
Neuroprotective and Anti-Neuroinflammatory Properties of Vignae Radiatae Semen in Neuronal HT22 and Microglial BV2 Cell Lines

Korean Medicine (KM)-Application Center, Korea Institute of Oriental Medicine, 70, Cheomdanro, Dong-gu, Daegu 41062, Republic of Korea
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2022, 14(24), 5265; https://doi.org/10.3390/nu14245265
Submission received: 2 November 2022
/
Revised: 7 December 2022
/
Accepted: 8 December 2022
/
Published: 10 December 2022
(This article belongs to the Special Issue Oxidative Stress and Protective Effects of Natural Products in Health)
Abstract
:The important factors in the pathogenesis of neurodegenerative disorders include oxidative stress and neuron-glia system inflammation. Vignae Radiatae Semen (VRS) exhibits antihypertensive, anticancer, anti-melanogenesis, hepatoprotective, and immunomodulatory properties. However, the neuroprotective effects and anti-neuroinflammatory activities of VRS ethanol extract (VRSE) remained unknown. Thus, this study aimed to investigate the neuroprotective and anti-inflammatory activities of VRSE against hydrogen peroxide (H2O2)-induced neuronal cell death in mouse hippocampal HT22 cells and lipopolysaccharide (LPS)-stimulated BV2 microglial activation, respectively. This study revealed that VRSE pretreatment had significantly prevented H2O2-induced neuronal cell death and attenuated reactive oxygen species generations in HT22 cells. Additionally, VRSE attenuated the apoptosis protein expression while increasing the anti-apoptotic protein expression. Further, VRSE showed significant inhibitory effects on LPS-induced pro-inflammatory cytokines in BV2 microglia. Moreover, VRSE pretreatment significantly activated the tropomyosin-related kinase receptor B/cAMP response element-binding protein, brain-derived neurotrophic factor and nuclear factor erythroid 2-related factor 2, and heme oxygenase-1 signaling pathways in HT22 cells exposed to H2O2 and inhibited the activation of the mitogen-activated protein kinase and nuclear factor-κB mechanism in BV2 cells stimulated with LPS. Therefore, VRSE exerts therapeutic potential against neurodegenerative diseases related to oxidative stress and pathological inflammatory responses.
1. Introduction
Oxidative stress and excessive inflammatory responses are closely related to the pathogenesis of the various neurodegenerative disorders of the central nervous system (CNS) [1,2]. Oxidative stress is characterized by a disequilibrium between the oxygen-derived radical generation and the antioxidant potential of organisms, thereby damaging cellular components and altering cellular function, ultimately contributing to cell death [3,4]. Reactive oxygen species (ROS) at low levels are important in neuronal signaling and physiology. However, ROS overproduction will damage proteins, DNA, and lipid membranes, leading to cellular destruction and cell death [5,6]. Microglia are major resident immune cells that play an important role in mediating inflammatory responses in the CNS. Sustained excessive activation of microglia contributes to neuronal damage and inflammatory mediator releases, such as tumor necrosis factor (TNF)-α, interleukin (IL)-6, and IL-1β, in response to lipopolysaccharide (LPS) stimulation. Overproduction of these mediators may contribute to neurodegenerative disorder progression [7,8]. Additionally, ROS production increases by an excessive inflammatory response, and oxidative stress triggers the microglia activation. Therefore, oxidative stress and excessive inflammatory response may be the major factors that cause or exacerbate neurodegenerative diseases.
Reducing the risk of oxidative stress caused by ROS overproduction and the excessive inflammatory response is a key target for treating or preventing neurodegenerative diseases. Brain-derived neurotrophic factor (BDNF) is a neurotrophin within the central and peripheral nervous systems, is concerned with neuronal cell preservation in in vivo systems, and is known to play an important role [9]. BDNF elicits its physiological function through binding to its receptor, tropomyosin-related kinase receptor B (TrkB), which initiates multiple signaling cascades [10,11,12]. Additionally, BDNF/TrkB signaling activation contributes to the phosphorylation and activation of transcription factor cAMP response element-binding protein (CREB) [13]. Another mechanism that performs neuroprotective action is correlated with the nuclear factor erythroid 2-related factor 2 (Nrf-2) signaling pathway. The Nrf-2, as a transcription factor, regulates the expression of many antioxidant enzymes, including NAD(P)H quinone oxidoreductase 1 (NQO-1), glutamate-cysteine ligase catalytic subunit (GCLC), and heme oxygenase (HO)-1 [14,15,16]. Many studies revealed that Nrf-2-mediated pathways can scavenge free radicals and efficiently protect neuronal cells from oxidative injury [17,18]. Therefore, it can be used as a strategy for pharmacological agent development that targets TrkB/CREB/BDNF pathways and the Nrf-2/antioxidant enzyme for neurodegenerative disease treatment and prevention.
