The T3SS Effector Protease NleC Is Active within Citrobacter rodentium

College of Veterinary Medicine, Kansas State University, Manhattan, KS 66506, USA
*
Author to whom correspondence should be addressed.
Pathogens 2021, 10(5), 589; https://doi.org/10.3390/pathogens10050589
Submission received: 15 April 2021
/
Revised: 6 May 2021
/
Accepted: 10 May 2021
/
Published: 12 May 2021
(This article belongs to the Section Bacterial Pathogens)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Whether type III secretion system (T3SS) effector proteins encoded by Gram-negative bacterial pathogens have intra-bacterial activities is an important and emerging area of investigation. Gram-negative bacteria interact with their mammalian hosts by using secretion systems to inject virulence proteins directly into infected host cells. Many of these injected protein effectors are enzymes that modify the structure and inhibit the function of mammalian proteins. The underlying dogma is that T3SS effectors are inactive until they are injected into host cells, where they then fold into their active conformations. We previously observed that the T3SS effectors NleB and SseK1 glycosylate Citrobacter rodentium and Salmonella enterica proteins, respectively, leading to enhanced resistance to environmental stress. Here, we sought to extend these studies to determine whether the T3SS effector protease NleC is also active within C. rodentium. To do this, we expressed the best-characterized mammalian substrate of NleC, the NF-κB p65 subunit in C. rodentium and monitored its proteolytic cleavage as a function of NleC activity. Intra-bacterial p65 cleavage was strictly dependent upon NleC. A p65 mutant lacking the known CE cleavage motif was resistant to NleC. Thus, we conclude that, in addition to NleB, NleC is also enzymatically active within C. rodentium.
1. Introduction
Gram-negative bacteria interact with host cells and inject virulence factors (effectors) through type III secretion system (T3SS) machineries [1]. Many Escherichia coli and Citrobacter rodentium T3SS effectors inhibit pro-inflammatory pathways, including NleB [2], NleH [3], EspL [4], NleD [5], and NleE [6].
NleC is a zinc metalloprotease T3SS effector that cleaves the NF-κB p65 subunit and thus inhibits p65 nuclear translocation. This dampens the host innate immune response, particularly the production of inflammatory cytokines [7,8,9,10,11]. NleC proteolytic activity is lost upon mutagenesis of the consensus zinc metalloprotease motif 183HEIIH187 [11]. Although NleC is secreted through the T3SS machinery, it also has the capacity to behave as a short-trip toxin and be internalized independently of the T3SS [12]. Other substrates of NleC other than p65 have been described and include the Rel family member p50 [8] and the acetyltransferase p300 [13]. It is thought that NleC specifically targets the NF-κB pathway instead of the MAPK pathway since it shows no activity against STAT or ERK [7,9].
Although two distinct p65 recognition and cleavage sites were initially described [7,11,14], it is now established that the cleavage site is C38/E39 [15,16]. NleC recognition of its substrates is also influenced by residues that are distant from the cleavage site [15,17].
The traditional view of T3SS effector secretion is that the effectors are inactive until injected into host cells, where they then fold into their active conformations and interact with host targets [18]. However, our recent work has challenged this dogma. We discovered that the T3SS effector NleB is active not only within the host, but also within C. rodentium. NleB activity with C. rodentium results in glycosylation of the glutathione synthetase (GshB) to enhance GshB activity and cause increased production of glutathione, leading to protection from oxidative stress [19]. We also observed that the NleB ortholog SseK1 is active within Salmonella enterica, where it enhances methylglyoxal detoxification [20]. As these data suggested that T3SS effectors may be active within the bacterium, here we desired to investigate the potential intra-bacterial effector of an effector with a different enzymatic activity. To do so, we investigated whether this phenomenon also applied to the C. rodentium zinc metalloprotease protease NleC.
