Low Folate and Selenium in the Mouse Maternal Diet Alters Liver Gene Expression Patterns in the Offspring after Weaning

 ,
, 
Abstract
:1. Introduction
2. Experimental Section
2.1. Animal Experiments
2.2. Sample Collection

{kind=link}
{kind=link}
| Macronutrient | Diet “L” (0.4 mg Folate + 0.08 mg Se/kg) | Diet “H” (2.4 mg Folate+ 0.44 mg Se/kg) | |||
|---|---|---|---|---|---|
| gm % | kcal% | gm % | kcal% | ||
| Protein | 20 | 17 | 20 | 17 | |
| Carbohydrate | 49 | 42 | 50 | 43 | |
| Fat | 21 | 40 | 21 | 40 | |
| Total | 99 | 100 | |||
| kcal/g | 4.68 | 4.68 | |||
| Ingredient | gm | kcal | gm | kcal | |
| Casein | 195 | 780 | 195 | 780 | |
| dl-methionine | 3 | 12 | 3 | 12 | |
| Corn Starch | 50 | 200 | 50 | 200 | |
| Matodextrin 10 | 100 | 400 | 100 | 400 | |
| Sucrose | 341 | 1364 | 341 | 1364 | |
| Cellulose BW200 | 50 | 0 | 50 | 0 | |
| Milk Fat Anhydrous | 200 | 1800 | 200 | 1800 | |
| Corn Oil | 10 | 90 | 10 | 90 | |
| Mineral Mix S10001 † | 0 | 35 | |||
| Sodium Selenite (45.7% Se) | 0.000067 | 0.00058 | |||
| Mineral mix S19101 (no Se) | 35 | 0 | |||
| Calcium Carbonate | 4 | 4 | |||
| Vitamin Mix V10001 ‡ | 0 | 10 | 40 | ||
| Vitamin mix V14901 (no Fo) | 10 | 40 | 0 | ||
| Folic Acid | 0.00032 | 0.00032 | |||
| Choline Bitartrate | 2 | 2 | |||
| Cholesterol USP | 1.5 | 1.5 | |||
| Ethoxyquin | 0.04 | 0.04 | |||
| FD&C Yellow Dye #5 | 0.05 | 0 | |||
| FD&C Red Dye #40 | 0 | 0.05 | |||
| Total | 1001.59 | 4686 | 1001.59 | 4686 | |
2.3. RNA and DNA Isolation
2.4. Microarray Hybridisation and Analysis
3. Results
3.1. Food Intake and Body Weight
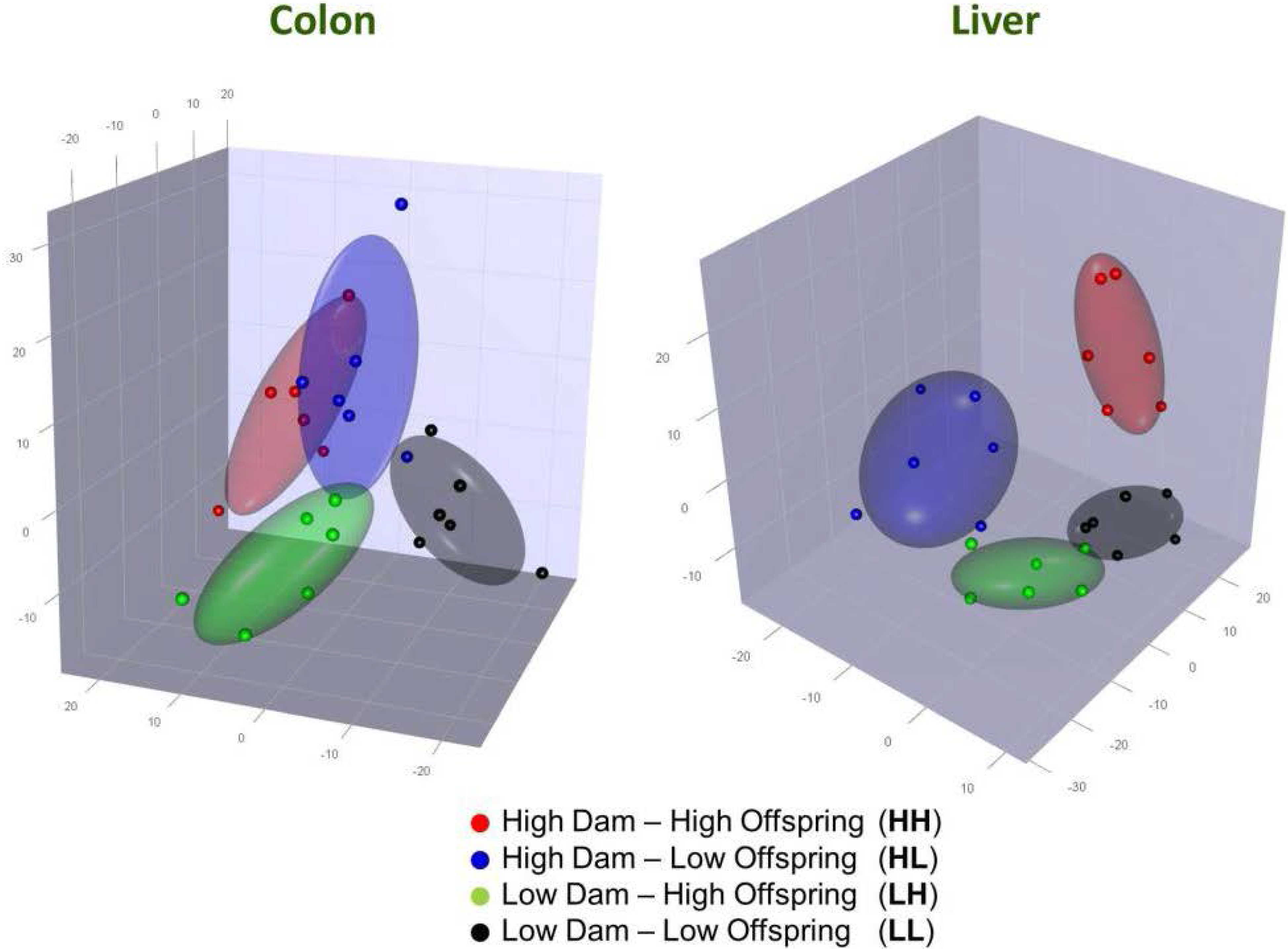
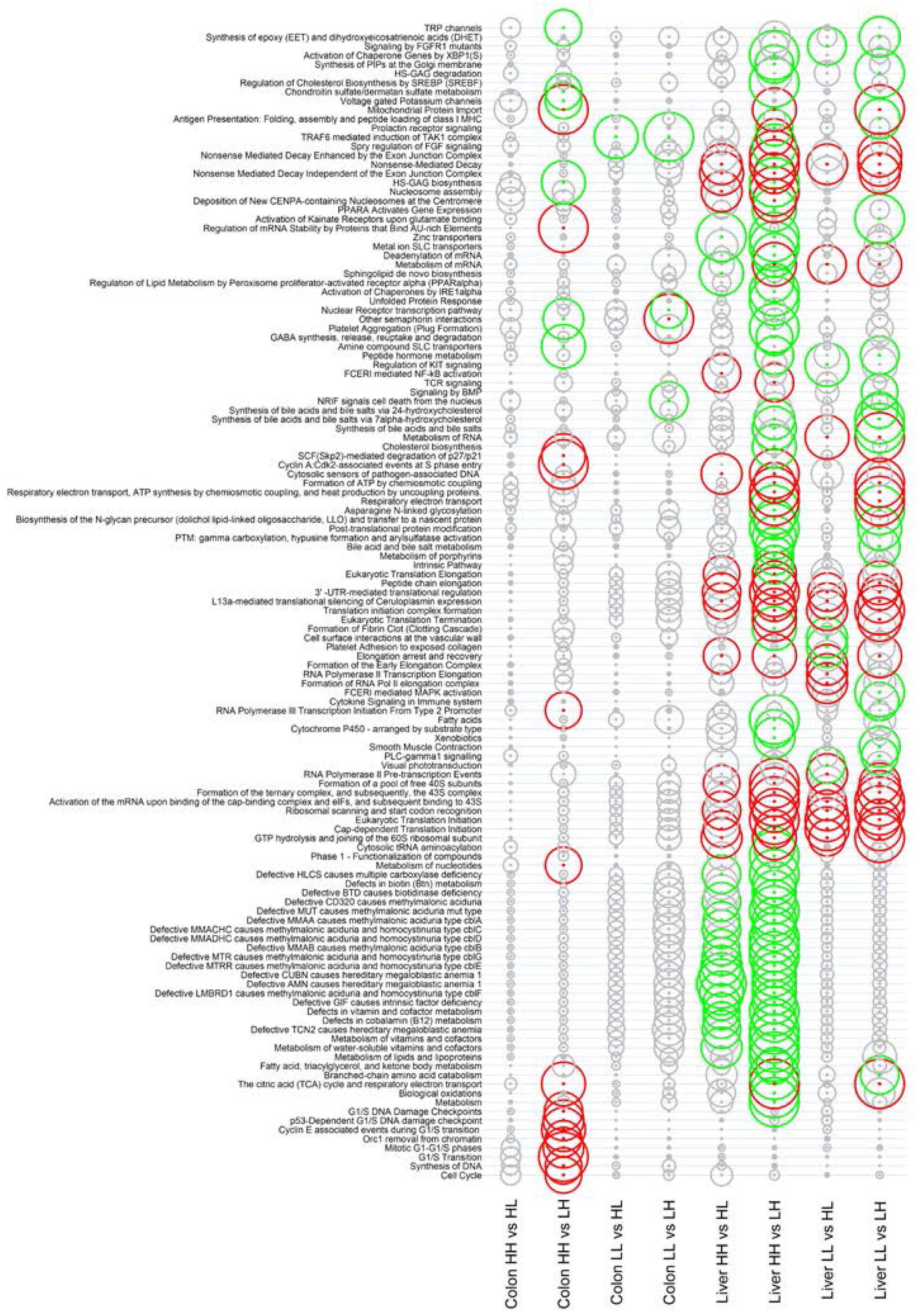
3.2. Gene Expression in the Liver

| Change in Gene Expression | Comparison | ||||
|---|---|---|---|---|---|
| HH vs. HL* | LH vs. LL | HH vs. LL | HL vs. LL | HH vs. LH* | |
| Lower expression | 50 | 18 | 13 | 0 | 358 |
| No change | 43,269 | 43,356 | 43,303 | 43,379 | 42,659 |
| Higher expression | 60 | 5 | 63 | 0 | 362 |
| Total DE genes | 110 | 23 | 76 | 0 | 720 |
3.3. Gene Expression in the Colon
4. Discussion
5. Conclusions

6. Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McKay, J.A.; Xie, L.; Manus, C.; Langie, S.A.; Maxwell, R.J.; Ford, D.; Mathers, J.C. Metabolic effects of a high-fat diet post-weaning after low maternal dietary folate during pregnancy and lactation. Mol. Nutr. Food Res. 2014, 58, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Poston, L.; Igosheva, N.; Mistry, H.D.; Seed, P.T.; Shennan, A.H.; Rana, S.; Karumanchi, S.A.; Chappell, L.C. Role of oxidative stress and antioxidant supplementation in pregnancy disorders. Am. J. Clin. Nutr. 2011, 94, 1980S–1985S. [Google Scholar] [CrossRef] [PubMed]
- Parisi, F.; Laoreti, A.; Cetin, I. Multiple micronutrient needs in pregnancy in industrialized countries. Ann. Nutr. Metab. 2014, 65, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Sloboda, D.M.; Howie, G.J.; Pleasants, A.; Gluckman, P.D.; Vickers, M.H. Pre- and postnatal nutritional histories influence reproductive maturation and ovarian function in the rat. PLoS ONE 2009, 4, e6744. [Google Scholar] [CrossRef] [PubMed]
- Cordain, L.; Eaton, S.B.; Sebastian, A.; Mann, N.; Lindeberg, S.; Watkins, B.A.; O’Keefe, J.H.; Brand-Miller, J. Origins and evolution of the Western diet: Health implications for the 21st century. Am. J. Clin. Nutr. 2005, 81, 341–354. [Google Scholar] [PubMed]
- Ernst, B.; Thurnheer, M.; Schmid, S.M.; Schultes, B. Evidence for the necessity to systematically assess micronutrient status prior to bariatric surgery. Obes. Surg. 2009, 19, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Kimmons, J.E.; Blanck, H.M.; Tohill, B.C.; Zhang, J.; Khan, L.K. Associations between body mass index and the prevalence of low micronutrient levels among US adults. MedGenMed. 2006, 8, 59. [Google Scholar] [PubMed]
- Madan, A.K.; Orth, W.S.; Tichansky, D.S.; Ternovits, C.A. Vitamin and trace mineral levels after laparoscopic gastric bypass. Obes. Surg. 2006, 16, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Damms-Machado, A.; Weser, G.; Bischoff, S.C. Micronutrient deficiency in obese subjects undergoing low calorie diet. Nutr. J. 2012, 11, 34. [Google Scholar] [CrossRef] [PubMed]