Neuroinflammation is an inflammatory process of the neural cells and is triggered by various biological stimuli, such as glial reactions and oxidative stress [19]. Nuclear factor (NF)-κB and mitogen-activated protein kinases (MAPK) are known as key inflammatory signaling pathways during inflammatory process progression. The major subunits of the NF-κB heterodimer include p65 and p50. Under stimulation, NF-κB translocates into nuclei to promote several inflammatory mediator expressions, including TNF-α, IL-6, and IL-1β [20,21]. The family members of MAPKs mainly consist of the extracellular signal-regulated kinase (ERK), p38, and c-Jun NH2-terminal kinase (JNK) [22], which are related to the regulation of inflammatory mediator production in stimulated microglia [23]. Therefore, targeting NF-κB and MAPK pathways might be an attractive therapeutic approach for neuroinflammatory disease prevention and treatment.
Vigna radiatus is a leguminous species that is mainly grown in Asia, and its seeds, Vignae Radiatae Semen (VRS), are used for food or medicinal purposes. VRS is one of the most important edible legumes and contains balanced nutrients, including protein, dietary fiber, minerals, vitamins, and significant lots of bioactive compounds [24]. VRS has been traditionally used as a medicinal herb for its detoxification activities, mentality recuperation, heat stroke alleviation, and gastrointestinal upset regulation. Previous studies reported that VRS is beneficial because of its hypoglycemic and hypolipidemic effects, as well as antihypertensive, anticancer, anti-melanogenesis, hepatoprotective, and immunomodulatory properties beyond meeting basic nutrient requirements [25,26,27,28,29,30]. However, the physiological activity and molecular mechanisms of HT22 hippocampal neuron-mediated neuroprotective and microglia-mediated anti-neuroinflammatory effects generated by VRS ethanol extract (VRSE) remained unknown. Hence, the current study aimed to determine the neuroprotective effects and anti-neuroinflammatory activities of VRSE in H2O2-exposed HT22 hippocampal cells and LPS-stimulated BV2 microglial cells. Furthermore, the chemical constituents of VRSE were investigated using high-performance liquid chromatography (HPLC) analysis.
2. Materials and Methods
2.1. Materials and Reagents
Dulbecco’s modified Eagle’s medium (DMEM), antibiotics, and fetal bovine serum (FBS) were purchased from Hyclone (Logan, UT, USA). All cell culture dishes and plates were obtained from SPL life sciences (Pocheon, Republic of Korea). LPS, bovine serum albumin (BSA), and dimethyl sulfoxide were purchased from Sigma–Aldrich (St. Louis, MO, USA). The Cell counting kit (CCK) was purchased from Dojindo Molecular Technologies (Kumamoto, Japan). The 2′,7′-dichlorofluorescein diacetate (H2DCFDA) was obtained from Invitrogen (Carlsbad, CA, USA). Various primary antibodies and horseradish peroxidase-conjugated secondary antibodies were obtained from Cell Signaling Technology, Inc. (Boston, MA, USA), Santa Cruz Biotechnology (Santa Cruz, CA, USA), and Thermo Fisher (Waltham, MA, USA). The polyvinylidene difluoride (PVDF) membrane was acquired from Millipore (Bedford, MA, USA). Enzyme-linked immunosorbent assay (ELISA) antibody sets were obtained from eBioscience (San Diego, CA, USA). Vitexin and isovitexin were purchased from Sigma–Aldrich. HPLC grade acetonitrile was obtained from Merck (KGaA, Darmstadt, Germany) and formic acid was purchased from Sigma–Aldrich. Tertiary distilled water was prepared by Puris-Evo-UP Water System with Evo-up Dio VFT and Evo-ROP Dico20 (Mirae ST Co., Ltd., Anyang, Republic of Korea).
