Are the Gut Bacteria Telling Us to Eat or Not to Eat? Reviewing the Role of Gut Microbiota in the Etiology, Disease Progression and Treatment of Eating Disorders



Abstract
:1. Introduction
2. Overview of Eating Disorders: Classic Etiology, Progression, and Treatment
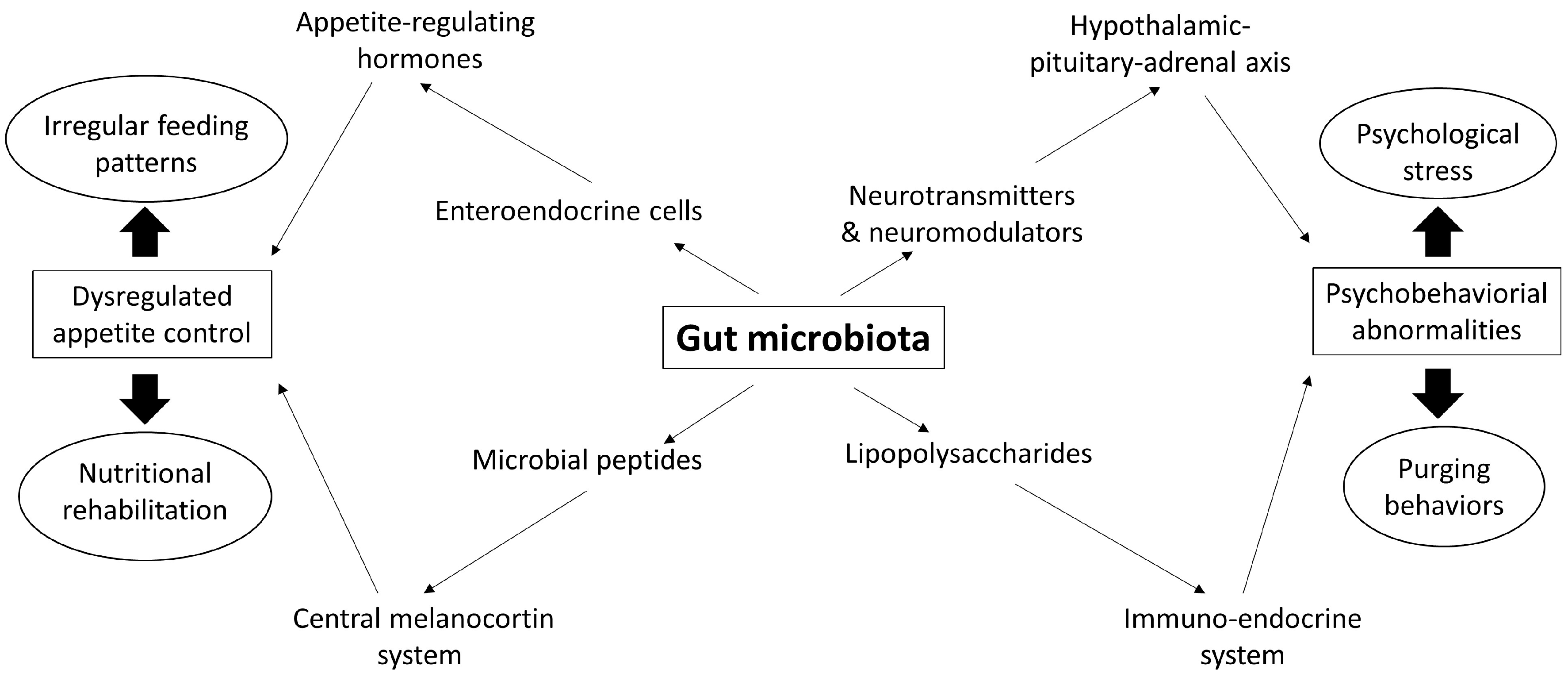
3. Gut Microbiota: A Missing Piece in Eating Disorders
3.1. Dysbiosis in Eating Disorders
3.2. The Microbiota-Gut-Brain Axis
3.2.1. Effects of Gut Microbiota on Appetite Control
3.2.2. Effects of Gut Microbiota on Brain Function and Behavior
3.2.3. Effects of the Brain on Gut Microbiota
3.3. Effects of Eating Disorder-Related Behaviors on Gut Microbiota
3.4. Effects of Nutritional Rehabilitation on Gut Microbiota
3.5. Mechanisms Underlying the Microbiota-Induced Dysmetabolism in Eating Disorders
3.5.1. Immune Function
3.5.2. Short-Chain Fatty Acid Production
3.5.3. Gut Barrier Function
4. Therapeutic Potential of Microbiome-Based Treatments for Eating Disorders
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mitchison, D.; Hay, P.; Slewa-Younan, S.; Mond, J. Time trends in population prevalence of eating disorder behaviors and their relationship to quality of life. PLoS ONE 2012, 7, e48450. [Google Scholar] [CrossRef]
- Mitchison, D.; Hay, P.; Slewa-Younan, S.; Mond, J. The changing demographic profile of eating disorder behaviors in the community. BMC Public Health 2014, 14, 943. [Google Scholar] [CrossRef] [PubMed]
- Stice, E.; Marti, C.N.; Rohde, P. Prevalence, incidence, impairment, and course of the proposed DSM-5 eating disorder diagnoses in an 8-year prospective community study of young women. J. Abnorm. Psychol. 2013, 122, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Samnaliev, M.; Noh, H.L.; Sonneville, K.R.; Austin, S.B. The economic burden of eating disorders and related mental health comorbidities: An exploratory analysis using the U.S. Medical Expenditures Panel Survey. Prev. Med. Rep. 2015, 2, 32–34. [Google Scholar] [CrossRef] [PubMed]
- Stuhldreher, N.; Konnopka, A.; Wild, B.; Herzog, W.; Zipfel, S.; Lowe, B.; Konig, H.H. Cost-of-illness studies and cost-effectiveness analyses in eating disorders: A systematic review. Int. J. Eat. Disord. 2012, 45, 476–491. [Google Scholar] [CrossRef] [PubMed]
- Zabala, M.J.; Macdonald, P.; Treasure, J. Appraisal of caregiving burden, expressed emotion and psychological distress in families of people with eating disorders: A systematic review. Eur. Eat. Disord. Rev. 2009, 17, 338–349. [Google Scholar] [CrossRef]
- Graap, H.; Bleich, S.; Herbst, F.; Scherzinger, C.; Trostmann, Y.; Wancata, J.; de Zwaan, M. The needs of carers: A comparison between eating disorders and schizophrenia. Soc. Psychiatry Psychiatr. Epidemiol. 2008, 43, 800–807. [Google Scholar] [CrossRef]
- American Psychiatric Association. The Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Arlington, VA, USA, 2013. [Google Scholar]
- Lilenfeld, L.R.; Kaye, W.H.; Greeno, C.G.; Merikangas, K.R.; Plotnicov, K.; Pollice, C.; Rao, R.; Strober, M.; Bulik, C.M.; Nagy, L. A controlled family study of anorexia nervosa and bulimia nervosa: Psychiatric disorders in first-degree relatives and effects of proband comorbidity. Arch. Gen. Psychiatry 1998, 55, 603–610. [Google Scholar] [CrossRef] [PubMed]
