

Oxidative Stress as an Underlying Mechanism of Bacteria-Inflicted Damage to Male Gametes

1
Institute of Biotechnology, Faculty of Biotechnology and Food Sciences, Slovak University of Agriculture in Nitra, Tr. A. Hlinku 2, 94976 Nitra, Slovakia
2
Institute of Applied Biology, Faculty of Biotechnology and Food Sciences, Slovak University of Agriculture in Nitra, Tr. A. Hlinku 2, 94976 Nitra, Slovakia
3
AgroBioTech Research Centre, Slovak University of Agriculture in Nitra, Tr. A. Hlinku 2, 94976 Nitra, Slovakia
*
Author to whom correspondence should be addressed.
Oxygen 2022, 2(4), 547-569; https://doi.org/10.3390/oxygen2040036
Submission received: 7 October 2022
/
Revised: 31 October 2022
/
Accepted: 3 November 2022
/
Published: 6 November 2022
(This article belongs to the Special Issue Review Papers in Oxygen)
Abstract
:Bacterial infestation of the male reproductive system with subsequent effects of bacteria on the structural integrity and functional activity of male gametes has become a significant factor in the etiology of male reproductive dysfunction. Bacteria may affect male fertility either by directly interacting with structures critical for sperm survival or indirectly by triggering a local immune response, leukocytospermia or reactive oxygen species (ROS) overproduction followed by oxidative stress development. This review aims to provide an overview of the currently available knowledge on bacteriospermia-associated sperm damage with a special emphasis on oxidative mechanisms underlying sperm deterioration caused by bacterial action. At the same time, we strive to summarize readily available alternatives to prevent or counteract alterations to spermatozoa caused by bacterial colonization of semen or by oxidative stress as an accompanying phenomenon of bacteriospermia.
1. Introduction
A global challenge of today’s era lies in the understanding, prevention, and management of ever-increasing male reproductive dysfunction. Particularly in mammals, male sub- or infertility is a complex and multifactorial condition that may be caused by a multitude of factors, ranging from genetic causes and systemic ailments to environmental exposure or nutrition [1]. Within this vast array of etiologies, bacteriospermia—the presence of bacteria in semen—is thought to be the causative agent in around 15% of all cases of subfertile males [1,2]. In clinical practice, bacteriospermia is acknowledged when the bacterial count exceeds 1.000 colony-forming units (CFU)/mL of semen [3]. The primary reason for bacterial colonization of ejaculates lies in an acute or chronic infection of the urogenital tract including diseases such as urethritis, prostatitis, epididymitis or orchitis, subsequently compromising proper spermatogenesis and sperm maturation [4]. Alternatively, bacteria may invade the reproductive system from urinary tract infection [5], by hematogenous dissemination [6] or by sexual intercourse [7]. In certain cases, bacteriospermia may be caused by the overgrowth of natural microflora present in the urogenital tract even in clinically healthy subjects [8,9]. Semen may be also infested by bacteria from external sources, such as skin, urine, feces [10] as well as laboratory tools, equipment or media needed for semen collection and/or processing for assisted reproduction techniques (ARTs) [11].
Bacteria present in semen may affect male gametes through adhesion [12] and agglutination events, which will ultimately result in sperm immobilization and inability to reach and fertilize the oocyte [13]. The released bacterial toxins may have direct cytotoxic effects on the male gamete, leading to increased sperm cell death [14]. Furthermore, bacterial infection may trigger persistent inflammation and leukocytic infiltration accompanied by the release of pro-inflammatory cytokines that will contribute to permanent damage of structures vital for sperm survival [9,15].
While the mechanisms of which bacteria may cause alterations to male reproductive cells are diverse, a convincing body of evidence indicates a notable involvement of reactive oxygen species (ROS) in the phenomena associated with bacteriospermia. As such, understanding the role of oxidative processes underlying bacteria-inflicted damage to spermatozoa may assist in the development of adequate intervention strategies to mitigate bacteriospermia-associated male subfertility.
2. Sources of ROS in Bacteriospermia
The involvement of ROS in male reproduction with beneficial as well as adverse effects has become indisputable in modern andrology. Carefully controlled physiological ROS levels play a crucial role in signaling pathways involved in sperm production and maturation, as well as during capacitation, hyperactivation, acrosome reaction and sperm-egg fusion. Nevertheless, ROS accumulation has been previously acknowledged to act either as a primary cause or an accompanying phenomenon in a vast array of male reproductive pathophysiologies [16,17,18,19]. Correspondingly, earlier reports on bacteriospermia have frequently observed increased oxidative damage resulting from a shift in the prooxidant–antioxidant milieu in favor of prooxidants, favoring further ROS release and action [20,21,22,23].
By and large, three sources of ROS are most relevant to bacteriospermia: (1) immature, damaged or dead spermatozoa, (2) bacterial metabolism, and (3) activated leukocytes.
2.1. Spermatozoa
The process of sperm maturation involves a series of events that take place during the sperm transition from the seminiferous epithelium to the epididymis. These processes include structural remodeling, male germ cell differentiation, acquisition of the ability to move and to accomplish acrosome reaction, all of which are dependent on changes in the architecture and fluidity of the plasma membrane [24]. In morphologically normal spermatozoa, cytoplasmic deposits located in the midpiece are extruded to allow for subsequent cell elongation and nuclear condensation to occur during spermiogenesis [25] (Figure 1a).
If sperm maturation is not carried out properly, spermatozoa will retain the cytoplasm in the form of proximal or distal droplets, containing large amounts of cytoplasmic enzymes such as lactate dehydrogenase, superoxide dismutase (SOD) and glucose-6-phosphate dehydrogenase [24,25,26], all of which have been shown to be directly involved in the cellular oxidative balance reviewed by [27] (Figure 1b). In particular, glucose-6-phosphate dehydrogenase has been repeatedly associated with an increased risk for peroxidative damage, since it produces nicotinamide adenine dinucleotide phosphate (NADPH). Subsequently, ROS is generated from NADPH via the intramembrane-located NADPH oxidase 5 (NOX 5) [28], a prime enzyme responsible for ROS production in the acrosomal, equatorial or post-acrosomal regions of morphologically abnormal spermatozoa [29]. In addition, significant associations have been reported between ROS production and the stage of sperm development, with particularly high ROS levels being observed in immature spermatozoa [30]. Correspondingly, increased ROS production has been reported in patients suffering from teratozoospermia [31] or in subjects with a high occurrence of cytoplasmic droplets [25,26].
On the other hand, it has been suggested that apart from immature or morphologically abnormal sperm, increased amounts of damaged or dead male gametes may represent a threat to still viable and functional spermatozoa. The risks associated with the presence of dying spermatozoa lie in an increased release of intracellular content into their surroundings, containing large amounts of ROS, which may then attack the polyunsaturated fatty acids (PUFAs) located in the membrane of viable male gametes. Such compromised cells will then exhibit phosphatidylserine externalization, loss of membrane fluidity and an increased occurrence of DNA breaks, all of which will ultimately result in the activation of apoptotic or even necrotic cell death (Figure 1c). In this sense, cell necrosis represents a larger risk for the loss of viable spermatozoa, since any disturbance to intracellular homeostasis is succeeded by the rupture of membranous structures and a subsequent contamination of the extracellular space with toxic metabolites, sustaining the chain reaction of damage to other functional male gametes [32]. Moreover, an increased occurrence of dead spermatozoa has been suggested to trigger the immune system and to contribute to the formation of sperm granulomas. These abnormal structures consist of spermatozoa, macrophages, and other immune cells, eventually developing into a mass attached to the vas deferens, causing an obstruction and raising the intraluminal pressure [33].
- (a)
- Mitochondria are the primary source of ROS in structurally and functionally normal spermatozoa. Superoxide (O2•−) is created by electron leakage within NADH dehydrogenase (complex I) and ubiquinol cytochrome C oxidoreductase (complex III). The radical is quickly scavenged by superoxide dismutase (SOD) to generate hydrogen peroxide (H2O2). Alternatively, O2•− and H2O2 are generated by the NADPH-oxidase isoform 5 (NOX5) located in the plasma membrane and are activated through NADPH and calcium ions (Ca2+). Modified from [29]. CI—complex I, CII—complex II (succinate dehydrogenase), CIII—complex III, CIV—complex IV (cytochrome C oxidase), CV—complex V (ATP synthase).
- (b)
- Immature or abnormal spermatozoa present with cytoplasmic droplets containing glucose-6-phosphate dehydrogenase (G6PD), which produces NADPH serving as a substrate for the creation of O2•− by NADPH oxidase systems (NADPH Ox). SOD present in the droplets catalyzes the conversion of O2•− to H2O2. In the meantime, lactate dehydrogenase produces the reduced form of nicotinamide adenine dinucleotide (NADH), which then proceeds to be oxidized by the NADH dehydrogenase (complex I) in the mitochondria. Modified from [28].
- (c)
- In necrotic spermatozoa, the intracellular content including ROS will be released into the environment, subsequently contaminating the surrounding cells with toxic metabolites.
Created with BioRender.com (supplementary: Confirmation of Publication and Licensing Rights) (accessed on 28 October 2022).
2.2. Bacteria
The extent of ROS production by bacterial action is largely affected by the bacterial load, diversity, and metabolism of bacteria representing the seminal microflora [34]. In aerobic bacteria, more than 90% of cellular oxygen will be consumed by oxidative phosphorylation to produce ATP and water. Similarly to eukaryotic cells, a small portion of molecular oxygen captures 0.1%–1% of electrons present in the transport chain, continually forming superoxide radical (O2•−) by NADPH oxidase, which is the primary source of ROS in bacteria [35] (Figure 2a).
While obligate anaerobes are not equipped to perform aerobic respiration, it has been speculated that these can deploy low-potential electron transfer pathways to produce ROS [36]. O2•− can be generated abiotically from the reaction of O2 with ferrous ion-like metal ions [37,38]. Correspondingly to higher organisms, hydrogen peroxide (H2O2) in bacteria is generated as part of the dismutation of O2•− catalyzed by SOD [38] (Figure 2a). These two ROS are prevalent reactive intermediates produced by the bacterial cell, and the presence of antioxidant enzymes (such as SOD, catalase, thioxiredoxin or thiol peroxidase) in several bacterial species fortifies the evidence that bacteria themselves are capable of producing, regulating and sustaining certain levels of free radicals [39].
Additionally, O2•− as well as H2O2 have been observed to be actively released by numerous potentially uropathogenic bacteria [40,41]. As suggested by Fraczek et al. [22], additional ROS may subsequently cause significant damage to the sperm membranes, evidenced by increased malondialdehyde (MDA) levels in spermatozoa subjected to in vitro culture in the presence of Campylobacter ureolyticus, Staphylococcus haemolyticus, and Escherichia coli. Similar experiments of in vitro bacteriospermia designed by Ďuračka et al. [41] also revealed that selected staphylococci contributed to increased ROS levels in a sperm-bacteria co-culture, leading to decreased sperm motility, mitochondrial membrane potential and DNA integrity.
2.3. Leukocytes
Under physiological conditions, the primary source of ROS in semen is polymorphonuclear neutrophilic granulocytes, which are activated by an ongoing inflammatory process. The primary role of such peroxidase-positive leukocytes is to dispose of the pathogen by an increased release of ROS into the source of inflammation [42]. In this case, high amounts of O2•− and H2O2 are produced by a series of reactions referred to as respiratory burst. This phenomenon is mediated by the NADPH-oxidase complex, which is rapidly assembled following neutrophil activation. As discussed previously, NADPH oxidase produces O2•− similarly to NOX5 found in immature spermatozoa [28,29,43]. Subsequent dismutation catalyzed by SOD takes place to create H2O2. Both ROS may then be released individually, or these may interact with products of other microbicidal systems to generate ozone, a hydroxyl radical or singlet oxygen [43] (Figure 2b). While the primary role of respiratory burst lies in an additional line of protection to the cells against a pathogen, excessive infiltration and inappropriate activation of leukocytes may result in the release of excessive ROS and a subsequent deterioration of sperm function, as repeatedly observed in subjects suffering from leukocytospermia [9,10,44,45].
- (a)
- O2•− is created by NADPH oxidase localized on the bacterial plasma membrane. The molecule is further processed by SOD to generate H2O2. Additional ROS may be generated by the Fenton and Haber–Weiss reaction catalyzed by iron. Modified from [37].
- (b)
- Respiratory burst is activated by the NADPH-oxidase complex, which will produce O2•−. Subsequently, SOD will dismutate the radical to generate H2O2.
Created with BioRender.com (supplementary: Confirmation of Publication and Licensing Rights) (accessed on 28 October 2022).
3. Adhesion and Agglutination Events
The ability of bacteria to adhere to the cell surface is a crucial prerequisite for subsequent bacterial colonization of ejaculate since adhesion events may lead to alterations to the sperm morphology and motion behavior [46]. Furthermore, bacterial species attached to the sperm structures may release signaling molecules to attract other bacteria to form agglutinating complexes that will completely immobilize male gametes [47]. Further development of agglutination structures may then stimulate the release of extracellular polymeric substances and initiate biofilm formation [46,48]. Finally, successful bacterial adherence may stimulate the secretion of bacterial endotoxins with cytotoxic effects on sperm functional activity and fertilization ability [49].
3.1. Bacterial Adhesion
Bacteria accomplish adhesion events with polymeric fibers called “fimbriae” or “pilli” that serve to establish contact among the cellular surfaces and serve as virulence factors able to mediate the formation of bacterial aggregates by the recognition of receptors located on the host cell [49,50]. In the case of G− bacteria, their pilli possess a high affinity to mannose receptors found in the sperm membrane [47,51]. Two major types of pilli have been studied in uropathogenic G− bacteria, namely Type 1 fimbrinae with a high affinity to mannose located in the sperm head [47] and a mannose-resistant P fimbriae, which recognizes a-D-galpl-4-9-D-galp (gal gal) located predominantly in the sperm tail [52].
With respect to G+ bacteria (particularly Lactobacillus and Corynebacterium), the most common fimbrinae involved in bacterial colonization is SpaCBA with a versatile ability to bind to host cells, mucin, and mucous collagen. It has been revealed that bacterial aggregation enabled by this pillus only affects sperm motility without any impact on the morphology or vitality of male reproductive cells [46,47].
Several factors play a role in bacterial adherence, particularly the physical properties of the bacterial cell wall, area of contact, charge distribution or hydrophobicity [46]. While the exact molecular pathways driving the adhesion events are not completely understood yet, pilot studies indicate at least a partial involvement of oxidative mechanisms in enabling proper bacterial anchoring into the sperm membrane.
