
The Interaction between Psychological Stress and Iron Status on Early-Life Neurodevelopmental Outcomes

1
Department of Psychiatry and Human Behavior, Warren Alpert Medical School, Brown University, Providence, RI 02912, USA
2
Center for Behavioral and Preventive Medicine, The Miriam Hospital, Providence, RI 02906, USA
3
Division of Neonatology, Department of Pediatrics, University of Minnesota Medical School, Minneapolis, MN 55455, USA
*
Author to whom correspondence should be addressed.
Nutrients 2023, 15(17), 3798; https://doi.org/10.3390/nu15173798
Submission received: 28 July 2023
/
Revised: 18 August 2023
/
Accepted: 22 August 2023
/
Published: 30 August 2023
(This article belongs to the Special Issue Iron Deficiency and Iron-Related Disorders)
Abstract
:This review presents evidence from animal and human studies demonstrating the possible connection and significant impact of poor iron status and psychological distress on neurocognitive development during pregnancy and the neonatal period, with implications for long-term cognition. Stress and iron deficiency are independently prevalent and thus are frequently comorbid. While iron deficiency and early-life stress independently contribute to long-term neurodevelopmental alterations, their combined effects remain underexplored. Psychological stress responses may engage similar pathways as infectious stress, which alters fundamental iron metabolism processes and cause functional tissue-level iron deficiency. Psychological stress, analogous to but to a lesser degree than infectious stress, activates the hypothalamic–pituitary–adrenocortical (HPA) axis and increases proinflammatory cytokines. Chronic or severe stress is associated with dysregulated HPA axis functioning and a proinflammatory state. This dysregulation may disrupt iron absorption and utilization, likely mediated by the IL-6 activation of hepcidin, a molecule that impedes iron absorption and redistributes total body iron. This narrative review highlights suggestive studies investigating the relationship between psychological stress and iron status and outlines hypothesized mechanistic pathways connecting psychological stress exposure and iron metabolism. We examine findings regarding the overlapping impacts of early stress exposure to iron deficiency and children’s neurocognitive development. We propose that studying the influence of psychological stress on iron metabolism is crucial for comprehending neurocognitive development in children exposed to prenatal and early postnatal stressors and for children at risk of early iron insufficiency. We recommend future directions for dual-exposure studies exploring iron as a potential mediating pathway between early stress and offspring neurodevelopment, offering opportunities for targeted interventions.
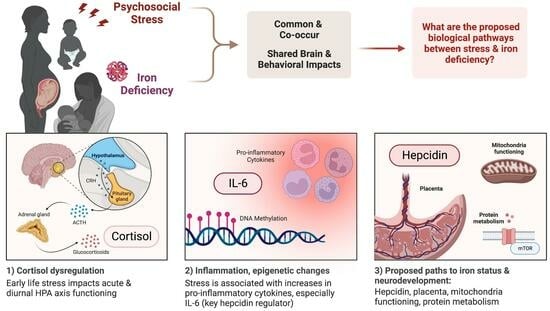
1. Introduction
The micronutrient iron is critical for the developing brain [1]. Iron deficiency’s impact on the developing brain has been reviewed extensively and includes acute effects on neurodevelopment alongside increased risk to lifelong mental health [2]. In total, 20–30% of all pregnant women have iron deficiency anemia (IDA), and 40% of children under five years of age are iron-deficient (ID; [3]). Though intervention through iron supplementation can support neurodevelopment, there is growing concern that oral iron treatment might not reverse all neurodevelopmental risks of ID, especially in the context of systemic inflammation [4,5]. While intervention research has focused on reducing inflammation through reducing pathways to infectious stress (e.g., integrated water, sanitation, and hygiene interventions), the contributions of how non-infectious psychological stress have yet to be rigorously evaluated. Non-infectious psychological stress responses utilize many of the same pathways as infectious stress to alter basic processes of nutrient metabolism, including absorption and prioritization [6]. Like infectious stress, psychological stress activates and, if chronic, dysregulates the hypothalamic–pituitary–adrenocortical (HPA) axis and increases proinflammatory cytokines [7]. The dysregulated neuroendocrine pathways that arise from chronic psychological stress can potentially disrupt iron absorption, distribution, and utilization, most likely through the activation of hepcidin by the proinflammatory cytokine IL-6, which is elevated in both infectious stress and psychological stress. Even with adequate iron intake, the activation of hepcidin by stress may disrupt maternal, fetal, and infant iron transport and distribution. Furthermore, iron deficiency and psychological stress independently affect similar neurodevelopmental outcomes. If maternal and infant stress and neuroendocrine dysregulation alter iron prioritization and loading through hepcidin, the resolution would suggest a non-nutritional—i.e., reduction in stress—instead of a nutritional solution—i.e., providing more iron.
This paper serves as a narrative review of the current literature to outline the effect of psychological stress pre- and neonatally and the risk it may carry for offspring iron status and neurodevelopment. Our central hypothesis is that chronic psychological stress, through the dysregulation of the HPA axis and proinflammatory mechanisms, negatively impacts maternal, fetal, and infant iron status, resulting in risk to early neurodevelopment. As the populations most at risk of iron deficiency are also those experiencing high burdens of psychological stress, we will first review psychological stress and the growing literature that examines associations between psychological stress and iron status. Second, we will review our hypothesized biological pathways between psychological stress and iron absorption and use, focusing on the HPA axis, inflammation, and hepcidin. Then, we will highlight the shared neurodevelopmental outcomes of pre- and early-life stress and iron deficiency. Finally, we will outline the current outstanding questions and future directions in this field of research. Our goal with this review is to create a framework for future studies to elucidate the effects of prenatal and neonatal stress and HPA mechanisms on the risk of functional iron deficiency in the mother and offspring iron outcomes. Ultimately, future research will help to identify outcome-specific biological mechanisms, markers, and modifiable risk factors as intervention targets.
2. Psychological Stress and Iron Status
2.1. Psychological Stress—Definitions and Prevalence
For our review, we focus on psychological stressors as either severely acute or chronic exposures that would threaten human survival throughout evolution [8] and require the individual’s stress-mediating systems to make physiological compensations to support allostasis [9]. This definition includes real and interpreted physical danger from another individual, whether through physical assault, abuse, or neglect, and social conflict and isolation [8]. For pregnant people, infants, and young children, stress exposure during these times is associated with altered offspring development [10]. Between 65% to 70% of pregnant women in the United States report at least one stressful life event during pregnancy [11,12], with higher rates for non-Hispanic Black women and American Indian/Alaska Native women [13]. Additionally, one in five women experiences multiple stressful life events while pregnant. Given the prevalence of maternal stress, low to moderate stress levels are likely adaptive. High or chronic levels likely carry risks. Around the world, ~300 million children aged two to four years experience stress in the form of physical and psychological violence [14], and infants under two are especially vulnerable [15]. One in four children under five lives with a mother who is a victim of intimate partner violence [16]. These stress exposures are especially relevant to iron deficiency, as populations and areas with a high prevalence of iron deficiency are also areas where many pregnant people and young children are exposed to chronic stressors such as food insecurity, poverty, and conflict [17,18,19,20].
2.2. Studies on Stress and Iron Status
2.2.1. Prenatal Stress and Offspring Iron Status
Exposure to stress, including measures of objective external stressors and self-reports of psychological distress, has been directly related to offspring iron status. A study of mothers living in an area under rocket attack in the first trimester of pregnancy had offspring with lower cord blood ferritin than the offspring of mothers who were pregnant after the rocket attacks ended [21]. Higher levels of maternal self-report subjective stress were also associated with lower cord blood ferritin concentration, especially for the group of mothers exposed to rocket-attack stress [21]. A study of 493 mother–infant dyads found that mothers with higher levels of prenatal perceived stress, violence exposure, and anxiety symptoms had infants with lower cord blood ferritin concentrations [22]. The impact of stress during pregnancy extends into infancy. Pregnant women who experienced higher stress levels during pregnancy had one-year-old infants at a higher risk of having low plasma ferritin [23]. Additionally, a study of pregnant rhesus macaques found that monkeys exposed to a laboratory stressor during pregnancy, particularly during the second trimester, had offspring with lower iron levels as they grew compared to unexposed offspring [24]. In multiple studies, evidence is accumulating to support how psychological stress prenatally places offspring at risk of worse iron status at birth and in infancy.
2.2.2. Postnatal Stress and Iron Status
Preclinical studies show evidence for changes in iron metabolism following postnatal stress exposure. Studies with rodents have shown that exposure to psychological stress for 1–2 weeks leads to decreased serum iron levels, increased hepatic iron content, and reduced protein expression of the iron exporter ferroportin (FPN) in the duodenum and liver [25,26,27,28]. In adult rodents, rodents exposed to psychological stress exhibit decreases in serum iron, hemoglobin, ferritin, and erythropoietin concentrations [26]. Psychological stress also decreases iron absorption and impairs iron transporter expression in the small intestine of adult rats [29]. Exposure to acute and chronic stressors in rodents reduces whole-blood iron concentration [25]. In contrast, the administration of dexamethasone for three weeks increases serum iron levels and reduces liver content and the expression of transferrin receptor 1 (TFR1) [30], while repeat restraint stress upregulates liver TFR2 expression and leads to liver iron accumulation [31]. As these studies were conducted using adult animals, it is unclear how these findings would relate to humans, especially prenatally and during infancy. Only a few studies have investigated the impact of stress on iron levels in humans. For instance, a study conducted on Navy SEAL trainees revealed that a week of psychological stress disrupted their iron levels [32]. Additionally, Chilean children exposed to accumulated family-level stressors during infancy were found to have poorer iron levels and were more likely to be diagnosed with IDA [33]. However, one drawback of the current research in this field is that there is a lack of studies conducted on infants, children, or animals in their early developmental stages.