2.2. Preparation of VRSE
VRS was purchased as a dried herb from Yeongcheonhyundai Herbal Market (Yeongcheon, Korea) and authenticated by Prof. KiHwan Bae of the College of Pharmacy, Chungnam National University (Daejeon, Republic of Korea). All voucher specimens were deposited in an herbal bank at the Korean Medicine-Application Center, Korea Institute of Oriental Medicine (voucher number: E26). The dried herb (30.0 g) was extracted in 70% ethanol (390 mL) at 40 °C in a shaking incubator (100 rpm) for 24 h. The extract solution was filtered through a 150 mm filter paper (Whatman, Piscataway, NJ, USA) and concentrated using a rotary vacuum evaporator (Buchi, Tokyo, Japan). Samples were then freeze-dried and kept in desiccators at −20 °C before use. The sample yield was 7.4693%, and 2.2408 g of extract was obtained.
2.3. Cell Culture
The murine hippocampal HT22 cell and microglial cell line BV2 were purchased from the American Type Culture Collection (Manassas, VA, USA) and grown in a DMEM medium that contains 10% of FBS and 1% of antibiotics. Each cell was maintained in a humidified incubator (37 °C, 95% of air, and 5% of CO2) during cultivation. The culture medium was renewed every 2 days, and cells were grown to 80–85% confluence for the experiments.
2.4. Cell Viability
HT22 cells (5 × 103 cells/well) were seeded and cultured into a 96-well plate for 24 h. BV2 cells were seeded into 96-well plates at a density of 5 × 104 cells/well and cultured for 18 h. After pre-incubation, VRSE or H2O2 was added to the cells, which were incubated for 24 h at 37 °C with 5% of CO2. Each well was added with 10 μL of CCK solution and incubated for 1 h. The absorbance of each well at 450 nm was measured using a microplate reader (SpectraMax i3, Molecular Devices, San Jose, CA, USA). Morphological changes of HT22 cells were analyzed by a holotomography (HT-X1, Tomocube, Daejeon, Republic of Korea).
2.5. Intracellular ROS Determination
The measurement of intracellular ROS levels was examined using H2DCFDA following our previous report [31]. The cells were washed twice in phosphate-buffered saline (PBS) and were then stained with 10 μM of H2DCFDA for 30 min at 37 °C in the dark after VRSE and H2O2 treatment. The stained cells were washed twice with PBS. A fluorescence microplate reader (SpectraMax i3, Molecular Devices, San Jose, CA, USA) at an excitation wavelength of 488 nm and emission wavelength of 525 nm was used to analyze dihydroethidium fluorescence. A fluorescence microscope (Eclipse Ti, Nikon, Tokyo, Japan) was used to obtain representative fluorescence images.
2.6. Western Blotting
Cell proteins were extracted using radioimmunoprecipitation assay lysis buffer (Millipore) by adding protease and phosphatase inhibitor cocktail (Roche, Basel, Switzerland). Protein samples were separated via electrophoresis in a sodium dodecyl sulfate-polyacrylamide gel and transferred onto a PVDF membrane. The membranes were blocked with 3% BSA at room temperature (RT) for 1 h and were incubated with their respective primary antibodies overnight at 4 °C. Primary antibodies were discarded, and the blots were washed with tris-buffered saline with 0.1% of Tween 20 four times. Membranes were incubated with horseradish-labeled species-specific secondary antibodies at RT for 1 h. ChemiDocTM Touch Imaging System (Bio-Rad, Hercules, CA, USA) was used to quantify the relative protein expression intensity. The band intensity was analyzed using Image J software (version 1.53k) with control value normalization. Table 1 shows the information regarding various primary and secondary antibodies.
2.7. Preparation of Cytosolic and Nuclear Fractions
Nuclear Extraction Kit (Thermo Fisher) was used to obtain the cytosolic and nuclear fractions. Each extracted fraction was lysed according to the protocol by the manufacturer’s instructions. The fractions were stored at −80 °C before use.
2.8. Cytokine Determination
The cytokine level in the culture medium was determined using an ELISA antibody set according to the manufacturer’s instructions. For ELISA, BV2 cells were seeded into a 24-well plate (2.5 × 105 cells/well) and incubated overnight. The cells were pretreated with various VRSE concentrations for 1 h and further challenged with LPS for an additional 24 h at 37 °C with 5% of CO2 [32]. After incubation, the TNF-α and IL-6 cytokine levels were measured using the collected supernatant of the medium as previously reported [32].