Integrins are transmembrane receptors that are intricately involved in important cell functions, such as cell adhesion or migration by regulating direct cell-to-cell associations or their interplay with extracellular matrix (ECM) proteins [53]. In addition to somatic cells, integrins have also been reported on the sperm surface and are known to play an important role in sperm membrane remodeling, organization of sperm protein complexes and sperm-oocyte fusion [54]. Nevertheless, activation of integrins by their specific ligands opens a transmembrane link for the transmission of molecular signals or mechanical forces across the plasma membrane in both directions. Several integrin receptors may be exploited by pathogenic agents to enable first contact with the host cell [53]. Furthermore, a growing list of recent reports demonstrates that integrin engagement by ECM proteins activates several intracellular cascades accompanied by induced ROS production promoting tight cell adhesion and cytoskeleton organization [55].
Although the underlying molecular mechanism of integrin-initiated ROS production remains to be elucidated, it has been hypothesized that the interplay between integrins and ECM proteins triggers ROS production by promoting changes in the mitochondrial redox function and activation of several oxidoreductases including NADPH-oxidases, 5-lipoxygenase and cyclooxygenase-2 [56,57,58]. The GTPase Rac1 activated by integrin engagement is essential for the stimulation of all ROS-producing systems [56]. Conversely, there is evidence that ROS can also affect integrin-mediated inside-out signaling by inducing conformational changes required for the activation of integrins [58]. According to Taddei et al. [59] an immediate cell-to-cell interaction carried out by integrins induces structural changes to adjacent adhesion molecules that are accompanied by mitochondrial ROS in the early stage of adhesion while 5-lipoxygenase-derived ROS peak during later stages of cell attachment. Furthermore, Chiarugi et al. [57] proposed that modulation of integrin signaling and cell adhesion by ROS is partially mediated by upregulation of focal adhesion kinase through oxidative inhibition of low molecular weight protein tyrosine phosphatases. Since ROS are capable of interacting with a wide array of biomolecules, their impact on cellular signaling events during cell adhesion depends on the site of production [56] and on the modification of redox-sensitive molecules. As such, target molecules may include phosphatases and kinases, receptors, transcription factors, actin and actin-associated proteins [56,58,59,60].
Finally, a recent report by Strempel et al. [61] has unraveled that H2O2 and hypochlorite that are frequently used as disinfecting agents may induce adaptive signaling pathways in Pseudomonas aeruginosa, particularly by increasing intracellular levels of the second messenger c-di-GMP and diguanylate cyclase. This enzyme is pivotal in mediating the motility and interactions of Pseudomonas aeruginosa with macrophages, and plays major roles in the biofilm formation typical of late stages of bacterial adhesion and aggregation. As such, it may be worth to assume that even extracellular oxidative agents traditionally used to counteract bacterial growth may in fact contribute to the initiation of bacterial adaptive mechanisms, which may further aggravate bacterial contamination of biological samples.
3.2. Sperm Agglutination
Sperm agglutination regarded as a direct result of bacterial adherence to the cell surface is defined as a phenomenon when previously motile spermatozoa stick to each other. Such compromised male gametes may be connected to each other either in a head-to-head, tail-to-tail, or mid-piece-to-tail pattern. Nonspecific agglutination may also occur by the adherence of either nonmotile sperm to each other or of motile sperm to cell debris, mucus, or other cell types [62]. The type of agglutination is by and large predetermined by the type of fimbrinae responsible for the initial bacterial adherence: while head-to-head agglutination is primarily caused by Type 1 fimbrinae, P-fimbriae are responsible for tail-to-tail agglutination. If the sample is infested by different bacterial types, a mixed agglutination may occur [63,64].
Events associated with sperm immobilization and an inherent high agglutination capacity of bacteria will ultimately lead to the formation of a complex structure called a biofilm. The unique architecture of biofilm provides a more favorable environment for bacteria to grow and reproduce, by creating niches occupied by bacterial cells that are covered by layers of extracellular polymeric molecules preventing the entry of antibacterial remedies [64,65]. The bacterial load within the biofilm is usually very high, promoting the spread of drug resistance phenotypes [65].
Two molecules released by bacteria play a pivotal role in the promotion of sperm agglutination and immobilization—the sperm agglutinating factor (SAF) and the sperm immobilizing factor (SIF). SAFs have been primarily detected in Escherichia coli [66] or staphylococci [67,68], and are able to block sperm motility, cause morphological abnormalities and act as spermicides at higher concentrations. The motility inhibition lies in the ability of SAFs to inhibit Mg2+-dependent ATPase as well as to trigger membrane receptors responsible for the initiation of apoptosis [66,67,68].
SIFs are molecules previously isolated from Escherichia coli [69] and Staphylococcus aureus [70], which directly immobilize male gametes without causing their agglutination. These molecules also have a specific receptor on the sperm membrane, and besides inhibiting ATP synthesis, also promote a premature acrosome reaction by causing an ionic imbalance in the sperm cell [69]. Furthermore, it has been revealed that Staphylococcus aureus-derived SIFs may have a direct cytotoxic effect on the male reproductive cell [70]. Besides Escherichia coli and staphylococci, sperm agglutination or immobilization have also been reported in the presence of Chlamydia trachomatis [71], Mycoplasma [72] or Trichomonas vaginalis [73].
Agglutinated sperm present with a high proportion of morphological and acrosomal abnormalities. Since both SAFs and SIFs target primarily ATPase, sperm agglutination and/or immobilization may lead to a disruption in the mitochondrial activity necessary for sperm motion [62]. As suggested by Wang et al. [74], sperm agglutination favors thiol-rich, oxidative-stressed, and apoptotic or necrotic spermatozoa, which themselves become additional sources of ROS. Furthermore, copper-induced lipid peroxidation (LPO) has been implicated in the loss of cholesterol from the membranous structures with a subsequent membrane destabilization in agglutinated spermatozoa [75]. Consequently, rupture of the plasma and mitochondrial membranes of spermatozoa engulfed by bacterial aggregates may result in the release of cytochrome C and ROS, leading to cell death and a decreased semen quality [14].
4. Bacterial Toxins
In addition to detrimental effects induced by physical contact between male gametes and bacteria, damage to spermatozoa may be caused by an array of molecules that are being synthesized and released by bacteria, including lipopolysaccharide (LPS) and hemolysins (Figure 3).
4.1. Lipopolysaccharide
LPS is a major component of the outer membrane of G− bacteria [76]. The molecule is defined as a prototypical endotoxin, which triggers a significant immune response [77]. During bacteriospermia, LPS is primarily recognized by the Toll-like receptor 4 (TLR4) in the acrosomal and tail regions of spermatozoa [78], leading to the activation of nuclear factor-κB to commence the transcription of downstream inflammatory factors [79]. LPS has been consistently linked to reprotoxicity [78,80,81] by affecting the expression patterns of pro-apoptotic genes [78,82], and by decreasing the levels of cyclic adenosine monophosphate (cAMP), Ca2+, and the intensity of protein phosphorylation [80]. LPS is also a very potent prooxidant, exposure to which leads to increased levels of mitochondrial ROS and occurrence of cell death with a concomitant decrease in sperm motility [78,83]. He et al. [78] observed that LPS induces mitochondrial transcription factor A (TFAM) translocation, which activates mitochondrial DNA (mtDNA) replication and subsequent activation of the mitochondrial oxidative phosphorylation system (OXPHOS) in boar sperm. As a result, ROS production in the mitochondria is elevated, which leads to oxidative stress (OS), mitochondrial membrane LPO, and a decrease in sperm motility and viability [84]. In this sense, the activation of TLR4 and mitochondrial translocation of TFAM are considered to act as prime mechanisms underlying LPS action [78,85].
4.2. Hemolysins
Hemolysins are unique pore-forming toxins that cause alterations to the membrane integrity and thus contribute to sperm immobilization. Escherichia coli is the sole producer of α-hemolysin, which forms pores in the host cell membrane, ultimately leading to cellular lysis [86]. The molecule is receptor-independent, since it presents with a versatility to permeabilize lipid bilayers of different architectures and compositions, and subsequently disturbs the intracellular osmotic pressure by forming voltage-dependent ion channels [87]. As such, α-hemolysin is only released if the bacterium adheres to the sperm surface [46]. With respect to spermatozoa, hemolytic Escherichia coli immobilizes spermatozoa more quickly and effectively when compared with nonhemolytic strains. An important side effect of hemolysin toxicity lies in the accumulation of lipoxygenase products, increased release of ROS (particularly O2•−) and reactive nitrogen species [87]. This disturbance of the oxidative milieu is accompanied by a decline in the antioxidant capacity and sperm mitochondrial membrane potential initiated by cellular rupture [88].
β-hemolysin is a toxin produced by Enterococcus that presents with the same mechanism of action as α-hemolysin and that exerts its toxic effects primarily on the sperm head, neck, and the middle segment of the tail. The head seems to be the primary target of the toxin, since a premature acrosome reaction and an increased release of hydrolytic enzymes are typical characteristics of urogenital infections caused by enterococci [89,90].
Finally, bacteriospermia is often accompanied by elevated levels of enzymes that are not per se toxic to male gametes but serve to create an optimal environment for the bacterial colonization of semen to proceed. These include but are not limited to proteases [91], lipases [92], coagulase or coagulation factors [93].
- (a)
- Since hemolysins are receptor-independent, a direct contact between the bacterium and the sperm cell is needed for their secretion. Once hemolysins are released, these will form pores in the plasma membrane, leading to the loss of its semipermeability and subsequent alterations to the intracellular osmotic balance. Hemolysins will additionally promote ROS overproduction, leading to peroxidation of the membrane lipids followed by membrane disintegration.
- (b)
- Once LPS is released, it will be recognized by the Toll-like receptor 4 (TRL4) located on the sperm surface. Its activation and a subsequent release of ROS will then trigger nuclear factor-κB (NF-κB) to promote inflammation, as well as caspases to initiate apoptosis. Meanwhile, LPS will activate the mitochondrial oxidative phosphorylation system, leading to increased ROS production by the mitochondria, peroxidation of mitochondrial lipids and a subsequent mitochondrial rupture. Initiation of the apoptotic process as well as mitochondrial dysfunction will then result in the loss of sperm motility.
Created with BioRender.com (supplementary: Confirmation of Publication and Licensing Rights) (accessed on 28 October 2022).
5. Leukocytospermia
Semen is a heterogeneous biological fluid containing an array of cells other than spermatozoa. Several components representing molecular immunity are found in ejaculates, such as immunoglobulins, chemokines, cytokines, or growth factors [94]. Furthermore, almost every semen specimen will contain white blood cells including granulocytes, lymphocytes, or macrophages, which represent the principal defense mechanism against foreign organisms [95]. Supraphysiological amounts of leukocytes is a condition defined as leukocytospermia, which according to the World Health Organization (WHO), is acknowledged if the concentration of peroxidase-positive white blood cells exceeds 1 × 106/mL of semen [96]. This cut-off value is considered a potential indicator of an ongoing and often asymptomatic male genital infection [95], particularly if a bladder infection or urethritis have been previously excluded [97].
Symptomatic as well as “silent” urogenital infections accompanied by increased levels of leukocytes are frequently observed in clinical practice [98]. Nevertheless, a conclusive interconnection among leukocytospermia and sub-fertility needs to be reinforced since some reports observed a direct correlation between increased concentrations of leukocytes and alterations to the sperm motion, morphology, or chromatin stability [99], while others excluded any impact of leukocytes on the fertilization potential of spermatozoa, particularly in the case of ARTs [100,101]. Despite this controversy, it is acknowledged that the presence of leukocytes in semen, regardless of their final concentration, is associated with ROS overproduction that may be of negative influence on certain sperm quality parameters [96].
Different mechanisms of leukocyte-inflicted damage to male gametes are known, out of which three may be considered as pre-dominant: (1) spermiophagy, (2) secretion of pro-inflammatory cytokines and (3) extracellular traps (ETs). Alterations to the sperm structure or function may occur individually or simultaneously, during isolated leukocytospermia or by leukocytospermia diagnosed concurrently with bacteriospermia [102].
Spermiophagy occurs when white blood cells, inherently programmed to detect and dispose of damaged or dead sperm, become overactivated and thus surround, engulf, and destroy even healthy male gametes [98,102,103]. The engulfment of spermatozoa by the cytoplasm of phagocytic cells is preceded by a direct contact of both cells and adhesion of leukocytes to the sperm head, midpiece, and flagellum, which is further reinforced by the secretion of proinflammatory cytokines [102,103].
According to Fraczek and Kurpisz [102], cytokines act within a network, where the toxicity of one cytokine may be modulated in the presence of other immune molecules. Among the most common pro-inflammatory cytokines, tumor necrosis factor (TNF) α released during inflammation and/or infection is most frequently reported to induce sperm cell death or to contribute to the loss of DNA integrity [104,105]. The cytotoxic behavior of TNF α may be further mediated via ROS or nitric oxide [106] that is inversely correlated with sperm motility. Within a vast and heterogenous group of proinflammatory interleukins (ILs), IL-1b, IL-6, IL-8, IL-12, and IL-18 have been reported to be involved in inflammation-inflicted damage to spermatozoa in response to bacterial infestation of semen and a compromised motion behavior [18,105,107]. Similarly to TNF α, increased levels of ILs have been observed in subfertile subjects or in semen samples with ROS overload [8,42,100].
A recently observed immune response to the presence of bacteria is the creation of ETs by activated white blood cells. ETs represent complex 3D web-like scaffolds of DNA strands embedded with histones and other antimicrobial molecules, such as myeloperoxidase, lactoferrin, elastase, bactericidal permeability-increasing protein or cathepsin G [108,109]. Physical contact between a leukocyte and a sperm cell leads to fast activation of the white blood cell, releasing ET structures that will then engulf the male gamete, causing its immobilization. This phenomenon is aggravated by phagocytosis, degranulation, and cytokine release [110,111]. Recent studies have observed that ROS are intricately involved in most reaction cascades promoting the release of ETs. Evidence gathered from a variety of perspectives has shown that ET-osis involves ROS generation by NADPH oxidase. This event occurs either directly via molecular signaling that enables ET formation and release, or indirectly by affecting other factors that modulate the process. Oxidative mechanisms are likely to act as important players in the regulatory network that determines whether the process of ET release and activation will be beneficial or detrimental [112].
6. Oxidative Stress and its Impact on Sperm Function
Regardless of the causes for ROS overproduction, oxidative stress has become one of the leading contributors to alterations in the sperm architecture or functional behavior. Oxidative stress is defined as a phenomenon when oxidants “overpower” antioxidants, when fast chain reactions of peroxidation develop and when these processes exhibit pathological effects on the cell [113]. The impact of OS is proportional to the intensity of these events since the cell is capable of overcoming minor perturbations and returning to its original state.