- Kaidar-Person, O.; Person, B.; Szomstein, S.; Rosenthal, R.J. Nutritional deficiencies in morbidly obese patients: A new form of malnutrition? Part B: Minerals. Obes. Surg. 2008, 18, 1028–1034. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- Hughes, D.J.; Fedirko, V.; Jenab, M.; Schomburg, L.; Meplan, C.; Freisling, H.; Bueno-de-Mesquita, H.B.; Hybsier, S.; Becker, N.P.; Czuban, M.; et al. Selenium status is associated with colorectal cancer risk in the European prospective investigation of cancer and nutrition cohort. Int. J. Cancer 2015, 136, 1149–1161. [Google Scholar] [CrossRef] [PubMed]
- Connelly-Frost, A.; Poole, C.; Satia, J.A.; Kupper, L.L.; Millikan, R.C.; Sandler, R.S. Selenium, folate, and colon cancer. Nutr. Cancer 2009, 61, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Searle, E.; Kelly, L.; Johnsen, S.; Bodman-Smith, K.; Bath, S.C.; Mao, J.; Redman, C.W. Effect of selenium on markers of risk of pre-eclampsia in UK pregnant women: A randomised, controlled pilot trial. Br. J. Nutr. 2014, 112, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Skroder, H.M.; Hamadani, J.D.; Tofail, F.; Persson, L.A.; Vahter, M.E.; Kippler, M.J. Selenium status in pregnancy influences children’s cognitive function at 1.5 years of age. Clin. Nutr. 2014. [Google Scholar] [CrossRef]
- Ren, Y.; Wang, Q.; Shi, L.; Yue, W.; Zhang, C.; Lei, F. Effects of maternal and dietary selenium (Se-enriched yeast) on the expression of p34(cdc2) and CyclinB1 of germ cells of their offspring in goats. Anim. Reprod. Sci. 2011, 123, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Nogales, F.; Ojeda, M.L.; Fenutria, M.; Murillo, M.L.; Carreras, O. Role of selenium and glutathione peroxidase on development, growth, and oxidative balance in rat offspring. Reproduction 2013, 146, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Scholl, T.O.; Johnson, W.G. Folic acid: Influence on the outcome of pregnancy. Am. J. Clin. Nutr. 2000, 71, 1295S–1303S. [Google Scholar] [PubMed]
- McNulty, B.; Pentieva, K.; Marshall, B.; Ward, M.; Molloy, A.M.; Scott, J.M.; McNulty, H. Women’s compliance with current folic acid recommendations and achievement of optimal vitamin status for preventing neural tube defects. Hum. Reprod. 2011, 26, 1530–1536. [Google Scholar] [CrossRef] [PubMed]