2.9. VRSE and Standard Solution Preparation
Standard component purity was confirmed that all were >95%. Each constituent (vitexin and isovitexin) stock solution was prepared with HPLC grade methanol at the concentration of 1000 ppm (1 mg/mL) after dissolving the solution through a 0.2-μm PETE filter. The filtrate was diluted with methanol at the concentration for the prepared standard curve for each compound. VRSE at 100 mg was accurately weighed and dissolved in 1 mL of methanol:water (50:50, v/v) solvent. VRSE solution was filtered through a 0.2-μm PETE filter before HPLC injection.
2.10. HPLC Conditions
The VRSE was analyzed using the Dionex UltiMate 3000 HPLC system (Dionex Corp., Sunnyvale, CA, USA) consisting of a binary pump, an autosampler, a column oven, and a diode array UV/VIS detector. System control and data analysis were processed with Dionex Chromeleon. All analysis was conducted under the following conditions: Kinetex C18 column (100 × 4.60 mm, 2.6 μm, Phenomenex, Seoul, Republic of Korea), and the column oven temperature was kept at 40 °C, mobile phase consisted of 0.1% of formic acid (v/v) in water (A) and acetonitrile (B) gradient elution system to improve the chromatographic separation capacity. The gradient system was programmed as 13% B for 0−25 min and 13−100% B for 25−26 min with a flow rate of 0.6 mL/min. The total run time was 40 min, and the injection volume was 5 μL. The detection wavelength was 280 nm and all samples were injected triplicated.