2. Results
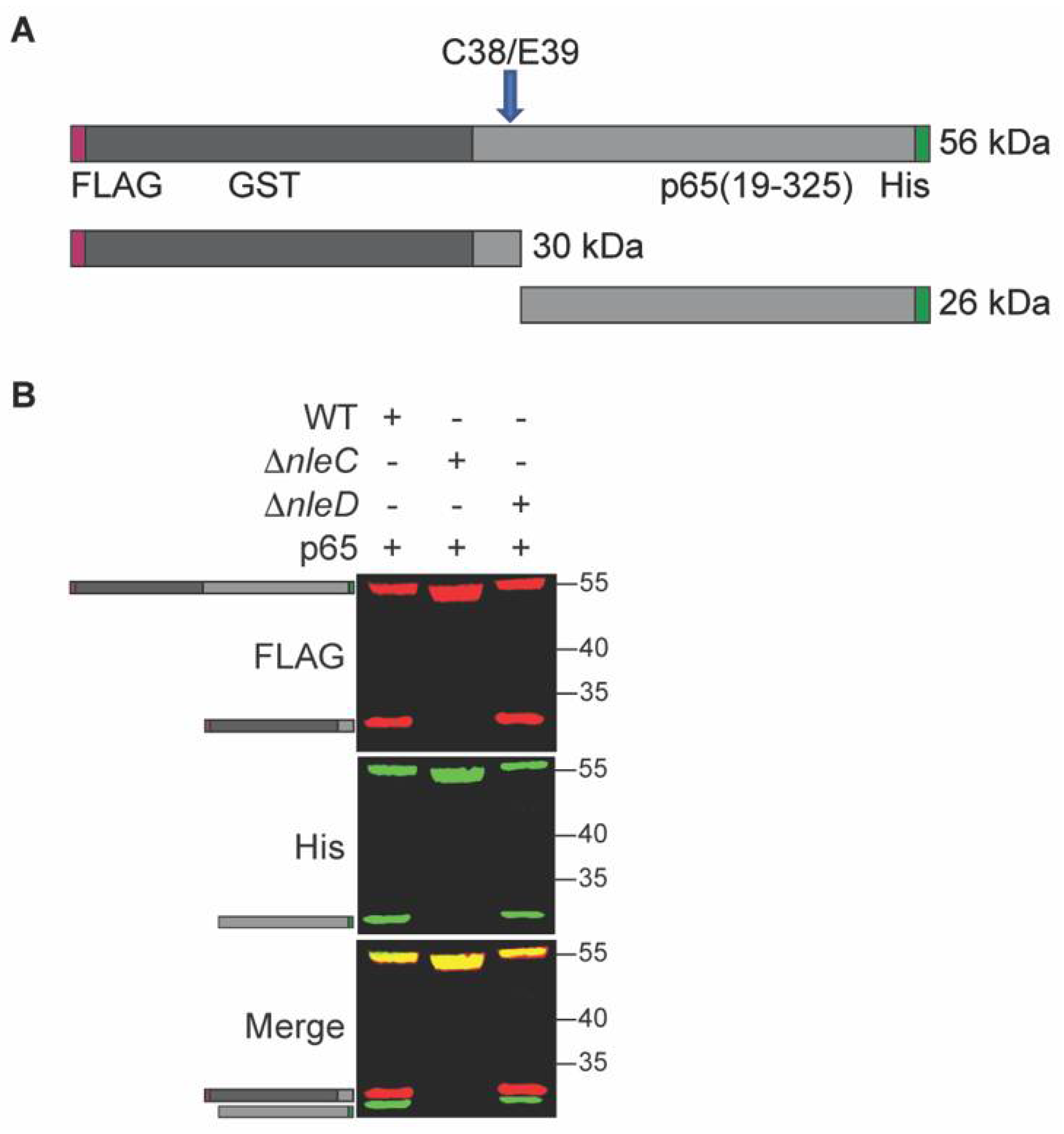
To investigate NleC activity inside C. rodentium, we cloned its known eukaryotic substrate p65 into a prokaryotic expression vector. Since the NleC cleavage site of p65 is near the N-terminus, we added a glutathione S-transferase (GST) tag to the N-terminus of p65 to increase the size of cleavage products. We also added a FLAG tag to the N-terminus to further aid detection of the cleavage product, as well as a C-terminal His-tag to aid with purification. Thus, the final recombinant p65 construct has a His-tag at the C-terminus and GST- and FLAG tags at the N-terminus. If cleavage by NleC occurs, the recombinant p65 protein is expected to be cleaved into two fragments: a 30 kDa N-terminal FLAG-tagged fragment and 26 kDa C-terminal His-tagged fragment (Figure 1A).
To investigate the NleC mediated cleavage of p65, p65 expression plasmids introduced into wild-type (WT) C. rodentium, or into deletion strains lacking NleC or NleD (ΔnleC), and (ΔnleD) strains. We observed that p65 was cleaved into the two expected fragments of appropriate molecular weights in the WT strain, but no such cleavage products were seen in the ΔnleC strain. Additionally, the same cleavage products were also seen in the ΔnleD strain, a strain lacking the T3SS effector NleD, a zinc metalloprotease that cleaves JNK [5] (Figure 1B). Thus, p65 cleavage within C. rodentium appears to be dependent upon NleC.