The complex and intricate cellular structure of male gametes predisposes them to be particularly vulnerable to OS, since their plasma membranes are characterized by large quantities of PUFAs, while containing only minor concentrations of antioxidant molecules in their intracellular compartments [17]. Increased ROS generation has been associated with a decline in sperm motility [18] either through the ability of H2O2 to diffuse across the membranes and inhibit the activity of several enzymes crucial for the sperm movement [114] or through inhibition of phosphorylation of axonemal proteins and subsequent sperm immobilization [115]. Furthermore, the motility loss is highly correlated with LPO, suggesting that oxidation of lipid structures is a major cause for alterations in the sperm motion behavior [116]. LPO of the sperm membrane is considered to be the key mechanism of ROS-induced sperm damage leading to subfertility [116], since PUFAs involved in the maintenance of membrane fluidity and transmembrane communication are most susceptible to oxidative insults [96]. Sperm LPO is a self-propagating process during which alkoxyl and peroxyl lipid radicals are formed, and subsequently act on other lipids in the membrane until all of them have undergone peroxidative changes [19]. These domino reactions will ultimately contribute to oxidative disintegration of DNA and proteins through lipoperoxides [19,116].
Within the molecule of DNA, bases and phosphodiester backbones are prone to oxidative insults. While sperm DNA protects itself against ROS by its specific compact organization and by antioxidants in the seminal plasma, spermatozoa lack any DNA repair mechanisms and rely on the oocyte to accomplish any repair following fertilization [117]. Out of different types of DNA damage caused by ROS, deletions, base modifications, DNA cross-links, occurrence of base-free sites, and chromosomal rearrangements are among the most commonly found in infertile males [118]. OS has also been linked to high frequencies of single- and double-strand DNA breaks and gene mutations that have been reported to occur, especially during sperm production and maturation [117,118]. Furthermore, mutations in mtDNA, which may also be affected by ROS, may compromise mitochondrial energy metabolism and sperm motility in vivo [119].
Oxidative insults to proteins may lead to in site-specific amino acid modifications, changes to the electric charge, disintegration of the peptide chain, and alterations to the susceptibility to proteolysis. Sulphur-containing amino acids are ought to be extraordinarily susceptible to changes in the oxidative balance. Such disturbance of the structure and function of sperm proteins may ultimately result in a decreased sperm ability to penetrate the zona pellucida and to fuse with the ovum [120].
Finally, ROS may trigger a chain of reactions that will ultimately lead to apoptosis. Under normal circumstances, programmed cell death is a natural process that helps to dispose of abnormal germ cells. Nevertheless, high ROS levels may disrupt the inner and outer mitochondrial membrane, accompanied by cytochrome C release and activation of caspases responsible for the promotion of apoptosis [121]. In the case of bacteriospermia, previous reports hypothesize that the principal pro-apoptotic mechanism lies in the activity of bacterial endotoxins, such as LPS or porins to interact with Toll-like receptors 2 and 4 located on the sperm surface [122]. Exposure of male gametes to these toxins may lead to ROS overproduction and subsequent mitochondrial depolarization followed by activation of caspase 3-mediated cell death [123]. Furthermore, the immune system may play a significant role in sperm apoptosis through the cytokine network, since it has been observed that IL-1b, IL-6, IL-8 or IL-18 could activate the Fas/Fas ligand complex on the sperm membrane, and subsequently initiate caspase 8-driven apoptotic machinery [14].
7. Clinical Effects of Bacteriospermia on Semen Quality
The effects of bacterial presence in semen on the resulting sperm structural integrity and functional activity are complex and multivariable as previously reported in several studies on humans [3,8,10,102,124,125] as well as domestic animals [23,90,126,127,128,129,130,131]. By and large, these reports agree that bacteriospermia may lead to the loss of sperm membrane stability [23,102,126,127,132], acrosome rupture and morphological alterations to the sperm head, mid-piece, and tail [23,48,102,127,133], and mitochondrial dysfunction accompanied by ATP depletion [23,126,127,133], all of which will ultimately result in the loss of sperm motility and a subsequent inability of spermatozoa to reach and fertilize the oocyte. A frequently observed phenomenon associated with the presence of bacteria in semen is sperm agglutination, DNA disintegration and early onset of cell death [14,41,126,127]. Furthermore, the presence of bacteria in semen has been frequently associated with the onset of leukocytospermia and increased levels of cytokines with cytotoxic properties [3,10,102]. Finally, preliminary reports have indicated that bacterial activity could modify the biochemical or physicochemical properties of semen, thus creating a less favorable environment for sperm survival [134,135]. Table 1 provides a summary of the most prominent effects of uropathogens on male gametes across currently available original papers published on the topic.
Recent studies on bacteriospermia have also provided a solid body of evidence, indicating a significant involvement of OS in bacteria-inflicted damage to male gametes. Interactions among bacteria and spermatozoa have been shown to trigger ROS overgeneration and subsequent oxidative insults, particularly to the membranous structures of the sperm cell, leading to the loss of sperm functionality in vivo or in vitro. Changes observed in the oxidative profile of semen and/or spermatozoa affected by the presence of bacteria are summarized in Table 2.
8. Management of Bacteriospermia in Practice: Strategies, Options and Alternatives
8.1. Antibiotics
Current management of bacteriospermia both in vivo as well as in vitro is by and large dependent upon antibiotics. The primary mechanism of action of these substances is based on inducing cell death by interfering with primary structural targets or by corrupting target-specific processes within the respective bacterial cell. Nevertheless, it is now well known that these target-specific interactions trigger stress responses that induce redox-related physiological alterations resulting in the formation ROS, which will further contribute to bacterial damage and/or death [152].
If bacteriospermia is caused by chronic or acute bacterial infections, these will be generally treated using a broad spectrum of antibiotics, among which ciprofloxacin, norfloxacin, and ofloxacin are most recommended in clinical practice [153]. In the meantime, supplementation of streptomycin, penicillin, spectinomycin, and lincomycin to semen extenders and cryopreservation media is required by law to avoid any potential transmission of disease among recipients [154].
Despite an indisputable contribution of traditional antibiotics to the prevention of bacteria-inflicted damage to male gametes, several reports have indicated that the inherently toxic properties of antibiotics could at least partially affect the male gametes themselves [155]. It has been recently reported that ciprofloxacin causes a rapid in vitro disintegration of sperm DNA [156], while gentamycin and ofloxacin administered in vivo may affect sperm motility and the extent of testicular apoptosis [157]. In this sense, there is a need to precisely define effective doses and eventual toxicity of antibiotics to male gametes in order to avoid any adverse effects.
Another aspect that must be taken into consideration lies in often-irrational antibiotic overuse in practice, which may endanger antibiotic susceptibility and lead to increased multidrug resistance in a wider range of bacterial species. A recent study has unraveled that more than 56% of all bacterial species found in boar semen were resistant to gentamycin, while only every fifth isolate was susceptible to gentamycin, lincomycin, penicillin, and neomycin [158]. In the meantime, Pseudomonas aeruginosa and Proteus mirabilis found in boar semen were resistant to spectinomycin, lincomycin, florfenicol, and streptomycin [159]. An alarming resistance of all isolates retrieved from bovine semen to penicillin has been reported, while the majority also presented with resistance patterns to tylosin and lincomycin [160]. With respect to humans, vancomycin and rifampicin were ineffective against G+ bacteria detected in Iraqi patients with fertility issues [161]. A similar study on infertile men unraveled the presence of Klebsiella pneumoniae, Escherichia coli, and Staphylococcus epidermidis with multidrug resistance patterns [162]. Furthermore, 90% of bacterial isolates found in in vitro fertilization (IVF) culture dishes were resistant to at least one of the antibiotics commonly used in ARTs [163], complementing a recent report on increased resistance of staphylococci in ART clinics [164].
8.2. Physical Removal of Bacteria
Since different concerns have arisen from an improper use of antibiotics in human or veterinarian andrology, several efforts have been made to develop procedures that would help with the physical removal of bacteria prior to further semen processing. Semen washing with an extra swim-up step has been reported to be more efficient in limiting the occurrence of potentially pathogenic bacteria in semen when compared to antibiotics [165]. Previous studies have also emphasized the advantages of density gradient centrifugation in removing bacteria and viruses from semen. Besides improving the microbial status of ejaculates, density gradient centrifugation also eliminates leukocytes or dead spermatozoa, significantly improving the quality of neat or thawed ejaculates even of suboptimal quality [41,90,165,166,167]. In addition, sperm filters have been developed for semen processing, causing less damage to the plasma membrane in comparison to centrifugation techniques [168]. Magnetic-activated cell-sorting (MACS), which identifies and eliminates apoptotic cells from semen using annexin V-conjugated superparamagnetic microbeads, could also become a suitable technique to reduce the proportion of damaged or dead spermatozoa prior to ARTs [169].
A new line of antimicrobial defense is represented by nanoparticles. Recent reports on iron oxide nanoparticles [170] or silver-carbon nanoparticles [171] applied to semen reveal that their use did not have any impact on sperm motility, viability, morphology or DNA integrity, while being effective in the elimination of several uropathogens including Escherichia coli, Staphylococcus aureus or Pseudomonas aeruginosa.
8.3. Alternative Antibacterial Supplements
A better prevention or management of bacteriospermia could also be accomplished by complementing conservative prescription medications with traditional remedies. Ethnopharmacological herbs such as copperleaf (Acalypha wilkesiana) [172], white weed (Ageratum conyzoides) [173], clove basil (Ocimum gratissimum) [174], pheasant-berry (Phylantus discoideus) [175], or Guinea pepper (Aframomum melegueta) [176] have been used for centuries now as remedies for male urogenital infections caused by pathogens such as Staphylococcus aureus, Escherichia coli, Klebsiella pneumoniae, Proteus mirabilis, Enterococcus faecium and Pseudomonas aeruginosa, all of which may additionally compromise semen quality. Herbal extracts and essential oils prepared from the above-mentioned plants may be used as floral beverages or functional foods with minimal known side effects on health. Furthermore, their use might help in reducing dependence on antibiotics and minimizing antibiotic resistance [177].
Furthermore, oxidative stress resulting from bacterial infestation of semen may be alleviated by products from Ajwain (Trachyspermum copticum), juniper (Juniperus communis), Gokshur (Tribulus terrestris), holy basil (Ocimum sanctum) or Ceylon cinnamon (Cinnamomum verum), which present with significant antioxidant properties [178]. Preliminary studies have indicated that administration of these remedies in the form of essential oils or extracts improved male steroidogenesis, which was accompanied by increased sperm motility, viability and DNA integrity [179,180].
Finally, the use of plant extracts [181] or pure natural biomolecules [90] as supplements to semen extenders could be yet another strategy to stabilize the oxidative milieu of in vitro processed spermatozoa while at the same time acting as antibacterial agents. A complex assessment of the biological activity of Schisandra chinensis revealed that the Omija extract was effective against Streptococcus pneumoniae and Enterococcus faecalis, while acting as a motion-promoting and metabolism-enhancing supplement to bovine sperm in vitro culture [182]. In the meantime, it was observed that marigold (Calendula officinalis) extract exhibited antibacterial activity against Staphylococcus aureus and Enterococcus faecalis [183] and at the same time provided significant antioxidant protection to extended bull semen [184]. In their experiments, Eini et al. [185] observed that tea tree (Melaleuca alternifolia) and rosemary (Rosmarinus officinalis) essential oils were capable of exerting similar effects to ampicillin on swine artificial insemination doses deprived of spermatozoa and spiked with Escherichia coli. In the meantime, moringa (Moringa oleifera) and ginger (Zingiber officinale) extracts supplemented with chilled banana shrimp spermatophores were able to maintain a higher percentage of viable sperm while inhibiting growth of pathogenic Vibrio and Pseudomonas [186]. With respect to pure biomolecules, a pivotal study by Duracka et al. [90] revealed that resveratrol, quercetin and curcumin provided protection to rabbit sperm structural integrity and functional activity during in vitro induced bacteriospermia by Enterococcus faecalis while exhibiting partial antibacterial activity against the bacterium.
While the preliminary results on alternative remedies against bacteriospermia collected from currently available literature are undoubtedly promising, precise and detail-oriented toxicological studies of these herbs, extracts and biomolecules are essential to elucidate their suitability for effective in vivo or in vitro management of male reproductive dysfunction caused by bacterial infection.
9. Conclusions
An increasing body of evidence strongly indicates that the presence and/or activity of bacteria in semen may exhibit detrimental effects on the structure and function of male gametes, rendering them to be ineffective in accomplishing their primary role of fertilizing the ovum. While the exact molecular machinery underlying bacteria-inflicted damage to spermatozoa is still not fully understood, it is feasible to state that oxidative mechanisms play a pivotal role in the etiology of bacteriospermia. Further studies unraveling the involvement of reactive oxygen species in bacterial infestation of ejaculates and consequences resulting from this condition will be needed to enable the development of new techniques to diagnose bacteriospermia in a more effective manner, followed by suitable strategies to manage the after-effects of bacterial action and/or oxidative damage to male reproductive cells.
Author Contributions
Conceptualization, E.T.; methodology, E.T. and M.Ď.; software, E.T.; validation, E.T. and F.B.; formal analysis, E.T., F.B. and M.Ď.; resources, E.T.; data curation, E.T., F.B. and M.Ď.; writing—original draft preparation, E.T., F.B. and M.Ď.; writing—review and editing, E.T., F.B. and M.Ď.; visualization, F.B.; supervision, E.T.; project administration, E.T.; funding acquisition, E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This publication was supported by the projects VEGA 1/0239/20, APVV-21-0095, KEGA 008SPU-4/2021 and by the Operational program Integrated Infrastructure within the project: Demand-driven research for sustainable and innovative food, Drive4SIFood 313011V336, cofinanced by the European Regional Development Fund.
