Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases

Abstract
:1. Introduction

{kind=link}
| Term | Interpretation |
|---|---|
| Lactase Persistent; LP | The dominant genetic trait in adults with |
| continuous ability to digest lactose throughout adulthood | |
| Lactase Non Peristent; LNP | The natural decline in intestinal lactase to <10 u/g |
| of tissue which leaves adults with minimal ability to | |
| digest lactose | |
| Lactase Deficiency; LD | Reduction of intestinal lactase enzyme from |
| either genetic (LNP) or any secondary causes | |
| due to diseases of the proximal small bowel mucosa | |
| Lactose Maldigestion; LM | Inability to digest lactose for any cause primary |
| (LNP) or secondary causes resulting in undigested | |
| lactose reaching the colon | |
| Lactose Intolerance; LI | Symptoms resulting from the ingestion of lactose |
| including flatus, gas, bloating, cramps, diarrhea | |
| and rarely vomiting. Currently, symptoms must | |
| not be present when an inert placebo is exchanged | |
| for lactose | |
| Lactose Sensitivity; | Symptoms with or without symptoms of LI and |
| the systemic features depression, headache, fatigue |
2. Symptomatic Improvement with Regular Lactose Consumption in LNP People
| Author | No. | LM in | LM | LM | BH2 Auc | BH2* Auc | BH2* Auc |
|---|---|---|---|---|---|---|---|
| Included | Preg N | Prepart | Postpart | Pregnancy | Prepart | Postpart | |
| Villar [28] | 114 | 62/118 (54%) | 35/118 (29.7%) | 116 ± 9.6 ppm + | 54 ± 7.3 ppm | ||
| Szilagyi [29] + | 28 | 16/28 (57.1%) | 20/28 (71%) | 3816.9 ± 577.4 ppm | 6490 ± 925 ppm |
| Area under the curve for beath H2 | Symptom Score (Max = 42) | Delayed Symtom Score (no max) | Lactose Intestinal Transit time (min) | Oral cecal Transit time (min) | |
|---|---|---|---|---|---|
| Baseline (n = 16) | 10,243 ± 1607 | 18.7 ± 3 | 8 ± 1.3 | 43.1 ± 6.8 | 56.9 ± 5.9 |
| 8 mg loperimide (n = 14) | 8527 ± 1456.7 | 8.5 ± 2.1* | 5.5 ± 1.5† | 48.2 ± 7.4 | 82.1 ± 13.9* |
| 12 mg Loperimide (n = 16) | 7685 ± 935.6** | 10.3 ± 2.4* | 3.1 ± 1* | 63.1 ± 9.7* | 90.3 ± 11.1* |
3. Probiotics, Prebiotics and Health Benefits for the Host
| Effect | Reference |
|---|---|
| Increases Fecal bulk and Laxation | [67] |
| Reduction of Intestinal Transit time | [67] |
| Microbial Substrate leading to SCFA | [68] |
| Butyrate preferred colonic nutrient | [68] |
| Anti carcinogen and inflammatory SCFA | [69] |
| Immune Modulation | [70] |
| Selective Stimulation of Lactic acid bacteria | [71,72] |
| Lactic acids stimulate other butyrogenic producing bacteria, altering metabolome and metagenome | [72,73] |
| Facilitated Calcium and other electrolyte absorption | [74] |
| Gastrointestinal/Nongastrointestinal | Prebiotic | Outcome | Reference |
|---|---|---|---|
| Infectious Gastroenteritis | OFS | Amoebic Gastroenteritis | [75] |
| Antibiotic Associated Diarrhea | OFS/Inulin | No effect | [75] |
| Irritable Bowel Syndrome | GOS/FOS | Modest benefit | [76,77,78] |
| Inflammatory Bowel Disease | Mixed Prebiotics | Possible Maintenance in UC | [46,79] |
| Colorectal Cancer | lactulose | decreased poly pformation | [80,81,82] |
| Lactose Intolerance | DS/OS | Improved symptoms Bifidobacteria expand | [32,33,34] |
| Necrotizing Enterocolitis | FOS/GOS | noclinical benefit | [83] |
| Cirrhosis (hepatic encephalopathy) | lactose(LNP), lactulose | Improved coma grade | [85,86] |
| Constipation | Fiber | no specific prebiotics | [67] |
| Acute Upper Respiratory Infections | GOS/FOS CT | rate lower with diet | [75] |
| Obesity, Metabolic Syndrome | FOS/inulin | improved satiety | [87] |
| Diabetes Type 2 | FOS/Inulin | improved glucose/insulin | [87] |
| Pediatric Eczema | GOS/FOS | reduced eczema | [88,89] |
| Pediatric Atopic Dermatitis | OS/Pectin RCT | reduced vs. control | [89] |
| Urinary Infections | lactulose | modest reduction | [90] |
4. Microbial and Metabolomic Effects of Lactose
| In Vitro | Reference |
|---|---|
| Hydrogen production is reduced and is not due to low pH | [100] |
| Bifidobacteria or Lactobacilli efficiently metabolize lactose | [93,94] |
| Lactose decreases bacterial ammonia production | [95] |
| In a model colon lactose increases growth of Bifidobacteria | [101] |
| A prebiotic Index for lactose has been calculated as 5.75 | [102] |
| In Vivo (Animal) | |
| Lactose induces Lactobacilli in Colon (pigs) | [103] |
| Lactose enhances metabolome of microflora (rats) Human | [104] |
| Lactose improves hepatic encephalopathy in lactose maldigester cirrhotics | [84] |
| Lactose improves some features of lactose intolerance in lactose maldigesters | [10] |
| Measurable increase in fecal β- galactosidase after regular lactose or lactulose consumption | [10,42] |
| Lactose induced expansion of Lactobacilli and Bifidobacteria. Decreased Bacteroides and Clostridia species | [105] |
| Selective increase in fecal Bifidobacteria in lactose maldigesters but not lactose digesters after 2 weeks of lactose ingestion. | [106] |
5. Effects of Dairy Foods on Diseases

| Risk Decreased | No effect on Risk | Risk Increased |
|---|---|---|
| Colorectal Cancer [114] (19 cohort) | Ovarian [137] (cohort) | Prostate [130] (32 cohort) |
| TDF 0.83 (0.78–0.88) | Low fat DF 0.76 (0.54–1.06) | TDF 1.07 (1.02–1.12) |
| Milk 0.91 (0.85–0.94) | Lactose 0.87 (0.69–1.11) * | Milk 1.03 (1.00–1.07) |
| Cheese 0.96 (0.83–1.12) | N 764 women | N TDF; 38,107 of 848,395 participants Milk; 11,392 of 556,146 participants |