- Sillender, M.; Pring, D.W. How effective was the Health Education Authority’s folic acid campaign? J. Obstet. Gynaecol. 2000, 20, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.G.; Singh, G.; Burrows, R.F. Evidence for suboptimal use of periconceptional folic acid supplements globally. BJOG 2004, 111, 399–408. [Google Scholar] [CrossRef] [PubMed]
- Department of Health. Folic Acid and the Prevention of Disease. Report on Health and Social Subjects; DOH Health Publication Unit: Heywood, Australia, 2002; p. 50. [Google Scholar]
- Burdge, G.C.; Lillycrop, K.A.; Jackson, A.A.; Gluckman, P.D.; Hanson, M.A. The nature of the growth pattern and of the metabolic response to fasting in the rat are dependent upon the dietary protein and folic acid intakes of their pregnant dams and post-weaning fat consumption. Br. J. Nutr. 2008, 99, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Geillinger, K.E.; Rathmann, D.; Kohrle, J.; Fiamoncini, J.; Daniel, H.; Kipp, A.P. Hepatic metabolite profiles in mice with a suboptimal selenium status. J. Nutr. Biochem. 2014, 25, 914–922. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.A.; Williams, E.A.; Mathers, J.C. Gender-specific modulation of tumorigenesis by folic acid supply in the Apc mouse during early neonatal life. Br. J. Nutr. 2008, 99, 550–558. [Google Scholar] [CrossRef] [PubMed]
- McKay, J.A.; Williams, E.A.; Mathers, J.C. Effect of maternal and post-weaning folate supply on gene-specific DNA methylation in the small intestine of weaning and adult apc and wild type mice. Front. Genet. 2011, 2, 23. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.M.; Neville, T.L.; Reed, J.J.; Taylor, J.B.; Reynolds, L.P.; Redmer, D.A.; Hammer, C.J.; Vonnahme, K.A.; Caton, J.S. Maternal nutritional plane and selenium supply during gestation impact visceral organ mass and intestinal growth and vascularity of neonatal lamb offspring. J. Anim. Sci. 2013, 91, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Pericleous, M.; Mandair, D.; Caplin, M.E. Diet and supplements and their impact on colorectal cancer. J. Gastrointest. Oncol. 2013, 4, 409–423. [Google Scholar] [PubMed]