Acknowledgments
We wish to thank the CeRA Team of Excellence for their support.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Okonofua, F.E.; Ntoimo, L.F.C.; Omonkhua, A.; Ayodeji, O.; Olafusi, C.; Unuabonah, E.; Ohenhen, V. Causes and Risk Factors for Male Infertility: A Scoping Review of Published Studies. Int. J. Gen. Med. 2022, 15, 5985–5997. [Google Scholar] [CrossRef]
- Das, S.; Roychoudhury, S.; Dey, A.; Jha, N.K.; Kumar, D.; Roychoudhury, S.; Slama, P.; Kesari, K.K. Bacteriospermia and Male Infertility: Role of Oxidative Stress. Adv. Exp. Med. Biol. 2022, 1358, 141–163. [Google Scholar] [PubMed]
- Rusz, A.; Pilatz, A.; Wagenlehner, F.; Linn, T.; Diemer, T.; Schuppe, H.C.; Lohmeyer, J.; Hossain, H.; Weidner, W. Influence of urogenital infections and inflammation on semen quality and male fertility. World J. Urol. 2012, 30, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Schuppe, H.C.; Pilatz, A.; Hossain, H.; Diemer, T.; Wagenlehner, F.; Weidner, W. Urogenital Infection as a Risk Factor for Male Infertility. Dtsch. Arztebl. Int. 2017, 114, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solomon, M.; Henkel, R. Semen culture and the assessment of genitourinary tract infections. Indian J. Urol. 2017, 3, 188–193. [Google Scholar]
- Bryan, E.R.; Kollipara, A.; Trim, L.K.; Armitage, C.W.; Carey, A.J.; Mihalas, B.; Redgrove, K.A.; McLaughlin, E.A.; Beagley, K.W. Hematogenous dissemination of Chlamydia muridarum from the urethra in macrophages causes testicular infection and sperm DNA damage. Biol. Reprod. 2019, 101, 748–759. [Google Scholar] [CrossRef]
- Ma, Z.S. Microbiome Transmission During Sexual Intercourse Appears Stochastic and Supports the Red Queen Hypothesis. Front. Microbiol. 2022, 12, 789983. [Google Scholar] [CrossRef]
- Tvrdá, E.; Lovíšek, D.; Gálová, E.; Schwarzová, M.; Kováčiková, E.; Kunová, S.; Žiarovská, J.; Kačániová, M. Possible Implications of Bacteriospermia on the Sperm Quality, Oxidative Characteristics, and Seminal Cytokine Network in Normozoospermic Men. Int. J. Mol. Sci. 2022, 23, 8678. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Hryhorowicz, M.; Gill, K.; Zarzycka, M.; Gaczarzewicz, D.; Jedrzejczak, P.; Bilinska, B.; Piasecka, M.; Kurpisz, M. The effect of bacteriospermia and leukocytospermia on conventional and nonconventional semen parameters in healthy young normozoospermic males. J. Reprod. Immunol. 2016, 118, 18–27. [Google Scholar] [CrossRef]
- Domes, T.; Lo, K.C.; Grober, E.D.; Mullen, J.B.; Mazzulli, T.; Jarvi, K. The incidence and effect of bacteriospermia and elevated seminal leukocytes on semen parameters. Fertil. Steril. 2012, 97, 1050–1055. [Google Scholar] [CrossRef]
- Ciornei, Ş.; Drugociu, D.; Ciornei, L.M.; Mareş, M.; Roşca, P. Total Aseptization of Boar Semen, to Increase the Biosecurity of Reproduction in Swine. Molecules 2021, 26, 6183. [Google Scholar] [CrossRef] [PubMed]
- Prabha, V.; Kaur, S. Isolation and purification of sperm immobilizing/agglutinating factors from bacteria and their corresponding receptors from human spermatozoa. In Chromatography—The Most Versatile Method of Chemical Analysis, 1st ed.; de Azevedo Calderon, L., Ed.; InTech Open: London, UK, 2012; pp. 295–310. [Google Scholar]
- Hauck, C.R. Cell adhesion receptors—Signaling capacity and exploitation by bacterial pathogens. Med. Microbiol. Immunol. 2002, 191, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Fraczek, M.; Hryhorowicz, M.; Gaczarzewicz, D.; Szumala-Kakol, A.; Kolanowski, T.J.; Beutin, L.; Kurpisz, M. Can apoptosis and necrosis coexist in ejaculated human spermatozoa during in vitro semen bacterial infection? J. Assist. Reprod. Genet. 2015, 32, 771–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackner, J.E.; Herwig, R.; Schmidbauer, J.; Schatzl, G.; Kratzik, C.; Marberger, M. Correlation of leukocytospermia with clinical infection and the positive effect of antiinflammatory treatment on semen quality. Fertil. Steril. 2006, 86, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Baskaran, S.; Finelli, R.; Agarwal, A.; Henkel, R. Reactive oxygen species in male reproduction: A boon or a bane? Andrologia 2021, 53, e13577. [Google Scholar] [CrossRef]
- Dutta, S.; Majzoub, A.; Agarwal, A. Oxidative stress and sperm function: A systematic review on evaluation and management. Arab J. Urol. 2019, 17, 87–97. [Google Scholar] [CrossRef] [Green Version]
- Kurkowska, W.; Bogacz, A.; Janiszewska, M.; Gabryś, E.; Tiszler, M.; Bellanti, F.; Kasperczyk, S.; Machoń-Grecka, A.; Dobrakowski, M.; Kasperczyk, A. Oxidative Stress is Associated with Reduced Sperm Motility in Normal Semen. Am. J. Mens Health 2020, 14, 1557988320939731. [Google Scholar] [CrossRef]
- Hosseinzadeh Colagar, A.; Karimi, F.; Jorsaraei, S.G. Correlation of sperm parameters with semen lipid peroxidation and total antioxidants levels in astheno- and oligoasheno- teratospermic men. Iran Red. Crescent. Med. J. 2013, 15, 780–785. [Google Scholar] [CrossRef] [Green Version]
- Marchiani, S.; Baccani, I.; Tamburrino, L.; Mattiuz, G.; Nicolò, S.; Bonaiuto, C.; Panico, C.; Vignozzi, L.; Antonelli, A.; Rossolini, G.M.; et al. Effects of common Gram-negative pathogens causing male genitourinary-tract infections on human sperm functions. Sci. Rep. 2021, 11, 19177. [Google Scholar] [CrossRef]
- Folliero, V.; Santonastaso, M.; Dell’Annunziata, F.; De Franciscis, P.; Boccia, G.; Colacurci, N.; De Filippis, A.; Galdiero, M.; Franci, G. Impact of Escherichia coli Outer Membrane Vesicles on Sperm Function. Pathogens 2022, 11, 782. [Google Scholar] [CrossRef]
- Fraczek, M.; Szumala-Kakol, A.; Jedrzejczak, P.; Kamieniczna, M.; Kurpisz, M. Bacteria trigger oxygen radical release and sperm lipid peroxidation in in vitro model of semen inflammation. Fertil. Steril. 2007, 88, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Kačániová, M.; Baláži, A.; Vašíček, J.; Vozaf, J.; Jurčík, R.; Ďuračka, M.; Žiarovská, J.; Kováč, J.; Chrenek, P. The Impact of Bacteriocenoses on Sperm Vitality, Immunological and Oxidative Characteristics of Ram Ejaculates: Does the Breed Play a Role? Animals 2022, 12, 54. [Google Scholar] [CrossRef] [PubMed]
- Cooper, T.G. Cytoplasmic droplets: The good, the bad or just confusing? Hum. Reprod. 2005, 20, 9–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gatimel, N.; Moreau, J.; Parinaud, J.; Léandri, R.D. Sperm morphology: Assessment, pathophysiology, clinical relevance, and state of the art in 2017. Andrology 2017, 5, 845–862. [Google Scholar] [CrossRef] [Green Version]
- Rengan, A.K.; Agarwal, A.; van der Linde, M.; du Plessis, S.S. An investigation of excess residual cytoplasm in human spermatozoa and its distinction from the cytoplasmic droplet. Reprod. Biol. Endocrinol. 2012, 10, 92. [Google Scholar] [CrossRef] [Green Version]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS Generation and Antioxidant Defense Systems in Normal and Malignant Cells. Oxid. Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellini, C.; D’Andrea, S.; Cordeschi, G.; Totaro, M.; Parisi, A.; Di Emidio, G.; Tatone, C.; Francavilla, S.; Barbonetti, A. Pathophysiology of Mitochondrial Dysfunction in Human Spermatozoa: Focus on Energetic Metabolism, Oxidative Stress and Apoptosis. Antioxidants 2021, 10, 695. [Google Scholar] [CrossRef]
- Villaverde, A.I.S.B.; Netherton, J.; Baker, M.A. From Past to Present: The Link Between Reactive Oxygen Species in Sperm and Male Infertility. Antioxidants 2019, 8, 616. [Google Scholar] [CrossRef] [Green Version]
- Gil-Guzman, E.; Ollero, M.; Lopez, M.C.; Sharma, R.K.; Alvarez, J.G.; Thomas, A.J., Jr.; Agarwal, A. Differential production of reactive oxygen species by subsets of human spermatozoa at different stages of maturation. Hum. Reprod. 2001, 16, 1922–1930. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Tvrda, E.; Sharma, R. Relationship amongst teratozoospermia, seminal oxidative stress and male infertility. Reprod. Biol. Endocrinol. 2014, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Roca, J.; Parrilla, I.; Gil, M.A.; Cuello, C.; Martinez, E.A.; Rodriguez-Martinez, H. Non-viable sperm in the ejaculate: Lethal escorts for contemporary viable sperm. Anim. Reprod. Sci. 2016, 169, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Samplaski, M.K.; Rodman, J.C.S.; Perry, J.M.; Marks, M.B.F.; Zollman, R.; Asanad, K.; Marks, S.F. Sperm granulomas: Predictive factors and impacts on patency post vasectomy reversal. Andrologia 2022, 54, e14439. [Google Scholar] [CrossRef] [PubMed]
- Imlay, J.A. Where in the world do bacteria experience oxidative stress? Environ. Microbiol. 2019, 21, 521–530. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Sethu, R.; Imlay, J.A. Endogenous superoxide is a key effector of the oxygen sensitivity of a model obligate anaerobe. Proc. Natl. Acad. Sci. USA 2018, 115, E3266–E3275. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ran, X.; Zhou, M.; Wang, K.; Wang, H.; Wang, Y. Oxidative stress and antioxidant mechanisms of obligate anaerobes involved in biological waste treatment processes: A review. Sci. Total Environ. 2022, 838, 156454. [Google Scholar] [CrossRef] [PubMed]
- Staerck, C.; Gastebois, A.; Vandeputte, P.; Calenda, A.; Larcher, G.; Gillmann, L.; Papon, N.; Bouchara, J.P.; Fleury, M.J.J. Microbial antioxidant defense enzymes. Microb. Pathog. 2017, 110, 56–65. [Google Scholar] [CrossRef]
- Chakraborty, S.P.; Pramanik, P.; Roy, S. Staphylococcus aureus Infection induced oxidative imbalance in neutrophils: Possible protective role of nanoconjugated vancomycin. ISRN Pharmacol. 2012, 2012, 435214. [Google Scholar] [CrossRef] [Green Version]
- Meier, B.; Habermehl, G.G. Evidence for superoxide dismutase and catalase in mollicutes and release of reactive oxygen species. Free Radic. Res. Commun. 1991, 12–13, 451–454. [Google Scholar] [CrossRef]
- Huycke, M.M.; Moore, D.; Joyce, W.; Wise, P.; Shepard, L.; Kotake, Y.; Gilmore, M.S. Extracellular superoxide production by Enterococcus faecalis requires demethylmenaquinone and is attenuated by functional terminal quinol oxidases. Mol. Microbiol. 2001, 42, 729–740. [Google Scholar] [CrossRef]
- Ďuračka, M.; Husarčíková, K.; Jančov, M.; Galovičová, L.; Kačániová, M.; Lukáč, N.; Tvrdá, E. Staphylococcus-Induced Bacteriospermia In Vitro: Consequences on the Bovine Spermatozoa Quality, Extracellular Calcium and Magnesium Content. Animals 2021, 11, 3309. [Google Scholar] [CrossRef]
- Henkel, R.; Offor, U.; Fisher, D. The role of infections and leukocytes in male infertility. Andrologia 2021, 53, e13743. [Google Scholar] [CrossRef] [PubMed]
- Robinson, J.M. Reactive oxygen species in phagocytic leukocytes. Histochem. Cell. Biol. 2008, 130, 281–297. [Google Scholar] [CrossRef] [Green Version]
- Saleh, R.A.; Agarwal, A.; Kandirali, E.; Sharma, R.K.; Thomas, A.J.; Nada, E.A.; Evenson, D.P.; Alvarez, J.G. Leukocytospermia is associated with increased reactive oxygen species production by human spermatozoa. Fertil. Steril. 2002, 78, 1215–1224. [Google Scholar] [CrossRef]
- Qiao, X.; Zeng, R.; Yang, Z.; Xu, L.; Ma, Q.; Yang, Y.; Bai, Y.; Yang, Y.; Bai, P. Effects of leukocytospermia on the outcomes of assisted reproductive technology. Andrologia 2022, 54, e14403. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Dai, J.; Chen, T. Role of Lactobacillus in female infertility via modulating sperm agglutination and immobilization. Front. Cell. Infect. Microbiol. 2021, 10, 620529. [Google Scholar] [CrossRef]