3. Shared Neurodevelopmental Outcomes of Pre-Natal and Early-Life Stress and Iron Deficiency
The high demand for iron in infancy coincides with the rapid growth and development of brain structure and functions that require iron, including the hippocampus, cortical regions, neuronal and glial energy metabolism, myelin synthesis, and neurotransmission [34]. Iron is also essential for synthesizing serotonin, norepinephrine, and dopamine neurotransmitters [34]. Offspring of mothers with iron deficiency anemia (IDA) are at high risk for low birth weight, prematurity, small for gestational age, and poor neurodevelopment [2]. IDA in pregnancy can negatively impact the iron endowment of the neonate, which may cause irreversible harm to neurodevelopment [4,17]. Postnatally, iron-sufficient newborn infants are especially at risk of becoming iron-deficient (ID) between 6 and 12 months of age, when prenatal iron stores become depleted [2,5]. Iron deficiency is particularly damaging during the critical 6–24-month postnatal window of rapid brain development, as several key brain areas require iron for normal development [1].
Animal studies show associations between altered brain metabolism, myelination [35,36,37,38,39], neurotransmitter function [40], and early-life iron deficiency. Early ID is also associated with alterations to the developing hippocampus [41], with pervasive and long-lasting iron deficiency-induced metabolic [42] and dendritic structure changes [43]. Neurophysiologic studies of the effects of iron deficiency have found differences in the speed of neural transmission in the auditory system, recognition memory, longer auditory brainstem response latencies, and longer visual evoked potential latencies (for review, see [34]). Infants at high risk for ID show poorer recognition memory, possibly due to iron’s effects on the hippocampus and central nervous system. Iron deficiency during infancy is associated with children’s socioemotional and behavioral problems and lower cognitive abilities [40]. Thus, inadequate iron can negatively impact neurodevelopment across several domains and in different brain regions.
The consequences of fetal iron under-loading include abnormal acute neonatal brain function and long-term associations with mental health and psychopathology. Fetal iron under-loading during gestation carries the risk of earlier onset of postnatal iron deficiency, as the newborn is born with lower stores of iron which are depleted earlier. In turn, postnatal iron deficiency anemia is associated with both acute and long-term neurobehavioral abnormalities. Insufficient iron in the fetal period is associated with brain function that includes worse recognition memory [44,45], slower speed of neural processing [46], and poor bonding and maternal interaction [47]. Long-term, fetal iron deficiency is associated with an increased risk of autism spectrum disorder diagnosis (first trimester ID) [48], schizophrenia (second trimester ID) [49], and other neurocognitive impacts, including impaired memory performance (third trimester ID) [50,51]. Postnatal iron deficiency is associated with motor dysfunction [52], social-emotional behavior [52], and an increased risk of depression and anxiety in adulthood [53]. Research in rodent models shows that early iron deficiency causes specific anatomical, physiological, and molecular brain changes in different regions [42]. Iron deficiency during prenatal and neonatal stages impacts the developing hippocampus, a crucial structure for learning, memory, and the neuroendocrine stress response [43,54].
Similar to insufficient fetal iron loading, prenatal stress impairs learning [55], increases anxiety and depressive behaviors [56] (reviewed in [10]), and affects dopaminergic and hippocampal development [57,58]. Exposure to stress during prenatal development is linked to cortical thinning and reduced cognitive functioning in offspring [59]. Prenatal stress exposure is also associated with emotional reactivity in preschool-age offspring [60]. Over the lifespan, fetal exposure to maternal stress has been associated with an increased risk for autism spectrum disorder (ASD), schizophrenia [61], anxiety, depression, and ADHD [62,63]. Postnatal stress is also associated with neurobehavioral outcomes similar to iron deficiency, including impacts on learning, socioemotional development, and increased risk for mood disorders and psychopathology. Severe stress limited to infancy in the form of early institutional care impairs attention regulation and executive functioning (EF) (reviewed in [64,65,66]), thought to be due to changes in prefrontal-striatal and anterior cingulate circuitry [67]. Rodents exposed to early-life stress display alterations in synaptic signaling and epigenetics in the hippocampus and amygdala, linked to increased anxiety and depressive-like behaviors (reviewed in [68,69,70,71,72,73,74,75]). Rodents subjected to postnatal stress experience reduced dendritic arborization in both the PFC and hippocampus (reviewed in [68,76,77]). Studies have shown a connection between changes in hippocampal synaptic plasticity caused by stress and decreased spatial memory learning in rodents [68,78,79]. The neurodevelopmental outcomes of fetal and early postnatal iron insufficiency share similarities with those arising from fetal and early exposure to psychological stress, summarized in Table 1. Still, to our knowledge, no study has elucidated both exposures’ combined contributions to neurodevelopment.
4. Hypothesized Biological Pathways between Psychological Stress and Iron Status
Biological and sociological reasons exist for the co-occurrence of psychological stress and ID [82]. For instance, young children experiencing stress in the form of poverty may be at higher risk of iron deficiency due to food insecurity or diets low in iron (e.g., [83]). Maternal ID also commonly occurs among communities affected by poverty, which contributes to fetal iron under-loading. The rate of ID in pregnancy in the US is 42% [84], with higher rates in less-resourced populations [3]. Behaviorally, stress has been found to change how individuals eat, and pregnant women and children under stress may eat with a preference for high-fat, high-sugar foods that are likely lower in iron (reviewed in [85,86]). While a lack of access to or reduced preference for iron-rich foods is an important factor to consider, more relevant to this review are the biological changes associated with psychological stress that may impact iron status even in the context of adequate intake. The stress-induced biological changes we will focus on are the HPA axis, inflammation, hepcidin, actions on protein, and other potential mechanisms of note, outlined in Figure 1.
4.1. Stress and the HPA Axis
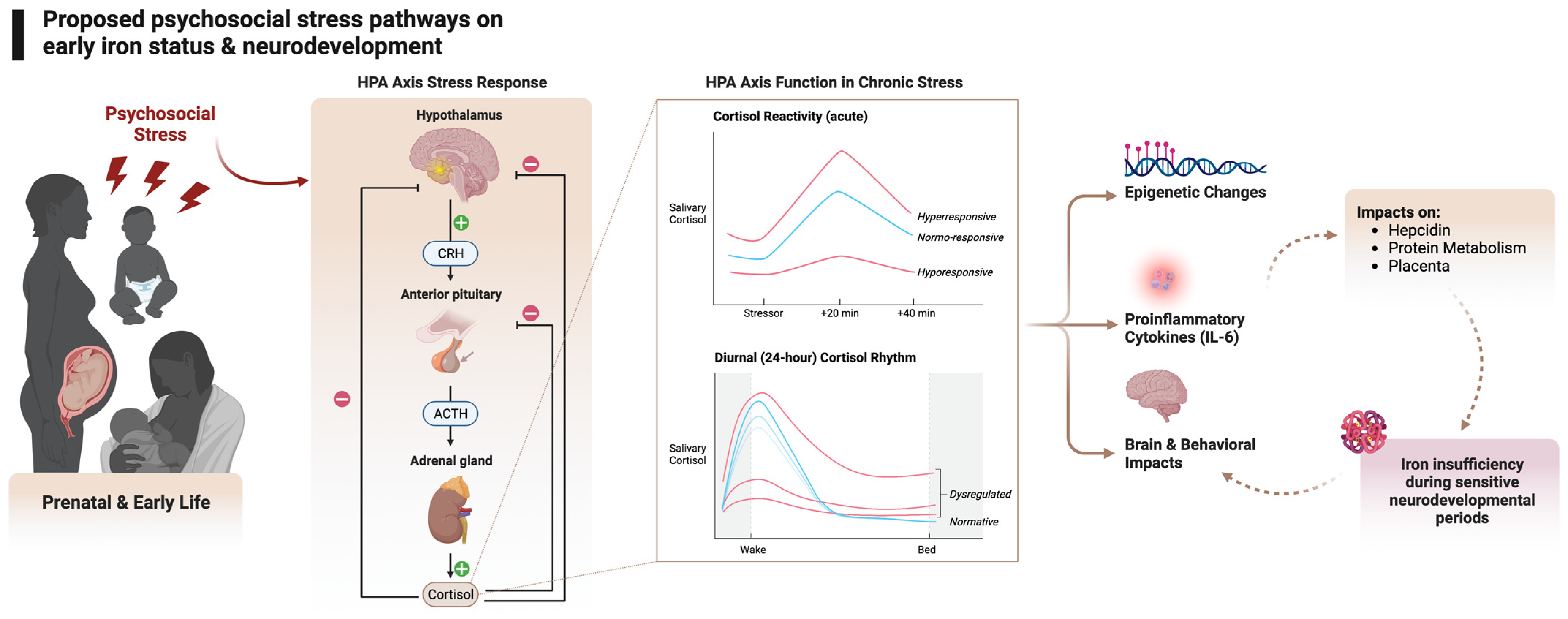
The hypothalamic–pituitary–adrenal (HPA) axis is a complex physiological system that plays a crucial role in our response to stress. The HPA axis involves multiple systems, including the autonomic, neuroendocrine, metabolic, and immune systems, which work in tandem to help manage stress and maintain homeostasis. One of the key functions of the HPA axis is the release of glucocorticoids (i.e., cortisol) in response to potential threats or danger [88]. The ability to mount a stress response and return to baseline is essential for survival: the mature HPA axis ideally responds to stress with a peak in cortisol and a swift return to baseline [89]. The HPA axis also exhibits a diurnal rhythm, with a rise in cortisol levels at waking and a peak 30–45 min after waking, followed by a decline across the day and a nadir in cortisol levels around bedtime [90,91,92,93]. Maintaining this diurnal rhythm is essential for mental and physical health, and chronic stress has been found to disturb the diurnal cortisol rhythm across multiple populations [90].
Current research suggests that experiencing moderate stress levels can benefit our overall functioning [94]. However, prolonged exposure to high stress levels can negatively impact physical and mental health [95,96,97]. The relationship between cortisol and optimal functioning follows an inverted U-shaped curve: cortisol that is too low and too high either in response to a stressor or in the diurnal rhythm tends to impair functioning [10]. Extended periods of low cortisol levels can impair restorative functions if the mineralocorticoid receptors are not fully occupied. On the other hand, extended periods of high cortisol levels can result in an overabundance of stress- or energy-depleting functions mediated by mineralocorticoid receptors due to a shift in the ratio of glucocorticoid receptor to mineralocorticoid receptor occupation (reviewed in [89]).