| NTDF; 11,579 cases vs. 1,170,942 controls Milk; 5011 cases vs. 655,483 controls | Origin United States | Origin United Kingdom |
| OriginUnited Kingdom | ||
| Stomach [123] (17 case-control, 6 cohort) | Bladder [125] ** (14 studies) | Testicular [142] (case-control) |
| TDF 0.76 (0.64–0.91) | Milk 0.89 (0.77–1.02) | Milk 1.37 (1.12–1.68) |
| Not observed in Asians | N 7966 cases of 324,241 participants | Galactose 2.01 (1.41–2.86) |
| N 3256 cases vs. 17,026 controls | Origin China | N 269 cases vs. 797 controls |
| OriginChina | Origin Germany | |
| Breast [145] (18 cohort) | ||
| TDF 0.85 (0.76–0.95) | ||
| N 24,187 cases vs. 163,471 controls | ||
| Origin China | ||
| Pancreas [124] (case-control) | ||
| Low fat milk 0.51 (0.3–0.84) | ||
| N 532 patients vs. 1701 controls | ||
| Origin United States | ||
| Crohn’s Disease [151] (case-control) | ||
| Milk 0.82 (0.69–0.97) | ||
| N 218 cases vs. 812 controls | ||
| Origin United Kingdom | ||
| Obesity [154] (nation-wide survey) | ||
| TDF 0.51 (0.3–0.54) | ||
| N 1352 cases | ||
| Origin Luxembourg | ||
| Dibetes Type II [158] (17 cohort) | ||
| TDF | ||
| N 22,877 cases vs. 526,487 controls | ||
| Origin United Kingdom |
6. Summary
Conflicts of Interest
References
- Swallow, D.M. Genetics of lactase persistence and lactose intolerance. Ann. Hum. Genet. 2003, 37, 197–219. [Google Scholar] [CrossRef] [PubMed]
- Tishkoff, S.A.; Reed, F.A.; Ranciaro, A.; Voight, B.F.; Babbitt, C.C.; Silverman, J.S. Convergent adaptation of human lactase persistence in Africa and Europe. Nat. Genet. 2007, 39, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Prentice, A.M. Dairy products in global public health. Am. J. Clin. Nutr. 2014, 99, 1212S–1216S. [Google Scholar] [CrossRef] [PubMed]
- Holden, C.; Mace, R. Phylogenetic analysis of the evolution of lactose digestion in adults. Hum. Biol. 1997, 69, 605–628. [Google Scholar] [CrossRef] [PubMed]
- Flatz, G.; Rotthauwe, H.W. Lactose nutrition and natural selection. Lancet 1973, 2, 76–77. [Google Scholar] [CrossRef]
- Bersaglieri, T.; Sabeti, P.C.; Patterson, N.; Vanderploeg, T.; Schaffner, S.F.; Drake, J.A. Genetic signatures of strong recent positive selection at the lactase gene. Am. J. Hum. Genet. 2004, 74, 1111–1120. [Google Scholar] [CrossRef] [PubMed]
- Gerbault, P.; Liebert, A.; Swallow, D.M.; Thomas, M.G. Lactose malabsorption and nutrition. In Advanced Nutrition and Dietetics in Gastroenterology, 1st ed.; Lomer, M., Ed.; John Wiley and Sons Ltd: Oxford, UK, 2014; pp. 202–209. [Google Scholar]
- Shrier, I.; Szilagyi, A.; Correa, J.A. Impact of lactose containing foods and the genetics of lactase on diseases: An analytical review of population data. Cancer Nutr. 2008, 60, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Scrimshaw, N.S.; Murray, E.B. Prevalence of lactose maldigestion. Am. J. Clin. Nutr. 1988, 48, 1086–1098. [Google Scholar]
- Hertzler, S.R.; Savaiano, D.A. Colonic adaptation to daily lactose feeding in lactose maldigesters reduces lactose intolerance. Am. J. Clin. Nutr. 1996, 64, 232–236. [Google Scholar] [PubMed]
- Suchy, F.J.; Brannon, P.M.; Carpenter, T.O.; Fernandez, J.R.; Gilsanz, V.; Gould, J.B.; Hall, K.; Hui, S.L.; Lupton, J.; Mennella, J.; et al. National Institutes of Health Consensus Development Conference: Lactose intolerance and health. Ann. Intern. Med. 2010, 152, 792–796. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Venema, K.; Priebe, M.G.; Welling, G.W.; Brummer, R.J.; Vonk, R.J. The colonic metabolism in lactose intolerance. Eur. J. Clin. Investig. 2008, 38, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Shaukat, A.; Levitt, M.D.; Taylor, B.C.; MacDonald, R.; Shamliyan, T.A.; Kane, R.L.; Wilt, T.J. Systematic review: Effective management strategies for lactose intolerance. Ann. Intern. Med. 2010, 152, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Eadala, P.; Waud, J.P.; Matthews, S.B.; Green, J.T.; Campbell, A.K. Quantifying the “hidden” lactose in drugs used for the treatment of gastrointestinal conditions. Aliment. Pharmacol. Ther. 2009, 29, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Montalto, M.; Gallo, A.; Santoro, L.; D’Onofrio, F.; Curigliano, V.; Covino, M.; Cammarota, G.; Grieco, A.; Gasbarrini, A.; Gasbarrini, G. Low-dose lactose in drugs neither increases breath hydrogen excretion nor causes gastrointestinal symptoms. Aliment. Pharmacol. Ther. 2008, 28, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Matthews, S.B.; Waud, J.P.; Roberts, A.G.; Campbell, A.K. Systemic lactose intolerance: A new perspective on an old problem. Postgrad. Med. J. 2005, 81, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Eadala, P.; Matthews, S.B.; Waud, J.P.; Green, J.T.; Campbell, A.K. Association of lactose sensitivity with inflammatory bowel disease—Demonstrated by analysis of genetic polymorphism, breath gases and symptoms. Aliment. Pharmacol. Ther. 2011, 34, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Gibson, P.R.; Shepherd, S.J. Evidence-based dietary management of functional gastrointestinal symptoms: The FODMAP approach. J. Gastroenterol. Hepatol. 2010, 25, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Casellas, F.; Aparici, A.; Casaus, M.; Rodríguez, P.; Malagelada, J.R. Subjective perception of lactose intolerance does not always indicate lactose malabsorption. Clin. Gastroenterol. Hepatol. 2010, 8, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.L.; Savaiano, D.A.; Levitt, M.D. A comparison after the consumption of milk or lactose-hydrolyzed milk by people with self-reported severe lactose intolerance. N. Engl. J. Med. 1995, 333, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Suarez, F.L.; Savaiano, D.; Arbisi, P.; Levitt, M.D. Tolerance to the daily ingestion of two cups of milk by individuals claiming lactose intolerance. Am. J. Clin. Nutr. 1997, 65, 1502–1506. [Google Scholar] [PubMed]
