Genet. Evol. 2012, 12, 240–253. [Google Scholar] [CrossRef]
- Grisard, E.C.; Campbell, D.A.; Romanha, A.J. Mini-exon gene sequence polymorphism among Trypanosoma rangeli strains isolated from distinct geographical regions. Parasitology 1999, 118, 375–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuervo, C.; Lopez, M.C.; Puerta, C. The Trypanosoma rangeli histone H2A gene sequence serves as a differential marker for KP1 strains. Infect. Genet. Evol. 2006, 6, 401–409. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Turner, W.R.; Larsen, F.W.; Brooks, T.M.; Gascon, C. Global biodiversity conservation: The critical role of hotspots. In Biodiversity Hotspots; Zachos, F.E., Habel, J.C., Eds.; Springer Publishers: London, UK, 2011; pp. 3–22. [Google Scholar]
- Junk, W.J.; Cunha, C.N.; Wantzen, K.M.; Petermann, P.; Strussmann, C.; Marques, M.I.; Adis, J. Biodiversity and its conservation in the Pantanal of Mato Grosso, Brazil. Aquat. Sci. 2006, 68, 278–309. [Google Scholar] [CrossRef]
- Jansen, A.M.; Xavier, S.C.D.C.; Roque, A.L.R. Landmarks of the Knowledge and Trypanosoma cruzi Biology in the Wild Environment. Front. Cell. Infect. Microbiol. 2020, 6, 10. [Google Scholar] [CrossRef]
- Sincero, T.C.; Stoco, P.H.; Steindel, M.; Vallejo, G.A.; Grisard, E.C. Trypanosoma rangeli displays a clonal population structure, revealing a subdivision of KP1(−) strains and the ancestry of the Amazonian group. Int. J. Parasitol. 2015, 45, 225–235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lisboa, C.V.; Mangia, R.H.; Luz, S.L.; Kluczkovski, A., Jr.; Ferreira, L.F.; Ribeiro, C.T.; Fernandes, O.; Jansen, A.M. Stable infection of primates with Trypanosoma cruzi I and II. Parasitology 2006, 133, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Xavier, S.C.; Roque, A.L.; Lima, V.D.S.; Monteiro, K.J.; Otaviano, J.C.; Ferreira da Silva, L.F.; Jansen, A.M. Lower richness of small wild mammal species and chagas disease risk. PLoS Negl. Trop. Dis. 2012, 6, e1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malavazi, P.F.N.S.; Daudt, C.; Melchior, L.A.K.; Meneguetti, D.U.O.; Xavier, S.C.C.; Jansen, A.M.; Souza, S.F.; Roque, A.L.R. Trypanosomes of vectors and domestic dogs in Trypanosoma cruzi transmission areas from Brazilian southwestern amazon: New mammalian host for Trypanosoma janseni. Acta Trop. 2020, 210, 105504. [Google Scholar] [CrossRef]
- Vallejo, G.A.; Guhl, F.; Chiari, E.; Macedo, A.M. Species specific detection of Trypanosoma cruzi and Trypanosoma rangeli in vector and mammal hosts by polymerase chain reaction amplification of kinetoplast minicircle DNA. Acta Trop. 1999, 72, 203–212. [Google Scholar] [CrossRef]
- Borghesan, T.C.; Ferreira, R.C.; Takata, C.S.; Campaner, M.; Borda, C.C.; Paiva, F.; Milder, R.V.; Teixeira, M.M.; Camargo, E.P. Molecular phylogenetic redefinition of Herpetomonas (Kinetoplastea, Trypanosomatidae), a genus of insect parasites associated with flies. Protist 2013, 164, 129–152. [Google Scholar] [CrossRef]
- Noyes, H.A.; Stevens, J.R.; Teixeira, M.; Phelan, J.; Holz, P. A nested PCR for the ssrRNA gene detects Trypanosoma binneyi in the platypus and Trypanosoma sp. in wombats and kangaroos in Australia. Int. J. Parasitol. 1999, 29, 331–339. [Google Scholar] [CrossRef]
- Smith, A.; Clark, P.; Averis, S.; Lymbery, A.J.; Wayne, A.F.; Morris, K.D.; Thompson, R.C.A. Trypanosomes in a declining species of threatened Australian marsupial, the brush-tailed bettong Bettongia penicillata (Marsupialia: Potoroidae). Parasitology 2008, 135, 1329–1335. [Google Scholar] [CrossRef] [Green Version]
