
Serum Samples from Co-Infected and Domestic Cat Field Isolates Nonspecifically Bind FIV and Other Antigens in Enzyme-Linked Immunosorbent Assays


Abstract
:1. Introduction
2. Results
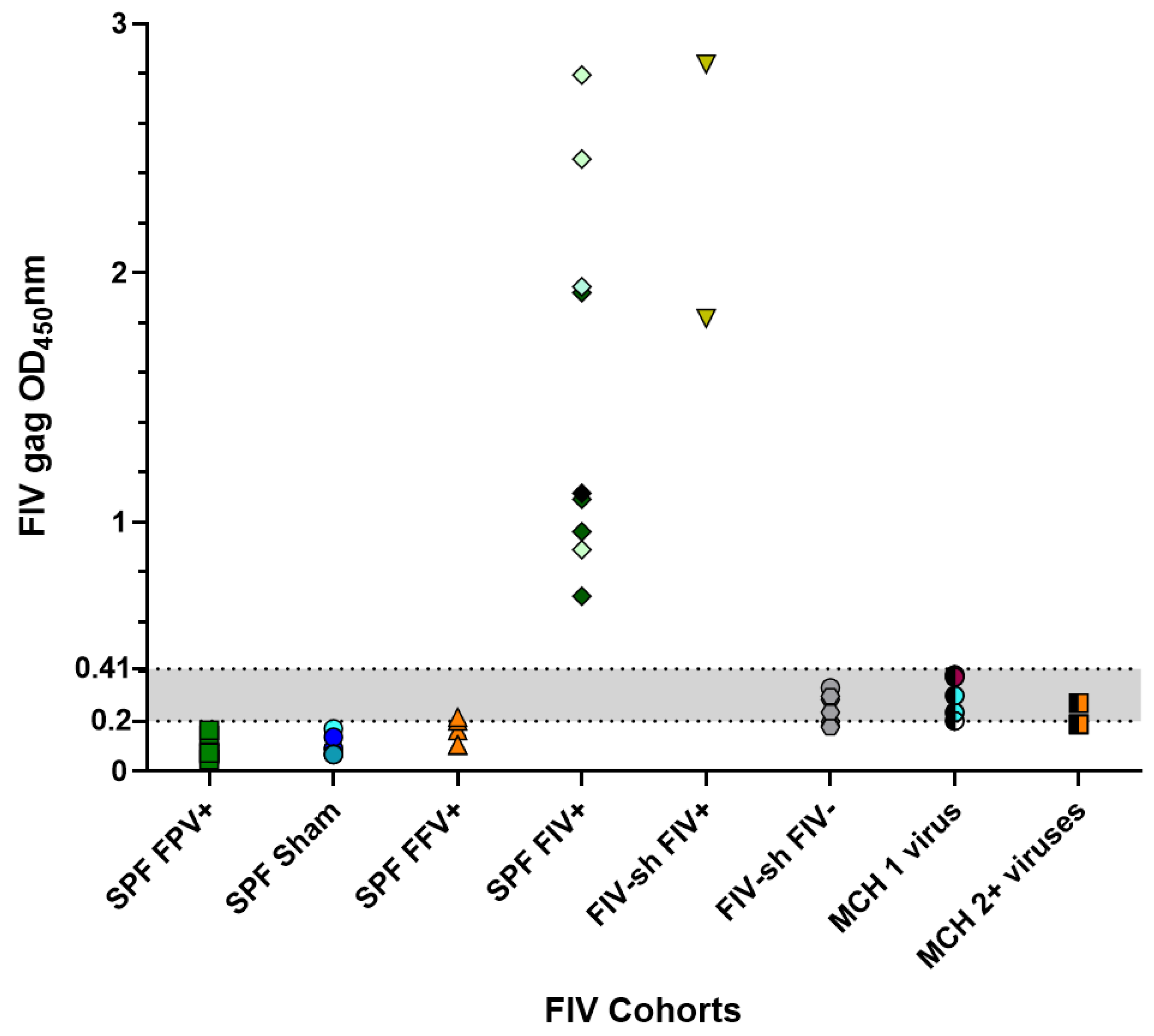
2.1. FIV SPF, Sham and Field Negative Results
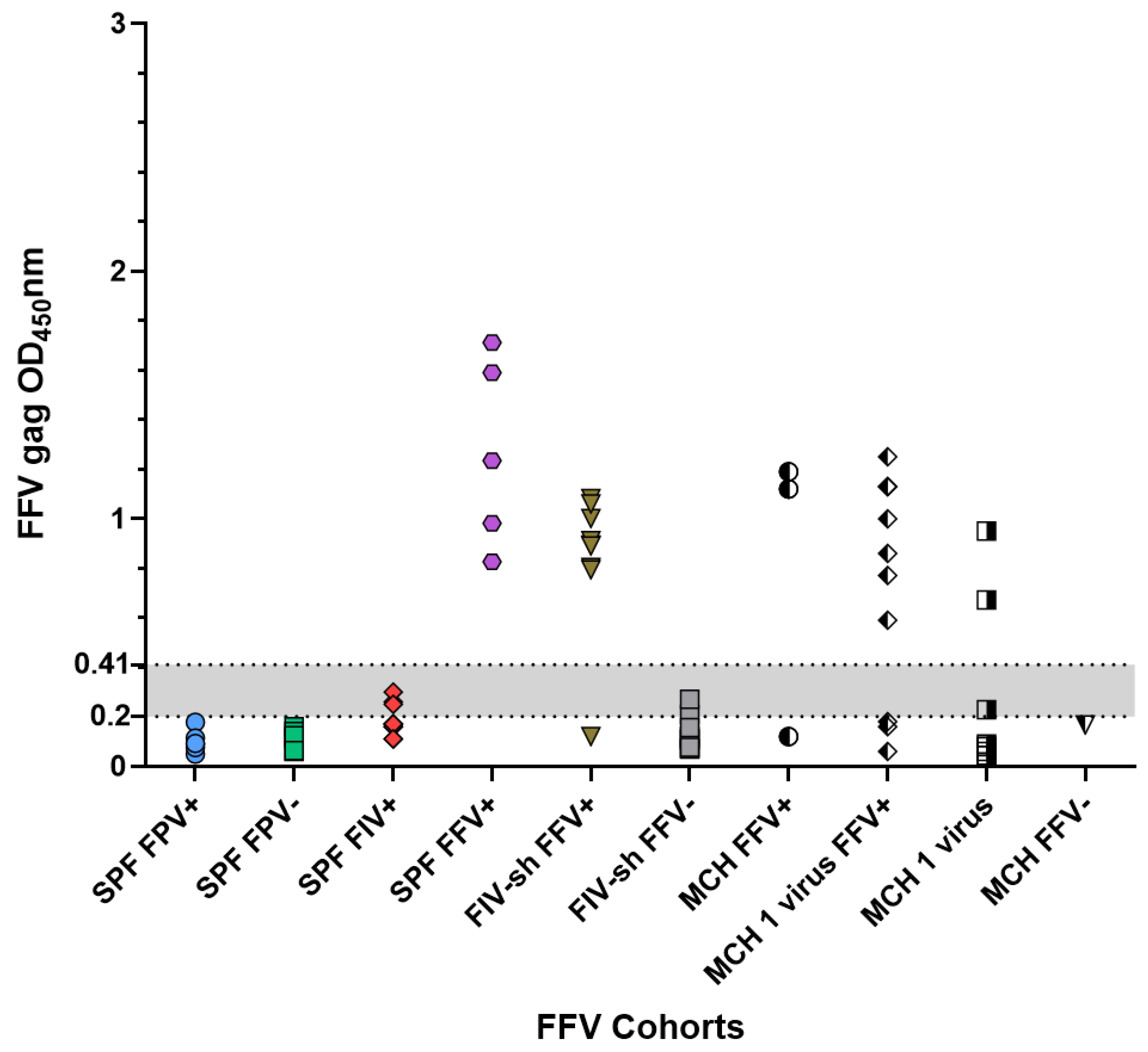
2.2. FFV SPF, Sham and Field Negative Results
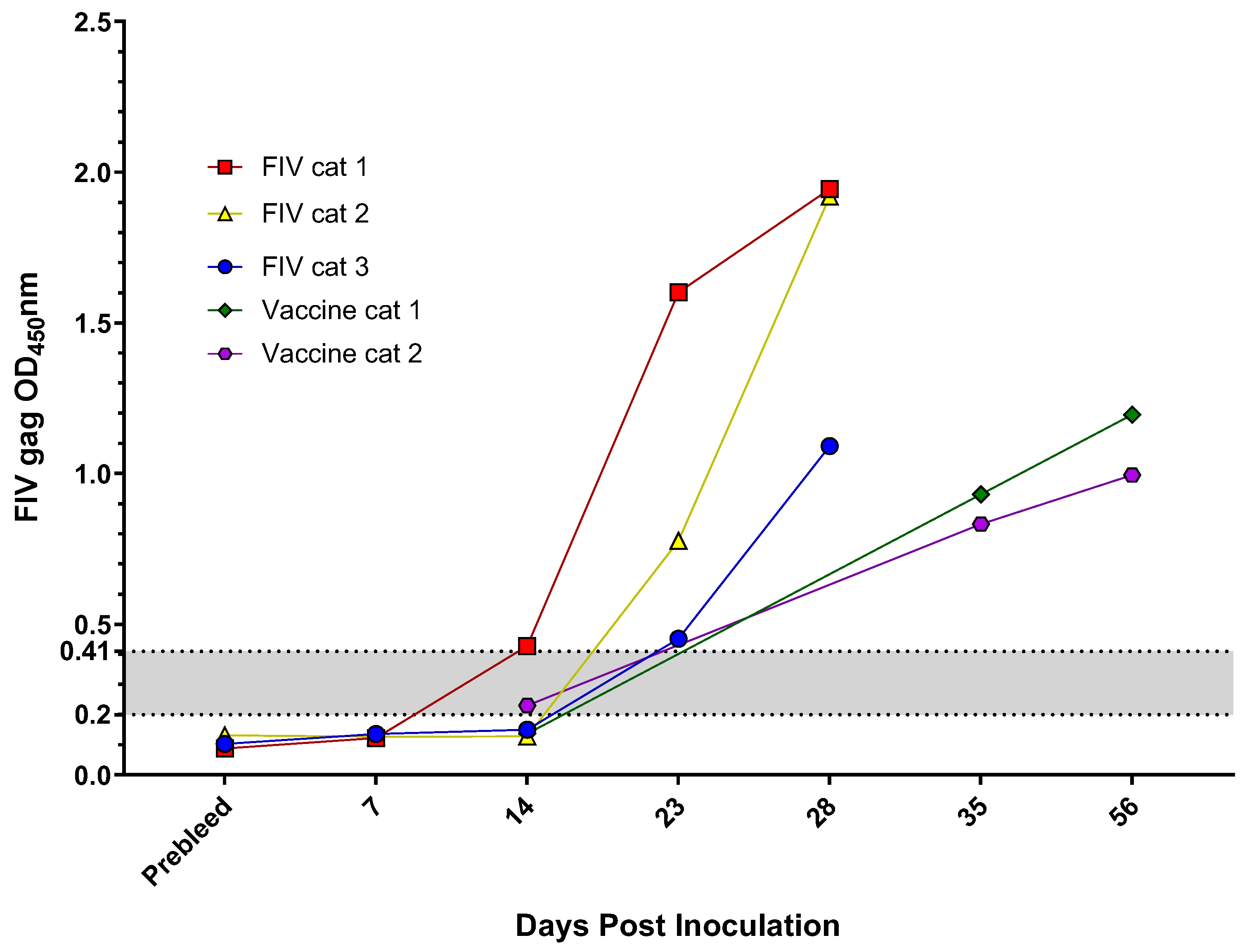
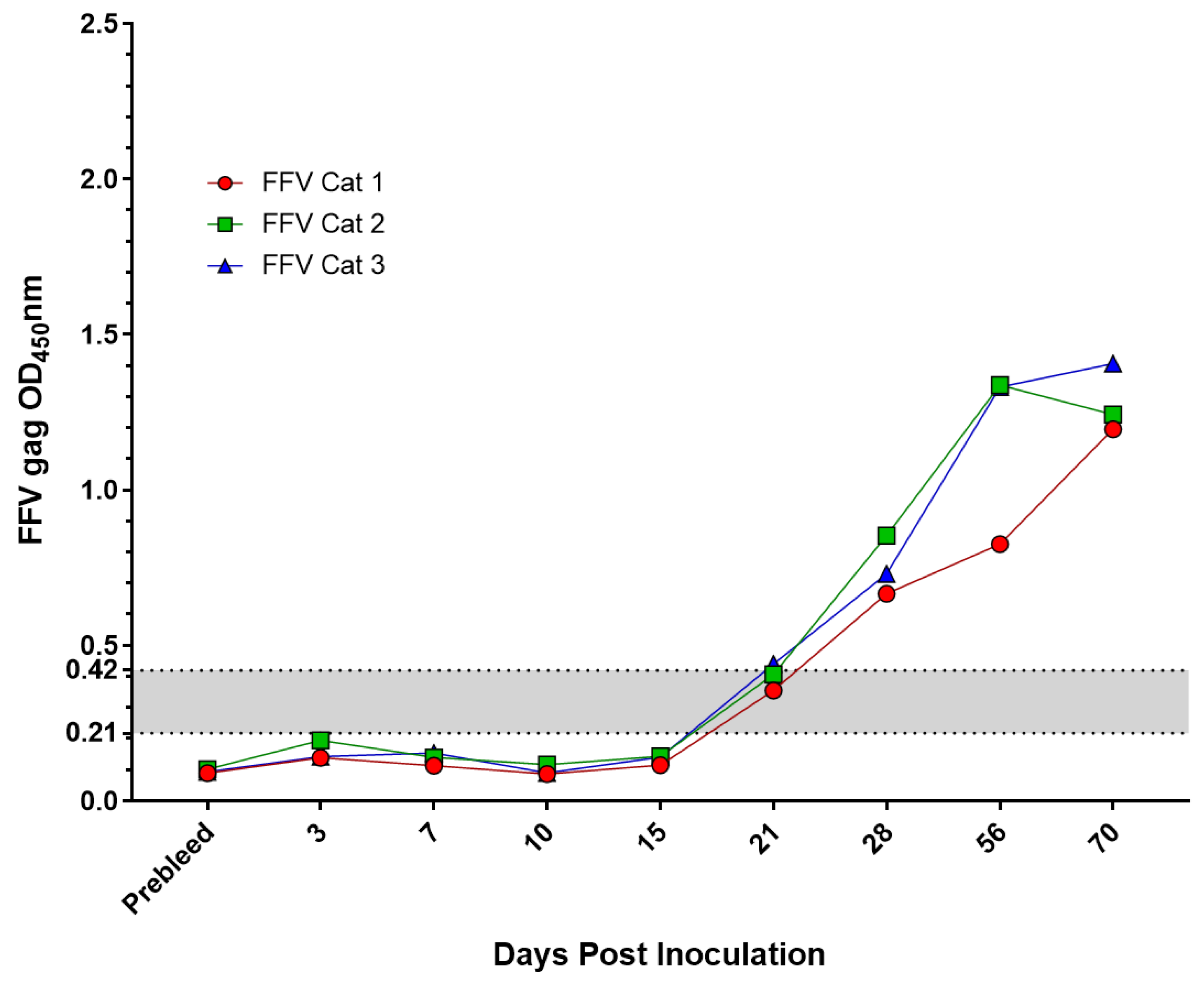
2.3. Experimentally Inoculated or Vaccinated FIV and FFV Results
2.4. Various Blocking Solutions Did Not Have an Effect on Nonspecific Binding in FIV ELISA
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. FIV and FFV Antigen Preparation
4.3. ELISA Protocol
4.4. Calculation of OD Indicating Seropositivity
4.5. Statistical Analysis
4.6. Blocking Experiments
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Güven, E.; Duus, K.; Lydolph, M.C.; Jørgensen, C.S.; Laursen, I.; Houen, G. Non-specific binding in solid phase immunoassays for autoantibodies correlates with inflammation markers. J. Immunol. Methods 2014, 403, 26–36. [Google Scholar] [CrossRef] [Green Version]
- Jacobson, R.H. Validation of serological assays for diagnosis of infectious diseases. Rev. Sci. Et Tech. (Int. Off. Epizoot.) 1998, 17, 469–526. [Google Scholar] [CrossRef]
- Miller, J.J.; Levinson, S.S. Interferences in immunoassays. In Immunoassay; Diamandis, E.P., Christopoulos, T.K., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 165–190. [Google Scholar]