- Savaiano, D.A.; Boushey, C.J.; McCabe, G.P. Lactose intolerance symptoms assessed by meta-analysis: A grain of truth that leads to exaggeration. J. Nutr. 2006, 136, 1107–1113. [Google Scholar] [PubMed]
- Vernia, P.; di Camillo, M.; Foglietta, T.; Avallone, V.E.; de Carolis, A. Diagnosis of lactose intolerance and the “nocebo” effect: The role of negative expectations. Dig. Liver Dis. 2010, 42, 616–619. [Google Scholar] [CrossRef] [PubMed]
- Briet, F.; Pochart, P.; Marteau, P.; Flourie, B.; Arrigoni, E.; Rambaud, J.C. Improved clinical tolerance to chronic lactose ingestion in subjects with lactose intolerance: A placebo effect? Gut 1997, 41, 632–635. [Google Scholar] [CrossRef] [PubMed]
- Habte, D.; Sterby, G.; Jijalmassen, B. Lactose malabsorption in Ethiopian children. Acta Paediatr.Scand. 1973, 62, 649–654. [Google Scholar] [CrossRef] [PubMed]
- Sadre, M.; Karbasi, K. Lactose intolerance in Iran. Am. J. Clin. Nutr. 1979, 32, 1948–1954. [Google Scholar] [PubMed]
- Johnson, A.O.; Semenya, J.G.; Buchowski, M.S.; Enwonwu, C.O.; Scrimshaw, N.S. Adaptation of lactose maldigesters to continued milk intakes. Am. J. Clin. Nutr. 1993, 58, 879–881. [Google Scholar] [PubMed]
- Villar, J.; Kestler, E.; Castillo, P.; Juarez, A.; Menendez, R.; Solomons, N.W. Improved lactose digestion during pregnancy: A case of physiologic adaptation? Obstet. Gynecol. 1988, 71, 697–700. [Google Scholar] [PubMed]
- Szilagyi, A.; Salomon, R.; Martin, M.; Fokeef, K.; Seidman, E. Improved lactose handling in late phase pregnancy. Clin. Investig. Med. 1996, 74, 416–426. [Google Scholar]
- Paige, D.M.; Witter, F.R.; Bronner, Y.L.; Kessler, L.A.; Perman, J.A.; Paige, T.R. Lactose digestion in pregnant African-Americans. Public Health Nutr. 2003, 6, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Ladas, S.; Papanikos, J.; Arapakis, G. Lactose malabsorption in Greek adults: Correlation of small bowel transit time with the severity of lactose intolerance. Gut 1982, 23, 968–973. [Google Scholar] [CrossRef] [PubMed]
- Salomon, R.; Levy, E.; Levesque, D.; Szilagyi, A.; Seidman, E. Caco-2 cell dissaccharidase activities are unaffected by gestational hormones. Can. J. Physiol. Pharmacol. 1996, 74, 1126–1131. [Google Scholar] [CrossRef] [PubMed]
- Lazzara, F.; Szilagyi, A.; Lerman, A. Lactose intolerance in hyperthyroidism: A possible unmasking effect. Am. J. Gastroenterol. 1985, 80, 846. [Google Scholar]
- Szilagyi, A.; Lerman, S.; Barr, R.G.; Stern, J.; MacMullan, S. Influence of hypothyroidism on gastric emptying, oral cecal transit and lactose absorption. Clin. Investig. Med. 1992, 15, A44. [Google Scholar]
- Asik, M.; Gunes, F.; Binnetoglu, E.; Eroglu, M.; Bozkurt, N.; Sen, H.; Akbal, E.; Bakar, C.; Beyazit, Y.; Ukinc, K. Decrease in TSH levels after lactose restriction in Hashimoto’s thyroiditis patients with lactose intolerance. Endocrine 2014, 46, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Lebenthal, A.; Lebenthal, E. The ontogeny of the small intestinal epithelium. J. Parenter. Enteral Nutr. 1999, 23 (Suppl. 5), S3–S6. [Google Scholar] [CrossRef]
- Galluser, M.; Belkhou, R.; Freund, J.N.; Duluc, I.; Torp, N.; Danielsen, M.; Raul, F. Adaptation of intestinal hydrolases to starvation in rats: Effect of thyroid function. J. Comp. Physiol. B 1991, 161, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Lerman, S.; Barr, R.G.; Stern, J.; Colacone, A.; McMullan, S. Reversible lactose malabsorption and intolerance in Graves’ disease. Clin. Investig. Med. 1991, 14, 188–197. [Google Scholar]
- Yaylali, O.; Kirac, S.; Yilmaz, M.; Akin, F.; Yuksel, D.; Demirkan, N.; Akdag, B. Does hypothyroidism affect gastrointestinal motility? Gastroenterol. Res. Pract. 2009. ID529802. [Google Scholar] [CrossRef] [PubMed]
- Daher, R.; Yazbeck, T.; Jaoude, J.B.; Abboud, B. Consequences of dysthyroidism on the digestive tract and viscera. World J. Gastroenterol. 2009, 15, 2834–2838. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Salomon, R.; Seidman, E. Influence of Loperamide on lactose handling and oral caecal transit time. Aliment. Pharmacol. Ther. 1996, 10, 765–770. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Rivard, J.; Fokeeff, K. Improved parameters of lactose maldigestion using lactulose. Dig. Dis. Sci. 2001, 46, 1509–1519. [Google Scholar] [CrossRef] [PubMed]
- Savaiano, D.A.; Ritter, A.J.; Klaenhammer, T.B.; James, G.M.; Longcore, A.T.; Chandler, J.R.; Walker, W.A.; Foyt, H.L. Improving lactose digestion and symptoms of lactose intolerance with a novel galacto-oligosaccharide (RP-G28): A randomized, double-blind clinical trial. Nutr. J. 2013, 12. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi, A.; Cohen, A.; Vinokuroff, C.; Ahmad, D.; Nathwani, U.; Yesovitch, S. De-adapt and re-adapt with lactose but no cross-adapt to lactulose: A case of occult colonic bacterial adaptation. Can. J. Gastroenterol. 2004, 18, 677–680. [Google Scholar] [PubMed]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [PubMed]