- Strober, M.; Freeman, R.; Lampert, C.; Diamond, J.; Kaye, W. Controlled family study of anorexia nervosa and bulimia nervosa: Evidence of shared liability and transmission of partial syndromes. Am. J. Psychiatry 2000, 157, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Trace, S.E.; Baker, J.H.; Penas-Lledo, E.; Bulik, C.M. The genetics of eating disorders. Annu. Rev. Clin. Psychol. 2013, 9, 589–620. [Google Scholar] [CrossRef] [PubMed]
- Bulik, C.M.; Sullivan, P.F.; Tozzi, F.; Furberg, H.; Lichtenstein, P.; Pedersen, N.L. Prevalence, heritability, and prospective risk factors for anorexia nervosa. Arch. Gen. Psychiatry 2006, 63, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Burt, S.A.; McGue, M.; Iacono, W.G. Changes in genetic and environmental influences on disordered eating across adolescence: A longitudinal twin study. Arch. Gen. Psychiatry 2007, 64, 1409–1415. [Google Scholar] [CrossRef] [PubMed]
- Klump, K.L.; Keel, P.K.; Sisk, C.; Burt, S.A. Preliminary evidence that estradiol moderates genetic influences on disordered eating attitudes and behaviors during puberty. Psychol. Med. 2010, 40, 1745–1753. [Google Scholar] [CrossRef] [PubMed]
- Culbert, K.M.; Burt, S.A.; McGue, M.; Iacono, W.G.; Klump, K.L. Puberty and the genetic diathesis of disordered eating attitudes and behaviors. J. Abnorm. Psychol. 2009, 118, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Bailer, U.F.; Kaye, W.H. A review of neuropeptide and neuroendocrine dysregulation in anorexia and bulimia nervosa. Curr. Drug Targets CNS Neurol. Disord. 2003, 2, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Kaye, W.H.; Wierenga, C.E.; Bailer, U.F.; Simmons, A.N.; Bischoff-Grethe, A. Nothing tastes as good as skinny feels: The neurobiology of anorexia nervosa. Trends Neurosci. 2013, 36, 110–120. [Google Scholar] [CrossRef] [PubMed]
- Stice, E. Interactive and Mediational Etiologic Models of Eating Disorder Onset: Evidence from Prospective Studies. Annu. Rev. Clin. Psychol. 2016, 12, 359–381. [Google Scholar] [CrossRef] [PubMed]
- Bakalar, J.L.; Shank, L.M.; Vannucci, A.; Radin, R.M.; Tanofsky-Kraff, M. Recent Advances in Developmental and Risk Factor Research on Eating Disorders. Curr. Psychiatry Rep. 2015, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Torsello, A.; Brambilla, F.; Tamiazzo, L.; Bulgarelli, I.; Rapetti, D.; Bresciani, E.; Locatelli, V. Central dysregulations in the control of energy homeostasis and endocrine alterations in anorexia and bulimia nervosa. J. Endocrinol. Investig. 2007, 30, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Zipfel, S.; Sammet, I.; Rapps, N.; Herzog, W.; Herpertz, S.; Martens, U. Gastrointestinal disturbances in eating disorders: Clinical and neurobiological aspects. Auton. Neurosci. 2006, 129, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Campbell, K.; Peebles, R. Eating disorders in children and adolescents: State of the art review. Pediatrics 2014, 134, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Nazar, B.P.; Gregor, L.K.; Albano, G.; Marchica, A.; Coco, G.L.; Cardi, V.; Treasure, J. Early Response to treatment in Eating Disorders: A Systematic Review and a Diagnostic Test Accuracy Meta-Analysis. Eur. Eat. Disord. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- American Academy of Pediatrics; Committee on Adolescence. Identifying and treating eating disorders. Pediatrics 2003, 111, 204–211. [Google Scholar]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Budhram, A.; Parvathy, S.; Kremenchutzky, M.; Silverman, M. Breaking down the gut microbiome composition in multiple sclerosis. Mult. Scler. 2016, 23, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Pistollato, F.; Sumalla Cano, S.; Elio, I.; Masias Vergara, M.; Giampieri, F.; Battino, M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr. Rev. 2016, 74, 624–634. [Google Scholar] [CrossRef] [PubMed]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef] [PubMed]
- Morita, C.; Tsuji, H.; Hata, T.; Gondo, M.; Takakura, S.; Kawai, K.; Yoshihara, K.; Ogata, K.; Nomoto, K.; Miyazaki, K.; et al. Gut Dysbiosis in Patients with Anorexia Nervosa. PLoS ONE 2015, 10, e0145274. [Google Scholar] [CrossRef] [PubMed]
- Kleiman, S.C.; Watson, H.J.; Bulik-Sullivan, E.C.; Huh, E.Y.; Tarantino, L.M.; Bulik, C.M.; Carroll, I.M. The Intestinal Microbiota in Acute Anorexia Nervosa and During Renourishment: Relationship to Depression, Anxiety, and Eating Disorder Psychopathology. Psychosom. Med. 2015, 77, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef] [PubMed]
- Mack, I.; Cuntz, U.; Gramer, C.; Niedermaier, S.; Pohl, C.; Schwiertz, A.; Zimmermann, K.; Zipfel, S.; Enck, P.; Penders, J. Weight gain in anorexia nervosa does not ameliorate the faecal microbiota, branched chain fatty acid profiles, and gastrointestinal complaints. Sci. Rep. 2016, 6, 26752. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.G.; Bloom, S.R. Gut hormones and the regulation of energy homeostasis. Nature 2006, 444, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Holzer, P.; Reichmann, F.; Farzi, A. Neuropeptide, Y, peptide YY and pancreatic polypeptide in the gut-brain axis. Neuropeptides 2012, 46, 261–274. [Google Scholar] [CrossRef] [PubMed]
- Levy, R.L.; Olden, K.W.; Naliboff, B.D.; Bradley, L.A.; Francisconi, C.; Drossman, D.A.; Creed, F. Psychosocial aspects of the functional gastrointestinal disorders. Gastroenterology 2006, 130, 1447–1458. [Google Scholar] [CrossRef] [PubMed]
- Garakani, A.; Win, T.; Virk, S.; Gupta, S.; Kaplan, D.; Masand, P.S. Comorbidity of irritable bowel syndrome in psychiatric patients: A review. Am. J. Ther. 2003, 10, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Raybould, H.E. Gut chemosensing: Interactions between gut endocrine cells and visceral afferents. Auton. Neurosci. 2010, 153, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.D.; Chen, J.; Hoffmann, C.; Bittinger, K.; Chen, Y.Y.; Keilbaugh, S.A.; Bewtra, M.; Knights, D.; Walters, W.A.; Knight, R.; et al. Linking long-term dietary patterns with gut microbial enterotypes. Science 2011, 334, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Alcock, J.; Maley, C.C.; Aktipis, C.A. Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays 2014, 36, 940–949. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Gray, A.M.; Erickson, M.A.; Salameh, T.S.; Damodarasamy, M.; Sheibani, N.; Meabon, J.S.; Wing, E.E.; Morofuji, Y.; Cook, D.G.; et al. Lipopolysaccharide-induced blood-brain barrier disruption: Roles of cyclooxygenase, oxidative stress, neuroinflammation, and elements of the neurovascular unit. J. Neuroinflamm. 2015, 12, 223. [Google Scholar] [CrossRef] [PubMed]