- Schrijver, R.S.; Kramps, J.A. Critical factors affecting the diagnostic reliability of enzyme-linked immunosorbent assay formats. Rev. Sci. Et Tech. (Int. Off. Epizoot.) 1998, 17, 550–561. [Google Scholar] [CrossRef]
- Miller, C.; Abdo, Z.; Ericsson, A.; Elder, J.; VandeWoude, S. Applications of the FIV Model to Study HIV Pathogenesis. Viruses 2018, 10, 206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westman, M.E.; Malik, R.; Norris, J.M. Diagnosing feline immunodeficiency virus (FIV) and feline leukaemia virus (FeLV) infection: An update for clinicians. Aust. Vet. J. 2019, 97, 47–55. [Google Scholar] [CrossRef]
- Kechejian, S.; Dannemiller, N.; Kraberger, S.; Ledesma Feliciano, C.; Löchelt, M.; Carver, S.; VandeWoude, S. Feline foamy virus seroprevalence and demographic risk factors in stray domestic cat populations in Colorado, Southern California and Florida, USA. JFMS Open Rep. 2019, 5, 2055116919873736. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Feliciano, C.; Troyer, R.M.; Zheng, X.; Miller, C.; Cianciolo, R.; Bordicchia, M.; Dannemiller, N.; Gagne, R.; Beatty, J.; Quimby, J.; et al. Feline Foamy Virus Infection: Characterization of Experimental Infection and Prevalence of Natural Infection in Domestic Cats with and without Chronic Kidney Disease. Viruses 2019, 11, 662. [Google Scholar] [CrossRef] [Green Version]