2.11. Statistical Analysis
The data are presented as the mean ± standard error of the mean for three independent experiments. Statistical analysis of the experimental results was performed using the one-way analysis variance followed by Dunnett’s test, after comparing the H2O2 or LPS sample and each treated sample. A p-value of <0.05 represented a statistical significance. GraphPad Prism version 5.02 (GraphPad Software, Inc., 188 San Diego, CA, USA) was used for statistical analyses and mapping.
3. Results
3.1. Effects of VRSE on H2O2-Induced Neurotoxicity in HT22 Cells
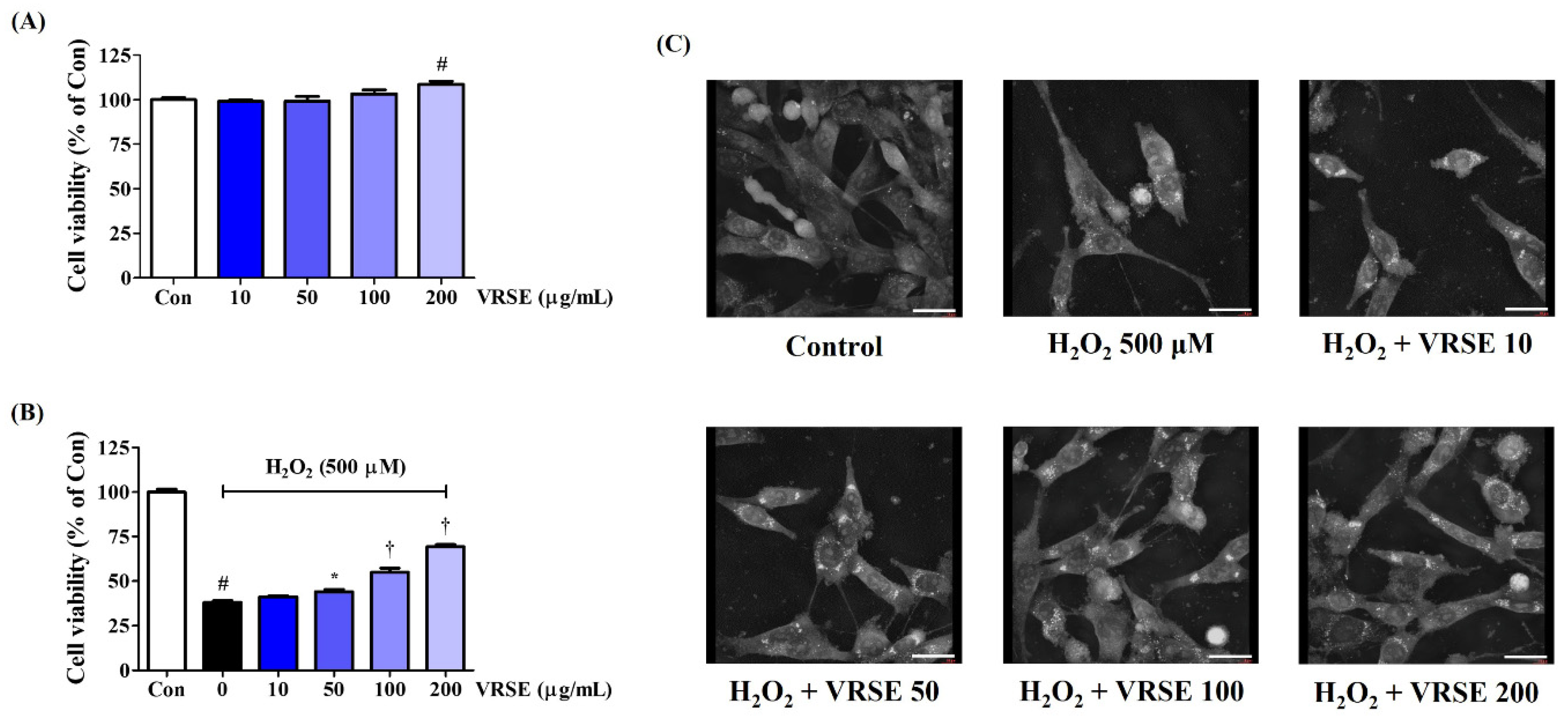
We assessed the effect of VRSE on the viability of HT22 mouse hippocampal neurons using the CCK assay to investigate the neuroprotective effects. VRSE treatment of up to 200 μg/mL alone for 24 h did not show any cytotoxicity on HT22 cells, as shown in Figure 1A. Thus, VRSE concentrations of <200 μg/mL were then investigated for neuroprotection activities. First, an additional CCK assay was performed to verify the possible protective effects of VRSE against the H2O2-induced neurotoxicity in HT22 