- Wisse, B.E.; Ogimoto, K.; Tang, J.; Harris, M.K., Jr.; Raines, E.W.; Schwartz, M.W. Evidence that lipopolysaccharide-induced anorexia depends upon central, rather than peripheral, inflammatory signals. Endocrinology 2007, 148, 5230–5237. [Google Scholar] [CrossRef] [PubMed]
- Von Meyenburg, C.; Hrupka, B.H.; Arsenijevic, D.; Schwartz, G.J.; Landmann, R.; Langhans, W. Role for CD14, TLR2, and TLR4 in bacterial product-induced anorexia. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R298–R305. [Google Scholar] [CrossRef] [PubMed]
- Ruud, J.; Wilhelms, D.B.; Nilsson, A.; Eskilsson, A.; Tang, Y.J.; Strohle, P.; Caesar, R.; Schwaninger, M.; Wunderlich, T.; Backhed, F.; et al. Inflammation- and tumor-induced anorexia and weight loss require MyD88 in hematopoietic/myeloid cells but not in brain endothelial or neural cells. FASEB J. 2013, 27, 1973–1980. [Google Scholar] [CrossRef] [PubMed]
- Fetissov, S.O.; Hallman, J.; Oreland, L.; Af Klinteberg, B.; Grenback, E.; Hulting, A.L.; Hokfelt, T. Autoantibodies against alpha -MSH, ACTH, and LHRH in anorexia and bulimia nervosa patients. Proc. Natl. Acad. Sci. USA 2002, 99, 17155–17160. [Google Scholar] [CrossRef] [PubMed]
- Fetissov, S.O.; Harro, J.; Jaanisk, M.; Jarv, A.; Podar, I.; Allik, J.; Nilsson, I.; Sakthivel, P.; Lefvert, A.K.; Hokfelt, T. Autoantibodies against neuropeptides are associated with psychological traits in eating disorders. Proc. Natl. Acad. Sci. USA 2005, 102, 14865–14870. [Google Scholar] [CrossRef] [PubMed]
- Breton, J.; Tennoune, N.; Lucas, N.; Francois, M.; Legrand, R.; Jacquemot, J.; Goichon, A.; Guerin, C.; Peltier, J.; Pestel-Caron, M.; et al. Gut Commensal E. coli Proteins Activate Host Satiety Pathways following Nutrient-Induced Bacterial Growth. Cell Metab. 2016, 23, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Tennoune, N.; Chan, P.; Breton, J.; Legrand, R.; Chabane, Y.N.; Akkermann, K.; Jarv, A.; Ouelaa, W.; Takagi, K.; Ghouzali, I.; et al. Bacterial ClpB heat-shock protein, an antigen-mimetic of the anorexigenic peptide alpha-MSH, at the origin of eating disorders. Transl. Psychiatry 2014, 4, e458. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J.; et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol. Psychiatry 2016, 21, 786–796. [Google Scholar] [CrossRef] [PubMed]
- Vuong, H.E.; Hsiao, E.Y. Emerging Roles for the Gut Microbiome in Autism Spectrum Disorder. Biol. Psychiatry 2017, 81, 411–423. [Google Scholar] [CrossRef] [PubMed]
- Hoban, A.E.; Moloney, R.D.; Golubeva, A.V.; McVey Neufeld, K.A.; O’Sullivan, O.; Patterson, E.; Stanton, C.; Dinan, T.G.; Clarke, G.; Cryan, J.F. Behavioural and neurochemical consequences of chronic gut microbiota depletion during adulthood in the rat. Neuroscience 2016, 339, 463–477. [Google Scholar] [CrossRef] [PubMed]