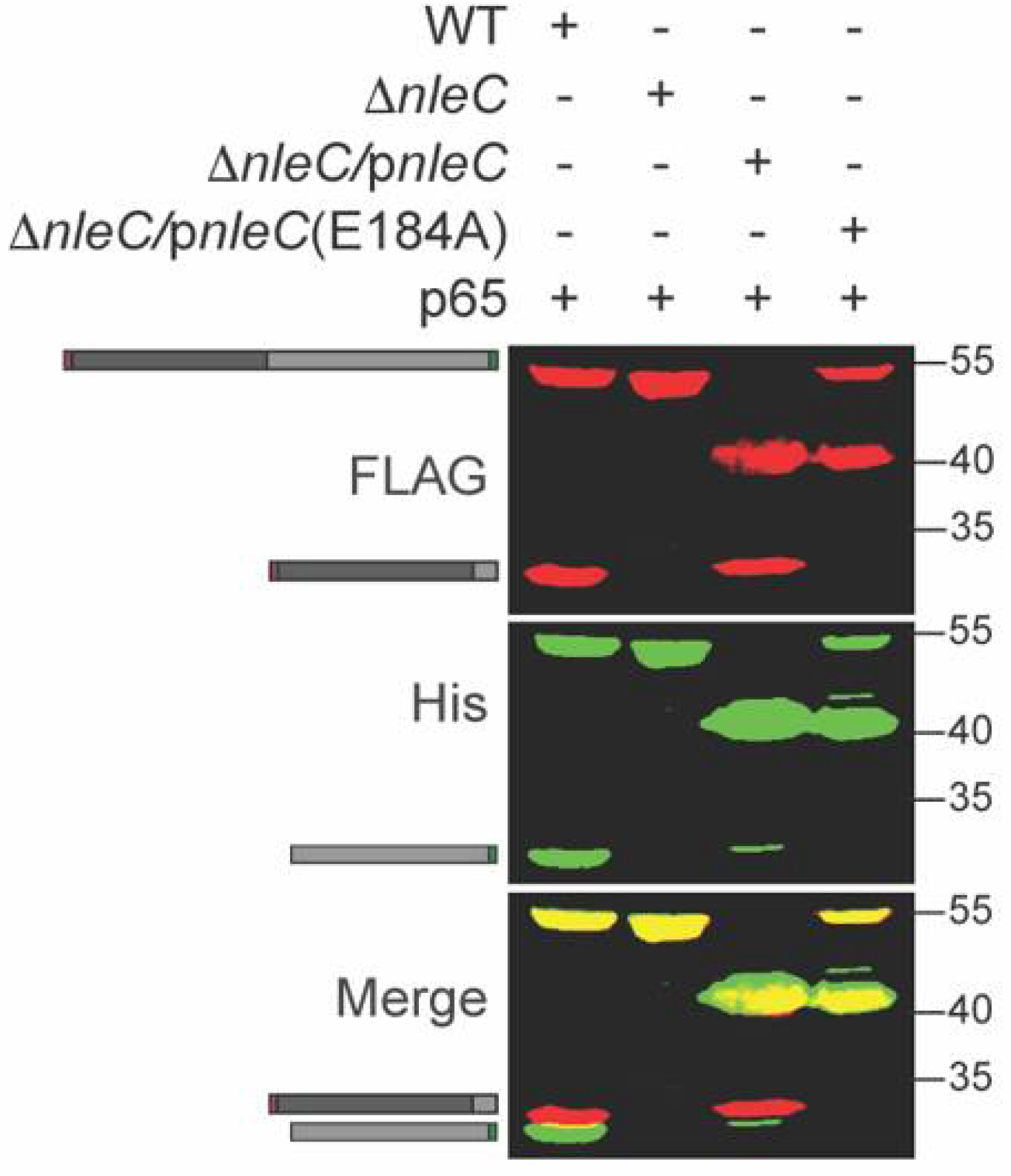
NleC enzymatic activity is significantly reduced in the host cell when the critical NleC residue E184 around the Zn2+ binding pocket is mutated to A184 [7]. To further evaluate p65 cleavage specificity and its dependence upon NleC activity, we co-transformed ΔnleC C. rodentium with a p65 expression plasmid and either WT NleC or NleC E184A complementation plasmids. The recombinant p65 was cleaved by WT NleC but not by NleC E184A (Figure 2).
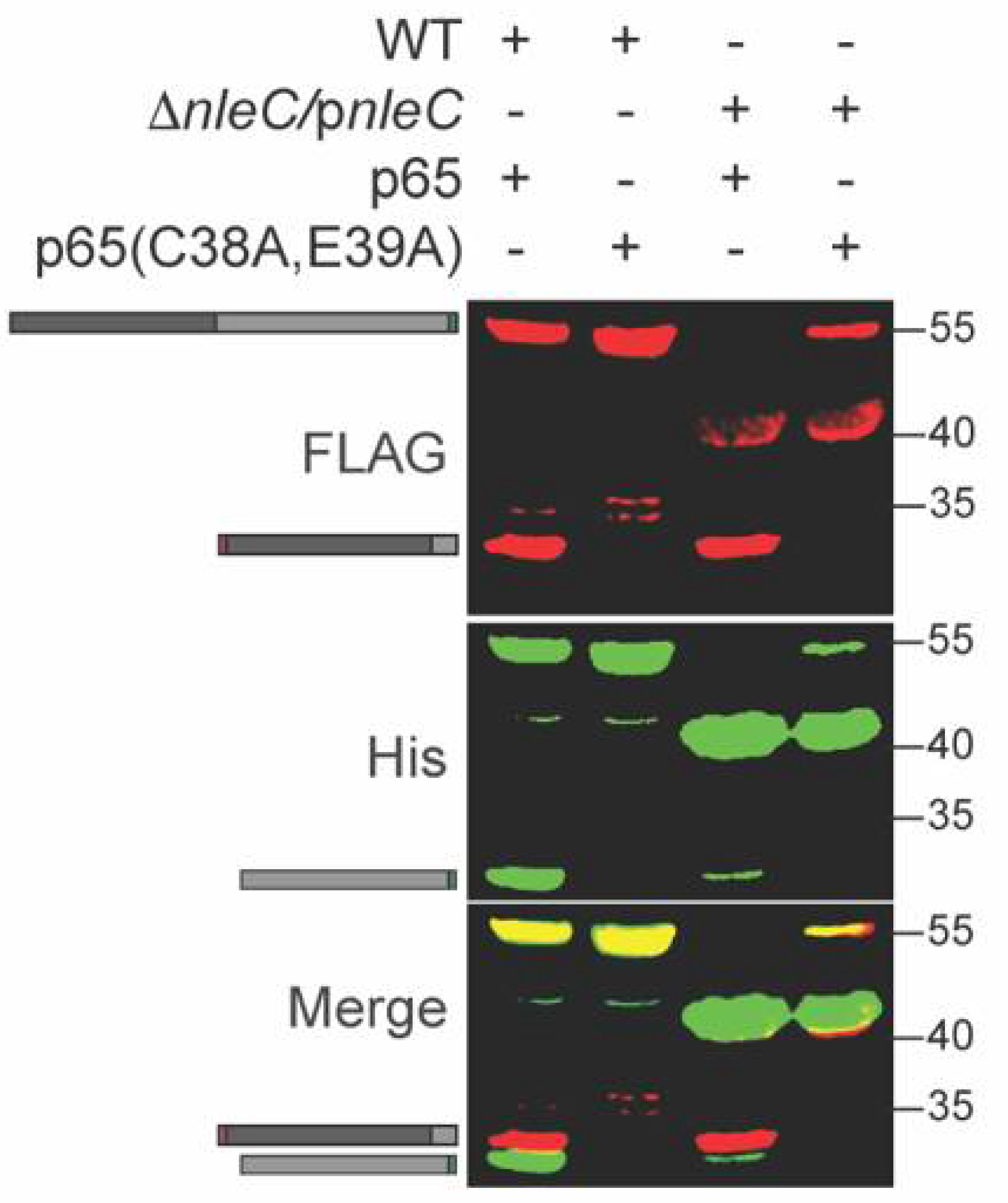
p65 is cleaved by NleC between C38 and E39 [15]. To determine whether this cleavage specificity was retained within C. rodentium, we mutated the p65 expression plasmid to change C38/E39 to alanine residues. We expressed this plasmid in WT C. rodentium and observed that the p65 C38A, E39A mutant was no longer cleaved by NleC (Figure 3). In addition, we also purified the His-tagged (C-terminal) fragment of the cleavage product (Figure 3, lane 1) and performed Edman degradation analyses. The five N-terminal amino acids we identified corresponded to the expected sequence (EGRSA) of the C-terminal cleavage product (data not shown). Thus, we concluded that NleC is active within C. rodentium.
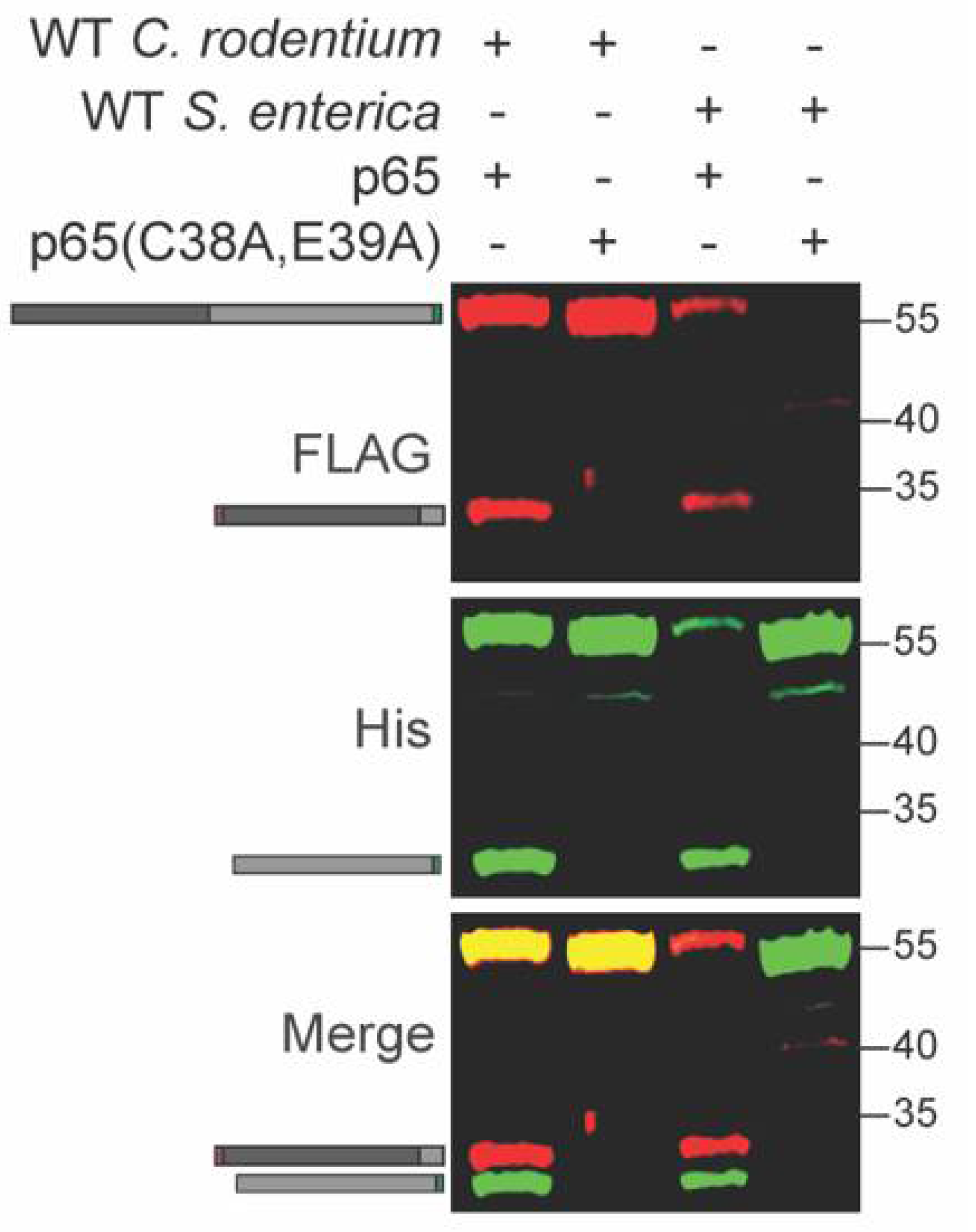
Salmonella enterica also induces p65 cleavage in host cells [21,22]. We tested whether p65 is cleaved inside S. enterica in a similar fashion to C. rodentium. We expressed the recombinant WT and C38A/E39A p65 constructs in S. enterica and observed cleavage of only the WT p65 construct, similarly to what was observed for C. rodentium (Figure 4 and Table S1). S. enterica harbors at least three distinct T3SS effectors, GogA, PipA, and GtgA, that cleave p65 inside host cells [22]. At least one of these effectors may be active within S. enterica.