- Morgan, X.C.; Huttenhower, C. Meta’omic analytic techniques for studying the intestinal microbiome. Gastoeneterology 2014, 146, 1437–1448. [Google Scholar] [CrossRef] [PubMed]
- Hollister, E.B.; Gao, X.; Versalovic, J. Compositional and functional features of gastrointestinal microbiome and their effects on human health. Gastroenterology 2014, 146, 1449–1458. [Google Scholar] [CrossRef] [PubMed]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the gut microbiota in disease. Microb. Ecol. Health Dis. 2015, 26, 26191. [Google Scholar] [CrossRef] [PubMed]
- Albenberg, L.G.; Wu, G.D. Diet and the intestinal microbiome: Associations, functions, and implications for health and disease. Gastroenterology 2014, 146, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.E.; le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, M.L.; Romanuk, T.N. A meta-analysis of probiotic efficacy for gastrointestinal diseases. PLoS ONE 2012, 7, e34938. [Google Scholar] [CrossRef] [PubMed]
- Videlock, E.J.; Cremonini, F. Meta-analysis: Probiotics in antibiotic-associated diarrhoea. Aliment. Pharmacol. Ther. 2012, 35, 1355–1369. [Google Scholar] [CrossRef] [PubMed]
- Tung, J.M.; Dolovich, L.R.; Lee, C.H. Prevention of Clostridium difficile infection with Saccharomyces boulardii: A systematic review. Can. J. Gastroenterol. 2009, 23, 817–821. [Google Scholar] [PubMed]
- Hoveyda, N.; Heneghan, C.; Mahtani, K.R.; Perera, R.; Roberts, N.; Glasziou, P. A systematic review and meta-analysis: Probiotics in the treatment of irritable bowel syndrome. BMC Gastroenterol. 2009, 9. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Zuo, Z.X.; Mao, A.P. Effect of probiotics on inducing remission and maintaining therapy in ulcerative colitis, Crohn’s disease, and pouchitis: Meta-analysis of randomized controlled trials. Inflamm. Bowel Dis. 2014, 20, 21–35. [Google Scholar] [CrossRef] [PubMed]
- Michail, S.K.; Stolfi, A.; Johnson, T.; Onady, G.M. Efficacy of probiotics in the treatment of pediatric atopic dermatitis: A meta-analysis of randomized controlled trials. Ann. Allergy AsthmaImmunol. 2008, 101, 508–516. [Google Scholar] [CrossRef]
- Lindsay, K.L.; Walsh, C.A.; Brennan, L.; McAuliffe, F.M. Probiotics in pregnancy and maternal outcomes: A systematic review. J. Matern. Fetal Neonatal Med. 2013, 26, 772–778. [Google Scholar] [CrossRef] [PubMed]
- Doege, K.; Grajecki, D.; Zyriax, B.C.; Detinkina, E.; Zu Eulenburg, C.; Buhling, K.J. Impact of maternal supplementation with probiotics during pregnancy on atopic eczema in childhood: A meta-analysis. Br. J. Nutr. 2012, 107, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Razmpoosh, E.; Javadi, M.; Ejtahed, H.S.; Mirmiran, P. Probiotics as beneficial agents on the management of diabetes mellitus: A systematic review. Diabetes Metab. Res. Rev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Almeida, C.C.; Lorena, S.L.; Pavan, C.R.; Akasaka, H.M.; Mesquita, M.A. Beneficial effects oflong-term consumption of a probiotic combination of Lactobacillus casei Shirota and Bifidobacteriumbreve Yakult may persist after suspension of therapy in lactose-intolerant patients. Nutr. Clin. Pract. 2012, 27, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Malolepszy, P.; Shrier, I.; Szilagyi, A. Adaptation to lactose intolerance is not achieved by long term ingestion of a multi species containing probiotic. Int. J. ProbioticsPrebiotics. 2006, 1, 113–120. [Google Scholar]
- Macfarlane, S.; Macfarlane, G.T.; Cummings, J.H. Review Article: Prebiotics in the gastrointestinal tract. Aliment. Pharmacol. Ther. 2006, 24, 701–714. [Google Scholar] [CrossRef] [PubMed]
- Ursell, L.K.; Haiser, H.J.; van Treuren, W.; Garg, N.; Reddivan, L.; Vanamala, J.; Dorrestein, P.C.; Turnbaugh, P.J.; Knight, R. The intestinal metabolome: An intersection between microbiota and host. Gastroenterology 2014, 146, 1470–1476. [Google Scholar] [CrossRef] [PubMed]
- McNulty, N.P.; Yatsunenko, T.; Hsiao, A.; Faith, J.J.; Muegge, B.D.; Goodman, A.L.; Henrissat, B.; Oozeer, R.; Cools-Portier, S.; Gobert, G.; et al. The impact of a consortium of fermented milk strains on the gut microbiome of gnotobiotic mice and monozygotic twins. Sci. Transl. Med. 2011, 3. [Google Scholar] [CrossRef] [PubMed]
- Young, S.L.; Simon, M.A.; Baird, M.A.; Tannock, G.W.; Bibiloni, R.; Spencely, K.; Lane, J.M.; Fitzharris, P.; Crane, J.; Town, I.; et al. Bifidobacterial species differentially affect expression of cell surface markers and cytokines of dendritic cells harvested from cord blood. Clin. Diagn. Lab. Immunol. 2004, 11, 686–690. [Google Scholar] [PubMed]
- De Filippo, C.; Cavalieri, D.; di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
- Slavin, J. Fiber and prebiotics: Mechanisms and health benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef] [PubMed]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, J.; Brummer, R.-J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Frei, R.; Akdis, M.; O’Mahony, P. Prebiotics, probiotics, synbiotics and the immune system: Experimental data and clinical evidence. Curr. Opin. Gastroenterol. 2015, 31, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Bouhnik, Y.; Attar, A.; Joly, F.A.; Riottot, M.; Dyard, F.; Flourié, B. Lactulose ingestion increases faecal bifidobacterial counts: A randomised double-blind study in healthy humans. Eur. J. Clin. Nutr. 2004, 58, 462–466. [Google Scholar] [CrossRef] [PubMed]