- Luczynski, P.; McVey Neufeld, K.A.; Oriach, C.S.; Clarke, G.; Dinan, T.G.; Cryan, J.F. Growing up in a Bubble: Using Germ-Free Animals to Assess the Influence of the Gut Microbiota on Brain and Behavior. Int. J. Neuropsychopharmacol. 2016, 19. [Google Scholar] [CrossRef] [PubMed]
- Bercik, P.; Denou, E.; Collins, J.; Jackson, W.; Lu, J.; Jury, J.; Deng, Y.; Blennerhassett, P.; Macri, J.; McCoy, K.D.; et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology 2011, 141, 599–609, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Lo Sauro, C.; Ravaldi, C.; Cabras, P.L.; Faravelli, C.; Ricca, V. Stress, hypothalamic-pituitary-adrenal axis and eating disorders. Neuropsychobiology 2008, 57, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice. J. Physiol. 2004, 558 Pt 1, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Martich, G.D.; Boujoukos, A.J.; Suffredini, A.F. Response of man to endotoxin. Immunobiology 1993, 187, 403–416. [Google Scholar] [CrossRef]
- Redl, H.; Bahrami, S.; Schlag, G.; Traber, D.L. Clinical detection of LPS and animal models of endotoxemia. Immunobiology 1993, 187, 330–345. [Google Scholar] [CrossRef]
- Tilders, F.J.; DeRijk, R.H.; Van Dam, A.M.; Vincent, V.A.; Schotanus, K.; Persoons, J.H. Activation of the hypothalamus-pituitary-adrenal axis by bacterial endotoxins: Routes and intermediate signals. Psychoneuroendocrinology 1994, 19, 209–232. [Google Scholar] [CrossRef]
- Vedder, H.; Schreiber, W.; Yassouridis, A.; Gudewill, S.; Galanos, C.; Pollmacher, T. Dose-dependence of bacterial lipopolysaccharide (LPS) effects on peak response and time course of the immune-endocrine host response in humans. Inflamm. Res. 1999, 48, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Erickson, M.A. The blood-brain barrier and immune function and dysfunction. Neurobiol. Dis. 2010, 37, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.H.; Pothoulakis, C.; Mayer, E.A. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.T.; Sperandio, V. Inter-kingdom signalling: Communication between bacteria and their hosts. Nat. Rev. Microbiol. 2008, 6, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Cogan, T.A.; Thomas, A.O.; Rees, L.E.; Taylor, A.H.; Jepson, M.A.; Williams, P.H.; Ketley, J.; Humphrey, T.J. Norepinephrine increases the pathogenic potential of Campylobacter jejuni. Gut 2007, 56, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, S.M.; Marchesi, J.R.; Scully, P.; Codling, C.; Ceolho, A.M.; Quigley, E.M.; Cryan, J.F.; Dinan, T.G. Early life stress alters behavior, immunity, and microbiota in rats: Implications for irritable bowel syndrome and psychiatric illnesses. Biol. Psychiatry 2009, 65, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rodenas, C.L.; Bergonzelli, G.E.; Nutten, S.; Schumann, A.; Cherbut, C.; Turini, M.; Ornstein, K.; Rochat, F.; Corthesy-Theulaz, I. Nutritional approach to restore impaired intestinal barrier function and growth after neonatal stress in rats. J. Pediatr. Gastroenterol. Nutr. 2006, 43, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.T.; Coe, C.L. Maternal separation disrupts the integrity of the intestinal microflora in infant rhesus monkeys. Dev. Psychobiol. 1999, 35, 146–155. [Google Scholar] [CrossRef]