- Daniels, L.A. Selenium metabolism and bioavailability. Biol. Trace Elem. Res. 1996, 54, 185–199. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, M.L.; Nogales, F.; Jotty, K.; Delgado, M.J.; Guerrero-Leon, M.M.; Murillo, M.L.; Carreras, O. Folic acid and selenite during reproduction, gestation and lactation protect against ethanol changed Se bioavailability. Alcohol Alcohol. 2010, 45, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Chanson, A.; Sayd, T.; Rock, E.; Chambon, C.; Sante-Lhoutellier, V.; Potier de Courcy, G.; Brachet, P. Proteomic analysis reveals changes in the liver protein pattern of rats exposed to dietary folate deficiency. J. Nutr. 2005, 135, 2524–2529. [Google Scholar] [PubMed]
- Schaible, T.D.; Harris, R.A.; Dowd, S.E.; Smith, C.W.; Kellermayer, R. Maternal methyl-donor supplementation induces prolonged murine offspring colitis susceptibility in association with mucosal epigenetic and microbiomic changes. Hum. Mol. Genet. 2011, 20, 1687–1696. [Google Scholar] [CrossRef] [PubMed]
- Bermingham, E.N.; Bassett, S.A.; Young, W.; Roy, N.C.; McNabb, W.C.; Cooney, J.M.; Brewster, D.T.; Laing, W.A.; Barnett, M.P. Post-weaning selenium and folate supplementation affects gene and protein expression and global DNA methylation in mice fed high-fat diets. BMC Med. Genom. 2013, 6, 7. [Google Scholar] [CrossRef]
- Joseph, S.B.; McKilligin, E.; Pei, L.; Watson, M.A.; Collins, A.R.; Laffitte, B.A.; Chen, M.; Noh, G.; Goodman, J.; Hagger, G.N.; et al. Synthetic LXR ligand inhibits the development of atherosclerosis in mice. Proc. Natl. Acad. Sci. USA 2002, 99, 7604–7609. [Google Scholar] [CrossRef] [PubMed]
- Bradley, M.N.; Hong, C.; Chen, M.; Joseph, S.B.; Wilpitz, D.C.; Wang, X.; Lusis, A.J.; Collins, A.R.; Hsueh, W.A.; Collins, J.L.; et al. Ligand activation of LXRβ reverses atherosclerosis and cellular cholesterol overload in mice lacking LXRα and apoE. J. Clin. Investig. 2007, 117, 2337–2346. [Google Scholar] [CrossRef] [PubMed]
- Davis, C.; Uthus, E. Dietary folate and selenium affect dimethylhydrazine-induced aberrant crypt formation, global DNA methylation and one-carbon metabolism in rats. J. Nutr. 2003, 133, 2907–2914. [Google Scholar] [PubMed]
- Ghoshal, K.; Li, X.; Datta, J.; Bai, S.; Pogribny, I.; Poribnh, M.; Huang, Y.; Young, D.; Jacob, S. A folate- and methyl-deficient diet alters the expression of DNA methyltransferases and methyl CpG binding proteins invlolved in epigenetic gene silencing in livers of F344 rats. J. Nutr. 2006, 136, 1522–1527. [Google Scholar] [PubMed]