How chronic stress impacts HPA axis functioning depends on which stage of development the organism experiences the stress in (for review, see [98]). In pregnancy, severe acute or chronic stress can impact the mother’s HPA axis and the offspring’s developing brain and HPA axis [99] (for review, see [86,100]). Some maternal cortisol does reach the fetal compartment, and maternal stress may decrease placental 11β-HSD-2 activity, allowing more maternal cortisol to enter fetal circulation as it crosses the placenta ([101], for review, see [102]). At birth, humans have a functioning HPA axis that exhibits an immature diurnal rhythm and an HPA axis that can react to environmental changes, arousal levels, and distress [103,104]. This axis continues to mature across development. In this way, chronic or severe stress exposure during the prenatal or postnatal period can impact the HPA axis and subsequently influence iron metabolism. Preclinical work has found a connection between stress hormones and iron metabolism in the brain: in vitro corticosterone application dysregulates iron metabolism in hippocampal neurons [105]. To our knowledge, the only study in humans that has examined this connection provides preliminary evidence that maternal HPA axis regulation in response to an acute stressor is associated with iron status in the third trimester of pregnancy [106]. We hypothesize that the primary actor in this cascade involves inflammatory mechanisms, which we will review in the next section.
4.2. Stress and Inflammation
Decades of research have established connections between stress and immune function, including inflammation ([107], for reviews, see [8,108,109]). There are multiple pathways by which stress-mediating systems communicate with the peripheral immune system and can induce a proinflammatory state that can influence iron status. These pathways include the HPA axis, the sympathetic nervous system (SNS), the vagus nerve, and meningeal lymphatic vessels. Psychological stress triggers an inflammatory response centrally and peripherally (e.g., [110,111]). Increases in levels of circulating proinflammatory cytokines and expression of proinflammatory genes typically follow exposure to an acute stressor (minutes to hours) [112,113]. Similar to the increase in innate immune activity and proinflammatory cytokines during infectious stress, this increase in a proinflammatory profile is considered adaptive in the short term [108] but can be maladaptive—especially for iron status—in the long term.
Chronic stress is also associated with increased inflammation, decreases in antiviral immunity, changes in adaptive immunity [107,109], and increased expression of inflammation-related genes, which have been hypothesized to shift an organism into a proinflammatory phenotype [114]. Early life stress increases the signaling and upregulation of genes associated with inflammation in primates, children, and adults [115,116,117,118]. Stress exposure during childhood is also associated with increased circulating markers of inflammation in pediatric populations [119] and adult populations with a history of childhood stress exposure [120]. Adolescents who experienced higher levels of childhood stress have exhibited increased inflammatory gene expression [121], which aligns with similar findings found in adults (as reviewed in [108]). Stress early in life may result in an epigenetic modification in the GR gene responsible for regulating the body’s inflammatory response, most notably in the neural transcriptome of the hypothalamus and amygdala [122]. Additionally, stress can epigenetically reprogram immune cells and impact their production from the bone marrow, resulting in an imbalance in the body’s immune response [123,124]. In pregnant individuals, studies of mothers with high levels of psychological stress and low social support find associations between stress and elevated proinflammatory cytokines levels across gestation [86,125]. How stress impacts the developing immune system and leads to a proinflammatory phenotype in human neonates and infants is still an active area of research [126]. Nevertheless, the evidence suggests that non-infectious psychological stress may operate along similar pathways of infection and inflammation [127], leading to the risk of IDA even in the context of adequate iron intake.
4.3. Stress, Inflammation, and Hepcidin
Hepcidin may be the key factor in the relationship between psychological stress, inflammation, and iron status. Hepcidin, its receptor, and iron transporter ferroportin work in concert to control iron’s dietary absorption, storage, and tissue distribution. Hepcidin is upregulated in response to inflammatory states to decrease iron availability and control infection. Hepcidin and ferroportin expression are modulated during infection and inflammation—and potentially psychological stress—to reduce iron availability. Iron supply for red blood cell precursors is also restricted, contributing to the anemia associated with infections and inflammatory conditions [128]. Hepcidin signaling causes functional iron deficiency (redistribution of iron into the reticuloendothelial system, away from the red cells) on top of iron deficiency from other causes, such as inadequate dietary iron, or pregnancy and early infancy’s physiologic demands for iron. In chronic stress, the hepcidin-mediated reduction in gut absorption may also worsen total body iron deficiency. IL-6 is a key proinflammatory cytokine that rises after an acute stressor, is found in studies of chronic stress [108,112], and is concurrently the cytokine required for inducing hepcidin during inflammation [129]. Thus, increased hepcidin activity would increase the risk of iron deficiency when increased inflammation is due to psychological stress. Preliminary evidence supports the stress-inflammation-hepcidin pathway. In a study of adult rats, psychological stress from a communication box paradigm induced hypoferremia through the IL-6–hepcidin axis [27]. Psychological stress increased IL-6 and hepcidin expression, and the changes were reversed by IL-6 monoclonal antibody injection [27]. In humans, a study of 370 adolescent boys from Hyderabad, India, found that higher levels of self-reported life event stressors were associated with elevated IL-6 and hepcidin concentration [130]. To our knowledge, no studies have investigated this link in pregnant or infant populations.
4.4. Other Mechanisms
Prenatally, the placenta plays a crucial role in regulating fetal development and the intrauterine environment. Maternal stress can impact the placental methylome, which may affect fetal outcomes and increase the risk of fetal iron-deficiency anemia [131,132]. However, the placenta’s role in fetal IDA risk in the context of maternal stress and maternal IDA remains understudied. There could be a connection between psychological stress, iron levels, mitochondrial functioning, and neurodevelopment. In non-pregnancy data, both stress and iron deficiency can affect the functioning of mitochondria, which are responsible for regulating and signaling metabolism in cells and have been implicated in developing psychopathology [133]. Iron is essential for neuronal energy metabolism [134]. For instance, early-life iron deficiency can disrupt the size, motility, and energy capacity of mitochondria in developing hippocampal neurons [135], as well as impair mitochondrial energetics and the transcriptional regulation of mitochondrial quality control genes in adult animals that were previously iron-deficient [136]. In a rodent model of prenatal stress, male offspring displayed depressive-like symptoms associated with a reduction in PGC-1α protein, a regulator of mitochondrial biogenesis, in the frontal cortex and hippocampus [137]. A human study also showed that prenatal stress can affect placental mitochondrial DNA gene expression, and increased expression of MT-ND2 was subsequently linked to infant temperament [138]. Furthermore, a recent study in mice found that chronic social stress can disrupt iron metabolism and enhance hepatic mitochondrial function and ATP production [139].
Another possible connection between psychological stress and iron status involves the disruption of protein metabolism. In cases of infectious stress, steroids and proinflammatory cytokines can disrupt protein metabolism, causing the body to go into a catabolic state to produce fuel for fight-or-flight responses. This means that proteins are broken down for gluconeogenesis instead of being used for building tissue, including the brain’s structural proteins like dendrites and synapses, as well as other soluble proteins like neurotransmitters and growth factors (which can be affected by stress/sepsis and reduce IGF-1 synthesis). Additionally, proteins are necessary to transport iron around the body (as hemoglobin or cytochromes to generate ATP). Linear growth stunting is one illustrative example of how stress and inflammation can suppress protein synthesis. Postnatal linear growth stunting in infants and toddlers is a likely somatic instantiation of inflammation (e.g., [140]). In a parallel population, in preterm infants in the NICU, stunting is due to a lack of nutrition (protein-energy) and the amount of steroid, antibiotic, and inflammation exposure [141]. Much of these effects are mediated through effects on mammalian Target of Rapamycin (mTOR) pathway signaling [142], which integrates nutrient availability and growth factor status to regulate neuronal growth and differentiation. Therefore, the connection between psychological stress and iron may also be linked to stress and inflammation’s effects on protein. Additional pathways between stress and iron metabolism are likely and warrant further research.
5. Conclusions and Future Directions
Chronic psychological stress and HPA axis dysregulation in pregnant and non-pregnant individuals are associated with increased levels of circulating proinflammatory cytokines IL-6 and CRP. IL-6 directly regulates hepcidin synthesis. The neurodevelopmental consequences of early stress and iron deficiency may reflect mirrored impacts or interactions between the neuroendocrine response to stress and iron metabolism. Therefore, a future area of research that needs attention is the dysregulated neuroendocrine pathways that arise from psychological stress that may disrupt iron absorption and utilization even in the context of adequate intake [143,144].
Several outstanding questions will need to be addressed in future research. In studies of adult rodents, it is unclear why and how stress-related imbalances in iron homeostasis are linked to the type, duration, and severity of stressors. Prenatally, psychological stress in monkeys early in pregnancy (second trimester) affected iron status in infant offspring [24], and dietary intervention with moderate iron supplementation during pregnancy did not prevent the offspring from developing iron deficiency [145]. The second trimester is also highlighted in the association between prenatal iron deficiency and the risk of offspring schizophrenia in epidemiological work in humans [49]. Future research must address the timing, type, and duration of psychological stress and the subsequent impacts on iron metabolism.
Furthermore, despite suggestive connections between psychological stress and iron status in preclinical animal work (e.g., [24]) and cohorts of pregnant women [21,22,23], no study has examined how hypothalamic–pituitary–adrenocortical (HPA) axis dysregulation arising from chronic psychological stress affects maternal risk for iron deficiency and offspring iron status. In studies of pregnant rhesus macaques, the magnitude of the maternal cortisol response over pregnancy was not associated with offspring iron status [24]. Though one preliminary study in humans found evidence for associations between maternal cortisol response to an acute stressor and iron status in the third trimester [106], the precise relationship between the HPA axis and iron regulation in pregnancy and infancy remains an open area of research. There are also still questions about whether psychological stress causes an increase in proinflammatory cytokines that leads to changes in hepcidin and protein regulation, which affects iron status. Additionally, it is unclear if there is a threshold for the relationship between HPA axis regulation, inflammation, and iron metabolism changes or if it is a continuous relationship, and if so, what the biomarker for that threshold is. Further research is needed to fully understand the complicated relationship between psychological stress, inflammation, and iron metabolism.