3. Discussion
NleC is a zinc metalloprotease that cleaves the NF-κB p65 subunit and dampens the immune response of the host cell. Despite the traditional view of T3SS effectors being active only after their translocation into the host, here we demonstrated that the C. rodentium T3SS effector NleC is active inside the bacterium. The implication of this novel finding is far-reaching. As demonstrated by our previous findings of the C. rodentium T3SS effector NleB glycosylating the bacterial substrate GshB and therefore contributing to the increased survival of C. rodentium under environmental stress [19], as well as S. enterica SseK1 glycosylating multiple bacterial substrates involved in methylglyoxal (MGO) detoxification [20], NleC may have its own endogenous bacterial targets. Compared with Locus of Enterocyte Effacement (LEE)-encoded effectors, non-LEE-encoded effectors such as NleB and NleC are potentially more recent acquisitions to the pathogens and their roles as effectors are still evolving [23]. This indicates that non-LEE effectors potentially have a dual role in modulating both pathogen and host cellular pathways. Therefore, NleC activity in C. rodentium could play a significant role in modulating bacterial regulation of virulence, response to environmental stress, and immune evasion.
The traditional model of T3SS effector translocation is that the effectors are chaperoned within the bacterium until they are secreted. Indeed, chaperones for multiple effectors have been identified and characterized [24,25,26,27,28]. To our knowledge, no chaperone for NleC has been identified, indicating that NleC might not be chaperoned before its secretion, potentially explaining its activity inside the bacteria. Similarly, no chaperone has been identified for either NleB1 [19] or SseK1.
The potential intra-bacterial targets of NleC are currently unknown, and this absence of knowledge potentially limits the biological impact of our findings. However, some insight could be obtained from bioinformatics analyses. For example, we performed an initial bioinformatics-based search for bacterial NleC substrates by using the known p65 cleavage site as a query. One potential hit is GrlR, a regulator of LEE expression [29] that harbors both the conserved NleC cleavage site and a DNA-binding domain. In theory, NleC could cleave GrlR and thus relieve the repression of LEE expression, which in turn could increase the expression and/or secretion of NleC and other effectors. Another study found that NleC recognizes and bind its target protein by DNA mimicry, which indicates that some other targets of NleC may be proteins with nucleic acid binding capacity [16]. NleC may thus indirectly regulate transcription and affect bacterial responses to environmental changes. Proteomic techniques such as Terminal Amine Isotopic labeling of Substrates (TAILS) [30] might also reasonably be employed to identify proteins whose abundance is dependent upon NleC activity.
4. Materials and Methods
4.1. Strains and Molecular Cloning
Plasmids were constructed in pET15 and pET28 backgrounds by using ABC cloning [31]. p65 and NleC point mutants were created by using site-directed mutagenesis. C. rodentium ΔnleC and ΔnleD strains were constructed by using lambda red recombination [32] with pKD3 and pKD119 plasmids. A recombination cassette harboring a selectable marker flanked by ~50 bp of complementary sequences from the 5′ and 3′ ends of the gene to be deleted was created by using PCR with pKD3 as a template. The nleC recombination cassette was amplified by using forward (5′-GCTCCCATTGCTCCTAATCGTGCTGAAAATGCCTATGCGGATGAAATCTAACAATGCGCT) and reverse (5′-GCCAATCCAGGAAAATCATGTTGCTGGATAAATCCGTATCGGTCGAGGTGGCCCGGCTCC) primers. The nleD recombination cassette was amplified by using forward (5′-GCTCAATGTCAGATACAGATATCGAGTCTCTTGTAAAAGCAAGCCACTGGAGCACCTCAA) and reverse (5′-CGGGTAGGAGGTTCTACGGGGCATCCCAATCTCTTCGTGGACGGGGAGAGCCTGAGCAAA) primers. Plasmids were introduced into C. rodentium and S. enterica strains by using electroporation.