- Falony, G.; Lazidou, K.; Verschaeren, A.; Weckx, S.; Maes, D.; de Vuyst, L. In vitro analysis of fermentation of prebiotic inulin-type fructans by bifidobacterium species reveals four different phenotypes. Appl. Environ. Microbiol. 2009, 75, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Falony, G.; Calmeyn, T.; Leroy, F.; de Vuyst, L. Coculture fermentations of bifidobacterium species and bacteroides thetaiotaomicron reveal a mechanistic insight into the prebiotic effect of inulin-type fructans. Appl. Environ. Microbiol. 2009, 75, 2312–2319. [Google Scholar] [CrossRef] [PubMed]
- Duncan, S.H.; Louis, P.; Flint, H.J. Lactate—Utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 2004, 70, 5810–5817. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, R. Dairy products, yogurts, and bone health. Am. J. Clin. Nutr. 2014, 99, 1256S–1262S. [Google Scholar] [CrossRef] [PubMed]
- Weichert, S.; Schroten, H.; Adam, R. The role of prebiotics in prevention and treatment of childhood infectious diseases. Pediatr. Infect. Dis. J. 2012, 31, 859–862. [Google Scholar] [CrossRef] [PubMed]
- Ford, A.C.; Quigley, E.M.M.; Lacy, B.E.; Lembo, A.J.; Saito, Y.A.; Schiller, L.R.; Soffer, E.E.; Spiegel, B.M.R.; Moayyedi, P. Efficacy of prebiotics, probiotics and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: Systematic review and meta-analysis. Am. J. Gastroenterol. 2014, 109, 1547–1561. [Google Scholar] [CrossRef] [PubMed]
- Paineau, D.; Payen, F.; Panserieu, S.; Coulombier, G.; Sobaszek, A.; Lartigau, I.; Brabet, M.; Galmiche, J.P.; Tripodi, D.; Sacher-Huvelin, S.; et al. The effects of regular consumption of short-chainfructo-oligosaccharides on digestive comfort of subjects with minor functional bowel disorders. Br. J. Nutr. 2008, 99, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Silk, D.B.; Davis, A.; Vulevic, J.; Tzortzis, G.; Gibson, G.R. Clinical trial: The effects of a trans-galacto-oligosaccharide prebiotic on faecal microbiota and symptoms in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2009, 29, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Ghouri, Y.A.; Richards, D.M.; Rahimi, E.F.; Krill, J.T.; Jelinek, K.A.; DuPont, A.W. Systematic review of randomized controlled trials of probiotics, prebiotics and synbiotics in inflammatory bowel disease. Clin. Exp. Gastroenterol. 2014, 7, 473–487. [Google Scholar] [PubMed]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothan, C.; Patel, A.T.; Purama, R.K.; Dave, J.M.; Vyas, B.R.M. Potential of probiotics, prebiotics and synbiotics for management of colorectal cancer. Gut Microbes 2013, 4, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.J.; Robien, K.; Slavin, J.L. Effector prebiotics on biomarkers of colorectal cancer in humans: A systematic review. Nutr. Rev. 2012, 70, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Roncucci, L.; di Donato, P.; Carati, L.; Ferrari, A.; Perini, M.; Bertoni, G.; Bedogni, G.; Paris, B.; Svanoni, F.; Girola, M.; et al. Antioxidant vitamins or lactulose for the prevention of the recurrence of colorectal adenomas. Dis. Colon Rectum 1993, 36, 227–234. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.M.; Denning, P.W. Therapeutic use of prebiotics, probiotics and postbiotics to prevent necrotizing enterocolitis: What is the current evidence? Clin. Perinatol. 2013, 40, 11–25. [Google Scholar] [CrossRef] [PubMed]
- Uribe, M.; Márquez, M.A.; Garcí-Ramos, G.; Escobedo, V.; Murillo, H.; Guevara, L.; Lisker, R. Treatment of chronic portal-systemic encephalopathy with lactose in lactase-deficient patients. Dig. Dis. Sci. 1980, 25, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Uribe, M.; Toledo, H.; Perez, F.; Vargas, F.; Gil, S.; Garcia-Ramos, G.; Ravelli, G.P.; Guevara, L. Lactitol, a second-generation disaccharide for treatment of chronic portal-systemic encephalopathy. A double-blind, crossover, randomized clinical trial. Dig. Dis. Sci. 1987, 32, 1345–1353. [Google Scholar] [CrossRef] [PubMed]
- Wen, J.; Liu, Q.; Song, J.; Tong, M.; Peng, T.; Liang, H. Lactulose is highly potential in prophylaxis of hepatic encephalopathy in patients with cirrhosis and upper gastrointestinal bleeding: Results of a controlled randomized trial. Digestion 2013, 87, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Kellow, N.J.; Coughlan, M.T.; Reid, C.M. Metabolic benefits of dietary prebiotics in human subjects: A systematic review of randomized controlled trials. Br. J. Nutr. 2014, 111, 1147–1161. [Google Scholar] [CrossRef] [PubMed]
- Osborn, D.A.; Sinn, J.K. Prebiotics in infants for prevention of allergy. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- Baquerizo Nole, K.L.; Yim, E.; Keri, J.E. Probiotics and prebiotics in dermatology. J. Am. Acad. Dermatol. 2014, 71, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Battle, M.; Martin, T.; Fulton, J. Lactulose may help prevent urinary tract infections. Br. Med. J. 2001, 323, 930–937. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.M.; Vanhoutvin, S.A.; Troost, F.J.; Rijkers, G.; de Bruïne, A.; Bast, A.; Venema, K.; Brummer, R.J. Effect of butyrate enemas on inflammation and antioxidant status in the colonic mucosa of patients with ulcerative colitis in remission. Clin. Nutr. 2010, 29, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Guillemot, F.; Colombel, J.F.; Neut, C.; Verplanck, N.; Lecomte, M.; Romond, C.; Paris, J.C.; Cortot, A. Treatment of diversion colitis by short-chain fatty acids. Prospective and double-blind study. Dis. Colon Rectum 1991, 34, 861–864. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Savaiano, D.A. In vitro lactose fermentation by human colonic bacteria is modified by lactobacillus acidophilus supplementation. J. Nutr. 1997, 1237, 1489–1495. [Google Scholar]