Evaluating maternal chronic stress as a novel treatment mechanism could prevent maternal and offspring IDA and subsequent health problems. This line of research has the potential to change the clinical infrastructure required to solve prenatal maternal iron deficiency and risk to offspring in high-stress populations, requiring a different allocation of resources. Studying the effects of maternal psychological stress, neuroendocrine regulation, and inflammatory biomarkers on iron status could help identify novel intervention targets focused on psychological stress for preventing IDA and subsequent neurodevelopmental sequelae. Overall, further research on the relationship between maternal chronic stress and iron deficiency has the potential to benefit the health and well-being of both pregnant people and their children.
Author Contributions
Conceptualization, B.M.R. and M.K.G.; writing—original draft preparation, B.M.R.; writing—review and editing, B.M.R. and M.K.G.; visualization, B.M.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the US National Institutes of Health (NIH) and Eunice Kennedy Shriver National Institute of Child Health and Human Development (NICHD), grant K99HD109373 to BMR and grant R01HD094809 to MKG. This work is also supported by the Canadian Institute for Advanced Research (CIFAR) program in Child and Brain Development (to BMR). The listed authors are solely responsible for this paper’s content; the conclusions do not reflect the official view of the NIH or CIFAR.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Our appreciation to Megan Gunnar and Sarah Cusick for fruitful discussions. We apologize to those of our colleagues whose papers could not be cited in this review owing to space limitations.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Cusick, S.E.; Georgieff, M.K. The Role of Nutrition in Brain Development: The Golden Opportunity of the “First 1000 Days”. J. Pediatr. 2016, 175, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Iron assessment to protect the developing brain. Am. J. Clin. Nutr. 2017, 106, 1588S–1593S. [Google Scholar] [CrossRef]
- Stevens, G.A.; Finucane, M.M.; De-Regil, L.M.; Paciorek, C.J.; Flaxman, S.R.; Branca, F.; Pena-Rosas, J.P.; Bhutta, Z.A.; Ezzati, M.; Nutrition Impact Model Study Group. Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: A systematic analysis of population-representative data. Lancet Glob. Health 2013, 1, e16–e25. [Google Scholar] [CrossRef]
- Cusick, S.; Georgieff, M.; Rao, R. Approaches for Reducing the Risk of Early-Life Iron Deficiency-Induced Brain Dysfunction in Children. Nutrients 2018, 10, 227. [Google Scholar] [CrossRef]
- Georgieff, M.K. Iron deficiency in pregnancy. Am. J. Obstet. Gynecol. 2020, 223, 516–524. [Google Scholar] [CrossRef]
- Lopresti, A.L. The Effects of Psychological and Environmental Stress on Micronutrient Concentrations in the Body: A Review of the Evidence. Adv. Nutr. 2020, 11, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Hantsoo, L.; Kornfield, S.; Anguera, M.C.; Epperson, C.N. Inflammation: A Proposed Intermediary Between Maternal Stress and Offspring Neuropsychiatric Risk. Biol. Psychiatry 2019, 85, 97–106. [Google Scholar] [CrossRef]
- Slavich, G.M. Social Safety Theory: A Biologically Based Evolutionary Perspective on Life Stress, Health, and Behavior. Annu. Rev. Clin. Psychol. 2020, 16, 265–295. [Google Scholar] [CrossRef]
- McEwen, B.S. Allostasis and allostatic load: Implications for neuropsychopharmacology. Neuropsychopharmacology 2000, 22, 108–124. [Google Scholar] [CrossRef]
- Lupien, S.J.; McEwen, B.S.; Gunnar, M.R.; Heim, C. Effects of stress throughout the lifespan on the brain, behaviour and cognition. Nat. Rev. Neurosci. 2009, 10, 434–445. [Google Scholar] [CrossRef]
- Herrick, H.W.B. The Effect of Stressful Life Events on Postpartum Depression: Results from the 1997–1998 North Carolina Pregnancy Risk Assessment Monitoring System (PRAMS); North Carolina State Center for Health Statistics: Raleigh, NC, USA, 2000. [Google Scholar]
- Whitehead, N.; Hill, H.A.; Brogan, D.J.; Blackmore-Prince, C. Exploration of Threshold Analysis in the Relation between Stressful Life Events and Preterm Delivery. Am. J. Epidemiol. 2002, 155, 117–124. [Google Scholar] [CrossRef]
- Lu, M.C.; Chen, B. Racial and ethnic disparities in preterm birth: The role of stressful life events. Am. J. Obstet. Gynecol. 2004, 191, 691–699. [Google Scholar] [CrossRef]
- World Health Organization. Global Status Report on Preventing Violence against Children; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- U.S. Department of Health & Human Services; Administration on Children, Youth and Families; Children’s Bureau. Child Maltreatment 2021; U.S. Department of Health & Human Services: Washington, DC, USA, 2021. [Google Scholar]
- United Nations Children’s Fund. A Familiar Face: Violence in the Lives of Children and Adolescents; UNICEF: New York, NY, USA, 2017. [Google Scholar]
- Brannon, P.M.; Taylor, C.L. Iron Supplementation during Pregnancy and Infancy: Uncertainties and Implications for Research and Policy. Nutrients 2017, 9, 1327. [Google Scholar] [CrossRef]
- Rahman, M.M.; Abe, S.K.; Rahman, M.S.; Kanda, M.; Narita, S.; Bilano, V.; Ota, E.; Gilmour, S.; Shibuya, K. Maternal anemia and risk of adverse birth and health outcomes in low- and middle-income countries: Systematic review and meta-analysis1,2. Am. J. Clin. Nutr. 2016, 103, 495–504. [Google Scholar] [CrossRef] [PubMed]
- Black, M.M.; Quigg, A.M.; Hurley, K.M.; Pepper, M.R. Iron deficiency and iron-deficiency anemia in the first two years of life: Strategies to prevent loss of developmental potential. Nutr. Rev. 2011, 69 (Suppl. S1), S64–S70. [Google Scholar] [CrossRef] [PubMed]
- Black, R.E.; Allen, L.H.; Bhutta, Z.A.; Caulfield, L.E.; de Onis, M.; Ezzati, M.; Mathers, C.; Rivera, J.; Maternal and Child Undernutrition Study Group. Maternal and child undernutrition: Global and regional exposures and health consequences. Lancet 2008, 371, 243–260. [Google Scholar] [CrossRef]
- Armony-Sivan, R.; Aviner, S.; Cojocaru, L.; Fytlovitch, S.; Ben-Alon, D.; Eliassy, A.; Babkoff, H.; Lozoff, B.; Anteby, E. Prenatal maternal stress predicts cord-blood ferritin concentration. J. Perinat. Med. 2013, 41, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.K.; Tamayo-Ortiz, M.; Cantoral, A.; Schnaas, L.; Osorio-Valencia, E.; Wright, R.J.; Tellez-Rojo, M.M.; Wright, R.O. Maternal Prenatal Psychological Stress and Prepregnancy BMI Associations with Fetal Iron Status. Curr. Dev. Nutr. 2020, 4, nzaa018. [Google Scholar] [CrossRef]
- Rendina, D.N.; Blohowiak, S.E.; Coe, C.L.; Kling, P.J. Maternal Perceived Stress during Pregnancy Increases Risk for Low Neonatal Iron at Delivery and Depletion of Storage Iron at One Year. J. Pediatr. 2018, 200, 166–173.e2. [Google Scholar] [CrossRef]
- Coe, C.L.; Lubach, G.R.; Shirtcliff, E.A. Maternal stress during pregnancy predisposes for iron deficiency in infant monkeys impacting innate immunity. Pediatr. Res. 2007, 61, 520–524. [Google Scholar] [CrossRef]
- Teng, W.F.; Sun, W.M.; Shi, L.F.; Hou, D.D.; Liu, H. Effects of restraint stress on iron, zinc, calcium, and magnesium whole blood levels in mice. Biol. Trace Elem. Res. 2008, 121, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Zhou, J.; Huang, X.; Li, M. Effects of psychological stress on serum iron and erythropoiesis. Int. J. Hematol. 2008, 88, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Chen, J.B.; Wang, W.Y.; Wang, L.; Ma, L.; Shen, H.; Li, M. Psychological stress induces hypoferremia through the IL-6-hepcidin axis in rats. Biochem. Biophys. Res. Commun. 2008, 373, 90–93. [Google Scholar] [CrossRef]
- Li, Y.J.; Zheng, Y.Y.; Qian, J.X.; Chen, X.M.; Shen, Z.L.; Tao, L.P.; Li, H.X.; Qin, H.H.; Li, M.; Shen, H. Preventive Effects of Zinc Against Psychological Stress-Induced Iron Dyshomeostasis, Erythropoiesis Inhibition, and Oxidative Stress Status in Rats. Biol. Trace Elem. Res. 2012, 147, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.B.; Shen, H.; Chen, C.J.; Wang, W.Y.; Yu, S.Y.; Zhao, M.; Li, M. The effect of psychological stress on iron absorption in rats. BMC Gastroenterol. 2009, 9, 83. [Google Scholar] [CrossRef]
- Li, H.F.; Jiang, S.X.; Yang, C.; Yang, S.; He, B.; Ma, W.Q.; Zhao, R.Q. Long-Term Dexamethasone Exposure Down-Regulates Hepatic TFR1 and Reduces Liver Iron Concentration in Rats. Nutrients 2017, 9, 617. [Google Scholar] [CrossRef]
- Guo, S.H.; Yang, C.; Jiang, S.X.; Ni, Y.D.; Zhao, R.Q.; Ma, W.Q. Repeated Restraint Stress Enhances Hepatic TFR2 Expression and Induces Hepatic Iron Accumulation in Rats. Biol. Trace Elem. Res. 2020, 196, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Smoak, B.L.; Patterson, K.Y.; LeMay, L.G.; Veillon, C.; Deuster, P.A. Biochemical indices of selected trace minerals in men: Effect of stress. Am. J. Clin. Nutr. 1991, 53, 126–131. [Google Scholar] [CrossRef]