- Sprague, W.S.; Troyer, R.M.; Zheng, X.; Wood, B.A.; Macmillan, M.; Carver, S.; VandeWoude, S. Prior Puma Lentivirus Infection Modifies Early Immune Responses and Attenuates Feline Immunodeficiency Virus Infection in Cats. Viruses 2018, 10, 210. [Google Scholar] [CrossRef] [Green Version]
- Ledesma-Feliciano, C.; Hagen, S.; Troyer, R.; Zheng, X.; Musselman, E.; Slavkovic Lukic, D.; Franke, A.-M.; Maeda, D.; Zielonka, J.; Münk, C.; et al. Replacement of feline foamy virus bet by feline immunodeficiency virus vif yields replicative virus with novel vaccine candidate potential. Retrovirology 2018, 15, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Bęczkowski, P.M.; Litster, A.; Lin, T.L.; Mellor, D.J.; Willett, B.J.; Hosie, M.J. Contrasting clinical outcomes in two cohorts of cats naturally infected with feline immunodeficiency virus (FIV). Vet. Microbiol. 2015, 176, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, J.A.; Chiu, E.S.; Kraberger, S.J.; Roelke-Parker, M.; Lowery, I.; Erbeck, K.; Troyer, R.; Carver, S.; VandeWoude, S. Feline Leukemia Virus (FeLV) Disease Outcomes in a Domestic Cat Breeding Colony: Relationship to Endogenous FeLV and Other Chronic Viral Infections. J. Virol. 2018, 92, e00649-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, K.; Griessmayr, P.; Schulz, B.; Greene, C.E.; Vidyashankar, A.N.; Jarrett, O.; Egberink, H.F. Quality of different in-clinic test systems for feline immunodeficiency virus and feline leukaemia virus infection. J. Feline Med. Surg. 2007, 9, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.N.; Keating, P.; Hosie, M.J.; Mackett, M.; Stephens, E.B.; Beatty, J.A.; Neil, J.C.; Jarrett, O. Env-specific CTL predominate in cats protected from feline immunodeficiency virus infection by vaccination. J. Immunol. 1996, 157, 3658–3665. [Google Scholar] [PubMed]
- Lecollinet, S.; Richardson, J. Vaccination against the feline immunodeficiency virus: The road not taken. Comp. Immunol. Microbiol. Infect. Dis. 2008, 31, 167–190. [Google Scholar] [CrossRef]
- Wood, B.A.; Carver, S.; Troyer, R.M.; Elder, J.H.; VandeWoude, S. Domestic cat microsphere immunoassays: Detection of antibodies during feline immunodeficiency virus infection. J. Immunol Methods 2013, 396, 74–86. [Google Scholar] [CrossRef] [Green Version]
- Fields, B.N.; Knipe, D.M.; Howley, P.M. Fields Virology, 6th ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013. [Google Scholar]
- German, A.C.; Harbour, D.A.; Helps, C.R.; Gruffydd-Jones, T.J. Is feline foamy virus really apathogenic? Vet. Immunol. Immunopathol. 2008, 123, 114–118. [Google Scholar] [CrossRef]
- Winkler, I.G.; Löchelt, M.; Flower, R.L. Epidemiology of feline foamy virus and feline immunodeficiency virus infections in domestic and feral cats: A seroepidemiological study. J. Clin. Microbiol. 1999, 37, 2848–2851. [Google Scholar] [CrossRef] [Green Version]
- Linial, M. Why aren’t foamy viruses pathogenic? Trends Microbiol. 2000, 8, 284–289. [Google Scholar] [CrossRef]
