Codonopsis lanceolata Extract Prevents Diet-Induced Obesity in C57BL/6 Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Contents (mg/100 g Extract) |
|---|---|
| Phenolic acid | |
| Phloroglucinol | 924.58 |
| Gallic acid | 85.28 |
| Chlorogenic acid | 32.99 |
| Caffeic acid | 37.18 |
| p-Coumaric acid | 2.23 |
| Trans-ferulic acid | 16.40 |
| Total of phenolic acid | 1098.66 |
| Flavonoids | |
| (−)-Epigallocatechin | 1353.00 |
| Epigallocatechin gallate | 20.64 |
| Rutin hydrate | 391.08 |
| Luteolin | 12.12 |
| Total of flavonoids | 1776.84 |
2.2. Anti-Obesity Activity of Codonopsis lanceolata Root Extract
2.2.1. Study Design
2.2.2. Blood Chemistry
2.2.3. Specimen Collection
2.2.4. Determination of Fat Size of Adipose Tissue by Oil Red O Staining
2.2.5. Histology Preparation and Hematoxylin and Eosin Stain (H & E)
2.3. Statistical Analysis
3. Results
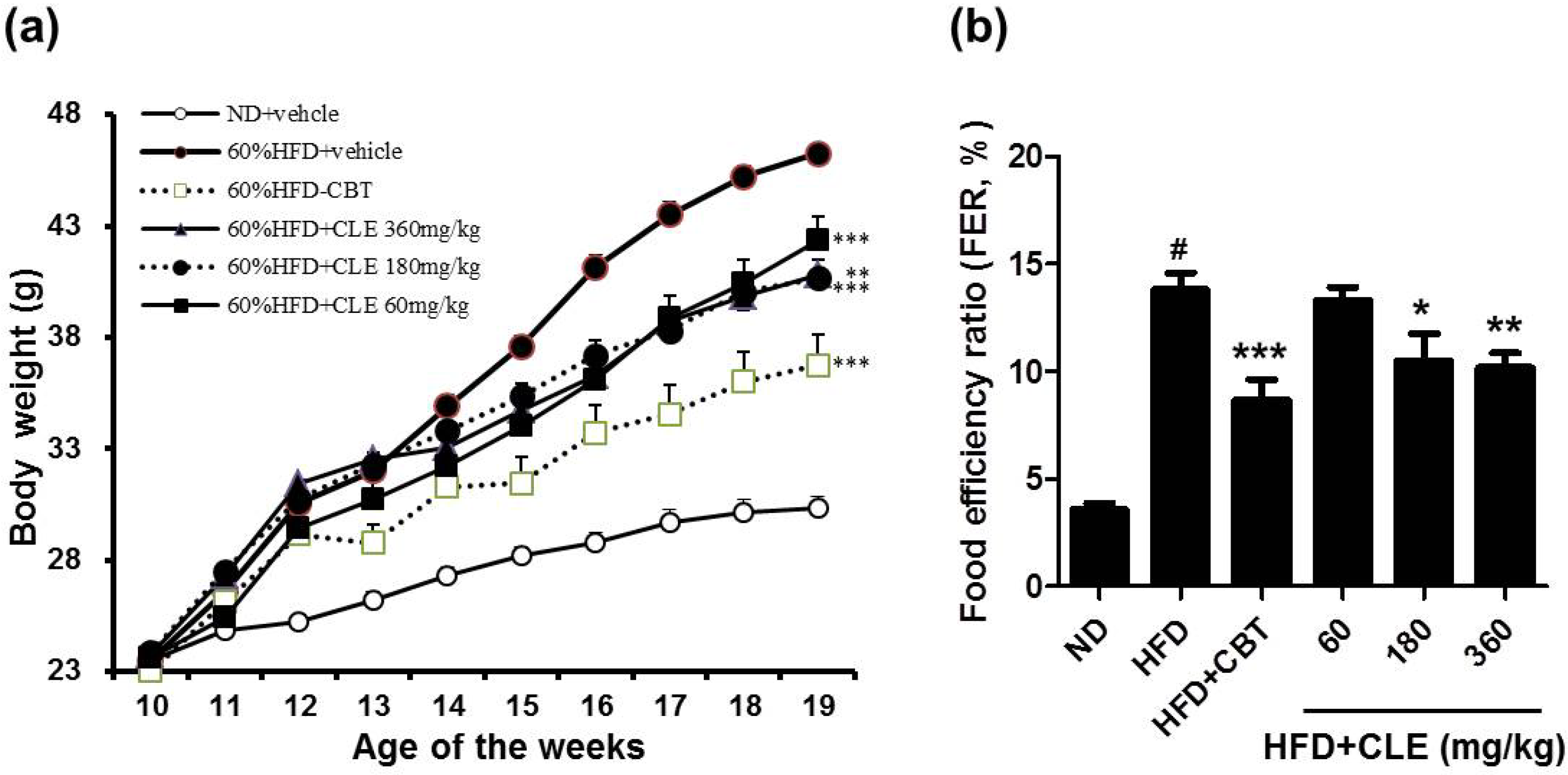
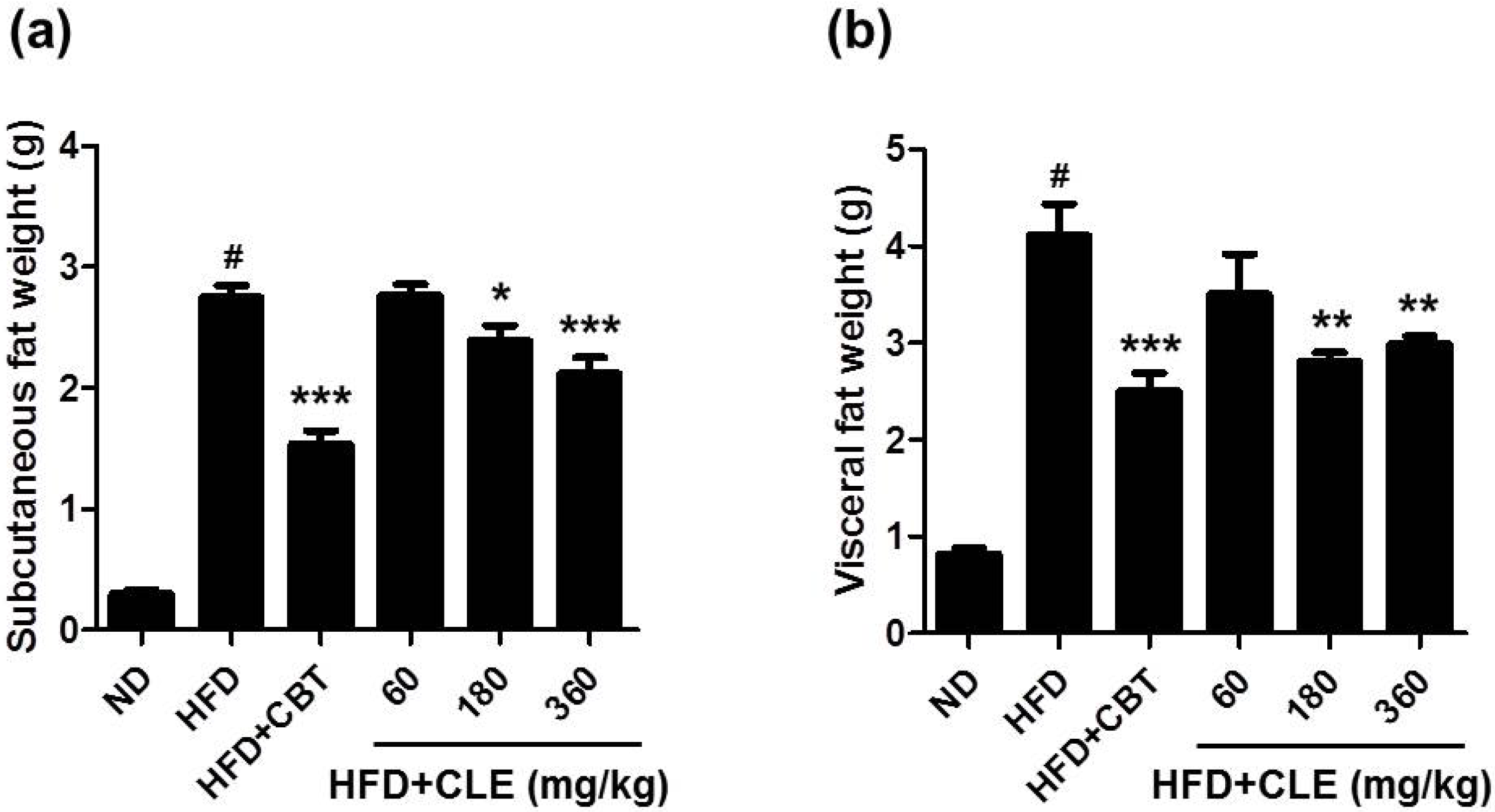
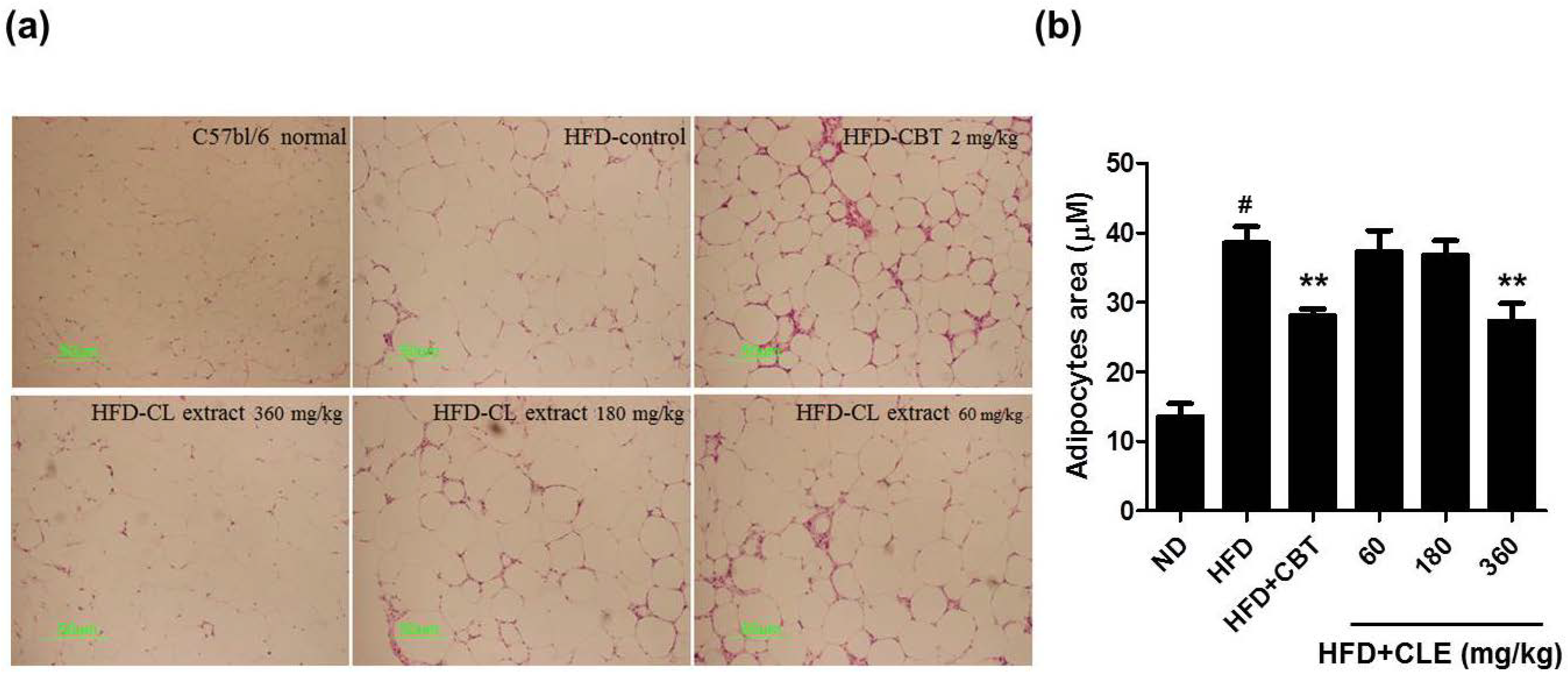
3.1. CLE Suppresses Body Weight Gain and Body Fat Accumulation in C57BL/6 Mice Fed HFD



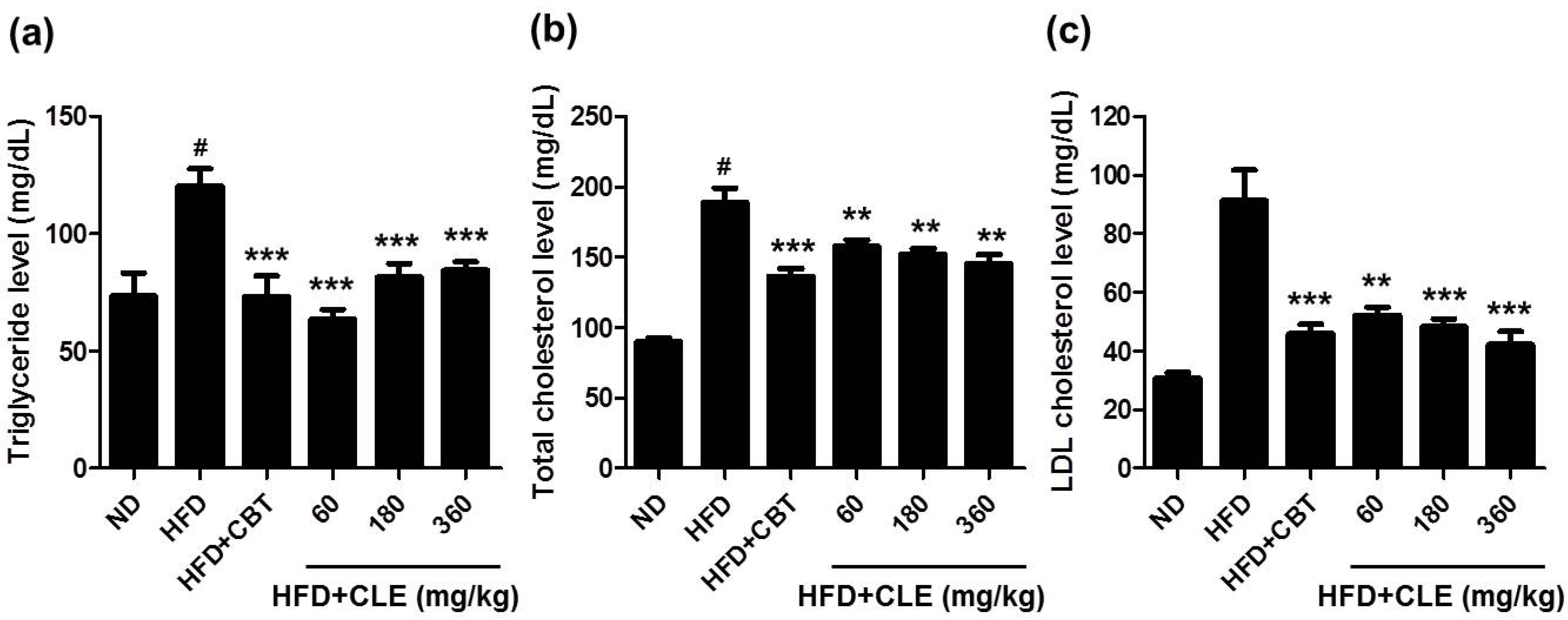
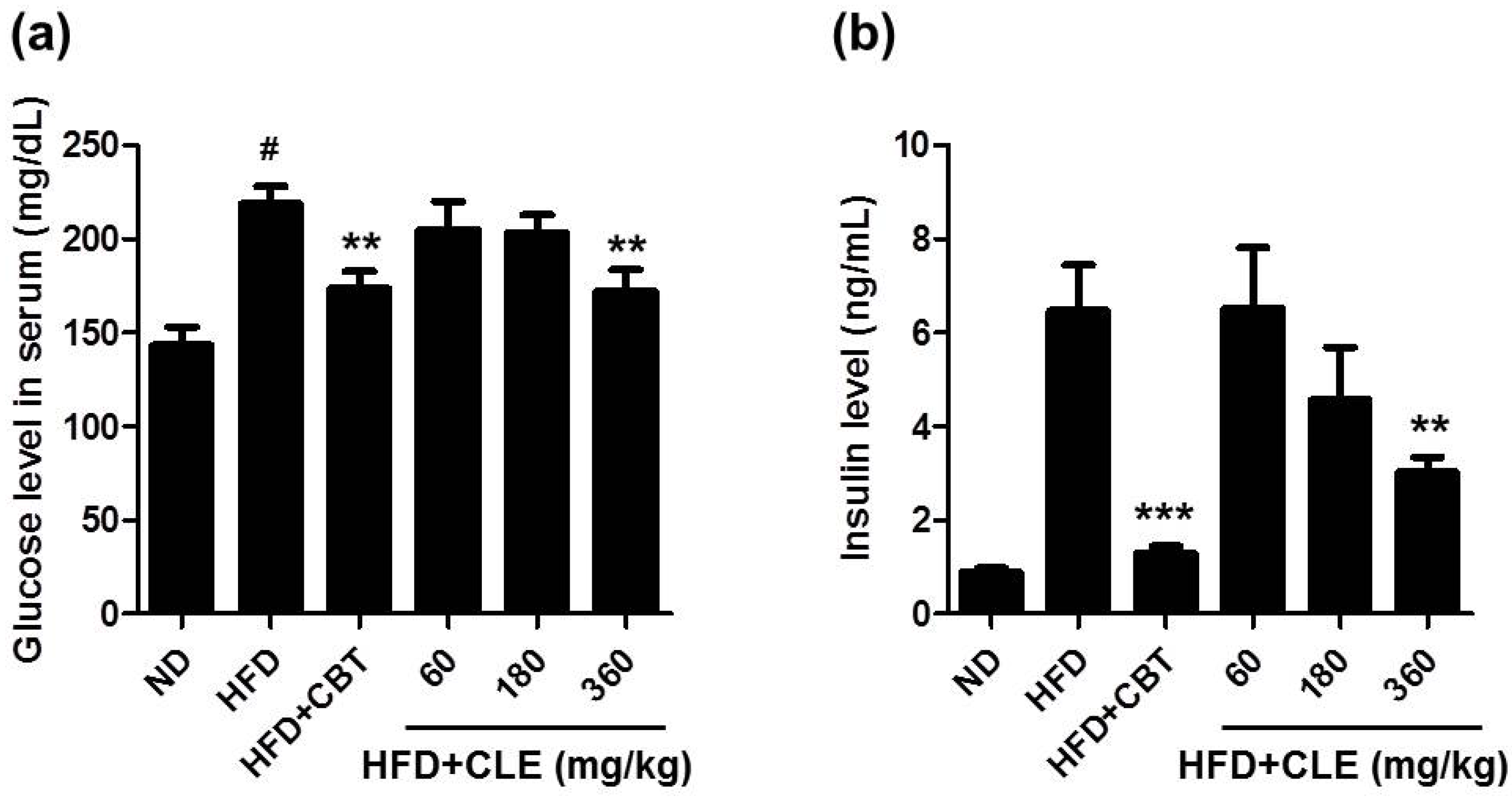
3.2. CLE Ameliorate Impaired Lipid, Glucose, and Insulin Homeostasis in HFD-Induced Obese Mice


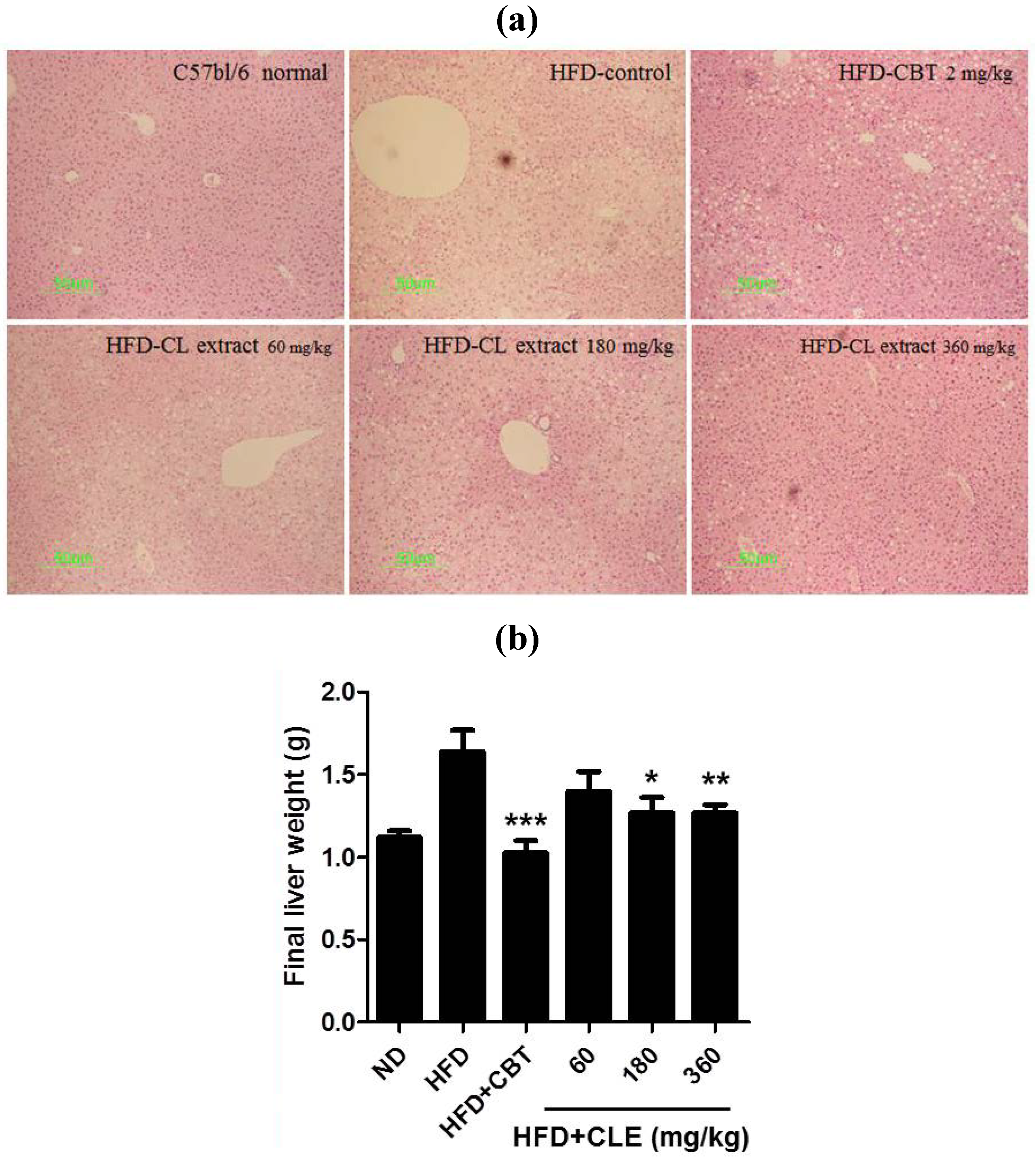
3.3. CLE Suppresses Hepatic Steatosis in HFD-Induced Obese Mice


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- De Onis, M.; Martinez-Costa, C.; Nunez, F.; Nguefack-Tsague, G.; Montal, A.; Brines, J. Association between WHO cut-offs for childhood overweight and obesity and cardiometabolic risk. Public Health Nutr. 2013, 16, 625–630. [Google Scholar] [PubMed]