- Reid, B.M.; East, P.; Blanco, E.; Doom, J.R.; Burrows, R.A.; Correa-Burrows, P.; Lozoff, B.; Gahagan, S. Early-life adversity is associated with poor iron status in infancy. Dev. Psychopathol. 2022, 1–12. [Google Scholar] [CrossRef]
- Lozoff, B.; Georgieff, M.K. Iron deficiency and brain development. Semin. Pediatr. Neurol. 2006, 13, 158–165. [Google Scholar] [CrossRef]
- Beard, J.L.; Connor, J.R. Iron status and neural functioning. Annu. Rev. Nutr. 2003, 23, 41–58. [Google Scholar] [CrossRef]
- Beard, J.L.; Wiesinger, J.A.; Connor, J.R. Pre- and postweaning iron deficiency alters myelination in Sprague-Dawley rats. Dev. Neurosci. 2003, 25, 308–315. [Google Scholar] [CrossRef] [PubMed]
- Kwik-Uribe, C.L.; Gietzen, D.; German, J.B.; Golub, M.S.; Keen, C.L. Chronic marginal iron intakes during early development in mice result in persistent changes in dopamine metabolism and myelin composition. J. Nutr. 2000, 130, 2821–2830. [Google Scholar] [CrossRef] [PubMed]
- Oloyede, O.B.; Folayan, A.T.; Odutuga, A.A. Effects of Low-Iron Status and Deficiency of Essential Fatty-Acids on Some Biochemical-Constituents of Rat-Brain. Biochem. Int. 1992, 27, 913–922. [Google Scholar] [PubMed]
- Yu, G.S.; Steinkirchner, T.M.; Rao, G.A.; Larkin, E.C. Effect of prenatal iron deficiency on myelination in rat pups. Am. J. Pathol. 1986, 125, 620–624. [Google Scholar]
- Lozoff, B.; Beard, J.; Connor, J.; Felt, B.; Georgieff, M.; Schallert, T. Long-Lasting Neural and Behavioral Effects of Iron Deficiency in Infancy. Nutr. Rev. 2006, 64 (Suppl. S2), S34–S43. [Google Scholar] [CrossRef] [PubMed]
- de Deungria, M.; Rao, R.; Wobken, J.D.; Luciana, M.; Nelson, C.A.; Georgieff, M.K. Perinatal iron deficiency decreases cytochrome c oxidase (CytOx) activity in selected regions of neonatal rat brain. Pediatr. Res. 2000, 48, 169–176. [Google Scholar] [CrossRef]
- Rao, R.; Tkac, I.; Townsend, E.L.; Gruetter, R.; Georgieff, M.K. Perinatal iron deficiency alters the neurochemical profile of the developing rat hippocampus. J. Nutr. 2003, 133, 3215–3221. [Google Scholar]
- Jorgenson, L.A.; Wobken, J.D.; Georgieff, M.K. Perinatal iron deficiency alters apical dendritic growth in hippocampal CA1 pyramidal neurons. Dev. Neurosci. 2003, 25, 412–420. [Google Scholar] [CrossRef]
- Siddappa, A.M.; Georgieff, M.K.; Wewerka, S.; Worwa, C.; Nelson, C.A.; DeRegnier, R.A. Iron deficiency alters auditory recognition memory in newborn infants of diabetic mothers. Pediatr. Res. 2004, 55, 1034–1041. [Google Scholar] [CrossRef]
- Geng, F.; Mai, X.; Zhan, J.; Xu, L.; Zhao, Z.; Georgieff, M.; Shao, J.; Lozoff, B. Impact of Fetal-Neonatal Iron Deficiency on Recognition Memory at 2 Months of Age. J. Pediatr. 2015, 167, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Amin, S.B.; Orlando, M.; Eddins, A.; MacDonald, M.; Monczynski, C.; Wang, H. In Utero Iron Status and Auditory Neural Maturation in Premature Infants as Evaluated by Auditory Brainstem Response. J. Pediatr. 2010, 156, 377–381. [Google Scholar] [CrossRef]
- Wachs, T.D.; Pollitt, E.; Cueto, S.; Jacoby, E. Relation of neonatal iron status to individual variability in neonatal temperament. Dev. Psychobiol. 2005, 46, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, R.J.; Tancredi, D.J.; Krakowiak, P.; Hansen, R.L.; Ozonoff, S. Maternal Intake of Supplemental Iron and Risk of Autism Spectrum Disorder. Am. J. Epidemiol. 2014, 180, 890–900. [Google Scholar] [CrossRef] [PubMed]
- Insel, B.J.; Schaefer, C.A.; McKeague, I.W.; Susser, E.S.; Brown, A.S. Maternal iron deficiency and the risk of schizophrenia in offspring. Arch. Gen. Psychiatry 2008, 65, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- Riggins, T.; Miller, N.C.; Bauer, P.J.; Georgieff, M.K.; Nelson, C.A. Consequences of Low Neonatal Iron Status Due to Maternal Diabetes Mellitus on Explicit Memory Performance in Childhood. Dev. Neuropsychol. 2009, 34, 762–779. [Google Scholar] [CrossRef] [PubMed]
- Jabès, A.; Thomas, K.M.; Langworthy, S.; Georgieff, M.K.; Nelson, C.A. Functional and Anatomic Consequences of Diabetic Pregnancy on Memory in Ten-Year-Old Children. J. Dev. Behav. Pediatr. 2015, 36, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Lozoff, B.; Clark, K.M.; Jing, Y.; Armony-Sivan, R.; Angelilli, M.L.; Jacobson, S.W. Dose-response relationships between iron deficiency with or without anemia and infant social-emotional behavior. J. Pediatr. 2008, 152, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Lukowski, A.F.; Koss, M.; Burden, M.J.; Jonides, J.; Nelson, C.A.; Kaciroti, N.; Jimenez, E.; Lozoff, B. Iron deficiency in infancy and neurocognitive functioning at 19 years: Evidence of long-term deficits in executive function and recognition memory. Nutr. Neurosci. 2010, 13, 54–70. [Google Scholar] [CrossRef] [PubMed]
- Jorgenson, L.A.; Sun, M.; O’Connor, M.; Georgieff, M.K. Fetal iron deficiency disrupts the maturation of synaptic function and efficacy in area CA1 of the developing rat hippocampus. Hippocampus 2005, 15, 1094–1102. [Google Scholar] [CrossRef]
- Vallée, M.; Maccari, S.; Dellu, F.; Simon, H.; Le Moal, M.; Mayo, W. Long-term effects of prenatal stress and postnatal handling on age-related glucocorticoid secretion and cognitive performance: A longitudinal study in the rat. Eur. J. Neurosci. 1999, 11, 2906–2916. [Google Scholar] [CrossRef] [PubMed]
- Vallée, M.; Mayo, W.; Dellu, F.; Le Moal, M.; Simon, H.; Maccari, S. Prenatal Stress Induces High Anxiety and Postnatal Handling Induces Low Anxiety in Adult Offspring: Correlation with Stress-Induced Corticosterone Secretion. J. Neurosci. 1997, 17, 2626–2636. [Google Scholar] [CrossRef] [PubMed]
- Piazza, P.V.; Le Moal, M. Pathophysiological Basis of Vulnerability to Drug Abuse: Role of an Interaction Between Stress, Glucocorticoids, and Dopaminergic Neurons. Annu. Rev. Pharmacol. Toxicol. 1996, 36, 359–378. [Google Scholar] [CrossRef] [PubMed]
- Lemaire, V.; Koehl, M.; Le Moal, M.; Abrous, D.N. Prenatal stress produces learning deficits associated with an inhibition of neurogenesis in the hippocampus. Proc. Natl. Acad. Sci. USA 2000, 97, 11032–11037. [Google Scholar] [CrossRef]
- Sandman, C.A.; Curran, M.M.; Davis, E.P.; Glynn, L.M.; Head, K.; Baram, T.Z. Cortical Thinning and Neuropsychiatric Outcomes in Children Exposed to Prenatal Adversity: A Role for Placental CRH? Am. J. Psychiatry 2018, 175, 471–479. [Google Scholar] [CrossRef]
- Swales, D.A.; Stout-Oswald, S.A.; Glynn, L.M.; Sandman, C.; Wing, D.A.; Davis, E.P. Exposure to traumatic events in childhood predicts cortisol production among high risk pregnant women. Biol. Psychol. 2018, 139, 186–192. [Google Scholar] [CrossRef]
- Chan, J.C.; Nugent, B.M.; Bale, T.L. Parental Advisory: Maternal and Paternal Stress Can Impact Offspring Neurodevelopment. Biol. Psychiatry 2018, 83, 886–894. [Google Scholar] [CrossRef]
- O’Donnell, K.J.; Glover, V.; Barker, E.D.; O’Connor, T.G. The persisting effect of maternal mood in pregnancy on childhood psychopathology. Dev. Psychopathol. 2014, 26, 393–403. [Google Scholar] [CrossRef]
- Van den Bergh, B.R.H.; van den Heuvel, M.I.; Lahti, M.; Braeken, M.; de Rooij, S.R.; Entringer, S.; Hoyer, D.; Roseboom, T.; Raikkonen, K.; King, S.; et al. Prenatal developmental origins of behavior and mental health: The influence of maternal stress in pregnancy. Neurosci. Biobehav. Rev. 2020, 117, 26–64. [Google Scholar] [CrossRef]
- Gunnar, M.; Reid, B.M. Early Deprivation Revisited: Contemporary Studies of the Impact on Young Children of Institutional Care. Annu. Rev. Dev. Psychol. 2019, 1, 93–118. [Google Scholar]
- Pollak, S.D.; Nelson, C.A.; Schlaak, M.; Roeber, B.; Wewerka, S.; Wiik, K.L.; Frenn, K.; Loman, M.M.; Gunnar, M.R. Neurodevelopmental effects of early deprivation in post-institutionalized children. Child Dev. 2010, 81, 224–236. [Google Scholar] [PubMed]
- Roy, P.; Rutter, M.; Pickles, A. Institutional care: Associations between overactivity and lack of selectivity in social relationships. J. Psychol. Psychiatry 2004, 45, 866–873. [Google Scholar]
- Burgess, P.W.; Stuss, D.T. Fifty Years of Prefrontal Cortex Research: Impact on Assessment. J. Int. Neuropsychol. Soc. 2017, 23, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Smith, K.E.; Pollak, S.D. Early life stress and neural development: Implications for understanding the developmental effects of COVID-19. Cogn. Affect. Behav. Neurosci. 2022, 22, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Turecki, G.; Meaney, M.J. Effects of the Social Environment and Stress on Glucocorticoid Receptor Gene Methylation: A Systematic Review. Biol. Psychiatry 2016, 79, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Brunson, K.L.; Kramar, E.; Lin, B.; Chen, Y.; Colgin, L.L.; Yanagihara, T.K.; Lynch, G.; Baram, T.Z. Mechanisms of late-onset cognitive decline after early-life stress. J. Neurosci. 2005, 25, 9328–9338. [Google Scholar] [CrossRef] [PubMed]