- Wolff, H.; Panhans, A.; Stolz, W.; Meurer, M. Adherence of Escherichia coli to sperm: A mannose mediated phenomenon leading to agglutination of sperm and E. coli. Fertil. Steril. 1993, 60, 154–158. [Google Scholar] [CrossRef]
- Rennemeier, C.; Frambach, T.; Hennicke, F.; Dietl, J.; Staib, P. Microbial quorum-sensing molecules induce acrosome loss and cell death in human spermatozoa. Infect. Immun. 2009, 77, 4990–4997. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhang, K.; Yao, Y.; Li, J.; Deng, S. Bacterial infections affect male fertility: A focus on the oxidative stress-autophagy axis. Front. Cell. Dev. Biol. 2021, 9, 727812. [Google Scholar] [CrossRef]
- Stones, D.H.; Krachler, A.M. Against the tide: The role of bacterial adhesion in host colonization. Biochem. Soc. Trans. 2016, 44, 1571–1580. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Chen, T.; Chen, Y.; Luo, T.; Tan, B.; Chen, H.; Xin, H. Evaluation of the inhibitory effects of vaginal microorganisms on sperm motility in vitro. Exp. Ther. Med. 2020, 19, 535–544. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, J.; Srivastava, S.; Singh, M. Pathogenomics of uropathogenic Escherichia coli. Indian J. Med. Microbiol. 2012, 30, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Scibelli, A.; Roperto, S.; Manna, L.; Pavone, L.M.; Tafuri, S.; Della Morte, R.; Staiano, N. Engagement of integrins as a cellular route of invasion by bacterial pathogens. Vet. J. 2007, 173, 482–491. [Google Scholar] [CrossRef]
- Merc, V.; Frolikova, M.; Komrskova, K. Role of Integrins in Sperm Activation and Fertilization. Int. J. Mol. Sci. 2021, 22, 11809. [Google Scholar] [CrossRef] [PubMed]
- Brakebusch, C.; Bouvard, D.; Stanchi, F.; Sakai, T.; Fassler, R. Integrins in invasive growth. J. Clin. Investig. 2002, 109, 999–1006. [Google Scholar] [CrossRef] [PubMed]
- Svineng, G.; Ravuri, C.; Rikardsen, O.; Huseby, N.E.; Winberg, J.O. The role of reactive oxygen species in integrin and matrix metalloproteinase expression and function. Connect. Tissue Res. 2008, 49, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Chiarugi, P.; Pani, G.; Giannoni, E.; Taddei, L.; Colavitti, R.; Raugei, G.; Symons, M.; Borrello, S.; Galeotti, T.; Ramponi, G. Reactive oxygen species as essential mediators of cell adhesion: The oxidative inhibition of a FAK tyrosine phosphatase is required for cell adhesion. J. Cell. Biol. 2003, 161, 933–944. [Google Scholar] [CrossRef]
- Zeller, K.S.; Riaz, A.; Sarve, H.; Li, J.; Tengholm, A.; Johansson, S. The role of mechanical force and ROS in integrin-dependent signals. PLoS ONE 2013, 8, e64897. [Google Scholar]
- Taddei, M.L.; Parri, M.; Mello, T.; Catalano, A.; Levine, A.D.; Raugei, G.; Ramponi, G.; Chiarugi, P. Integrin-mediated cell adhesion and spreading engage different sources of reactive oxygen species. Antioxid. Redox Signal. 2007, 9, 469–481. [Google Scholar] [CrossRef]
- Goitre, L.; Pergolizzi, B.; Ferro, E.; Trabalzini, L.; Retta, S.F. Molecular Crosstalk between Integrins and Cadherins: Do Reactive Oxygen Species Set the Talk? J. Signal. Transduct. 2012, 2012, 807682. [Google Scholar] [CrossRef] [Green Version]
- Strempel, N.; Nusser, M.; Neidig, A.; Brenner-Weiss, G.; Overhage, J. The Oxidative Stress Agent Hypochlorite Stimulates c-di-GMP Synthesis and Biofilm Formation in Pseudomonas aeruginosa. Front. Microbiol. 2017, 8, 2311. [Google Scholar] [CrossRef] [Green Version]
- Berger, G.K.; Smith-Harrison, L.I.; Sandlow, J.I. Sperm agglutination: Prevalence and contributory factors. Andrologia 2019, 51, e13254. [Google Scholar] [CrossRef] [PubMed]
- Tvrdá, E.; Ďuračka, M.; Benko, F.; Lukáč, N. Bacteriospermia—A formidable player in male subfertility. Open Life Sci. 2022, 17, 1001–1029. [Google Scholar] [CrossRef] [PubMed]
- Katsikogianni, M.; Missirlis, Y.F. Concise review of mechanisms of bacterial adhesion to biomaterials and of techniques used in estimating bacteria-material interactions. Eur. Cell. Mater. 2004, 8, 37–57. [Google Scholar] [CrossRef] [PubMed]
- Rabin, N.; Zheng, Y.; Opoku-Temeng, C.; Du, Y.; Bonsu, E.; Sintim, H.O. Biofilm formation mechanisms and targets for developing antibiofilm agents. Future Med. Chem. 2015, 7, 493–512. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Kaur, S.; Rishi, P.; Singh, S.K.; Prabha, V. Evidence for the occurrence of receptor in sperm for spermagglutinating factor isolated from Escherichia coli. Gynecol. Endocrinol. 2012, 34, 207–209. [Google Scholar]
- Pant, N.C.; Singh, R.; Gupta, V.; Chauhan, A.; Mavuduru, R.; Prabha, V.; Sharma, P. Contraceptive efficacy of sperm agglutinating factor from Staphylococcus warneri, isolated from the cervix of a woman with inexplicable infertility. Reprod. Biol. Endocrinol. 2019, 17, 85. [Google Scholar] [CrossRef] [Green Version]
- Ohri, M.; Prabha, V. Isolation of a sperm-agglutinating factor from Staphylococcus aureus isolated from a woman with unexplained infertility. Fertil. Steril. 2005, 84, 1539–1541. [Google Scholar] [CrossRef]
- Vander, H.; Gupta, S.; Kaur, S.; Kaur, K.; Prabha, V. Characterization of sperm immobilization factor from Escherichia coli and its receptor to study the underlying mechanism of sperm immobilization. Am. J. Biomed Sci. 2013, 5, 25–33. [Google Scholar] [CrossRef]
- Prabha, V.; Gupta, T.; Kaur, S.; Kaur, N.; Kala, S.; Singh, A. Isolation of a spermatozoal immobilization factor from Staphylococcus aureus filtrates. Can. J. Microbiol. 2009, 55, 874–878. [Google Scholar] [CrossRef]
- Sellami, H.; Znazen, A.; Sellami, A.; Mnif, H.; Louati, N.; Ben Zarrouk, S.; Keskes, L.; Rebai, T.; Gdoura, R.; Hammami, A. Molecular detection of Chlamydia trachomatis and other sexually transmitted bacteria in semen of male partners of infertile couples in Tunisia: The effect on semen parameters and spermatozoa apoptosis markers. PLoS ONE 2014, 9, e98903. [Google Scholar] [CrossRef]
- Gdoura, R.; Kchaou, W.; Chaari, C.; Znazen, A.; Keskes, L.; Rebai, T.; Gdoura, R.; Hammami, A. Ureaplasma urealyticum, Ureaplasma parvum, Mycoplasma hominis and Mycoplasma genitalium infections and semen quality of infertile men. BMC Infect. Dis. 2007, 7, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monga, M.; Roberts, J.A. Spermagglutination by bacteria: Receptor-specific interactions. J. Androl. 1994, 15, 151–156. [Google Scholar] [PubMed]
- Wang, T.E.; Yeh, L.Y.; Kuo-Kuang Lee, R.; Lu, C.H.; Yang, T.H.; Kuo, Y.W.; Joshi, R.; Tsai, P.S.; Li, S.H. Secretory mouse quiescin sulfhydryl oxidase 1 aggregates defected human and mouse spermatozoa in vitro and in vivo. iScience 2021, 24, 103167. [Google Scholar] [CrossRef] [PubMed]
- Leahy, T.; Rickard, J.P.; Aitken, R.J.; de Graaf, S.P. D-penicillamine prevents ram sperm agglutination by reducing the disulphide bonds of a copper-binding sperm protein. Reproduction 2016, 151, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [Green Version]
- Osborn, M.J.; Rosen, S.M.; Rothfield, L.; Zeleznick, L.; Horecker, B.L. Lipopolysaccharide of the Gram-negative cell wall. Science 1964, 145, 783–789. [Google Scholar] [CrossRef]
- He, B.; Guo, H.; Gong, Y.; Zhao, R. Lipopolysaccharide-induced mitochondrial dysfunction in boar sperm is mediated by activation of oxidative phosphorylation. Theriogenology 2017, 87, 1–8. [Google Scholar] [CrossRef]
- Gioannini, T.L.; Weiss, J.P. Regulation of interactions of Gramnegative bacterial endotoxins with mammalian cells. Immunol. Res. 2007, 39, 249–260. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, D.; He, Y.; Ding, Z.; Mao, F.; Luo, T.; Zhang, X. Lipopolysaccharide Compromises Human Sperm Function by Reducing Intracellular cAMP. Tohoku J. Exp. Med. 2016, 238, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Makvandi, A.; Kowsar, R.; Hajian, M.; Mahdavi, A.H.; Tanhaei Vash, N.; Nasr-Esfahani, M.H. Alpha lipoic acid reverses the negative effect of LPS on mouse spermatozoa and developmental competence of resultant embryos in vitro. Andrology 2019, 7, 350–356. [Google Scholar] [CrossRef]
- Hakimi, H.; Geary, I.; Pacey, A.; Eley, A. Spermicidal activity of bacterial lipopolysaccharide is only partly due to lipid A. J. Androl. 2006, 27, 774–779. [Google Scholar] [CrossRef]
- Urata, K.; Narahara, H.; Tanaka, Y.; Egashira, T.; Takayama, F.; Miyakawa, I. Effect of endotoxin-induced reactive oxygen species on sperm motility. Fertil. Steril. 2001, 76, 163–166. [Google Scholar] [CrossRef]
- Barbonetti, A.; Vassallo, M.R.; Costanzo, M.; Battista, N.; Maccarrone, M.; Francavilla, S.; Francavilla, F. Involvement of cannabinoid receptor-1 activation in mitochondrial depolarizing effect of lipopolysaccharide in human spermatozoa. Andrology 2014, 2, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Shi, D.; Li, X.; Gong, W.; Wu, F.; Guo, X.; Xiao, H.; Liu, L.; Zhou, H. TLR signalling affects sperm mitochondrial function and motility via phosphatidylinositol 3-kinase and glycogen synthase kinase-3α. Cell. Signal. 2016, 28, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Ristow, L.C.; Welch, R.A. Hemolysin of uropathogenic Escherichia coli: A cloak or a dagger? Biochim. Biophys. Acta 2016, 1858, 538–545. [Google Scholar] [CrossRef] [PubMed]
- Baronetti, J.L.; Villegas, N.A.; Aiassa, V.; Paraje, M.G.; Albesa, I. Hemolysin from Escherichia coli induces oxidative stress in blood. Toxicon 2013, 70, 15–20. [Google Scholar] [CrossRef]
- May, A.K.; Gleason, T.G.; Sawyer, R.G.; Pruett, T.L. Contribution of Escherichia coli alpha-hemolysin to bacterial virulence and to intraperitoneal alterations in peritonitis. Infect. Immun. 2000, 68, 176–183. [Google Scholar] [CrossRef] [Green Version]
- Qiang, H.; Jiang, M.S.; Lin, J.Y.; He, W.M. Influence of enterococci on human sperm membrane in vitro. Asian J. Androl. 2007, 9, 77–81. [Google Scholar] [CrossRef]
- Duracka, M.; Lukac, N.; Kacaniova, M.; Kantor, A.; Hleba, L.; Ondruska, L.; Tvrda, E. Antibiotics Versus Natural Biomolecules: The Case of In Vitro Induced Bacteriospermia by Enterococcus Faecalis in Rabbit Semen. Molecules 2019, 24, 4329. [Google Scholar] [CrossRef] [Green Version]
- Dubin, G.; Koziel, J.; Pyrc, K.; Wladyka, B.; Potempa, J. Bacterial proteases in disease—Role in intracellular survival, evasion of coagulation/ fibrinolysis innate defenses, toxicoses and viral infections. Curr. Pharm. Des. 2013, 19, 1090–1113. [Google Scholar] [CrossRef]
- Bender, J.; Flieger, A. Lipases as Pathogenicity Factors of Bacterial Pathogens of Humans. In Handbook of Hydrocarbon and Lipid Microbiology, 1st ed.; Timmis, K.N., Ed.; Springer: Berlin, Germany, 2010; pp. 3241–3258. [Google Scholar]
- Liu, Y.; Gao, W.; Yang, J.; Guo, H.; Zhang, J.; Ji, Y. Contribution of Coagulase and Its Regulator SaeRS to Lethality of CA-MRSA 923 Bacteremia. Pathogens 2021, 10, 1396. [Google Scholar] [CrossRef] [PubMed]
- Politch, J.A.; Tucker, L.; Bowman, F.P.; Anderson, D.J. Concentrations and significance of cytokines and other immunologic factors in semen of healthy fertile men. Hum. Reprod. 2007, 22, 2928–2935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lackner, J.; Schatzl, G.; Horvath, S.; Kratzik, C.; Marberger, M. Value of counting white blood cells (WBC) in semen samples to predict the presence of bacteria. Eur. Urol. 2006, 49, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Drevet, J.R.; Moazamian, A.; Gharagozloo, P. Male Infertility and Oxidative Stress: A Focus on the Underlying Mechanisms. Antioxidants 2022, 11, 306. [Google Scholar] [CrossRef]