- Rocha, V.Z.; Libby, P. Obesity, inflammation, and atherosclerosis. Nat. Rev. Cardiol. 2009, 6, 399–409. [Google Scholar]
- Thomas, S.A.; Matsumoto, A.M.; Palmiter, R.D. Noradrenaline is essential for mouse fetal development. Nature 1995, 374, 643–646. [Google Scholar] [PubMed]
- D’souza, A.M.; Beaudry, J.L.; Szigiato, A.A.; Trumble, S.J.; Snook, L.A.; Bonen, A.; Giacca, A.; Riddell, M.C. Consumption of a high-fat diet rapidly exacerbates the development of fatty liver disease that occurs with chronically elevated glucocorticoids. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 302, G850–G863. [Google Scholar] [CrossRef]
- Lichtenstein, A.H.; Kennedy, E.; Barrier, P.; Danford, D.; Ernst, N.D.; Grundy, S.M.; Leveille, G.A.; van Horn, L.; Williams, C.L.; Booth, S.L. Dietary fat consumption and health. Nutr. Rev. 1998, 56, S3–S19. [Google Scholar]
- Davey, R.C. The obesity epidemic: Too much food for thought? Br. J. Sports Med. 2004, 38, 360–363. [Google Scholar] [CrossRef]
- Halford, J.C. Pharmacotherapy for obesity. Appetite 2006, 46, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Siebenhofer, A.; Jeitler, K.; Horvath, K.; Berghold, A.; Siering, U.; Semlitsch, T. Long-term effects of weight-reducing drugs in hypertensive patients. Cochrane Database Syst. Rev. 2013, 3, CD007654. [Google Scholar] [CrossRef]