- Danielewicz, J.; Hess, G. Early life stress alters synaptic modification range in the rat lateral amygdala. Behav. Brain Res. 2014, 265, 32–37. [Google Scholar] [CrossRef]
- Ishikawa, J.; Nishimura, R.; Ishikawa, A. Early-life stress induces anxiety-like behaviors and activity imbalances in the medial prefrontal cortex and amygdala in adult rats. Eur. J. Neurosci. 2015, 41, 442–453. [Google Scholar] [CrossRef]
- Wei, L.; David, A.; Duman, R.S.; Anisman, H.; Kaffman, A. Early life stress increases anxiety-like behavior in Balbc mice despite a compensatory increase in levels of postnatal maternal care. Horm. Behav. 2010, 57, 396–404. [Google Scholar] [CrossRef]
- Bolton, J.L.; Molet, J.; Regev, L.; Chen, Y.C.; Rismanchi, N.; Haddad, E.; Yang, D.Z.; Obenaus, A.; Baram, T.Z. Anhedonia Following Early-Life Adversity Involves Aberrant Interaction of Reward and Anxiety Circuits and Is Reversed by Partial Silencing of Amygdala Corticotropin-Releasing Hormone Gene. Biol. Psychiatry 2018, 83, 137–147. [Google Scholar] [CrossRef]
- Berman, A.K.; Lott, R.B.; Donaldson, S.T. Periodic maternal deprivation may modulate offspring anxiety-like behavior through mechanisms involving neuroplasticity in the amygdala. Brain Res. Bull. 2014, 101, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Monroy, E.; Hernandez-Torres, E.; Flores, G. Maternal separation disrupts dendritic morphology of neurons in prefrontal cortex, hippocampus, and nucleus accumbens in male rat offspring. J. Chem. Neuroanat. 2010, 40, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Bagot, R.C.; van Hasselt, F.N.; Champagne, D.L.; Meaney, M.J.; Krugers, H.J.; Joels, M. Maternal care determines rapid effects of stress mediators on synaptic plasticity in adult rat hippocampal dentate gyrus. Neurobiol. Learn. Mem. 2009, 92, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Ivy, A.S.; Rex, C.S.; Chen, Y.C.; Dube, C.; Maras, P.M.; Grigoriadis, D.E.; Gall, C.M.; Lynch, G.; Baram, T.Z. Hippocampal Dysfunction and Cognitive Impairments Provoked by Chronic Early-Life Stress Involve Excessive Activation of CRH Receptors. J. Neurosci. 2010, 30, 13005–13015. [Google Scholar] [CrossRef]
- Oomen, C.A.; Soeters, H.; Audureau, N.; Vermunt, L.; van Hasselt, F.N.; Manders, E.M.M.; Joels, M.; Lucassen, P.J.; Krugers, H. Severe Early Life Stress Hampers Spatial Learning and Neurogenesis, but Improves Hippocampal Synaptic Plasticity and Emotional Learning under High-Stress Conditions in Adulthood. J. Neurosci. 2010, 30, 6635–6645. [Google Scholar] [CrossRef]
- Swales, D.A.; Winiarski, D.A.; Smith, A.K.; Stowe, Z.N.; Newport, D.J.; Brennan, P.A. Maternal depression and cortisol in pregnancy predict offspring emotional reactivity in the preschool period. Dev. Psychobiol. 2018, 60, 557–566. [Google Scholar] [CrossRef]
- Van den Bergh, B.R.; Van Calster, B.; Smits, T.; Van Huffel, S.; Lagae, L. Antenatal maternal anxiety is related to HPA-axis dysregulation and self-reported depressive symptoms in adolescence: A prospective study on the fetal origins of depressed mood. Neuropsychopharmacol. 2008, 33, 536–545. [Google Scholar] [CrossRef]
- Grantham-McGregor, S.; Ani, C. A review of studies on the effect of iron deficiency on cognitive development in children. J. Nutr. 2001, 131, 649S–666S. [Google Scholar]
- Skalicky, A.; Meyers, A.F.; Adams, W.G.; Yang, Z.; Cook, J.T.; Frank, D.A. Child food insecurity and iron deficiency anemia in low-income infants and toddlers in the United States. Matern. Child Health J. 2006, 10, 177–185. [Google Scholar] [CrossRef]
- Auerbach, M.; Abernathy, J.; Juul, S.; Short, V.; Derman, R. Prevalence of iron deficiency in first trimester, nonanemic pregnant women. J. Matern.-Fetal Neonatal Med. 2021, 34, 1002–1005. [Google Scholar] [CrossRef]
- Baskin, R.; Hill, B.; Jacka, F.N.; O’Neil, A.; Skouteris, H. The association between diet quality and mental health during the perinatal period. A systematic review. Appetite 2015, 91, 41–47. [Google Scholar] [CrossRef]
- Monk, C.; Lugo-Candelas, C.; Trumpff, C. Prenatal Developmental Origins of Future Psychopathology: Mechanisms and Pathways. Annu. Rev. Clin. Psychol. 2019, 15, 317–344. [Google Scholar] [CrossRef] [PubMed]
- Fontana, C. Adapted from “Hypothalamic-Pituitary-Adrenal Axis” by BioRender.com. Available online: https://app.biorender.com/biorender-templates (accessed on 21 August 2023).
- Herman, J.P.; Cullinan, W.E. Neurocircuitry of stress: Central control of the hypothalamo-pituitary-adrenocortical axis. Trends Neurosci. 1997, 20, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Engel, M.L.; Gunnar, M.R. The development of stress reactivity and regulation during human development. Int. Rev. Neurobiol. 2020, 150, 41–76. [Google Scholar] [PubMed]
- Adam, E.K.; Quinn, M.E.; Tavernier, R.; McQuillan, M.T.; Dahlke, K.A.; Gilbert, K.E. Diurnal cortisol slopes and mental and physical health outcomes: A systematic review and meta-analysis. Psychoneuroendocrinology 2017, 83, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Kirschbaum, C.; Hellhammer, D.H. Salivary cortisol in psychoneuroendocrine research: Recent developments and applications. Psychoneuroendocrinology 1994, 19, 313–333. [Google Scholar] [CrossRef]
- Levine, A.; Zagoory-Sharon, O.; Feldman, R.; Lewis, J.G.; Weller, A. Measuring cortisol in human psychobiological studies. Physiol. Behav. 2007, 90, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Miller, R.; Stalder, T.; Jarczok, M.; Almeida, D.M.; Badrick, E.; Bartels, M.; Boomsma, D.I.; Coe, C.L.; Dekker, M.C.; Donzella, B.; et al. The CIRCORT database: Reference ranges and seasonal changes in diurnal salivary cortisol derived from a meta-dataset comprised of 15 field studies. Psychoneuroendocrinology 2016, 73, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Lyons, D.M.; Parker, K.J. Stress inoculation-induced indications of resilience in monkeys. J. Trauma. Stress 2007, 20, 423–433. [Google Scholar] [CrossRef]
- Boyce, W.T.; Ellis, B.J. Biological sensitivity to context: I. An evolutionary-developmental theory of the origins and functions of stress reactivity. Dev. Psychopathol. 2005, 17, 271–301. [Google Scholar] [CrossRef]
- Lyons, D.M.; Parker, K.J.; Schatzberg, A.F. Animal models of early life stress: Implications for understanding resilience. Dev. Psychobiol. 2010, 52, 402–410. [Google Scholar] [CrossRef]
- Tronick, E. The Inherent Stress of Normal Daily Life and Social Interaction Leads to the Development of Coping and Resilience, and Variation in Resilience in Infants and Young Children: Comments on the Papers of Suomi and Klebanov & Brooks-Gunn. Ann. NY Acad. Sci. 2006, 1094, 83–104. [Google Scholar] [CrossRef]
- Leneman, K.B.; Gunnar, M.R. Developmental Timing of Stress Effects on the Brain. In The Oxford Handbook of Stress and Mental Health; Harkness, K.L., Hayden, E.P., Eds.; Oxford University Press: Oxford, UK, 2020. [Google Scholar] [CrossRef]
- Thayer, Z.M.; Wilson, M.A.; Kim, A.W.; Jaeggi, A.V. Impact of prenatal stress on offspring glucocorticoid levels: A phylogenetic meta-analysis across 14 vertebrate species. Sci. Rep. 2018, 8, 4942. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Davis, E.P. The effects of stress on early brain and behavioral development. In Cognitive Development; Rakic, P., Rubenstein, J., Tager-Flusberg, H.B., Eds.; Elsevier: New York, NY, USA, 2013; Volume 2, pp. 447–466. [Google Scholar]
- Gitau, R.; Cameron, A.; Fisk, N.M.; Glover, V. Fetal exposure to maternal cortisol. Lancet 1998, 352, 707–708. [Google Scholar] [CrossRef]
- Howland, M.A.; Sandman, C.A.; Glynn, L.M. Developmental origins of the human hypothalamic-pituitary-adrenal axis. Expert Rev. Endocrinol. Metab. 2017, 12, 321–339. [Google Scholar] [CrossRef]
- Gunnar, M.R.; Vazquez, D. Stress Neurobiology and Developmental Psychopathology. In Developmental Psychopathology: Vol. 2. Developmental Neuroscience; Cicchetti, D., Cohen, D., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; Volume 2. [Google Scholar]
- Gunnar, M.R. Reactivity of the hypothalamic-pituitary-adrenocortical system to stressors in normal infants and children. Pediatrics 1992, 90, 491–497. [Google Scholar]
- Wang, L.; Wang, H.; Li, L.; Li, W.; Dong, X.; Li, M.; Lv, L. Corticosterone induces dysregulation of iron metabolism in hippocampal neurons in vitro. Biol. Trace Elem. Res. 2010, 137, 88–95. [Google Scholar] [CrossRef]
- Reid, B.M.G.; Keenan, K. Association between Cortisol Stress Response and Iron Status in Pregnancy: Insights from a Study of Pregnant Black Women with Medicaid Insurance. In Proceedings of the International Society of Psychoneuroendocrinology Annual Conference, London, UK, 30 August–1 September 2023. [Google Scholar]
- Segerstrom, S.C.; Miller, G.E. Psychological stress and the human immune system: A meta-analytic study of 30 years of inquiry. Psychol. Bull. 2004, 130, 601–630. [Google Scholar] [CrossRef]