- Boitrelle, F.; Shah, R.; Saleh, R.; Henkel, R.; Kandil, H.; Chung, E.; Vogiatzi, P.; Zini, A.; Arafa, M.; Agarwal, A. The Sixth Edition of the WHO Manual for Human Semen Analysis: A Critical Review and SWOT Analysis. Life 2021, 11, 1368. [Google Scholar] [CrossRef]
- Khodamoradi, K.; Kuchakulla, M.; Narasimman, M.; Khosravizadeh, Z.; Ali, A.; Brackett, N.; Ibrahim, E.; Ramasamy, R. Laboratory and clinical management of leukocytospermia and hematospermia: A review. Ther. Adv. Reprod. Health 2020, 14, 2633494120922511. [Google Scholar] [CrossRef]
- Eldamnhoury, E.M.; Elatrash, G.A.; Rashwan, H.M.; El-Sakka, A.I. Association between leukocytospermia and semen interleukin-6 and tumor necrosis factor-alpha in infertile men. Andrology 2018, 6, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Lackner, J.E.; Märk, I.; Sator, K.; Huber, J.; Sator, M. Effect of leukocytospermia on fertilization and pregnancy rates of artificial reproductive technologies. Fertil. Steril. 2008, 90, 869–871. [Google Scholar] [CrossRef]
- Cavagna, M.; Oliveira, J.B.; Petersen, C.G.; Mauri, A.L.; Silva, L.F.; Massaro, F.C.; Baruffi, R.L.; Franco, J.G., Jr. The influence of leukocytospermia on the outcomes of assisted reproductive technology. Reprod. Biol. Endocrinol. 2012, 10, 44. [Google Scholar] [CrossRef] [Green Version]
- Fraczek, M.; Kurpisz, M. Mechanisms of the harmful effects of bacterial semen infection on ejaculated human spermatozoa: Potential inflammatory markers in semen. Folia Histochem. Cytobiol. 2015, 53, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Sanocka, D.; Fraczek, M.; Jedrzejczak, P.; Szumała-Kakol, A.; Kurpisz, M. Male genital tract infection: An influence of leukocytes and bacteria on semen. J. Reprod. Immunol. 2004, 62, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Lampiao, F.; du Plessis, S.S. TNF-alpha and IL-6 affect human sperm function by elevating nitric oxide production. Reprod. Biomed. Online 2008, 17, 628–631. [Google Scholar] [CrossRef]
- Perdichizzi, A.; Nicoletti, F.; La Vignera, S.; Barone, N.; D’agata, R.; Vicari, E.; Calogero, A.E. Effects of tumour necrosis factor-alpha on human sperm motility and apoptosis. J. Clin. Immunol. 2007, 27, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Dziadecki, W.; Celińska, A.; Fracki, S.; Bablok, L.; Barcz, E. Interleukin 1b and interleukin 18 and their connection with leukocytospermia in human semen. Centr. Eur. J. Immunol. 2010, 35, 157–161. [Google Scholar]
- Koçak, I.; Yenisey, C.; Dündar, M.; Okyay, P.; Serter, M. Relationship between seminal plasma interleukin-6 and tumor necrosis factor alpha levels with semen parameters in fertile and infertile men. Urol. Res. 2002, 30, 263–267. [Google Scholar]
- Mateo-Otero, Y.; Zambrano, F.; Catalán, J.; Sánchez, R.; Yeste, M.; Miro, J.; Fernandez-Fuertes, B. Seminal plasma, and not sperm, induces time and concentration-dependent neutrophil extracellular trap release in donkeys. Equine Vet. J. 2022, 54, 415–426. [Google Scholar] [CrossRef]
- Moya, C.; Rivera-Concha, R.; Pezo, F.; Uribe, P.; Schulz, M.; Sánchez, R.; Hermosilla, C.; Taubert, A.; Gärtner, U.; Zambrano, F. Adverse Effects of Single Neutrophil Extracellular Trap-Derived Components on Bovine Sperm Function. Animals 2022, 12, 1308. [Google Scholar] [CrossRef]
- Schulz, M.; Zambrano, F.; Schuppe, H.C.; Wagenlehner, F.; Taubert, A.; Gaertner, U.; Sánchez, R.; Hermosilla, C. Monocyte-derived extracellular trap (MET) formation induces aggregation and affects motility of human spermatozoa in vitro. Syst. Biol. Reprod. Med. 2019, 65, 357–366. [Google Scholar] [CrossRef]
- Zambrano, F.; Carrau, T.; Gärtner, U.; Seipp, A.; Taubert, A.; Felmer, R.; Sanchez, R.; Hermosilla, C. Leukocytes coincubated with human sperm trigger classic neutrophil extracellular traps formation, reducing sperm motility. Fertil. Steril. 2016, 106, 1053–1060. [Google Scholar] [CrossRef] [Green Version]
- Stoiber, W.; Obermayer, A.; Steinbacher, P.; Krautgartner, W.-D. The Role of Reactive Oxygen Species (ROS) in the Formation of Extracellular Traps (ETs) in Humans. Biomolecules 2015, 5, 702–723. [Google Scholar] [CrossRef] [Green Version]
- Pruchniak, M.P.; Aražna, M.; Demkow, U. Biochemistry of Oxidative Stress. Adv. Exp. Med. Biol. 2016, 878, 9–19. [Google Scholar]
- Pujianto, D.A.; Oktarina, M.; Sharma Sharaswati, I.A.; Yulhasri. Hydrogen Peroxide Has Adverse Effects on Human Sperm Quality Parameters, Induces Apoptosis, and Reduces Survival. J. Hum. Reprod. Sci. 2021, 14, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Peña, F.J.; O’Flaherty, C.; Ortiz Rodríguez, J.M.; Martín Cano, F.E.; Gaitskell-Phillips, G.L.; Gil, M.C.; Ortega Ferrusola, C. Redox Regulation and Oxidative Stress: The Particular Case of the Stallion Spermatozoa. Antioxidants 2019, 8, 567. [Google Scholar] [CrossRef] [Green Version]
- Moazamian, R.; Polhemus, A.; Connaughton, H.; Fraser, B.; Whiting, S.; Gharagozloo, P.; Aitken, R.J. Oxidative stress and human spermatozoa: Diagnostic and functional significance of aldehydes generated as a result of lipid peroxidation. Mol. Hum. Reprod. 2015, 21, 502–515. [Google Scholar] [CrossRef] [Green Version]
- Rashki Ghaleno, L.; Alizadeh, A.; Drevet, J.R.; Shahverdi, A.; Valojerdi, M.R. Oxidation of Sperm DNA and Male Infertility. Antioxidants 2021, 10, 97. [Google Scholar] [CrossRef]
- Hosen, M.B.; Islam, M.R.; Begum, F.; Kabir, Y.; Howlader, M.Z. Oxidative stress induced sperm DNA damage, a possible reason for male infertility. Iran. J. Reprod. Med. 2015, 13, 525–532. [Google Scholar] [PubMed]
- Durairajanayagam, D.; Singh, D.; Agarwal, A.; Henkel, R. Causes and consequences of sperm mitochondrial dysfunction. Andrologia 2021, 53, e13666. [Google Scholar] [CrossRef]
- Mostek, A.; Slowinska, M.; Judycka, S.; Karol, H.; Ciereszko, A.; Dietrich, M.A. Identification of oxidatively modified proteins due to cryopreservation of carp semen. J. Anim. Sci. 2018, 96, 1453–1465. [Google Scholar] [CrossRef] [Green Version]
- Asadi, A.; Ghahremani, R.; Abdolmaleki, A.; Rajaei, F. Role of sperm apoptosis and oxidative stress in male infertility: A narrative review. Int. J. Reprod. Biomed. 2021, 19, 493–504. [Google Scholar] [CrossRef]
- Fujita, Y.; Mihara, T.; Okazaki, T.; Shitanaka, M.; Kushino, R.; Ikeda, C.; Negishi, H.; Liu, Z.; Richards, J.S.; Shimada, M. Toll-like receptors (TLR) 2 and 4 on human sperm recognize bacterial endotoxins and mediate apoptosis. Hum. Reprod. 2011, 26, 2799–2806. [Google Scholar] [CrossRef] [Green Version]
- Satta, A.; Stivala, A.; Garozzo, A.; Morello, A.; Perdichizzi, A.; Vicari, E.; Salmeri, M.; Calogero, A.E. Experimental Chlamydia trachomatis infection causes apoptosis in human sperm. Hum. Reprod. 2006, 21, 134–137. [Google Scholar] [CrossRef] [Green Version]
- Pergialiotis, V.; Karampetsou, N.; Perrea, D.N.; Konstantopoulos, P.; Daskalakis, G. The impact of bacteriospermia on semen parameters: A meta-analysis. J. Family Reprod. Health 2018, 12, 73–83. [Google Scholar]
- Baud, D.; Pattaroni, C.; Vulliemoz, N.; Castella, V.; Marsland, B.J.; Stojanov, M. Sperm microbiota and its impact on semen parameters. Front. Microbiol. 2019, 10, 234. [Google Scholar] [CrossRef] [Green Version]
- Lenický, M.; Slanina, T.; Kačániová, M.; Galovičová, L.; Petrovičová, M.; Ďuračka, M.; Benko, F.; Kováč, J.; Tvrdá, E. Identification of Bacterial Profiles and Their Interactions with Selected Quality, Oxidative, and Immunological Parameters of Turkey Semen. Animals 2021, 11, 1771. [Google Scholar] [CrossRef]
- Ďuračka, M.; Belić, L.; Tokárová, K.; Žiarovská, J.; Kačániová, M.; Lukáč, N.; Tvrdá, E. Bacterial communities in bovine ejaculates and their impact on the semen quality. Syst. Biol. Reprod. Med. 2021, 67, 438–449. [Google Scholar] [CrossRef]
- Medo, J.; Žiarovská, J.; Ďuračka, M.; Tvrdá, E.; Baňas, Š.; Gábor, M.; Kyseľ, M.; Kačániová, M. Core Microbiome of Slovak Holstein Friesian Breeding Bulls’ Semen. Animals 2021, 11, 3331. [Google Scholar] [CrossRef]
- Otter, A. Bacterial isolates from the semen of rams with suspected infertility. Vet. Rec. 2008, 162, 623–624. [Google Scholar] [CrossRef]
- Tvrdá, E.; Bučko, O.; Rojková, K.; Ďuračka, M.; Kunová, S.; Kováč, J.; Benko, F.; Kačániová, M. The Efficiency of Selected Extenders against Bacterial Contamination of Boar Semen in a Swine Breeding Facility in Western Slovakia. Animals 2021, 11, 3320. [Google Scholar] [CrossRef]
- Quiñones-Pérez, C.; Martínez, A.; Crespo, F.; Vega-Pla, J.L. Comparative Semen Microbiota Composition of a Stallion in a Taylorella equigenitalis Carrier and Non-Carrier State. Animals 2020, 10, 868. [Google Scholar] [CrossRef]
- Bussalleu, E.; Yeste, M.; Sepúlveda, L.; Torner, E.; Pinart, E.; Bonet, S. Effects of different concentrations of enterotoxigenic and verotoxigenic E. coli on boar sperm quality. Anim. Reprod. Sci. 2011, 127, 176–182. [Google Scholar] [CrossRef]
- Fraczek, M.; Piasecka, M.; Gaczarzewicz, D.; Szumala-Kakol, A.; Kazienko, A.; Lenart, S.; Laszczynska, M.; Kurpisz, M. Membrane stability and mitochondrial activity of human-ejaculated spermatozoa during in vitro experimental infection with Escherichia coli, Staphylococcus haemolyticus and Bacteroides ureolyticus. Andrologia 2012, 44, 315–329. [Google Scholar] [CrossRef] [PubMed]
- Ďuračka, M.; Kováčik, A.; Kačániová, M.; Lukáč, N.; Tvrdá, E. Bacteria may deteriorate progressive motility of bovine spermatozoa and biochemical parameters of seminal plasma. J. Microbiol. Biotech. Food Sci. 2020, 9, 844–847. [Google Scholar] [CrossRef]
- Tvrda, E.; Fik, M.; Kovacik, A.; Duracka, M.; Kacaniova, M. Biochemical and bacteriological characterization of Dachshund semen: A correlation study. Reprod. Domest. Anim. 2022, 57, 60. [Google Scholar]
- Kumar, V.; Garg, N. Effect of Escherichia coli on Semen Quality of Infertile Human Male. Virol. Immunol. J. 2019, 3, 000214. [Google Scholar] [CrossRef]
- Pelliccione, F.; D’Angeli, A.; Cordeschi, G.; Mihalca, R.; Ciociola, F.; Necozione, S.; Francavilla, F.; Francavilla, S. Seminal macrophages in ejaculates from men with couple infertility. Int. J. Androl. 2009, 32, 623–628. [Google Scholar] [CrossRef]
- Kala, S.; Singh, A.; Prabha, V.; Singh, R.; Sharma, P. Escherichia coli attaches to human spermatozoa: Affecting sperm parameters. Arch. Appl. Sci. Res. 2011, 3, 618–623. [Google Scholar]
- Ahmadian Mahmoudabadi, M.; Ghorbanmehr, N.; Movahedin, M.; Elikaei, A. Effects of Escherichia coli and Staphylococcus aureus Inoculation on Human Sperm Parameters. Navid. No. 2020, 23, 1–13. [Google Scholar]
- El-Mulla, K.F.; Köhn, F.M.; Dandal, M.; el Beheiry, A.H.; Schiefer, H.G.; Weidner, W.; Schill, W.B. In vitro effect of Escherichia coli on human sperm acrosome reaction. Arch. Androl. 1996, 37, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Zuleta-González, M.C.; Zapata-Salazar, M.E.; Guerrero-Hurtado, L.S.; Puerta-Suárez, J.; Cardona-Maya, W.D. Klebsiella pneumoniae and Streptococcus agalactiae: Passengers in the sperm travel. Arch. Esp. Urol. 2019, 72, 939–947. [Google Scholar]
- Weng, S.L.; Chiu, C.M.; Lin, F.M.; Huang, W.C.; Liang, C.; Yang, T.; Yang, T.L.; Liu, C.Y.; Wu, W.Y.; Chang, Y.A.; et al. Bacterial communities in semen from men of infertile couples: Metagenomic sequencing reveals relationships of seminal microbiota to semen quality. PLoS ONE 2014, 9, e110152. [Google Scholar] [CrossRef] [Green Version]