- Katoh, S. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace aware data structure for phylogenomic inference from supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An advanced software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarisation in Bayesian phylogenetics using Tracer 1.7. Systemat. Biol. 2018. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.-J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Polzin, T.; Daneschmand, S.V. On Steiner trees and minimum spanning trees in hypergraphs. Oper. Res. Lett. 2003, 31, 12–20. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Order | Species | T. rangeli Infection/Total Examined | Infection Rate (%) | Biome | T. rangeli Lineage |
|---|---|---|---|---|---|
| Carnivora | Nasua nasua | 22/189 | 11.6 | Pantanal | A and B |
| Canis familiaris | 4/119 | 3.4 | Amazon Forest (n = 2) | A and E | |
| Atlantic Forest (n = 2 *) | E | ||||
| Procyon cancrivorus | 1/17 | 5.9 | Pantanal | E | |
| Cingulata | Priodontes maximus | 3 **/10 | 30 | Pantanal | E |
| Chiroptera | Carollia perspicillata | 3/279 | 1.1 | Amazon Forest (n = 1) | A |
| Atlantic Forest (n = 2) | B and D | ||||
| Didelphimorphia | Didelphis albiventris | 1/303 | 0.3 | Caatinga | A |
| Didelphis aurita | 1/271 | 0.4 | Atlantic Forest | D | |
| Didelphis marsupialis | 1/58 | 1.7 | Amazon Forest | A | |
| Philander opossum | 1/59 | 1.7 | Amazon Forest | E | |
| Primates | Alouatta belzebul | 1/6 | 16.7 | Amazon-Cerrado transition area | B |
| Alouatta caraya | 1/5 | 20 | Amazon-Cerrado transition area | B | |
| Sapajus libidinosus | 15/46 | 32.6 | Amazon-Cerrado transition area | A and B | |
| Saguinus bicolor bicolor | 1/24 | 4.2 | Amazon | B | |
| Rodentia | Coendou prehensilis | 1/5 | 20 | Amazon-Cerrado transition area | A |
| Trinomys dimidiatus | 1/1 | 100 | Atlantic Forest | D | |
| Total | 57/1392 | 4.1% | - | - |
| Order | Species | Number of Samples |
|---|---|---|
| Carnivora | Nasua nasua | 189 |
| Canis familiaris | 119 | |
| Procyon cancrivorus | 17 | |
| Cingulata | Priodontes maximus | 10 |
| Chiroptera | Carollia perspicillata | 279 |
| Didelphimorphia | Didelphis albiventris | 303 |
| Didelphis aurita | 271 | |
| Didelphis marsupialis | 58 | |
| Philander opossum | 59 | |
| Primates | Alouatta belzebul | 6 |
| Alouatta caraya | 5 | |
| Sapajus libidinosus | 46 | |
| Saguinus bicolor bicolor | 24 | |
| Rodentia | Coendou prehensilis | 5 |
| Trinomys dimidiatus | 1 | |
| Total | 15 | 1392 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dario, M.A.; Pavan, M.G.; Rodrigues, M.S.; Lisboa, C.V.; Kluyber, D.; Desbiez, A.L.J.; Herrera, H.M.; Roque, A.L.R.; Lima, L.; Teixeira, M.M.G.; et al. Trypanosoma rangeli Genetic, Mammalian Hosts, and Geographical Diversity from Five Brazilian Biomes. Pathogens 2021, 10, 736. https://doi.org/10.3390/pathogens10060736
Dario MA, Pavan MG, Rodrigues MS, Lisboa CV, Kluyber D, Desbiez ALJ, Herrera HM, Roque ALR, Lima L, Teixeira MMG, et al. Trypanosoma rangeli Genetic, Mammalian Hosts, and Geographical Diversity from Five Brazilian Biomes. Pathogens. 2021; 10(6):736. https://doi.org/10.3390/pathogens10060736
Chicago/Turabian StyleDario, Maria Augusta, Márcio Galvão Pavan, Marina Silva Rodrigues, Cristiane Varella Lisboa, Danilo Kluyber, Arnaud L. J. Desbiez, Heitor Miraglia Herrera, André Luiz Rodrigues Roque, Luciana Lima, Marta M. G. Teixeira, and et al. 2021. "Trypanosoma rangeli Genetic, Mammalian Hosts, and Geographical Diversity from Five Brazilian Biomes" Pathogens 10, no. 6: 736. https://doi.org/10.3390/pathogens10060736