- Baur, J.A.; Pearson, K.J.; Price, N.L.; Jamieson, H.A.; Lerin, C.; Kalra, A.; Prabhu, V.V.; Allard, J.S.; Lopez-Lluch, G.; Lewis, K.; et al. Resveratrol improves health and survival of mice on a high-calorie diet. Nature 2006, 444, 337–342. [Google Scholar]
- Cha, Y.S.; Rhee, S.J.; Heo, Y.R. Acanthopanax senticosus extract prepared from cultured cells decreases adiposity and obesity indices in C57BL/6J mice fed a high fat diet. J. Med. Food 2004, 7, 422–429. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, K.J.; Park, K.J.; Yoon, B.R.; Lim, J.H.; Lee, O.H. Buckwheat (Fagopyrum esculentum M.) sprout treated with methyl jasmonate (MeJA) improved anti-adipogenic activity associated with the oxidative stress system in 3T3-L1 adipocytes. Int. J. Mol. Sci. 2013, 14, 1428–1442. [Google Scholar]
- He, X.; Kim, S.S.; Park, S.J.; Seong, D.H.; Yoon, W.B.; Lee, H.Y.; Park, D.S.; Ahn, J. Combined effects of probiotic fermentation and high-pressure extraction on the antioxidant, antimicrobial, and antimutagenic activities of deodeok (Codonopsis lanceolata). J. Agric. Food Chem. 2010, 58, 1719–1725. [Google Scholar] [CrossRef]
- Wang, L.; Xu, M.L.; Hu, J.H.; Rasmussen, S.K.; Wang, M.H. Codonopsis lanceolata extract induces G0/G1 arrest and apoptosis in human colon tumor HT-29 cells—Involvement of ROS generation and polyamine depletion. Food Chem. Toxicol. 2011, 49, 149–154. [Google Scholar] [CrossRef]
- Ha do, T.; Nam Trung, T.; Bich Thu, N.; van On, T.; Hai Nam, N.; van Men, C.; Thi Phuong, T.; Bae, K. Adlay seed extract (Coix lachryma-jobi L.) decreased adipocyte differentiation and increased glucose uptake in 3T3-L1 cells. J. Med. Food 2010, 13, 1331–1339. [Google Scholar]