- Romen, F.; Pawlita, M.; Sehr, P.; Bachmann, S.; Schröder, J.; Lutz, H.; Löchelt, M. Antibodies against Gag are diagnostic markers for feline foamy virus infections while Env and Bet reactivity is undetectable in a substantial fraction of infected cats. Virology 2006, 345, 502–508. [Google Scholar] [CrossRef] [Green Version]
- Bleiholder, A.; Mühle, M.; Hechler, T.; Bevins, S.; vandeWoude, S.; Denner, J.; Löchelt, M. Pattern of seroreactivity against feline foamy virus proteins in domestic cats from Germany. Vet. Immunol Immunopathol 2011, 143, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Uhl, E.W.; Heaton-Jones, T.G.; Pu, R.; Yamamoto, J.K. FIV vaccine development and its importance to veterinary and human medicine: A review: FIV vaccine 2002 update and review. Vet. Immunol. Immunopathol. 2002, 90, 113–132. [Google Scholar] [CrossRef]
- Jacobson, R.H. Factors in selecting serum samples for use in determining the positive/negative threshold (cut-off) in ELISA. In Proceedings of the Final Research Co-Ordination Meeting on the Use of ELISA for Epidemiology and Control of Foot and Mouth Disease and Bovine Brucellosis in Latin America, Vienna, Austria, 14–18 April 1997. [Google Scholar]
- Zhang, L.; Zhang, J.J.; Kubiak, R.J.; Yang, H. Statistical methods and tool for cut point analysis in immunogenicity assays. J. Immunol. Methods 2013, 389, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Lardeux, F.; Torrico, G.; Aliaga, C. Calculation of the ELISA’s cut-off based on the change-point analysis method for detection of Trypanosoma cruzi infection in Bolivian dogs in the absence of controls. Mem. Do Inst. Oswaldo. Cruz. 2016, 111, 501–504. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Type | Sample Archive | Description | FIV ELISA | FFV ELISA | Reference (s) |
|---|---|---|---|---|---|
| Laboratory Cats | SPF FPV+ | Cats from a closed specific pathogen-free breeding colony naturally and inadvertently exposed to feline parvovirus. | 6 | 6 | [8,9,10] |
| SPF Sham | SPF cats in an FIV naïve control arm receiving culture supernatant from un-infected MYA-1 cells 34 days prior to sampling. SPF cats in the FFV naïve control arm received FFV-negative CrFK culture media 10 days prior to sampling. | 3 | 3 | [9,10] | |
| SPF FPV- | SPF cats prior to FPV exposure | 0 | 5 | [9,10] | |
| SPF FIV+ | SPF cats inoculated with 107.2 TCID50 FIV-C36. | 3 | 3 | [9] | |
| SPF-FIV-vx | SPF cats vaccinated with Fel-O-Vax (Boehringer Ingelheim, North Ryde, NSW, Australia) and boosted 2 weeks post prime. | 2 | 0 | Unpublished data | |
| SPF FFV+ | SPF cats vaccinated with 2.78 × 105 TCID50 FFV pCF-7. | 3 | 3 | [8,10] | |
| Field Cats | FIV shelter-adopted cats (FIV-sh) | Cats surrendered to an animal shelter which tested FIV-positive and were rehomed. | 9 | 18 | [11] |
| Private multi-cat household (MCH) | Cats homed in a large privately owned household. | 5 | 23 | [12] |