- Kiecolt-Glaser, J.K.; Preacher, K.J.; MacCallum, R.C.; Atkinson, C.; Malarkey, W.B.; Glaser, R. Chronic stress and age-related increases in the proinflammatory cytokine IL-6. Proc. Natl. Acad. Sci. USA 2003, 100, 9090–9095. [Google Scholar] [CrossRef] [PubMed]
- Gouin, J.P.; Glaser, R.; Malarkey, W.B.; Beversdorf, D.; Kiecolt-Glaser, J. Chronic stress, daily stressors, and circulating inflammatory markers. Health Psychol. 2012, 31, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Maukonen, J.; Saarela, M. Human gut microbiota: Does diet matter? Proc. Nutr. Soc. 2015, 74, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.S.; Walker, A.W.; Louis, P.; Parkhill, J.; Vermeiren, J.; Bosscher, D.; Duncan, S.H.; Flint, H.J. Modulation of the human gut microbiota by dietary fibres occurs at the species level. BMC Biol. 2016, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Dominianni, C.; Sinha, R.; Goedert, J.J.; Pei, Z.; Yang, L.; Hayes, R.B.; Ahn, J. Sex, body mass index, and dietary fiber intake influence the human gut microbiome. PLoS ONE 2015, 10, e0124599. [Google Scholar] [CrossRef] [PubMed]
- Secor, S.M.; Carey, H.V. Integrative Physiology of Fasting. Compr. Physiol. 2016, 6, 773–825. [Google Scholar] [PubMed]
- Kohl, K.D.; Amaya, J.; Passement, C.A.; Dearing, M.D.; McCue, M.D. Unique and shared responses of the gut microbiota to prolonged fasting: A comparative study across five classes of vertebrate hosts. FEMS Microbiol. Ecol. 2014, 90, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Monira, S.; Nakamura, S.; Gotoh, K.; Izutsu, K.; Watanabe, H.; Alam, N.H.; Endtz, H.P.; Cravioto, A.; Ali, S.I.; Nakaya, T.; et al. Gut microbiota of healthy and malnourished children in bangladesh. Front. Microbiol. 2011, 2, 228. [Google Scholar] [CrossRef]
- Smith, M.I.; Yatsunenko, T.; Manary, M.J.; Trehan, I.; Mkakosya, R.; Cheng, J.; Kau, A.L.; Rich, S.S.; Concannon, P.; Mychaleckyj, J.C.; et al. Gut microbiomes of Malawian twin pairs discordant for kwashiorkor. Science 2013, 339, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zeevi, D.; Levy, M.; Zilberman-Schapira, G.; Suez, J.; Tengeler, A.C.; Abramson, L.; Katz, M.N.; Korem, T.; Zmora, N.; et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell 2014, 159, 514–529. [Google Scholar] [CrossRef] [PubMed]
- Zarrinpar, A.; Chaix, A.; Yooseph, S.; Panda, S. Diet and feeding pattern affect the diurnal dynamics of the gut microbiome. Cell Metab. 2014, 20, 1006–1017. [Google Scholar] [CrossRef] [PubMed]
- Kasiraj, A.C.; Harmoinen, J.; Isaiah, A.; Westermarck, E.; Steiner, J.M.; Spillmann, T.; Suchodolski, J.S. The effects of feeding and withholding food on the canine small intestinal microbiota. FEMS Microbiol. Ecol. 2016, 92, fiw085. [Google Scholar] [CrossRef]