- Choi, H.K.; Won, E.K.; Jang, Y.P.; Choung, S.Y. Antiobesity effect of Codonopsis lanceolata in high-calorie/high-fat-diet-induced obese rats. Evid. Based Complement. Altern. Med. 2013, 2013, 210297. [Google Scholar] [CrossRef]
- Cho, K.; Kim, S.J.; Park, S.H.; Kim, S.; Park, T. Protective effect of Codonopsis lanceolata root extract against alcoholic fatty liver in the rat. J. Med. Food 2009, 12, 1293–1301. [Google Scholar] [CrossRef]
- Sclafani, A. Animal models of obesity: Classification and characterization. Int. J. Obes. 1984, 8, 491–508. [Google Scholar]
- Johnson, P.R.; Greenwood, M.R.; Horwitz, B.A.; Stern, J.S. Animal models of obesity: Genetic aspects. Annu. Rev. Nutr. 1991, 11, 325–353. [Google Scholar] [CrossRef] [PubMed]
- Rossmeisl, M.; Rim, J.S.; Koza, R.A.; Kozak, L.P. Variation in type 2 diabetes—Related traits in mouse strains susceptible to diet-induced obesity. Diabetes 2003, 52, 1958–1966. [Google Scholar] [PubMed]
- Speakman, J.; Hambly, C.; Mitchell, S.; Krol, E. Animal models of obesity. Obes. Rev. 2007, 8, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Kim, Y.H.; Kim, D.B.; Bang, W.S.; Lee, O.H. Acute and 4-week repeated-dose oral toxicity studies of Cirsium setidens in rats. Molecules 2014, 19, 7138–7151. [Google Scholar] [CrossRef]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS One 2014, 9, e96969. [Google Scholar] [CrossRef]