4.2. Western Blotting
Strains were grown in LB media with appropriate antibiotics (kanamycin and/or carbenicillin) for 4 h at 37 °C and then induced with 0.5 mM IPTG with growth for an additional 4 h at 30 °C. Cell lysates were derived from 1 mL of bacterial cultures via centrifugation and lysed in 100 µL SDS loading buffer by boiling for 5 min. Samples (10 µL) were used in Western blot assays with anti-His (1:1000 dilution; H1029, Sigma-Aldrich, St. Louis, SL, USA) and anti-FLAG (1:5000 dilution; F7425, Millipore, Burlington, USA) primary antibodies. IRDye 800CW goat-anti-mouse IgG (926-32210, LI-COR) and IRDye 680RD goat-anti-rabbit IgG (926-68071, LI-COR) secondary antibodies were used at 1:10,000 dilutions.
4.3. Edman Degradation
The C-terminal p65 cleavage product generated from WT C. rodentium was purified using standard His-tagged protein purification methods and processed according to the sample preparation guidelines for Edman degradation by the Protein Facility of the Iowa State University Office of Biotechnology.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/pathogens10050589/s1, Table S1: Edman degradation analysis of p65 C-terminal cleavage product.
Author Contributions
Conceptualization, P.R.H.; methodology, M.K.H. and S.E.Q.; investigation, M.K.H. and S.E.Q.; writing—original draft preparation, M.K.H.; writing—review and editing, P.R.H.; funding acquisition, P.R.H. All authors have read and agreed to the published version of the manuscript.
Funding
The described project was supported by grant number GM130448 from the National Institute of General Medical Sciences (NIGMS) of the National Institutes of Health. Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NIGMS.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and supplemental file.
Acknowledgments
We thank the Protein Facility of the Iowa State University Office of Biotechnology for performing Edman degradation assays.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Büttner, D. Protein Export According to Schedule: Architecture, Assembly, and Regulation of Type III Secretion Systems from Plant- and Animal-Pathogenic Bacteria. Microbiol. Mol. Biol. Rev. 2012, 76, 262–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, J.S.; Giogha, C.; Ong, S.Y.; Kennedy, C.L.; Kelly, M.; Robinson, K.S.; Lung, T.W.F.; Mansell, A.; Riedmaier, P.; Oates, C.V.L.; et al. A type III effector antagonizes death receptor signalling during bacterial gut infection. Nat. Cell Biol. 2013, 501, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wan, F.; Mateo, K.; Callegari, E.; Wang, D.; Deng, W.; Puente, J.; Li, F.; Finlay, B.B.; Lenardo, M.J.; et al. Bacterial effector binding to ribosomal protein s3 subverts NF-kappaB function. PLoS Pathog. 2009, 5, e1000708. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.S.; Giogha, C.; Mühlen, S.; Nachbur, U.; Pham, C.L.L.; Zhang, Y.; Hildebrand, J.M.; Oates, C.V.; Lung, T.W.F.; Ingle, D.; et al. EspL is a bacterial cysteine protease effector that cleaves RHIM proteins to block necroptosis and inflammation. Nat. Microbiol. 2017, 2, 16258. [Google Scholar] [CrossRef]
- Creuzburg, K.; Giogha, C.; Lung, T.W.F.; Scott, N.E.; Mühlen, S.; Hartland, E.L.; Pearson, J.S. The Type III Effector NleD from Enteropathogenic Escherichia coli Differentiates between Host Substrates p38 and JNK. Infect. Immun. 2016, 85. [Google Scholar] [CrossRef] [Green Version]
- Nadler, C.; Baruch, K.; Kobi, S.; Mills, E.; Haviv, G.; Farago, M.; Alkalay, I.; Bartfeld, S.; Meyer, T.F.; Ben-Neriah, Y.; et al. The type III secretion effector NleE inhibits NF-kappaB activation. PLoS Pathog. 2010, 6, e1000743. [Google Scholar] [CrossRef] [Green Version]
- Baruch, K.; Gur-Arie, L.; Nadler, C.; Koby, S.; Yerushalmi, G.; Ben-Neriah, Y.; Yogev, O.; Shaulian, E.; Guttman, C.; Zarivach, R.; et al. Metalloprotease type III effectors that specifically cleave JNK and NF-kappaB. EMBO J. 2011, 30, 221–231. [Google Scholar] [CrossRef]
- Muhlen, S.; Ruchaud-Sparagano, M.H.; Kenny, B. Proteasome-independent degradation of canonical NFkappaB complex components by the NleC protein of pathogenic Escherichia coli. J. Biol. Chem. 2011, 286, 5100–5107. [Google Scholar] [CrossRef] [Green Version]
- Pearson, J.S.; Riedmaier, P.; Marches, O.; Frankel, G.; Hartland, E.L. A type III effector protease NleC from enteropathogenic Escherichia coli targets NF-kappaB for degradation. Mol. Microbiol. 2011, 80, 219–230. [Google Scholar] [CrossRef] [Green Version]