cells. Our data revealed that H2O2 treatment alone at 500 μM decreased cell viability by approximately 70%. In contrast, VRSE treatment improved the cell viability of H2O2-exposed HT22 cells. At 100 and 200 μg/mL concentrations, the cell viability increased with statistical significance (Figure 1B). Additionally, H2O2-induced nuclear condensation and cell contraction were found by observing the cell morphology with a holotomography. In contrast, VRSE pretreatment had a concentration-dependent effect on maintaining cellular structure and inhibiting morphological changes (Figure 1C). Therefore, VRSE significantly mitigates neurotoxicity in H2O2-exposed HT22 cells.
3.2. VRSE Reduced Intracellular ROS Generation by H2O2
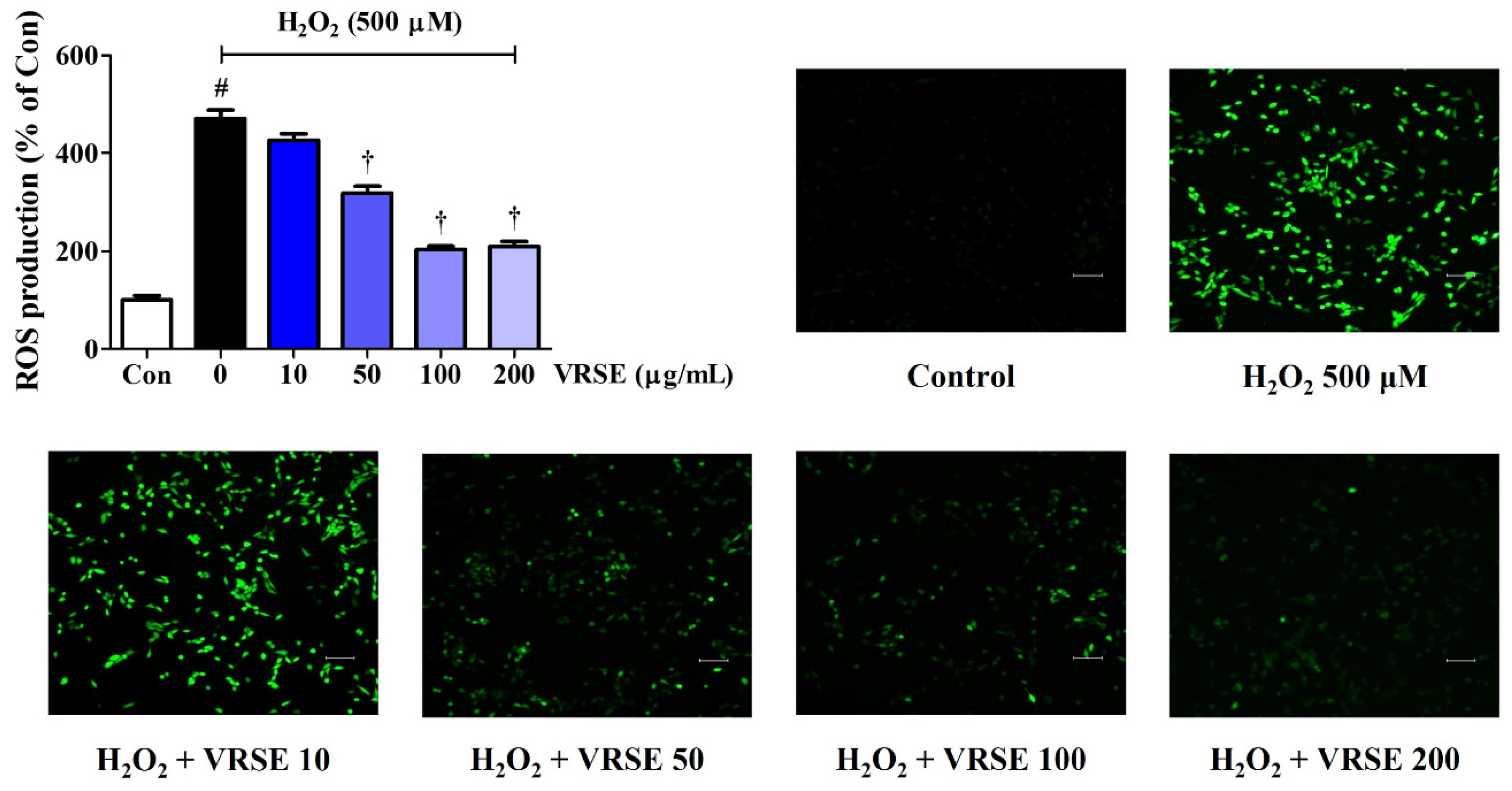
We also explored the effects of several VRSE concentrations (10, 50, 100, and 200 μg/mL) on ROS accumulation in HT22 cells using an H2DCFDA fluorescence assay. Our data revealed that H2O2 treatment alone significantly increased intracellular ROS levels, whereas VRSE pretreatment concentration dependently reduced ROS generation (Figure 2). The fluorescent images in Figure 2 also show an effect similar to that mentioned above, suggesting that VRSE could inhibit excessive ROS production induced by H2O2.
3.3. VRSE Suppress H2O2-Induced Apoptosis in HT22 Cells