- Inazawa, Y.; Nakatsu, M.; Yasugi, E.; Saeki, K.; You, A. Lipid droplet formation in human myeloid NB4 cells stimulated by all trans retinoic acid and granulocyte colony-stimulating factor: Possible involvement of peroxisome proliferator-activated receptor gamma. Cell Struct. Funct. 2003, 28, 487–493. [Google Scholar] [PubMed]
- Kim, K.J.; Godarova, A.; Seedle, K.; Kim, M.H.; Ince, T.A.; Wells, S.I.; Driscoll, J.J.; Godar, S. Rb suppresses collective invasion, circulation and metastasis of breast cancer cells in CD44-dependent manner. PLoS One 2013, 8, e80590. [Google Scholar] [CrossRef]
- Collins, S.; Martin, T.L.; Surwit, R.S.; Robidoux, J. Genetic vulnerability to diet-induced obesity in the C57BL/6J mouse: Physiological and molecular characteristics. Physiol. Behav. 2004, 81, 243–248. [Google Scholar] [PubMed]
- Wang, C.Y.; Liao, J.K. A mouse model of diet-induced obesity and insulin resistance. Methods Mol. Biol. 2012, 821, 421–433. [Google Scholar] [PubMed]
- Rolo, A.P.; Teodoro, J.S.; Palmeira, C.M. Role of oxidative stress in the pathogenesis of nonalcoholic steatohepatitis. Free Radic. Biol. Med. 2012, 52, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Nanji, A.A.; French, S.W.; Freeman, J.B. Serum alanine aminotransferase to aspartate aminotransferase ratio and degree of fatty liver in morbidly obese patients. Enzyme 1986, 36, 266–269. [Google Scholar] [PubMed]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Patch, C.S.; Sullivan, D.R.; Fenech, M.; Roodenrys, S.; Keogh, J.B.; Clifton, P.M.; Williams, P.G.; et al. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust. 2006, 185, S4–S24. [Google Scholar]