- Bower, J.E.; Kuhlman, K.R. Psychoneuroimmunology: An Introduction to Immune-to-Brain Communication and Its Implications for Clinical Psychology. Annu. Rev. Clin. Psychol. 2023, 19, 331–359. [Google Scholar] [CrossRef]
- Glaser, R.; Kiecolt-Glaser, J.K. Science and society—Stress-induced immune dysfunction: Implications for health. Nat. Rev. Immunol. 2005, 5, 243–251. [Google Scholar] [CrossRef]
- Weber, M.D.; Godbout, J.P.; Sheridan, J.F. Repeated Social Defeat, Neuroinflammation, and Behavior: Monocytes Carry the Signal. Neuropsychopharmacology 2017, 42, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Reader, B.F.; Jarrett, B.L.; McKim, D.B.; Wohleb, E.S.; Godbout, J.P.; Sheridan, J.F. Peripheral and central effects of repeated social defeat stress: Monocyte trafficking, microglial activation, and anxiety. Neuroscience 2015, 289, 429–442. [Google Scholar] [CrossRef] [PubMed]
- Marsland, A.L.; Walsh, C.; Lockwood, K.; John-Henderson, N.A. The effects of acute psychological stress on circulating and stimulated inflammatory markers: A systematic review and meta-analysis. Brain Behav. Immun. 2017, 64, 208–219. [Google Scholar] [CrossRef] [PubMed]
- MacCormack, J.K.; Gaudier-Diaz, M.M.; Armstrong-Carter, E.L.; Arevalo, J.M.G.; Meltzer-Brody, S.; Sloan, E.K.; Cole, S.W.; Muscatell, K.A. Beta-adrenergic blockade blunts inflammatory and antiviral/antibody gene expression responses to acute psychological stress. Neuropsychopharmacology 2021, 46, 756–762. [Google Scholar] [CrossRef]
- Cole, S.W. The Conserved Transcriptional Response to Adversity. Curr. Opin. Behav. Sci. 2019, 28, 31–37. [Google Scholar] [CrossRef]
- Cole, S.W.; Conti, G.; Arevalo, J.M.; Ruggiero, A.M.; Heckman, J.J.; Suomi, S.J. Transcriptional modulation of the developing immune system by early life social adversity. Proc. Natl. Acad. Sci. USA 2012, 109, 20578–20583. [Google Scholar] [CrossRef]
- Marie-Mitchell, A.; Cole, S.W. Adverse childhood experiences and transcriptional response in school-age children. Dev. Psychopathol. 2022, 34, 875–881. [Google Scholar] [CrossRef]
- Bower, J.E.; Kuhlman, K.R.; Ganz, P.A.; Irwin, M.R.; Crespi, C.M.; Cole, S.W. Childhood maltreatment and monocyte gene expression among women with breast cancer. Brain Behav. Immun. 2020, 88, 396–402. [Google Scholar] [CrossRef]
- Miller, G.E.; Chen, E.; Sze, J.; Marin, T.; Arevalo, J.M.; Doll, R.; Ma, R.; Cole, S.W. A functional genomic fingerprint of chronic stress in humans: Blunted glucocorticoid and increased NF-kappa B signaling. Biol. Psychiatry 2008, 64, 266–272. [Google Scholar] [CrossRef]
- Kuhlman, K.R.; Horn, S.R.; Chiang, J.J.; Bower, J.E. Early life adversity exposure and circulating markers of inflammation in children and adolescents: A systematic review and meta-analysis. Brain Behav. Immun. 2020, 86, 30–42. [Google Scholar] [CrossRef]
- Baumeister, D.; Akhtar, R.; Ciufolini, S.; Pariante, C.M.; Mondelli, V. Childhood trauma and adulthood inflammation: A meta-analysis of peripheral C-reactive protein, interleukin-6 and tumour necrosis factor-alpha. Mol. Psychiatry 2016, 21, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Kuhlman, K.R.; Cole, S.W.; Craske, M.G.; Fuligni, A.J.; Irwin, M.R.; Bower, J.E. Enhanced Immune Activation Following Acute Social Stress among Adolescents with Early-Life Adversity. Biol. Psychiatry Glob. Open Sci. 2023, 3, 213–221. [Google Scholar] [CrossRef] [PubMed]
- McGowan, P.O.; Sasaki, A.; D’Alessio, A.C.; Dymov, S.; Labonte, B.; Szyf, M.; Turecki, G.; Meaney, M.J. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat. Neurosci. 2009, 12, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Crisan, T.O.; Netea, M.G.; Joosten, L.A.B. Innate immune memory: Implications for host responses to damage-associated molecular patterns. Eur. J. Immunol. 2016, 46, 817–828. [Google Scholar] [CrossRef]
- Hanke, M.L.; Powell, N.D.; Stiner, L.M.; Bailey, M.T.; Sheridan, J.F. Beta adrenergic blockade decreases the immunomodulatory effects of social disruption stress. Brain Behav. Immun. 2012, 26, 1150–1159. [Google Scholar] [CrossRef]
- Coussons-Read, M.E.; Okun, M.L.; Nettles, C.D. Psychological stress increases inflammatory markers and alters cytokine production across pregnancy. Brain Behav. Immun. 2007, 21, 343–350. [Google Scholar] [CrossRef]
- Reid, B.; Danese, A. Challenges in researching the immune pathways between early life adversity and psychopathology. Dev. Psychopathol. 2020, 32, 1597–1624. [Google Scholar] [CrossRef]
- Cohen, S.; Janicki-Deverts, D.; Doyle, W.J.; Miller, G.E.; Frank, E.; Rabin, B.S.; Turner, R.B. Chronic stress, glucocorticoid receptor resistance, inflammation, and disease risk. Proc. Natl. Acad. Sci. USA 2012, 109, 5995–5999. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Nemeth, E. Iron Sequestration and Anemia of Inflammation. Semin. Hematol. 2009, 46, 387–393. [Google Scholar] [CrossRef]
- Nemeth, E.; Rivera, S.; Gabayan, V.; Keller, C.; Taudorf, S.; Pedersen, B.K.; Ganz, T. IL-6 mediates hypoferremia of inflammation by inducing the synthesis of the iron regulatory hormone hepcidin. J. Clin. Investig. 2004, 113, 1271–1276. [Google Scholar] [CrossRef] [PubMed]
- Augustine, L.F.; Nair, K.M.; Rao, S.F.; Rao, M.V.; Ravinder, P.; Balakrishna, N.; Laxmaiah, A.; Vazir, S. Adolescent Life-Event Stress in Boys Is Associated with Elevated IL-6 and Hepcidin but Not Hypoferremia. J. Am. Coll. Nutr. 2014, 33, 354–362. [Google Scholar] [CrossRef]
- Gambling, L.; Charania, Z.; Hannah, L.; Antipatis, C.; Lea, R.G.; McArdle, H.J. Effect of iron deficiency on placental cytokine expression and fetal growth in the pregnant rat. Biol. Reprod. 2002, 66, 516–523. [Google Scholar] [CrossRef]
- Cao, C.; Prado, M.A.; Sun, L.; Rockowitz, S.; Sliz, P.; Paulo, J.A.; Finley, D.; Fleming, M.D. Maternal Iron Deficiency Modulates Placental Transcriptome and Proteome in Mid-Gestation of Mouse Pregnancy. J. Nutr. 2021, 151, 1073–1083. [Google Scholar] [CrossRef]
- Manji, H.; Kato, T.; Di Prospero, N.A.; Ness, S.; Beal, M.F.; Krams, M.; Chen, G. Impaired mitochondrial function in psychiatric disorders. Nat. Rev. Neurosci. 2012, 13, 293–307. [Google Scholar] [CrossRef]
- Bastian, T.W. Potential Mechanisms Driving Mitochondrial Motility Impairments in Developing Iron-Deficient Neurons. J. Exp. Neurosci. 2019, 13, 117906951985835. [Google Scholar] [CrossRef] [PubMed]
- Bastian, T.W.; Von Hohenberg, W.C.; Georgieff, M.K.; Lanier, L.M. Chronic Energy Depletion due to Iron Deficiency Impairs Dendritic Mitochondrial Motility during Hippocampal Neuron Development. J. Neurosci. 2019, 39, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Bastian, T.W.; Rao, R.; Tran, P.V.; Georgieff, M.K. The Effects of Early-Life Iron Deficiency on Brain Energy Metabolism. Neurosci. Insights 2020, 15, 263310552093510. [Google Scholar] [CrossRef] [PubMed]
- Glombik, K.; Stachowicz, A.; Slusarczyk, J.; Trojan, E.; Budziszewska, B.; Suski, M.; Kubera, M.; Lason, W.; Wedzony, K.; Olszanecki, R.; et al. Maternal stress predicts altered biogenesis and the profile of mitochondrial proteins in the frontal cortex and hippocampus of adult offspring rats. Psychoneuroendocrinology 2015, 60, 151–162. [Google Scholar] [CrossRef] [PubMed]
- Lambertini, L.; Chen, J.; Nomura, Y. Mitochondrial Gene Expression Profiles Are Associated with Maternal Psychological Stress in Pregnancy and Infant Temperament. PLoS ONE 2015, 10, e0138929. [Google Scholar] [CrossRef]
- Guo, S.; Dong, Y.; Cheng, X.; Chen, Z.; Ni, Y.; Zhao, R.; Ma, W. Chronic Psychological Stress Disrupts Iron Metabolism and Enhances Hepatic Mitochondrial Function in Mice. Biol. Trace Elem. Res. 2023, 201, 1761–1771. [Google Scholar] [CrossRef]
- Prendergast, A.J.; Humphrey, J.H. The stunting syndrome in developing countries. Paediatr. Int. Child Health 2014, 34, 250–265. [Google Scholar] [CrossRef]
- Belfort, M.B.; Ramel, S.E. NICU Diet, Physical Growth and Nutrient Accretion, and Preterm Infant Brain Development. Neoreviews 2019, 20, e385–e396. [Google Scholar] [CrossRef]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR Signaling in Growth and Metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef]
- Monk, C.; Georgieff, M.K.; Osterholm, E.A. Research review: Maternal prenatal distress and poor nutrition—Mutually influencing risk factors affecting infant neurocognitive development. J. Child Psychol. Psychiatry 2013, 54, 115–130. [Google Scholar] [CrossRef]
- Suchdev, P.S.; Boivin, M.J.; Forsyth, B.W.; Georgieff, M.K.; Guerrant, R.L.; Nelson, C.A. Assessment of Neurodevelopment, Nutrition, and Inflammation From Fetal Life to Adolescence in Low-Resource Settings. Pediatrics 2017, 139, S23–S37. [Google Scholar] [CrossRef]
- Lubach, G.R.; Coe, C.L. Preconception maternal iron status is a risk factor for iron deficiency in infant rhesus monkeys (Macaca mulatta). J. Nutr. 2006, 136, 2345–2349. [Google Scholar]
Figure 1.