| ELISA | Sample Archive | Mean OD450 | OD450 range | Number of Samples in Indeterminant Range |
|---|---|---|---|---|
| FIV | SPF FPV+ cats | 0.095 | 0.046–0.16 | 0/6 |
| FIV | SPF Sham cats | 0.099 | 0.067–0.17 | 0/3 |
| FIV | Field cats | 0.23 | 0.103–0.39 | 10/15 (66.7%) |
| FFV | SPF FPV+ cats | 0.09 | 0.04–0.17 | 0/6 |
| FFV | SPF Sham cats | 0.08 | 0.06–0.12 | 0/3 |
| FFV | Field cats | 0.19 | 0.04–0.95 | 3/41(7.32%) |
| p-Value | ||
|---|---|---|
| FIV Sample Group | FFV Sample Group | |
| SPF vs Sham cats | 0.87 | 0.66 |
| SPF vs Field negative cats | <0.001 | 0.098 |
| Sham vs Field negative cats | <0.001 | 0.425 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moskaluk, A.; Nehring, M.; VandeWoude, S. Serum Samples from Co-Infected and Domestic Cat Field Isolates Nonspecifically Bind FIV and Other Antigens in Enzyme-Linked Immunosorbent Assays. Pathogens 2021, 10, 665. https://doi.org/10.3390/pathogens10060665
Moskaluk A, Nehring M, VandeWoude S. Serum Samples from Co-Infected and Domestic Cat Field Isolates Nonspecifically Bind FIV and Other Antigens in Enzyme-Linked Immunosorbent Assays. Pathogens. 2021; 10(6):665. https://doi.org/10.3390/pathogens10060665
Chicago/Turabian StyleMoskaluk, Alex, Mary Nehring, and Sue VandeWoude. 2021. "Serum Samples from Co-Infected and Domestic Cat Field Isolates Nonspecifically Bind FIV and Other Antigens in Enzyme-Linked Immunosorbent Assays" Pathogens 10, no. 6: 665. https://doi.org/10.3390/pathogens10060665