- Sato, Y.; Fukudo, S. Gastrointestinal symptoms and disorders in patients with eating disorders. Clin. J. Gastroenterol. 2015, 8, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Forney, K.J.; Buchman-Schmitt, J.M.; Keel, P.K.; Frank, G.K. The medical complications associated with purging. Int. J. Eat. Disord. 2016, 49, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Garber, A.K.; Sawyer, S.M.; Golden, N.H.; Guarda, A.S.; Katzman, D.K.; Kohn, M.R.; Le Grange, D.; Madden, S.; Whitelaw, M.; Redgrave, G.W. A systematic review of approaches to refeeding in patients with anorexia nervosa. Int. J. Eat. Disord. 2016, 49, 293–310. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K.; Judd, P.A.; Preedy, V.R.; Taylor, M.A. Enteral feeding: The effect on faecal output, the faecal microflora and SCFA concentrations. Proc. Nutr. Soc. 2004, 63, 105–113. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J. Tube feeding, the microbiota, and Clostridium difficile infection. World J. Gastroenterol. 2010, 16, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Whelan, K.; Judd, P.A.; Preedy, V.R.; Simmering, R.; Jann, A.; Taylor, M.A. Fructooligosaccharides and fiber partially prevent the alterations in fecal microbiota and short-chain fatty acid concentrations caused by standard enteral formula in healthy humans. J. Nutr. 2005, 135, 1896–1902. [Google Scholar] [PubMed]
- Raevuori, A.; Suokas, J.; Haukka, J.; Gissler, M.; Linna, M.; Grainger, M.; Suvisaari, J. Highly increased risk of type 2 diabetes in patients with binge eating disorder and bulimia nervosa. Int. J. Eat. Disord. 2015, 48, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Raevuori, A.; Haukka, J.; Vaarala, O.; Suvisaari, J.M.; Gissler, M.; Grainger, M.; Linna, M.S.; Suokas, J.T. The increased risk for autoimmune diseases in patients with eating disorders. PLoS ONE 2014, 9, e104845. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 535, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Correa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Morrison, D.J.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Rios-Covian, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de Los Reyes-Gavilan, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.Y.; Mitchell, A.J.; Holmes, A.J.; Denyer, G.S.; Gummesson, A.; Caterson, I.D.; Storlien, L.H. Role of the gut in visceral fat inflammation and metabolic disorders. Obesity 2011, 19, 2113–2120. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Delzenne, N.M. Interplay between obesity and associated metabolic disorders: New insights into the gut microbiota. Curr. Opin. Pharmacol. 2009, 9, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R.; Buschmann, M.M.; Romero-Calvo, I.; Sailer, A.; Shen, L. The role of molecular remodeling in differential regulation of tight junction permeability. Semin. Cell Dev. Biol. 2014, 36, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Jesus, P.; Ouelaa, W.; Francois, M.; Riachy, L.; Guerin, C.; Aziz, M.; Do Rego, J.C.; Dechelotte, P.; Fetissov, S.O.; Coeffier, M. Alteration of intestinal barrier function during activity-based anorexia in mice. Clin. Nutr. 2014, 33, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Monteleone, P.; Carratu, R.; Carteni, M.; Generoso, M.; Lamberti, M.; Magistris, L.D.; Brambilla, F.; Colurcio, B.; Secondulfo, M.; Maj, M. Intestinal permeability is decreased in anorexia nervosa. Mol. Psychiatry 2004, 9, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L. The gut microbiota and obesity: From correlation to causality. Nat. Rev. Microbiol. 2013, 11, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Quigley, E.M.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.; Moayyedi, P. Efficacy of prebiotics, probiotics, and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: Systematic review and meta-analysis. Am. J. Gastroenterol. 2014, 109, 1547–1561, quiz 1546, 1562. [Google Scholar] [CrossRef] [PubMed]