- Jiang, T.; Savaiano, D.A. Modification of colonic fermentation by bifidobacteria and pH in vitro (Impact on lactose metabolism, short-chain fatty acid, and lactate production). Dig. Dis. Sci. 1997, 42, 2370–2377. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Esquivel, M.; Maran, S.; Poo, J.L.; Munoz, R.M. In vitro and in vivo lactose and lactulose effects on colonic fermentation and portal-systemic encephalopathy parameters. Scand. J. Gastroenterol. 1997, 32 (Suppl. 222), 49–52. [Google Scholar]
- Szilagyi, A. Redefining lactose as a conditional prebiotic. Can. J. Gastroenterol. 2004, 18, 163–167. [Google Scholar] [PubMed]
- Gilat, T.; Russo, S.; Gelman-Malachi, E.; Aldor, T.A. Lactase in man: A nonadaptable enzyme. Gastroenterology 1972, 62, 1125–1127. [Google Scholar] [PubMed]
- Keusch, G.T.; Troncale, F.J.; Thavaramara, B.; Prinyanont, P.; Anderson, P.R.; Bhamarapravathi, N. Lactase deficiency in Thailand : Effects of prolonged lactose feeding. Am. J. Clin. Nutr. 1969, 22, 638–641. [Google Scholar] [PubMed]
- Bond, J.H.; Levitt, M.D. Quantitative measurement of lactose absorption. Gastroenterology 1976, 70, 1058–1062. [Google Scholar] [PubMed]
- Hertzler, S.R.; Savaiano, D.A.; Levitt, M.D. Fecal hydrogen production and consumption measurements. Response to daily lactose ingestion by lactose maldigesters. Dig. Dis. Sci. 1997, 42, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Makivuokko, H.A.; Saarinen, M.T.; Ouwehand, A.C.; Rautonen, N.E. Effects of lactose on colon microbial community structure and function in a four-stage semi-continuous culture system. Biosci. Biotechnol. Biochem. 2006, 70, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.L.; Gibson, G.R.; Rastall, R.A. Influence of disaccharide structure on prebiotic selectivity in vitro. J. Agric. Food Chem. 2005, 53, 5192–5199. [Google Scholar] [CrossRef] [PubMed]
- Daly, K.; Darby, A.C.; Hall, N.; Nau, A.; Bravo, D.; Shirazi-Beechey, S.P. Dietary supplementation with lactose or artificial sweetener enhances swine gut lactobacillus population abundance. Br. J. Nutr. 2014, 111, S30–S35. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, V.; Even, P.C.; Larue-Achagiotis, C.; Blouin, J.M.; Blachier, F.; Benamouzig, R.; Tome, D.; Davila, A.-M. Lactose absorption and colonic fermentation after host metabolism in rats. Br. J. Nutr. 2013, 110, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Kimura, M. Influence of lactose on faecal microflora in lactose maldigesters. Microb. Ecol. Health Dis. 1993, 6, 73–76. [Google Scholar] [CrossRef]
- Szilagyi, A.; Shrier, I.; Heilpern, D.; Je, J.S.; Park, S.H.; Chong, G.; Lalonde, C.; Cote, L.-F.; Lee, B.H. Differential impact of lactose/lactase phenotype on colonic microflora. Can. J. Gastroenterol. 2010, 24, 373–379. [Google Scholar] [PubMed]
- Palframan, R.; Gibson, G.R.; Rastall, R.A. Development of a quantitative tool for the comparison of the prebiotic effect of dietary oligosaccharides. Lett. Appl. Microbiol. 2003, 37, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Oku, T.; Nakamura, S.; Ichinose, M. Maximum permissive dose of lactose and lactitol for transitory diarrhea, and utilizable capacity for lactose in Japanese female adults. J. Nutr. Sci. Viaminol. 2005, 51, 51–57. [Google Scholar] [CrossRef]
- Hertzler, S.R.; Huynh, B.C.; Savaiano, D.A. How much lactose is low lactose? J. Am. Diet. Assoc. 1996, 96, 243–246. [Google Scholar] [CrossRef]
- Bouhnik, Y.; Vahedi, K.; Achour, L.; Attar, A.; Salfati, J.; Pochart, P.; Marteau, P.; Flourié, B.; Bornet, F.; Rambaud, J.C. Short-chain fructo-oligosaccharide administration dose-dependently increases fecal bifidobacteria in healthy humans. J. Nutr. 1999, 129, 113–116. [Google Scholar] [PubMed]
- Pereira, P.C. Milk nutritional composition and its role in human health. Nutrition 2014, 30, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Leitzmann, M.F.; Subar, A.F.; Hollenbeck, A.; Schatzkin, A. Dairy Food, Calcium, and Risk of Cancer in the NIH-AARP Diet and Health Study. Arch. Intern. Med. 2009, 169, 391–401. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.; Smith-Warner, S.A.; Spiegelman, D.; Beeson, W.L.; van den Brandt, P.A.; Colditz, G.A.; Folsom, A.R.; Fraser, G.E.; Freudenheim, J.L.; Giovannucci, E.; et al. Dairy foods, calcium, and colorectal cancer: A pooled analysis of 10 cohort studies. J. Natl. Cancer Inst. 2004, 96, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Lau, R.; Chan, D.S.; Vieira, R.; Greenwood, D.C.; Kampman, E.; Norat, T. Dairy products and colorectal cancer risk: A systematic review and meta-analysis of cohort studies. Ann. Oncol. 2012, 23, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Weingarten, M.A.; Zalmanovici Trestioreanu, A.; Yaphe, J. Dietary calcium supplementation for preventing colorectal cancer and adenomatous polyps. Cochrane Database Syst. Rev. 2008, 23. [Google Scholar] [CrossRef]
- Chandler, P.D.; Buring, J.E.; Manson, J.E.; Giovannucci, E.L.; Moorthy, M.V.; Zhang, S.; Lee, I.M.; Lin, J.H. Circulating Vitamin D Levels and Risk of Colorectal Cancer in Women. Cancer Prev. Res. 2015. [Google Scholar] [CrossRef] [PubMed]