- Sham, H.P.; Shames, S.R.; Croxen, M.A.; Ma, C.; Chan, J.M.; Khan, M.A.; Wickham, M.E.; Deng, W.; Finlay, B.B.; Vallance, B.A. Attaching and effacing bacterial effector NleC suppresses epithelial inflammatory responses by inhibiting NF-kappaB and p38 mitogen-activated protein kinase activation. Infect Immun. 2011, 79, 3552–3562. [Google Scholar] [CrossRef] [Green Version]
- Yen, H.; Ooka, T.; Iguchi, A.; Hayashi, T.; Sugimoto, N.; Tobe, T. NleC, a type III secretion protease, compromises NF-kappaB activation by targeting p65/RelA. PLoS Pathog. 2010, 6, e1001231. [Google Scholar] [CrossRef] [Green Version]
- Stolle, A.S.; Norkowski, S.; Körner, B.; Schmitz, J.; Lüken, L.; Frankenberg, M.; Rüter, C.; Schmidt, M.A. T3SS-Independent Uptake of the Short-Trip Toxin-Related Recombinant NleC Effector of Enteropathogenic Escherichia coli Leads to NF-kappaB p65 Cleavage. Front Cell Infect Microbiol. 2017, 7, 119. [Google Scholar] [CrossRef]
- Shames, S.R.; Bhavsar, A.P.; Croxen, M.A.; Law, R.J.; Mak, S.H.; Deng, W.; Li, Y.; Bidshari, R.; de Hoog, C.L.; Foster, L.J. The pathogenic Escherichia coli type III secreted protease NleC degrades the host acetyltransferase p300. Cell Microbiol. 2011, 13, 1542–1557. [Google Scholar] [CrossRef]
- Li, W.; Liu, Y.; Sheng, X.; Yin, P.; Hu, F.; Liu, Y.; Chen, C.; Li, Q.; Yan, C.; Wang, J. Structure and mechanism of a type III secretion protease, NleC. Acta Crystallogr. Sect. D Biol. Crystallogr. 2013, 70, 40–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodgson, A.; Wier, E.M.; Fu, K.; Sun, X.; Yu, H.; Zheng, W.; Sham, H.P.; Johnson, K.; Bailey, S.; Vallance, B.A.; et al. Metalloprotease NleC Suppresses Host NF-κB/Inflammatory Responses by Cleaving p65 and Interfering with the p65/RPS3 Interaction. PLOS Pathog. 2015, 11, e1004705. [Google Scholar] [CrossRef] [PubMed]
- Turco, M.M.; Sousa, M.C. The Structure and Specificity of the Type III Secretion System Effector NleC Suggest a DNA Mimicry Mechanism of Substrate Recognition. Biochemistry 2014, 53, 5131–5139. [Google Scholar] [CrossRef] [PubMed]
- Giogha, C.; Lung, T.W.F.; Mühlen, S.; Pearson, J.S.; Hartland, E.L. Substrate recognition by the zinc metalloprotease effector NleC from enteropathogenicEscherichia coli. Cell. Microbiol. 2015, 17, 1766–1778. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.F.; Cornelis, G.R. The multitalented type III chaperones: All you can do with 15 kDa. FEMS Microbiol. Lett. 2003, 219, 151–158. [Google Scholar] [CrossRef] [Green Version]
- El Qaidi, S.; Scott, N.E.; Hays, M.P.; Geisbrecht, B.V.; Watkins, S.; Hardwidge, P.R. An intra-bacterial activity for a T3SS effector. Sci. Rep. 2020, 10, 1073. [Google Scholar] [CrossRef] [Green Version]
- El Qaidi, S.; Scott, N.E.; Hardwidge, P.R. Arginine glycosylation enhances methylglyoxal detoxification. Sci. Rep. 2021, 11, 3834. [Google Scholar] [CrossRef]
- Jennings, E.; Esposito, D.; Rittinger, K.; Thurston, T.L.M. Structure-function analyses of the bacterial zinc metalloprotease effector protein GtgA uncover key residues required for deactivating NF-kappaB. J. Biol. Chem. 2018, 293, 15316–15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Kamanova, J.; Lara-Tejero, M.; Galan, J.E. A Family of Salmonella Type III Secretion Effector Proteins Selectively Targets the NF-kappaB Signaling Pathway to Preserve Host Homeostasis. PLoS Pathog. 2016, 12, e1005484. [Google Scholar] [CrossRef]
- Eswarappa, S.M.; Janice, J.; Balasundaram, S.V.; Chakravortty, D. Non-neutral evolution in non-LEE-encoded type III effectors of attaching and effacing Escherichia coli. Microbes Infect. 2013, 15, 147–151. [Google Scholar] [CrossRef]
- Abe, A.; De Grado, M.; Pfuetzner, R.A.; Sánchez-SanMartín, C.; DeVinney, R.; Puente, J.L.; Strynadka, N.C.; Finlay, B.B. Enteropathogenic Escherichia coli translocated intimin receptor, Tir, requires a specific chaperone for stable secretion. Mol. Microbiol. 1999, 33, 1162–1175. [Google Scholar] [CrossRef]
- Bronstein, P.A.; Miao, E.A.; Miller, S.I. InvB Is a Type III Secretion Chaperone Specific for SspA. J. Bacteriol. 2000, 182, 6638–6644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, C.A.; Zhang, K.; Andres, S.N.; Fang, Y.; Kaniuk, N.A.; Hannemann, M.; Brumell, J.H.; Foster, L.J.; Junop, M.S.; Coombes, B.K. Structural and Biochemical Characterization of SrcA, a Multi-Cargo Type III Secretion Chaperone in Salmonella Required for Pathogenic Association with a Host. PLoS Pathog. 2010, 6, e1000751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Page, A.-L.; Sansonetti, P.; Parsot, C. Spa15 of Shigella flexneri, a third type of chaperone in the type III secretion pathway. Mol. Microbiol. 2002, 43, 1533–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spaeth, K.E.; Chen, Y.S.; Valdivia, R.H. The Chlamydia type III secretion system C-ring engages a chaperone-effector protein complex. PLoS Pathog. 2009, 5, e1000579. [Google Scholar] [CrossRef]