- Thompson, K.M.; Haibach, H.; Evenson, J.K.; Sunde, R.A. Liver Selenium and Testis Phospholipid Hydroperoxide Glutathione Peroxidase Are Associated with Growth during Selenium Repletion of Second-Generation Se-Deficient Male Rats. J. Nutr. 1998, 128, 1289–1295. [Google Scholar] [PubMed]
- National Research Council Subcommittee on Laboratory Animal Nutrition. Nutrient Requirements of Laboratory Animals: Fourth Revised Edition, 1995; National Academy of Sciences: Washington, DC, USA, 1995; p. 173. [Google Scholar]
- Park, E.I.; Paisley, E.A.; Mangian, H.J.; Swartz, D.A.; Wu, M.X.; O’Morchoe, P.J.; Behr, S.R.; Visek, W.J.; Kaput, J. Lipid level and type alter stearoyl CoA desaturase mRNA abundance differently in mice with distinct susceptibilities to diet-influenced diseases. J. Nutr. 1997, 127, 566–573. [Google Scholar] [PubMed]
- Knoch, B.; Barnett, M.P.G.; Zhu, S.; Park, Z.A.; Nones, K.; Dommels, Y.E.M.; Knowles, S.O.; McNabb, W.C.; Roy, N.C. Genome-wide analysis of dietary eicosapentaenoic acid- and oleic acid-induced modulation of colon inflammation in interleukin-10 gene-deficient mice. J. Nutrigenet. Nutrigenomics 2009, 2, 9–28. [Google Scholar] [CrossRef] [PubMed]
- Roy, N.C.; Barnett, M.P.G.; Knoch, B.; Dommels, Y.E.M.; McNabb, W.C. Nutrigenomics applied to an animal model of Inflammatory Bowel Diseases: Transcriptomic analysis of the effects of eicosapentaenoic acid- and arachidonic acid-enriched diets. Mutat. Res. 2007, 622, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Smyth, G.K. Limma: Linear models for microarray data. In Bioinformatics and Computational Biology Solutions Using R and Bioconductor; Gentleman, R., Carey, V., Dudoit, S., Irizarry, R., Huber, W., Eds.; Springer: New York, NY, USA, 2005; pp. 397–420. [Google Scholar]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.H.; Pages, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Dejean, S.; Gonzalez, I.; Le Cao, K.A.; Monget, P.; Coquery, J.; Yao, F.Z.; Liquet, B. mixOmics: Omics Data Integration Project, R Package Version 4.1–4; 2013. Available online: http://CRAN.R-project.org/package=mixOmics (access on 5 May 2015).