- Bostick, R.M. Effects of supplemental vitamin D and calcium on normal colon tissue and circulating biomarkers of risk for colorectal neoplasms. J. Steroid Biochem. Mol. Biol. 2015, 148, 86–95. [Google Scholar] [CrossRef] [PubMed]
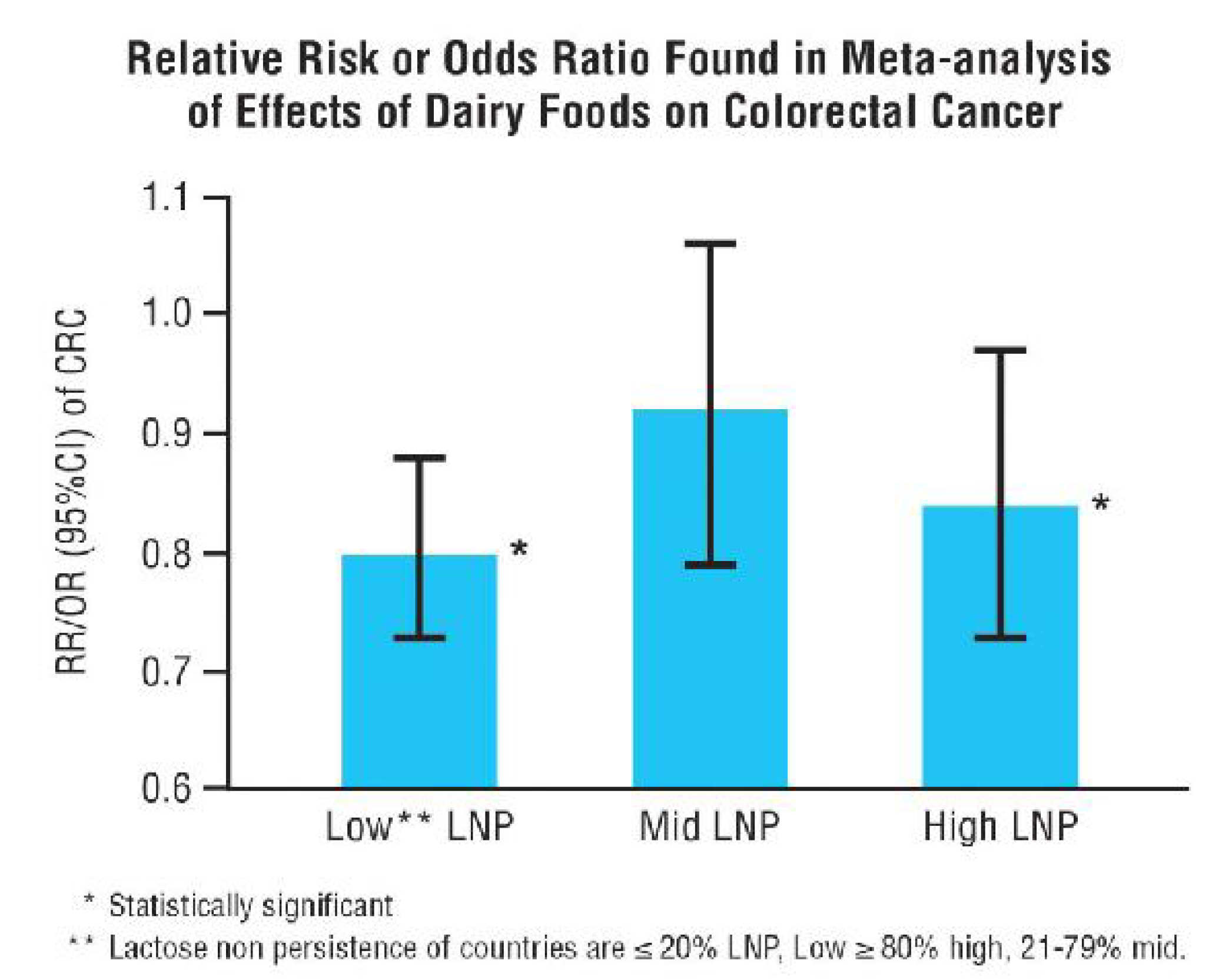
- Szilagyi, A.; Nathwani, U.; Vinukoroff, C.; Correa, A.; Shrier, I. The effect of lactose maldigestion on the relationship between dairy food intake and colorectal cancer: A systematic review. Nutr. Cancer 2006, 55, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Tian, S.B.; Yu, J.C.; Kang, W.M.; Ma, Z.Q.; Ye, X.; Cao, Z.J. Association between dairy intake and gastric cancer: A meta-analysis of observational studies. PLoS ONE 2014, 9, e101728. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Wu, H.Y.; Chuang, Y.C.; Chang, A.S.; Chao, H.H.; Chen, K.Y.; Chen, H.K.; Lai, G.M.; Huang, H.H.; Chen, C.J. Epidemiologic characteristics and multiple risk factors of stomach cancer in Taiwan. Anticancer Res. 1990, 10, 875–882. [Google Scholar] [PubMed]
- Pham, T.M.; Fujino, Y.; Kikuchi, S.; Tamakoshi, A.; Matsuda, S.; Yoshimura, T. Dietary patterns and risk of stomach cancer mortality: The Japan collaborative cohort study. Ann. Epidemiol. 2010, 20, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.X.; Qin, L.Q.; Wang, P.Y. Meta-analysis of the relationship between dairy product consumption and gastric cancer. Zhonghua Yu Fang Yi Xue Za Zhi 2009, 43, 193–196. [Google Scholar] [PubMed]
- Guo, Y.; Shan, Z.; Ren, H.; Chen, W. Dairy consumption and gastric cancer risk: A meta-analysisof epidemiological studies. Nutr. Cancer 2015, 67, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Gong, Z.; Holly, E.A.; Bracci, P.M. Dietary patterns and risk of pancreatic cancer in a large population-based case-control study in the San Francisco Bay Area. Nutr. Cancer 2013, 65, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; An, S.L.; Zhou, Y.; Liang, Z.K.; Jiao, Z.J.; Jing, Y.M.; Wan, P.; Shi, X.J.; Tan, W.L. Milk and dairy consumption and risk of bladder cancer: A meta-analysis. Urology 2011, 78, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.Q.; Dai, Y.; Lin, Y.W.; Qin, J.; Xie, L.P.; Zheng, X.Y. Milk consumption and bladder cancer risk: A meta-analysis of published epidemiological studies. Nutr. Cancer 2011, 63, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Keszei, A.P.; Schouten, L.J.; Goldbohm, R.A.; van den Brandt, P.A. Dairy intake and the risk of bladder cancer in the Netherlands Cohort Study on Diet and Cancer. Am. J. Epidemiol. 2010, 171, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Feyisetan, O.; Tracey, C.; Hellawell, G.O. Probiotics, dendritic cells and bladder cancer. BJU Int. 2012, 109, 1594–1597. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.C.; Orsini, N.; Wolk, A. Milk, milk products and lactose intake and ovarian cancer risk: A meta-analysis of epidemiological studies. Int. J. Cancer 2006, 118, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Navarro Rosenblatt, D.A.; Chan, D.S.; Vieira, A.R.; Vieira, R.; Greenwood, D.C.; Vatten, L.J.; Norat, T. Dairy products, calcium, and prostate cancer risk: A systematic review and meta-analysis of cohort studies. Am. J. Clin. Nutr. 2015, 101, 87–117. [Google Scholar] [CrossRef] [PubMed]
- Travis, R.C.; Appleby, P.N.; Siddiq, A.; Allen, N.E.; Kaaks, R.; Canzian, F.; Feller, S.; Tjønneland, A.; Føns Johnsen, N.; Overvad, K.; et al. Genetic variation in the lactase gene, dairy product intake and risk for prostate cancer in the European prospective investigation into cancer and nutrition. Int. J. Cancer 2013, 132, 1901–1910. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.J.; Birkett, N.J.; Johnson, K.C.; Shatenstein, B.; Ghadirian, P.; Krewski, D. Canadian Cancer Registries Epidemiology Research Group. Dietary risk factors for testicular carcinoma. Int. J. Cancer 2003, 106, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Cramer, D.W. Lactase persistence and milk consumption as determinants of ovarian cancer risk. Am. J. Epidemiol. 1989, 130, 904–910. [Google Scholar] [PubMed]
- Cramer, D.W.; Harlow, B.L.; Willett, W.C.; Welch, W.R.; Bell, D.A.; Scully, R.E.; Ng, W.G.; Knapp, R.C. Galactose consumption and metabolism in relation to the risk of ovarian cancer. Lancet 1989, 2, 66–71. [Google Scholar] [CrossRef]
- Faber, M.T.; Jensen, A.; Søgaard, M.; Høgdall, E.; Høgdall, C.; Blaakaer, J.; Kjaer, S.K. Use of dairy products, lactose, and calcium and risk of ovarian cancer—Results from a Danish case-controlstudy. Acta Oncol. 2012, 51, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Genkinger, J.M.; Hunter, D.J.; Spiegelman, D.; Anderson, K.E.; Arslan, A.; Beeson, W.L.; Buring, J.E.; Fraser, G.E.; Freudenheim, J.L.; Goldbohm, R.A.; et al. Dairy products and ovarian cancer: A pooled analysis of 12 cohort studies. Cancer Epidemiol. Biomarker. Prev. 2006, 15, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Merritt, M.A.; Poole, E.M.; Hankinson, S.E.; Willett, W.C.; Tworoger, S.S. Dairy food and nutrient intake in different life periods in relation to risk of ovarian cancer. Cancer Causes Control 2014, 25, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Merritt, M.A.; Cramer, D.W.; Vitonis, A.F.; Titus, L.J.; Terry, K.L. Dairy foods and nutrients in relation to risk of ovarian cancer and major histological subtypes. Int. J. Cancer 2013, 132, 1114–1124. [Google Scholar] [CrossRef] [PubMed]
- Skowronski, R.J.; Peehl, P.Y.; Feldman, D. Vitamin D and prostate cancer: 1,25 dihydroxyvitamin D3 receptors and actions in human prostate cancer cell lines. Endocrinology 1993, 132, 1952–1960. [Google Scholar] [PubMed]