- Curro, D.; Ianiro, G.; Pecere, S.; Bibbo, S.; Cammarota, G. Probiotics, fibre and herbal medicinal products for functional and inflammatory bowel disorders. Br. J. Pharmacol. 2016, 174, 1426–1449. [Google Scholar] [CrossRef] [PubMed]
- Wierdsma, N.J.; van Bodegraven, A.A.; Uitdehaag, B.M.; Arjaans, W.; Savelkoul, P.H.; Kruizenga, H.M.; van Bokhorst-de van der Schueren, M.A. Fructo-oligosaccharides and fibre in enteral nutrition has a beneficial influence on microbiota and gastrointestinal quality of life. Scand. J. Gastroenterol. 2009, 44, 804–812. [Google Scholar] [CrossRef] [PubMed]
- Pirbaglou, M.; Katz, J.; de Souza, R.J.; Stearns, J.C.; Motamed, M.; Ritvo, P. Probiotic supplementation can positively affect anxiety and depressive symptoms: A systematic review of randomized controlled trials. Nutr. Res. 2016, 36, 889–898. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Wang, K.; Hu, J. Effect of Probiotics on Depression: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Nutrients 2016, 8, 483. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Categories of Eating Disorders | Diagnostic Criteria (DSM-5) 1 | Eating Disorder Behaviors | |||||
|---|---|---|---|---|---|---|---|
| Restrict | Binge | Vomit | Laxative | Over-Exercise | Body, Weight & Shape Concerns | ||
| Anorexia Nervosa (AN) | Significant weight loss; fear of weight gain; body weight & shape concerns | + | +/− | +/− | +/− | +/− | + |
| Bulimia Nervosa (BN) | Regular binge eating; compensation behaviors (e.g., vomiting, laxative abuse); body weight & shape concerns | +/− | + | +/− | +/− | +/− | + |
| Binge Eating Disorder (BED) | Regular binge eating, at least 3 of 5 other negative features (e.g., eating large amounts when not hungry, eating alone due to embarrassment); significant distress | +/− | + | +/− | |||
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lam, Y.Y.; Maguire, S.; Palacios, T.; Caterson, I.D. Are the Gut Bacteria Telling Us to Eat or Not to Eat? Reviewing the Role of Gut Microbiota in the Etiology, Disease Progression and Treatment of Eating Disorders. Nutrients 2017, 9, 602. https://doi.org/10.3390/nu9060602
Lam YY, Maguire S, Palacios T, Caterson ID. Are the Gut Bacteria Telling Us to Eat or Not to Eat? Reviewing the Role of Gut Microbiota in the Etiology, Disease Progression and Treatment of Eating Disorders. Nutrients. 2017; 9(6):602. https://doi.org/10.3390/nu9060602
Chicago/Turabian StyleLam, Yan Y., Sarah Maguire, Talia Palacios, and Ian D. Caterson. 2017. "Are the Gut Bacteria Telling Us to Eat or Not to Eat? Reviewing the Role of Gut Microbiota in the Etiology, Disease Progression and Treatment of Eating Disorders" Nutrients 9, no. 6: 602. https://doi.org/10.3390/nu9060602