- Tundis, R.; Deguin, B.; Loizzo, M.R.; Bonesi, M.; Statti, G.A.; Tillequin, F.; Menichini, F. Potential antitumor agents: Flavones and their derivatives from Linaria reflexa Desf. Bioorg. Med. Chem. Lett. 2005, 15, 4757–4760. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, D.O.; Wightman, E.L. Herbal extracts and phytochemicals: Plant secondary metabolites and the enhancement of human brain function. Adv. Nutr. 2011, 2, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, M.; Ohta, S.; Komoto, N.; Ushijima, M.; Kodera, Y.; Hayama, M.; Shirota, O.; Sekita, S.; Kuroyanagi, M. Simultaneous determination of seven saponins in the roots of Codonopsis lanceolata by liquid chromatography-mass spectrometry. J. Nat. Med. 2009, 63, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.T.; Choi, J.; Jung, W.T.; Nam, J.H.; Jung, H.J.; Park, H.J. Structure of a new echinocystic acid bisdesmoside isolated from Codonopsis lanceolata roots and the cytotoxic activity of prosapogenins. J. Agric. Food Chem. 2002, 50, 4190–4193. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Jung, H.J.; Jeong, H.Y.; Kwon, H.J.; Ali, M.Y.; Choi, J.S. Phlorotannins isolated from the edible brown alga Ecklonia stolonifera exert anti-adipogenic activity on 3T3-L1 adipocytes by downregulating C/EBPα and PPARγ. Fitoterapia 2014, 92, 260–269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.T.; Chang, C.Q.; Liu, Y.; Chen, Z.M. Effect of chlorogenic acid on disordered glucose and lipid metabolism in db/db mice and its mechanism. Zhongguo Yi Xue Ke Xue Yuan Xue Bao 2011, 33, 281–286. [Google Scholar]