- Sepúlveda, L.; Bussalleu, E.; Yeste, M.; Bonet, S. Effect of Pseudomonas aeruginosa on sperm capacitation and protein phosphorylation of boar spermatozoa. Theriogenology 2016, 85, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.Y.; An, L.M.; Li, W.W.; Li, K.; Shao, Y.; Shang, X.J.; Yao, B.; Cui, Y.X.; Huang, Y.F. Ureaplasma urealyticum infection affects sperm plasma membrane integrity in infertile men. Zhonghua Nan Ke Xue 2011, 17, 1069–1072. [Google Scholar] [PubMed]
- Zhou, Y.H.; Ma, H.X.; Shi, X.X.; Liu, Y. Ureaplasma spp. in male infertility and its relationship with semen quality and seminal plasma components. J. Microbiol. Immunol. Infect. 2018, 51, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Al-Jebouri, M.; Mohamed, A. A Study on Infertility of Males Infected with Mycoplasma hominis with Reference to Sperm Morphology. Open J. Pathol. 2021, 11, 7–21. [Google Scholar] [CrossRef]
- Tjagur, S.; Mändar, R.; Poolamets, O.; Pomm, K.; Punab, M. Mycoplasma genitalium Provokes Seminal Inflammation among Infertile Males. Int. J. Mol. Sci. 2021, 22, 13467. [Google Scholar] [CrossRef]
- Gallegos, G.; Ramos, B.; Santiso, R.; Goyanes, V.; Gosálvez, J.; Fernández, J.L. Sperm DNA fragmentation in infertile men with genitourinary infection by Chlamydia trachomatis and Mycoplasma. Fertil. Steril. 2008, 90, 328–334. [Google Scholar] [CrossRef]
- Svenstrup, H.F.; Fedder, J.; Abraham-Peskir, J.; Birkelund, S.; Christiansen, G. Mycoplasma genitalium attaches to human spermatozoa. Hum Reprod. 2003, 18, 2103–2109. [Google Scholar] [CrossRef] [Green Version]
- Moazenchi, M.; Totonchi, M.; Salman Yazdi, R.; Hratian, K.; Mohseni Meybodi, M.A.; Ahmadi Panah, M.; Chehrazi, M.; Mohseni Meybodi, A. The impact of Chlamydia trachomatis infection on sperm parameters and male fertility: A comprehensive study. Int. J. STD AIDS. 2018, 29, 466–473. [Google Scholar] [CrossRef]
- Ammar-Keskes, L.; Gdoura, R.; Bouzid, F.; Ben Salah, F.; Sellami, D.; Hakim, H.; Hammami, A.; Rebaï, T.; Rekik, S.; Orfila, J. Retentissement de l’infection genitale a Chlamydia trachomatis sur le sperme chez les hommes consultant pour infertilite du couple. Andrologie 1998, 8, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, D.J.; Belenky, P.A.; Yang, J.H.; MacDonald, I.C.; Martell, J.D.; Takahashi, N.; Chan, C.T.; Lobritz, M.A.; Braff, D.; Schwarz, E.G.; et al. Antibiotics induce redox-related physiological alterations as part of their lethality. Proc. Natl. Acad. Sci. USA 2014, 111, E2100–E2109. [Google Scholar] [CrossRef] [Green Version]
- Veeraraghavan, B.; Bakthavatchalam, Y.D.; Sahni, R.D. Oral Antibiotics in Clinical Development for Community-Acquired Urinary Tract Infections. Infect Dis. Ther. 2021, 10, 1815–1835. [Google Scholar] [CrossRef]
- Santos, C.S.; Silva, A.R. Current and alternative trends in antibacterial agents used in mammalian semen technology. Anim. Reprod. 2020, 17, e20190111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.J.; Sharma, S. Effect Of Antibiotic Therapy On Sperm Quality. Eur. J. Mol. Clin. Med. 2020, 7, 4398–4403. [Google Scholar]
- Tímermans, A.; Vázquez, R.; Otero, F.; Gosálvez, J.; Johnston, S.; Fernández, J.L. Antibiotic toxicity on human spermatozoa assessed using the sperm DNA fragmentation dynamic assay. Andrologia 2022, 54, e14328. [Google Scholar] [CrossRef] [PubMed]
- Khaki, A.; Novin, M.G.; Khaki, A.A.; Nouri, M.; Sanati, E.; Nikmanesh, M. Comparative study of the effects of gentamicin, neomycin, streptomycin and ofloxacin antibiotics on sperm parameters and testis apoptosis in rats. Pak. J. Biol. Sci. 2008, 11, 1683–1689. [Google Scholar] [CrossRef] [Green Version]
- Costinar, L.; Herman, V.; Pitoiu, E.; Iancu, I.; Degi, J.; Hulea, A.; Pascu, C. Boar Semen Contamination: Identification of Gram-Negative Bacteria and Antimicrobial Resistance Profile. Animals 2022, 12, 43. [Google Scholar] [CrossRef]
- Dalmutt, A.C.; Moreno, L.Z.; Gomes, V.T.M.; Cunha, M.P.V.; Barbosa, M.R.F.; Sato, M.I.Z.; Pedroso, A.C.; Moreno, A.M. Characterization of bacterial contaminants of boar semen: Identification by MALDI-TOF mass spectrometry and antimicrobial susceptibility profiling. J. App. Anim. Res. 2020, 48, 559–565. [Google Scholar] [CrossRef]
- Goularte, K.L.; Voloski, F.; Redú, J.; Ferreira, C.; Vieira, A.D.; Duval, E.H.; Mondadori, R.G.; Lucia, T., Jr. Antibiotic resistance in microorganisms isolated in a bull semen stud. Reprod. Dom. Anim. 2020, 55, 318–324. [Google Scholar] [CrossRef]
- Al-Jebouri, M.M.; Mdish, S.A. Tracing of antibiotic-resistant bacteria isolated from semen of Iraqi males with primary infertility. Open J. Urol. 2019, 9, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Faisal, A.J.; Salman, H.A. Determination of semen quality and antibacterial susceptibility pattern of bacteria isolated from semen of Iraqi subjects. Microbiol. Biotechnol. Lett. 2021, 49, 587–593. [Google Scholar] [CrossRef]
- Kastrop, P.M.; de Graaf-Miltenburg, L.A.; Gutknecht, D.R.; Weima, S.M. Microbial contamination of embryo cultures in an ART laboratory: Sources and management. Hum. Reprod. 2007, 22, 2243–2248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machen, G.L.; Bird, E.T.; Brown, M.L.; Ingalsbe, D.A.; East, M.M.; Reyes, M.; Kuehl, T.J. Time trends for bacterial species and resistance patterns in semen in patients undergoing evaluation for male infertility. Proceedings (Bayl. Univ. Med. Cent.) 2018, 31, 165–167. [Google Scholar] [CrossRef] [PubMed]
- Fourie, J.; Loskutoff, N.; Huyser, C. Elimination of bacteria from human semen during sperm preparation using density gradient centrifugation with a novel tube insert. Andrologia 2012, 44, 513–517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Kass, Z.; Spergser, J.; Aurich, C.; Kuhl, J.; Schmidt, K.; Johannisson, A.; Morrell, J.M. Sperm Quality during Storage Is Not Affected by the Presence of Antibiotics in EquiPlus Semen Extender but Is Improved by Single Layer Centrifugation. Antibiotics 2018, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, A.H.; Ajina, T.; Ali, M.B.; Mehdi, M. Efficacy of density gradient centrifugation technique (DGC) in enhancing sperm cell DNA quality for assisted reproductive technique. Middle East Fertil. Soc. J. 2022, 27, 22. [Google Scholar] [CrossRef]
- Larentis, G.R.; Camozzato, G.C.; Bastos, H.B.A.; Gregory, R.M.; Mattos, R.C. Equine Sperm Selection by Synthetic Membrane Filter. J. Equine Vet. Sci. 2018, 63, 69–73. [Google Scholar] [CrossRef]
- Pacheco, A.; Blanco, A.; Bronet, F.; Cruz, M.; García-Fernández, J.; García-Velasco, J.A. Magnetic-Activated Cell Sorting (MACS): A Useful Sperm-Selection Technique in Cases of High Levels of Sperm DNA Fragmentation. J. Clin. Med. 2020, 9, 3976. [Google Scholar] [CrossRef]
- Tsakmakidis, I.A.; Samaras, T.; Anastasiadou, S.; Basioura, A.; Ntemka, A.; Michos, I.; Simeonidis, K.; Karagiannis, I.; Tsousis, G.; Angelakeris, M.; et al. Iron Oxide Nanoparticles as an Alternative to Antibiotics Additive on Extended Boar Semen. Nanomaterials 2020, 10, 1568. [Google Scholar] [CrossRef]
- Yousef, M.S.; Abdelhamid, H.N.; Hidalgo, M.; Fathy, R.; Gómez-Gascón, L.; Dorado, J. Antimicrobial activity of silver-carbon nanoparticles on the bacterial flora of bull semen. Theriogenology 2021, 161, 219–227. [Google Scholar] [CrossRef]
- Adesina, S.K.; Idowu, O.; Ogundaini, A.O.; Oladimeji, H.; Olugbade, T.A.; Onawunmi, G.O.; Pais, M. Antimicrobial constituents of the leaves of Acalypha wilkesiana and Aacalypha hispida. Phytother. Res. 2000, 14, 371–374. [Google Scholar] [CrossRef]
- Trinh, P.C.; Thao, L.T.; Ha, H.T.; Nguyen, T. DPPH-Scavenging and Antimicrobial Activities of Asteraceae Medicinal Plants on Uropathogenic Bacteria. Evid.-Based Complement. Altern. Med. 2020, 2020, 7807026. [Google Scholar] [CrossRef] [PubMed]
- Nweze, E.I.; Eze, E.E. Justification for the use of Ocimum gratissimum L in herbal medicine and its interaction with disc antibiotics. BMC Complement. Altern. Med. 2009, 9, 37. [Google Scholar] [CrossRef]
- Mensah, J.L.; Lagarde, I.; Ceschin, C.; Michel, G.; Gleye, J.; Fouraste, I. Antibacterial activity of the leaves of Phyllanthus discoideus. J. Ethnopharmacol. 1990, 28, 129–133. [Google Scholar] [CrossRef]
- Olajuyigbe, O.O.; Adedayo, O.; Coopoosamy, R.M. Antibacterial Activity of Defatted and Nondefatted Methanolic Extracts of Aframomum melegueta K. Schum. against Multidrug-Resistant Bacteria of Clinical Importance. Sci. World J. 2020, 2020, 4808432. [Google Scholar] [CrossRef] [PubMed]
- Cock, I.; Mavuso, N.; Van Vuuren, S. A Review of Plant-Based Therapies for the Treatment of Urinary Tract Infections in Traditional Southern African Medicine. Evid.-Based Complement. Altern. Med. 2021, 2021, 7341124. [Google Scholar] [CrossRef]
- Noh, S.; Go, A.; Kim, D.B.; Park, M.; Jeon, H.W.; Kim, B. Role of Antioxidant Natural Products in Management of Infertility: A Review of Their Medicinal Potential. Antioxidants 2020, 9, 957. [Google Scholar] [CrossRef]
- Sethi, J.; Yadav, M.; Sood, S.; Dahiya, K.; Singh, V. Effect of tulsi (Ocimum Sanctum Linn.) on sperm count and reproductive hormones in male albino rabbits. Int. J. Ayurveda Res. 2010, 1, 208–210. [Google Scholar] [CrossRef] [Green Version]
- Khaki, A. Effect of Cinnamomum zeylanicumon on Spermatogenesis. Iran. Red. Crescent. Med. J. 2015, 17, e18668. [Google Scholar] [CrossRef] [Green Version]
- Ros-Santaella, J.L.; Pintus, E. Plant Extracts as Alternative Additives for Sperm Preservation. Antioxidants 2021, 10, 772. [Google Scholar] [CrossRef]
- Tvrdá, E.; Michalko, J.; Árvay, J.; Vukovic, N.L.; Ivanišová, E.; Ďuračka, M.; Matušíková, I.; Kačániová, M. Characterization of the Omija (Schisandra chinensis) Extract and Its Effects on the Bovine Sperm Vitality and Oxidative Profile during In Vitro Storage. Evid. Based Complement. Alternat. Med. 2020, 2020, 7123780. [Google Scholar] [CrossRef]
- Efstratiou, E.; Hussain, A.I.; Nigam, P.S.; Moore, J.E.; Ayub, M.A.; Rao, J.R. Antimicrobial activity of Calendula officinalis petal extracts against fungi, as well as Gram-negative and Gram-positive clinical pathogens. Complement Ther. Clin. Pract. 2012, 18, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Bneko, F.; Palkovičová, V.; Ďuračka, M.; Árvay, J.; Lukáč, N.; Tvrdaá, E. Antioxidant Effects of Marigold (Calendula officinalis) Flower Extract on the Oxidative Balance of Bovine Spermatozoa. Contemp. Agric. 2019, 68, 92–102. [Google Scholar] [CrossRef]
- Elmi, A.; Prosperi, A.; Zannoni, A.; Bertocchi, M.; Scorpio, D.G.; Forni, M.; Foni, E.; Bacci, M.L.; Ventrella, D. Antimicrobial capabilities of non-spermicidal concentrations of tea tree (Melaleuca alternifolia) and rosemary (Rosmarinus officinalis) essential oils on the liquid phase of refrigerated swine seminal doses. Res. Vet. Sci. 2019, 127, 76–81. [Google Scholar] [CrossRef]
- Nimrat, S.; Noppakun, S.; Sripuak, K.; Boonthai, T.; Vuthiphandchai, V. Cryopreservation of banana shrimp (Fenneropenaeus merguiensis) spermatophores with supplementation of medicinal plant extracts: Development of a programmable controlled-rate method and a practical method. Aquaculture 2020, 515, 734537. [Google Scholar] [CrossRef]
Figure 1.
Sources of ROS in normal (a), immature (b) or dead (c) spermatozoa.

Figure 2.
Sources of ROS in aerobic bacteria (a) and leukocytes (b).

Figure 3.
Damage to the sperm cell caused by (a) hemolysins or (b) lipopolysaccharide (LPS).