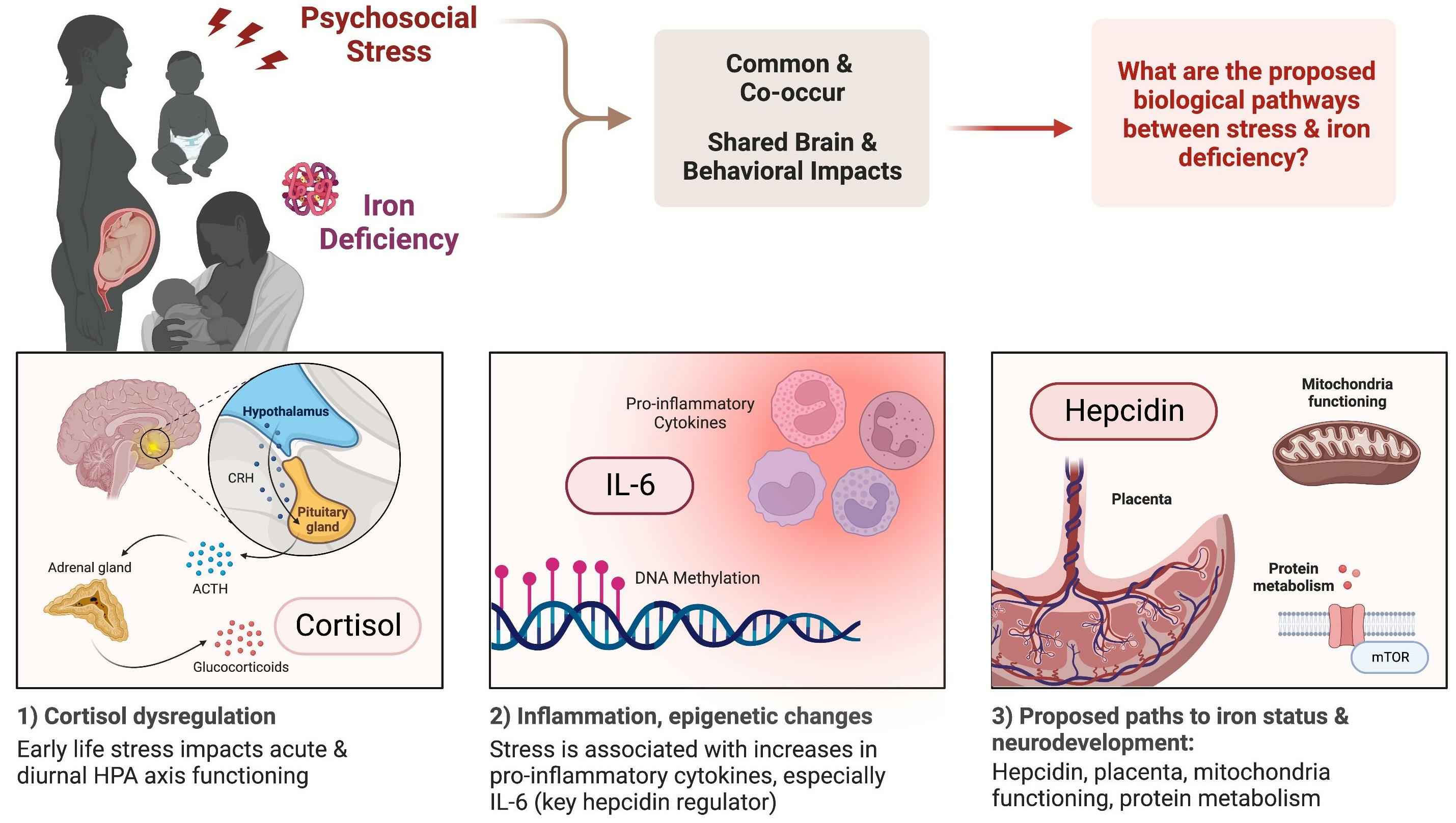
Exposure to psychological stress triggers the hypothalamic pituitary adrenocortical (HPA axis) response. The HPA axis response to a stressor begins with the release of corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP) by neurons in the medial parvocellular region of the paraventricular nucleus of the hypothalamus. These hormones stimulate the pituitary gland to secrete adrenocorticotropic hormone (ACTH), which triggers the adrenal cortex to produce glucocorticoids. These hormones bind to corticosteroid receptors (the glucocorticoid receptor (GR) and the mineralocorticoid receptor (MR)) throughout the brain and regulate gene expression, leading to various physiological and psychological effects [10]. The key to the HPA axis is feedback loops to maintain homeostasis. Once the perceived stressor has subsided, the feedback loops at multiple levels, including the hypothalamus, hippocampus, and frontal cortex, shut down the HPA axis and return the organism to homeostasis [10]. If a stressor is chronic, dysregulated cortisol output is seen in acute stress responses and the diurnal pattern of cortisol. These cortisol patterns are associated with epigenetic changes, increases in proinflammatory cytokines, and brain and behavior changes. We hypothesize that increased IL-6 from psychological stress impacts key pathways between iron metabolism and brain development. Figure created with Biorender.com [87].
Figure 1.
Exposure to psychological stress triggers the hypothalamic pituitary adrenocortical (HPA axis) response. The HPA axis response to a stressor begins with the release of corticotropin-releasing hormone (CRH) and arginine vasopressin (AVP) by neurons in the medial parvocellular region of the paraventricular nucleus of the hypothalamus. These hormones stimulate the pituitary gland to secrete adrenocorticotropic hormone (ACTH), which triggers the adrenal cortex to produce glucocorticoids. These hormones bind to corticosteroid receptors (the glucocorticoid receptor (GR) and the mineralocorticoid receptor (MR)) throughout the brain and regulate gene expression, leading to various physiological and psychological effects [10]. The key to the HPA axis is feedback loops to maintain homeostasis. Once the perceived stressor has subsided, the feedback loops at multiple levels, including the hypothalamus, hippocampus, and frontal cortex, shut down the HPA axis and return the organism to homeostasis [10]. If a stressor is chronic, dysregulated cortisol output is seen in acute stress responses and the diurnal pattern of cortisol. These cortisol patterns are associated with epigenetic changes, increases in proinflammatory cytokines, and brain and behavior changes. We hypothesize that increased IL-6 from psychological stress impacts key pathways between iron metabolism and brain development. Figure created with Biorender.com [87].


{kind=link}
{kind=link}
Table 1.
Shared neurodevelopmental consequences of insufficient iron and stress early in life.
| Insufficient Iron during Gestation and in Infancy | Maternal Stress Exposure and HPA Activation in Pregnancy, Infancy, Toddlerhood |
|---|---|
| Abnormal acute brain function | |
| Poor recognition memory * [44,45] Slower speed of neural processing * [46] Poor bonding and maternal interaction * [47] Impacts on hippocampus [41,43,54]; metabolic [42] and dendritic structure changes *† [43] Altered brain metabolism, myelination [35,36,37,38,39], neurotransmitter function *† [40] | ↑ cortical thinning and decrements in cognitive functioning * [59] Alterations in synaptic signaling and epigenetics in the hippocampus and amygdala, linked to ↑ anxiety and depressive behaviors † [69,70,71,72,73,74,75] ↓ Dendritic arborization in PFC, hippocampus † [76,77] Changes in hippocampal synaptic plasticity, ↓ spatial memory learning † [68,78,79] |
| Acute and long-term neurobehavioral abnormalities | |
| Motor Dysfunction † [52] Socio-Affective † [52] Neurocognitive, including ↓ memory performance (3rd trimester ID) * [50,51] | Impaired learning * [55] Impaired attention regulation and EF † [64,65,66] ↑ Anxiety and depressive behaviors * [56] ↑ Emotional reactivity in preschool-age offspring * [80] |
| Long-term mental health abnormalities | |
| ↑ Risk of ASD (1st trimester ID) * [48] ↑ Risk of schizophrenia (2nd trimester ID) * [49] | ↑ Risk for ASD schizophrenia [61] ↑ Risk of anxiety, *† depression, *† ADHD * [53,62,63,81] |
* = prenatal exposure, † = postnatal exposure, ID = iron deficiency, ↓ = decreased, ↑ = increased, ADHD = attention deficit hyperactivity disorder, PFC = prefrontal cortex, ASD = autism spectrum disorder, and EF = executive functioning.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Reid, B.M.; Georgieff, M.K. The Interaction between Psychological Stress and Iron Status on Early-Life Neurodevelopmental Outcomes. Nutrients 2023, 15, 3798. https://doi.org/10.3390/nu15173798
AMA Style
Reid BM, Georgieff MK. The Interaction between Psychological Stress and Iron Status on Early-Life Neurodevelopmental Outcomes. Nutrients. 2023; 15(17):3798. https://doi.org/10.3390/nu15173798
Chicago/Turabian StyleReid, Brie M., and Michael K. Georgieff. 2023. "The Interaction between Psychological Stress and Iron Status on Early-Life Neurodevelopmental Outcomes" Nutrients 15, no. 17: 3798. https://doi.org/10.3390/nu15173798
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.