- Shen, C.L.; Smith, B.J.; Lo, D.F.; Chyu, M.C.; Dunn, D.M.; Chen, C.H.; Kwun, I.S. Dietary polyphenols and mechanisms of osteoarthritis. J. Nutr. Biochem. 2012, 23, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Quon, M.J.; Kim, J.A. New insights into the mechanisms of polyphenols beyond antioxidant properties; lessons from the green tea polyphenol, epigallocatechin 3-gallate. Redox Biol. 2014, 2, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Klaus, S.; Pultz, S.; Thone-Reineke, C.; Wolfram, S. Epigallocatechin gallate attenuates diet-induced obesity in mice by decreasing energy absorption and increasing fat oxidation. Int. J. Obes. 2005, 29, 615–623. [Google Scholar] [CrossRef]
- Fujioka, S.; Matsuzawa, Y.; Tokunaga, K.; Tarui, S. Contribution of intra-abdominal fat accumulation to the impairment of glucose and lipid metabolism in human obesity. Metabolism 1987, 36, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Hokanson, J.E.; Austin, M.A. Plasma triglyceride level is a risk factor for cardiovascular disease independent of high-density lipoprotein cholesterol level: A meta-analysis of population-based prospective studies. J. Cardiovasc. Risk 1996, 3, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Bush, T.L.; Fried, L.P.; Barrett-Connor, E. Cholesterol, lipoproteins, and coronary heart disease in women. Clin. Chem. 1988, 34, B60–B70. [Google Scholar]