- Abé, H.; Miyahara, A.; Oshima, T.; Tashiro, K.; Ogura, Y.; Kuhara, S.; Ogasawara, N.; Hayashi, T.; Tobe, T. Global Regulation by Horizontally Transferred Regulators Establishes the Pathogenicity of Escherichia coli. DNA Res. 2008, 15, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Kleifeld, O.; Doucet, A.; Keller, U.A.D.; Prudova, A.; Schilling, O.; Kainthan, R.K.; Starr, A.E.; Foster, L.J.; Kizhakkedathu, J.N.; Overall, C.M. Isotopic labeling of terminal amines in complex samples identifies protein N-termini and protease cleavage products. Nat. Biotechnol. 2010, 28, 281–288. [Google Scholar] [CrossRef]
- Qaidi, S.E.; Hardwidge, P.R. ABC cloning: An efficient, simple, and rapid restriction/ligase-free method. MethodsX 2019, 6, 316–321. [Google Scholar] [CrossRef] [PubMed]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
NleC cleaves p65 in C. rodentium. (A). Schematic. p65 (residues 19-325) was cloned and expressed as a recombinant fusion to N-terminal FLAG (red)- and GST (grey)-tags and a C-terminal His-tag (green). The cleavage site is indicated with an arrow; the N-terminal and C-terminal cleavage products with epitope tags and their expected sizes are also shown. (B). p65 cleavage is dependent upon NleC. p65 was expressed in WT, ΔnleC, or ΔnleD C. rodentium. Protein lysates were analyzed using Western blotting. FLAG, His, and merged blot images are shown. Cartoons indicate the expected full-length and cleavage products as depicted in panel A.
Figure 1.
NleC cleaves p65 in C. rodentium. (A). Schematic. p65 (residues 19-325) was cloned and expressed as a recombinant fusion to N-terminal FLAG (red)- and GST (grey)-tags and a C-terminal His-tag (green). The cleavage site is indicated with an arrow; the N-terminal and C-terminal cleavage products with epitope tags and their expected sizes are also shown. (B). p65 cleavage is dependent upon NleC. p65 was expressed in WT, ΔnleC, or ΔnleD C. rodentium. Protein lysates were analyzed using Western blotting. FLAG, His, and merged blot images are shown. Cartoons indicate the expected full-length and cleavage products as depicted in panel A.

Figure 2.
Inactive NleC does not cleave p65. p65 was expressed in WT, ΔnleC, or ΔnleC complemented with either WT NleC or NleC E184A C. rodentium. Experiments were conducted as described in Figure 1 panel B. The 40 kDa protein band is the NleC protein produced via plasmid complementation.
Figure 2.
Inactive NleC does not cleave p65. p65 was expressed in WT, ΔnleC, or ΔnleC complemented with either WT NleC or NleC E184A C. rodentium. Experiments were conducted as described in Figure 1 panel B. The 40 kDa protein band is the NleC protein produced via plasmid complementation.

Figure 3.
NleC does not cleave the p65 (C38A/E39A) mutant. WT and mutant p65 were expressed in WT C. rodentium. Experiments were conducted as described in Figure 1 panel B.
Figure 3.
NleC does not cleave the p65 (C38A/E39A) mutant. WT and mutant p65 were expressed in WT C. rodentium. Experiments were conducted as described in Figure 1 panel B.

Figure 4.
p65 cleavage occurs within S. enterica. WT and mutant p65 were expressed in WT S. enterica. Experiments were conducted as described in Figure 1 panel B.
Figure 4.
p65 cleavage occurs within S. enterica. WT and mutant p65 were expressed in WT S. enterica. Experiments were conducted as described in Figure 1 panel B.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hasan, M.K.; El Qaidi, S.; Hardwidge, P.R. The T3SS Effector Protease NleC Is Active within Citrobacter rodentium. Pathogens 2021, 10, 589. https://doi.org/10.3390/pathogens10050589
AMA Style
Hasan MK, El Qaidi S, Hardwidge PR. The T3SS Effector Protease NleC Is Active within Citrobacter rodentium. Pathogens. 2021; 10(5):589. https://doi.org/10.3390/pathogens10050589
Chicago/Turabian StyleHasan, Md Kamrul, Samir El Qaidi, and Philip R. Hardwidge. 2021. "The T3SS Effector Protease NleC Is Active within Citrobacter rodentium" Pathogens 10, no. 5: 589. https://doi.org/10.3390/pathogens10050589
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.