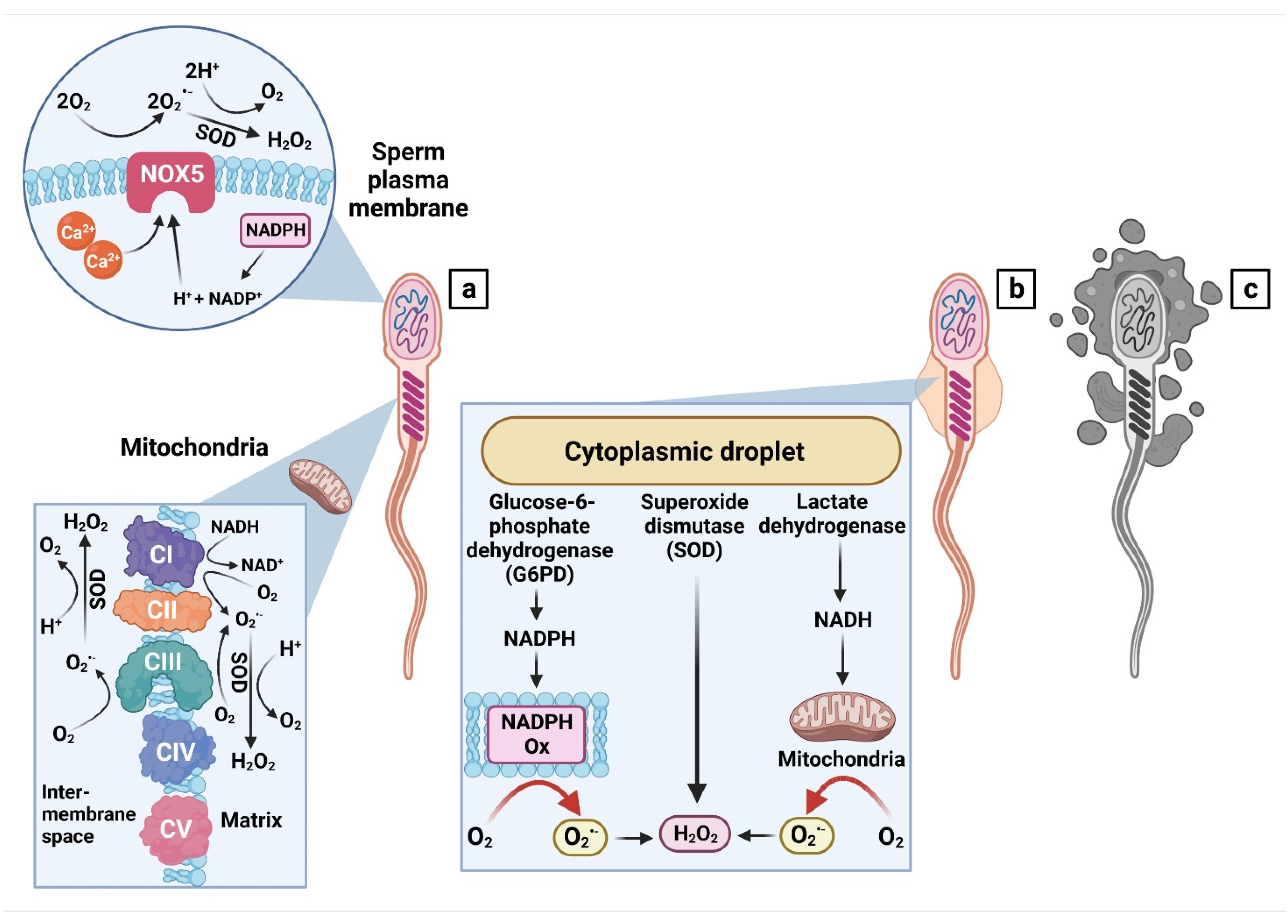{kind=link}
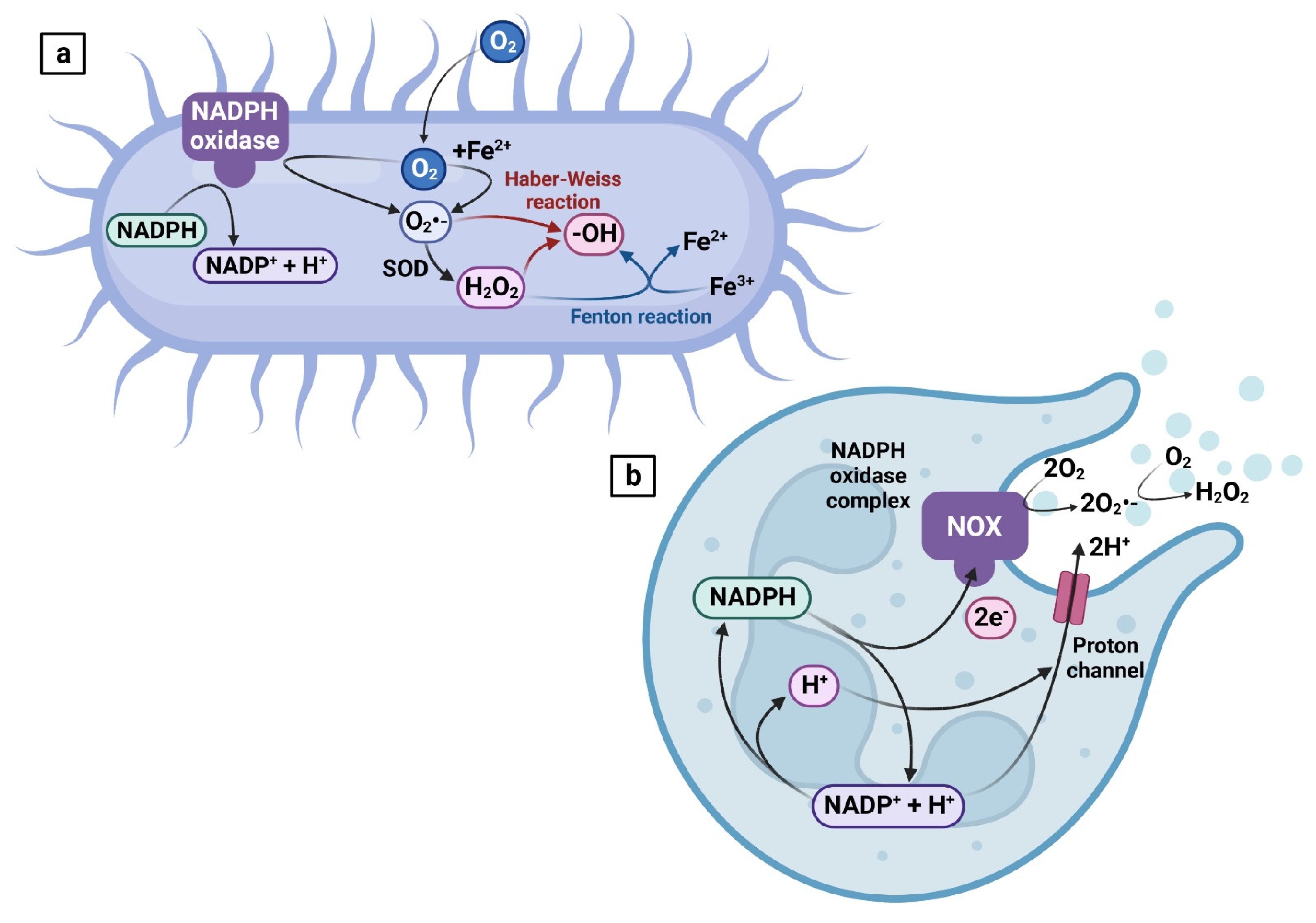{kind=link}
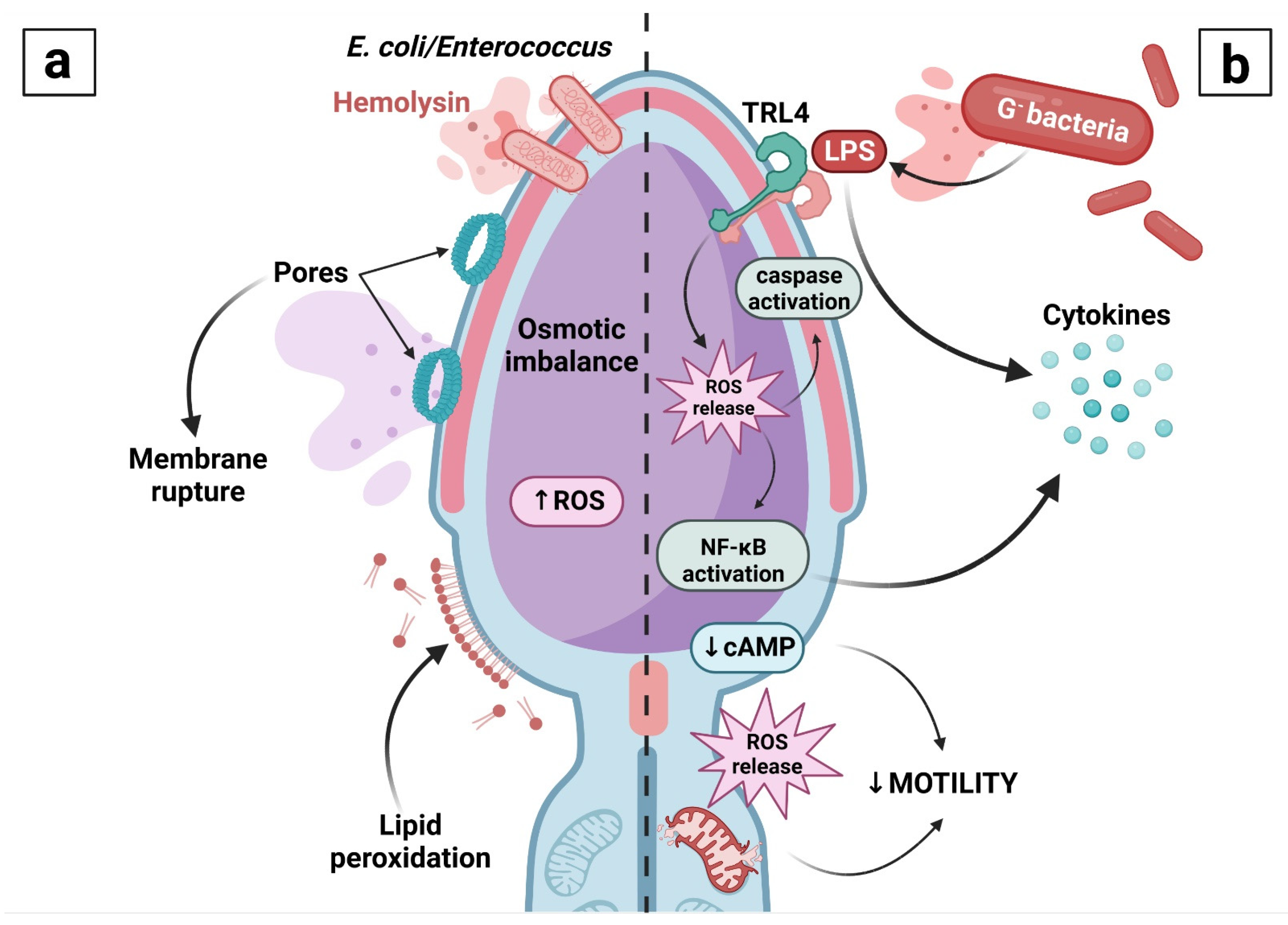{kind=link}
Table 1.
Leading consensual in vivo and in vitro effects of the most frequent uropathogenic bacteria on semen quality and/or sperm structural integrity and functional activity.
Table 1.
Leading consensual in vivo and in vitro effects of the most frequent uropathogenic bacteria on semen quality and/or sperm structural integrity and functional activity.
| Bacterium | In Vivo Observations | In Vitro Observations |
|---|---|---|
| Escherichia coli | ↓ sperm count [8,9,23,126,136,137] ↓ sperm motility [8,9,10,23,126,136,137,138] ↓ sperm viability [8,9,23,126,136] ↓ sperm morphology [9,136] ↓ acrosome integrity [8,23,126] ↑ DNA damage [8,9,23] ↓ chromatin integrity [8] ↑ leukocytospermia [8,9,23,44,126,137] ↑ concentration of cytokines [8,23,126] ↓ concentration of antibacterial proteins [8,126] | ↓ sperm motility and viability [20,21,133,139] ↓ sperm morphology [139] ↓ acrosome integrity [140] ↑ caspase 3 and 7 activity [20] |
| Klebsiella pneumoniae | ↓ sperm count, motility, viability and acrosome integrity [126] ↓ mitochondrial activity [126] ↑ DNA damage [8,9,126] ↑ leukocytospermia and cytokine levels [126] ↓ concentration of antibacterial proteins [126] | ↓ sperm motility [20,21,141] ↓ sperm viability [20,21] ↓ sperm morphology [21] ↑ caspase 3 and 7 activity [20] |
| Pseudomonas aeruginosa | ↓ sperm count, motility and morphology [128,142] ↓ sperm viability [122,128] | ↓ sperm motility [143] ↓ sperm viability [20] ↑ caspase 3 and 7 activity [20] |
| Enterococcus spp. | ↓ sperm count, motility and acrosome integrity [8,9] ↓ sperm viability [8,9] ↓ sperm morphology [9] ↓ mitochondrial activity [8,9,126] ↑ sperm apoptosis and phospholipid scrambling [9] ↑ DNA damage [8,9,126] ↓ chromatin integrity [8,86,126] ↑ leukocytospermia and cytokine levels [8,9,126] ↓ concentration of antibacterial proteins [8,126] | ↓ sperm motility and viability [20,90] ↓ acrosome integrity [90] ↓ mitochondrial activity [90] ↑ caspase 3 and 7 activity [20] ↓ chromatin integrity [90] |
| Staphylococcus spp. | ↓ sperm count and motility [8,9,23,127] ↓ sperm viability [8,9,127] ↓ sperm morphology [9] ↓ acrosome integrity [8,23,127] ↓ mitochondrial activity [8,9,23,127] ↑ sperm apoptosis and phospholipid scrambling [9] ↓ chromatin integrity [8,23,127] ↑ leukocytospermia [8,9,23,127] ↑ concentration of cytokines [8,23,127] ↓ concentration of antibacterial proteins [8] | ↓ sperm motility and viability [41,133,139] ↓ sperm morphology [139] ↓ acrosome integrity [41] ↓ mitochondrial activity [41,133] |
| Campylobacter ureolyticus | ↓ sperm count [9,137] ↓ sperm motility and viability [9] ↓ cell morphology [9,137] ↓ mitochondrial activity [9] ↑ sperm apoptosis, DNA damage and phospholipid scrambling [9] ↑ leukocytospermia [9,137] | N/A |
| Ureaplasma urealyticum | ↓ sperm count and motility [9,144,145] ↓ sperm viability [9] ↓ sperm morphology [9,144] ↓ mitochondrial activity [9] ↑ sperm apoptosis, DNA damage and phospholipid scrambling [9] ↑ leukocytospermia [9] | ↓ sperm motility and viability [133] |
| Mycoplasma spp. | ↓ sperm count and motility [9,146,147,148] ↓ sperm morphology [9,146,148] ↓ sperm viability [9,146] ↑ sperm apoptosis, DNA damage and phospholipid scrambling [9] ↓ chromatin integrity [148] ↑ leukocytospermia [9,147] ↑ concentration of cytokines [147] | ↓ sperm motility and morphology [149] |
| Chlamydia trachomatis | ↓ sperm count, motility and morphology [148,150,151] ↓ sperm viability [151] ↓ chromatin integrity [148,150] | N/A |
Table 2.
Leading consensual in vivo and in vitro effects of the most frequent uropathogenic bacteria on oxidative characteristics of semen and/or spermatozoa.
Table 2.
Leading consensual in vivo and in vitro effects of the most frequent uropathogenic bacteria on oxidative characteristics of semen and/or spermatozoa.
| Bacterium | In Vivo Observations | In Vitro Observations |
|---|---|---|
| Escherichia coli | ↑ intracellular ROS production [23,126] ↑ superoxide production [9] ↓ antioxidant capacity [8,23,126] ↑ LPO [8,9,23,126] ↑ protein oxidation [23,126] | ↑ intracellular ROS production [21,22] ↑ mitochondrial ROS production [20] ↑ LPO [22] |
| Klebsiella pneumoniae | ↑ intracellular ROS production [126] ↓ antioxidant capacity [126] ↑ LPO and protein oxidation [126] | ↑ mitochondrial ROS production [20] ↑ LPO [141] |
| Pseudomonas aeruginosa | N/A | ↑ mitochondrial ROS production [20] ↑ LPO [20] |
| Enterococcus spp. | ↑ intracellular ROS production [126] ↑ superoxide production [9] ↓ antioxidant capacity [8,126] ↑ LPO [8,9,126] ↑ protein oxidation [126] | ↑ intracellular ROS production [90] ↑ mitochondrial ROS production [20] |
| Staphylococcus spp. | ↑ superoxide production [9] ↓ antioxidant capacity [8,23,127] ↑ LPO [8,9,23,127] ↑ protein oxidation [23,127] | ↑ intracellular ROS production [22,41] ↑ LPO [22] |
| Campylobacter ureolyticus | ↑ superoxide production [9] ↑ LPO [9,137] | ↑ intracellular ROS production [22] ↑ LPO [22] |
| Ureaplasma urealyticum | ↑ superoxide production [9] ↑ LPO [9] | N/A |
| Mycoplasma spp. | ↑ superoxide production [9] ↑ LPO [9] | N/A |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tvrdá, E.; Benko, F.; Ďuračka, M. Oxidative Stress as an Underlying Mechanism of Bacteria-Inflicted Damage to Male Gametes. Oxygen 2022, 2, 547-569. https://doi.org/10.3390/oxygen2040036
AMA Style
Tvrdá E, Benko F, Ďuračka M. Oxidative Stress as an Underlying Mechanism of Bacteria-Inflicted Damage to Male Gametes. Oxygen. 2022; 2(4):547-569. https://doi.org/10.3390/oxygen2040036
Chicago/Turabian StyleTvrdá, Eva, Filip Benko, and Michal Ďuračka. 2022. "Oxidative Stress as an Underlying Mechanism of Bacteria-Inflicted Damage to Male Gametes" Oxygen 2, no. 4: 547-569. https://doi.org/10.3390/oxygen2040036