- Manninen, V.; Elo, M.O.; Frick, M.H.; Haapa, K.; Heinonen, O.P.; Heinsalmi, P.; Helo, P.; Huttunen, J.K.; Kaitaniemi, P.; Koskinen, P.; et al. Lipid alterations and decline in the incidence of coronary heart disease in the Helsinki Heart Study. JAMA 1988, 260, 641–651. [Google Scholar]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Bais, S.; Singh, G.S.; Sharma, R. Antiobesity and hypolipidemic activity of Moringa oleifera leaves against high fat diet-induced obesity in rats. Adv. Biol. 2014, 2014, 1–9. [Google Scholar] [CrossRef]
- Falck-Ytter, Y.; Younossi, Z.M.; Marchesini, G.; McCullough, A.J. Clinical features and natural history of nonalcoholic steatosis syndromes. Semin. Liver Dis. 2001, 21, 17–26. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.S.; Kim, K.-J.; Kim, Y.-H.; Kim, D.-B.; Shin, G.-H.; Cho, J.-H.; Kim, B.K.; Lee, B.-Y.; Lee, O.-H. Codonopsis lanceolata Extract Prevents Diet-Induced Obesity in C57BL/6 Mice. Nutrients 2014, 6, 4663-4677. https://doi.org/10.3390/nu6114663
Lee JS, Kim K-J, Kim Y-H, Kim D-B, Shin G-H, Cho J-H, Kim BK, Lee B-Y, Lee O-H. Codonopsis lanceolata Extract Prevents Diet-Induced Obesity in C57BL/6 Mice. Nutrients. 2014; 6(11):4663-4677. https://doi.org/10.3390/nu6114663
Chicago/Turabian StyleLee, Jong Seok, Kui-Jin Kim, Young-Hyun Kim, Dan-Bi Kim, Gi-Hae Shin, Ju-Hyun Cho, Bong Kyun Kim, Boo-Yong Lee, and Ok-Hwan Lee. 2014. "Codonopsis lanceolata Extract Prevents Diet-Induced Obesity in C57BL/6 Mice" Nutrients 6, no. 11: 4663-4677. https://doi.org/10.3390/nu6114663