- Perez-Enciso, M.; Tenenhaus, M. Prediction of clinical outcome with microarray data: A partial least squares discriminant analysis (PLS-DA) approach. Hum. Genet. 2003, 112, 581–592. [Google Scholar] [PubMed]
- Wu, D.; Lim, E.; Vaillant, F.; Asselin-Labat, M.L.; Visvader, J.E.; Smyth, G.K. ROAST: Rotation gene set tests for complex microarray experiments. Bioinformatics 2010, 26, 2176–2182. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Gentleman, R. Extensions to gene set enrichment. Bioinformatics 2007, 23, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed]
- Matthews, L.; Gopinath, G.; Gillespie, M.; Caudy, M.; Croft, D.; de Bono, B.; Garapati, P.; Hemish, J.; Hermjakob, H.; Jassal, B.; et al. Reactome knowledgebase of human biological pathways and processes. Nucleic Acids Res. 2009, 37, D619–D622. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.; Domrachev, M.; Lash, A.E. Gene Expression Omnibus: NCBI gene expression and hybridization array data repository. Nucleic Acids Res. 2002, 30, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Cornelis, M.C.; Fornage, M.; Foy, M.; Xun, P.; Gladyshev, V.N.; Morris, S.; Chasman, D.I.; Hu, F.B.; Rimm, E.B.; Kraft, P.; et al. Genome-wide association study of selenium concentrations. Hum. Mol. Genet. 2015, 24, 1469–1477. [Google Scholar] [CrossRef] [PubMed]
- Pagmantidis, V.; Meplan, C.; van Schothorst, E.M.; Keijer, J.; Hesketh, J.E. Supplementation of healthy volunteers with nutritionally relevant amounts of selenium increases the expression of lymphocyte protein biosynthesis genes. Am. J. Clin. Nutr. 2008, 87, 181–189. [Google Scholar] [PubMed]
- Brostrom, C.O.; Brostrom, M.A. Regulation of translational initiation during cellular responses to stress. Prog. Nucleic. Acid Res. Mol. Biol. 1998, 58, 79–125. [Google Scholar] [PubMed]
- Hesketh, J. Nutrigenomics and selenium: Gene expression patterns, physiological targets, and genetics. Ann. Rev. Nutr. 2008, 28, 157–177. [Google Scholar] [CrossRef]
- Cole-Ezea, P.; Swan, D.; Shanley, D.; Hesketh, J. Glutathione peroxidase 4 has a major role in protecting mitochondria from oxidative damage and maintaining oxidative phosphorylation complexes in gut epithelial cells. Free Radic. Biol. Med. 2012, 53, 488–497. [Google Scholar] [CrossRef]
- Brutsch, S.H.; Wang, C.C.; Li, L.; Stender, H.; Neziroglu, N.; Richter, C.; Kuhn, H.; Borchert, A. Expression of inactive glutathione peroxidase 4 leads to embryonic lethality, and inactivation of the alox15 gene does not rescue such knock-in mice. Antioxid. Redox Signal. 2015, 22, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Schafer, K.; Kyriakopoulos, A.; Gessner, H.; Grune, T.; Behne, D. Effects of selenium deficiency on fatty acid metabolism in rats fed fish oil-enriched diets. J. Trace Elem. Med. Biol. 2004, 18, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Opalinska, M.; Meisinger, C. Metabolic control via the mitochondrial protein import machinery. Curr. Opin. Cell Biol. 2014, 33, 42–48. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barnett, M.P.G.; Bermingham, E.N.; Young, W.; Bassett, S.A.; Hesketh, J.E.; Maciel-Dominguez, A.; McNabb, W.C.; Roy, N.C. Low Folate and Selenium in the Mouse Maternal Diet Alters Liver Gene Expression Patterns in the Offspring after Weaning. Nutrients 2015, 7, 3370-3386. https://doi.org/10.3390/nu7053370
Barnett MPG, Bermingham EN, Young W, Bassett SA, Hesketh JE, Maciel-Dominguez A, McNabb WC, Roy NC. Low Folate and Selenium in the Mouse Maternal Diet Alters Liver Gene Expression Patterns in the Offspring after Weaning. Nutrients. 2015; 7(5):3370-3386. https://doi.org/10.3390/nu7053370
Chicago/Turabian StyleBarnett, Matthew P.G., Emma N. Bermingham, Wayne Young, Shalome A. Bassett, John E. Hesketh, Anabel Maciel-Dominguez, Warren C. McNabb, and Nicole C. Roy. 2015. "Low Folate and Selenium in the Mouse Maternal Diet Alters Liver Gene Expression Patterns in the Offspring after Weaning" Nutrients 7, no. 5: 3370-3386. https://doi.org/10.3390/nu7053370