- Agarwal, M.M.; Rana, S.V.; Mandal, A.K.; Malhotra, S.; Khandelwal, N.; Kumar, S.; Acharya, N.C.; Singh, S.K. Lactose intolerance in prostate cancer patients: Incidence and associated factors. Scand. J. Gastroeneterol. 2008, 43, 270–276. [Google Scholar] [CrossRef]
- Davies, T.W.; Palmer, C.R.; Lipscombe, J.M. Adolescent milk, dairy product and fruit consumption and testicular cancer. Br. J. Cancer 1998, 74, 657–660. [Google Scholar] [CrossRef]
- Stang, A.; Ahrens, W.; Baumgrdt-Elms, C. Adolescent milk fat and galactose consumption and testicular cell cancer. Cancer Epidemiol. Biomark. Prev. 2006, 15, 2189–2195. [Google Scholar] [CrossRef] [PubMed]
- Ganmaa, D.; Sato, A. The possible role of female sex hormones in milk from pregnant cows in the development of breast, ovarian and corpus uteri cancers. Med. Hypotheses 2005, 65, 1028–1037. [Google Scholar] [CrossRef] [PubMed]
- Pala, V.; Krogh, V.; Berrino, F.; Sieri, S.; Grioni, S.; Tjønneland, A.; Olsen, A.; Jakobsen, M.U.; Overvad, K.; Clavel-Chapelon, F.; et al. Meat, eggs, dairy products, and risk of breast cancer in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Am. J. Clin. Nutr. 2009, 90, 602–612. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Y.; Zhang, L.; He, K.; Qin, L.Q. Dairy consumption and risk of breast cancer: A meta-analysis of prospective cohort studies. Breast Cancer Res. Treat. 2011, 127, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Sankaranarayanan, R.; Varghese, C.; Duffy, S.W.; Padmakumary, G.; Day, N.E.; Nair, M.K. A case-control study of diet and lung cancer in Kerala, south India. Int. J. Cancer 1994, 58, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Ji, J.; Sundquist, J.; Sundquist, K. Lactose intolerance and risk of lung, breast and ovarian cancers: Aetiological clues from a population-based study in Sweden. Br. J. Cancer 2015, 112, 149–152. [Google Scholar] [CrossRef] [PubMed]
- Asakura, H.; Suzuki, K.; Kitahora, T.; Morizane, T. Is there a link between food and intestinal microbes and the occurrence of Crohn’s disease an ulcerative colitis. J. Gastroenterol. Hepatol. 2008, 23, 1794–1801. [Google Scholar] [CrossRef] [PubMed]
- Joachim, G. The relationship between habits of food consumption and reported reactions to food in people with inflammatory bowel disease—Testing the limits. Nutr. Health 1999, 13, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Jantchou, P.; Morois, S.; Clavel-Chapelon, F.; Boutron-Ruault, M.C.; Carbonnel, F. Animal protein intake and risk of inflammatory bowel disease: The E3N prospective study. Am. J. Gastroenterol. 2010, 105, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Abubakar, I.; Myhill, D.J.; Hart, A.R.; Lake, I.R.; Harvey, I.; Rhodes, J.M.; Robinson, R.; Lobo, A.J.; Probert, C.S.; Hunter, P.R. A case-control study of drinking water and dairy products in Crohn’s disease—Further investigation of the possible role of Mycobacterium avium paratuberculosis. Am. J. Epidemiol. 2007, 165, 776–783. [Google Scholar] [CrossRef] [PubMed]
- Mishkin, B.; Yalovsky, M.; Mishkin, S. Increased prevalence of lactose malabsorption in Crohn’s disease patients at low risk for lactose malabsorption based on ethnic origin. Am. J. Gastroenterol. 1997, 92, 1148–1153. [Google Scholar] [PubMed]
- Szilagyi, A.; Rivard, J.; Shrier, J. Diminished efficacy of colonic adaptation to lactulose occurs in patients with inflammatory bowel disease in remission. Dig. Dis. Sci. 2002, 47, 2811–2822. [Google Scholar] [CrossRef] [PubMed]
- Crichton, G.E.; Alkerwi, A. Whole-fat dairy food intake is inversely associated with obesity prevalence: Findings from the Observation of Cardiovascular Risk Factors in Luxembourg study. Nutr. Res. 2014, 34, 936–943. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, T.A.; Bremner, A.P.; Bremer, H.K.; Seares, M.E.; Beilin, L.J.; Mori, T.A.; Lyons-Wall, P.; Devine, A.; Oddy, W.H. Dairy product consumption, dietary nutrient and energy density and associations with obesity in Australian adolescents. J. Hum. Nutr. Diet. 2014. [Google Scholar] [CrossRef] [PubMed]
- Hirahatake, K.M.; Slavin, J.L.; Maki, K.C.; Adams, S.H. Associations between dairy foods, diabetes, and metabolic health: Potential mechanisms and future directions. Metabolism 2014, 63, 618–627. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, I.M.; Li-Chan, E.C. Investigation of the putative associations between dairy consumption and incidence of type 1 and type 2 diabetes. Crit. Rev. Food Sci. Nutr. 2014, 54, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Aune, D.; Norat, T.; Romundstad, P.; Vatten, L.J. Dairy products and the risk of type 2 diabetes: A systematic review and dose-response meta-analysis of cohort studies. Am. J. Clin. Nutr. 2013, 98, 1066–1083. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szilagyi, A. Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases. Nutrients 2015, 7, 6751-6779. https://doi.org/10.3390/nu7085309
Szilagyi A. Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases. Nutrients. 2015; 7(8):6751-6779. https://doi.org/10.3390/nu7085309
Chicago/Turabian StyleSzilagyi, Andrew. 2015. "Adaptation to Lactose in Lactase Non Persistent People: Effects on Intolerance and the Relationship between Dairy Food Consumption and Evalution of Diseases" Nutrients 7, no. 8: 6751-6779. https://doi.org/10.3390/nu7085309