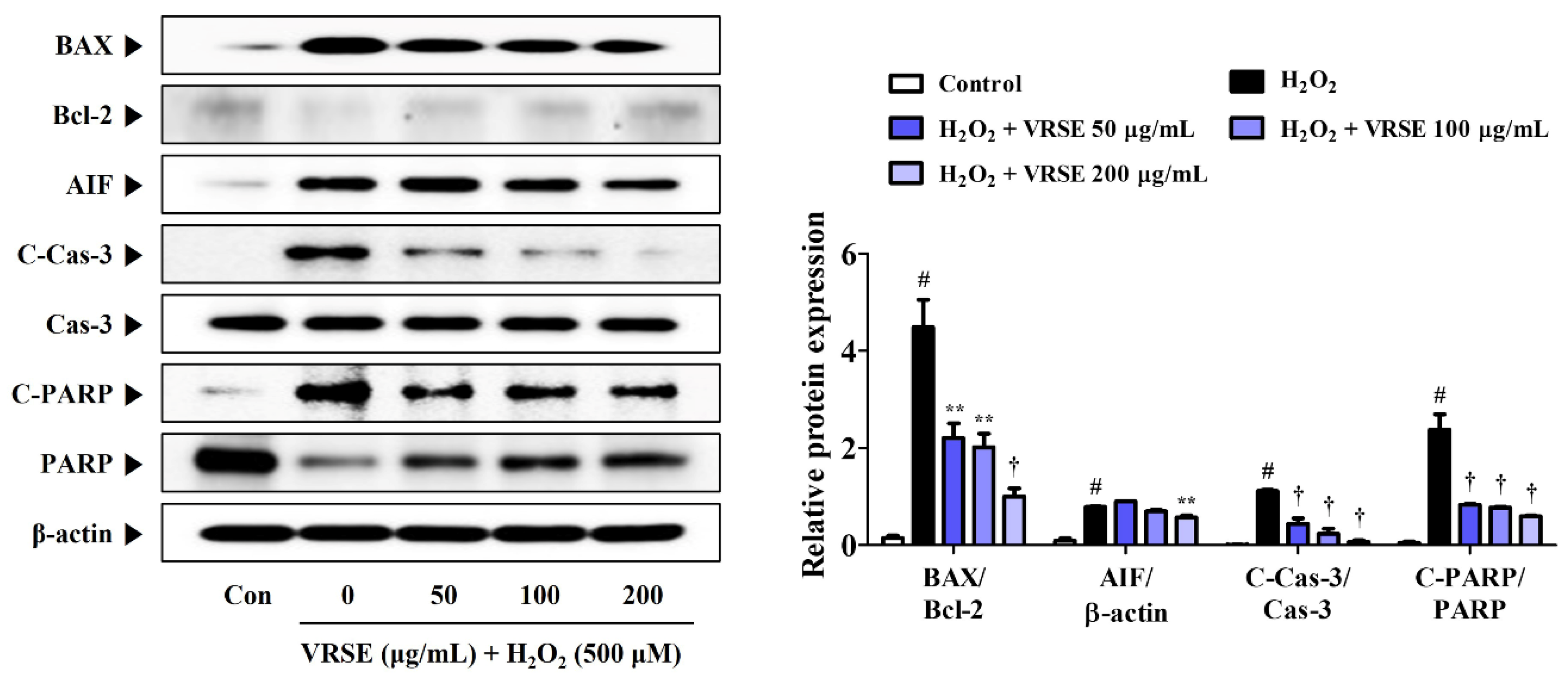
The expression of apoptosis-related proteins was determined using Western blot analysis to measure the prevention effects of VRSE on H2O2-induced apoptotic neurotoxicity in HT22 cells. H2O2 treatment increased the apoptotic marker expression in HT22 cells, as shown in Figure 3. In contrast, VRSE pretreatment notably diminished the apoptosis-related protein level, such as Bcl-2-associated X (BAX), apoptosis-inducing factor (AIF), cleaved-caspases-3, and cleaved-Poly (ADP-ribose) polymerase (PARP). Moreover, it enhanced the anti-apoptotic protein expression, such as B-cell lymphoma 2 (Bcl-2) and PARP compared with H2O2 alone-treated cells (Figure 3). These results revealed that VRSE pretreatment exerts a neuroprotective activity via H2O2-induced apoptotic cell death inhibition.
3.4. Effects of VRSE on the Production of Inflammatory Cytokines in LPS-Stimulated BV2 Cells
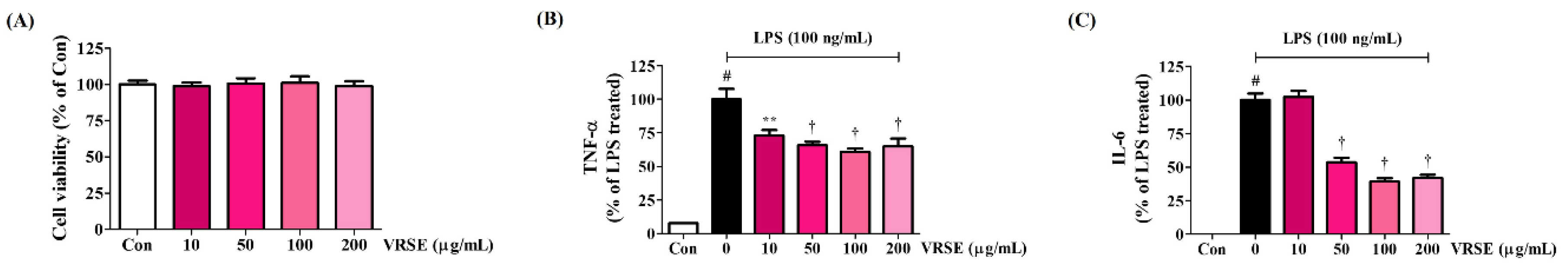
First, we investigated the effect of VRSE treatment on cell viability in BV2 microglia to explore the concentration at which potential cytotoxicity was eliminated. Hence, VRSE did not affect cell viability up to a concentration of 200 μg/mL, as in HT22 cells (Figure 4A). We measured the production of inflammatory cytokines, including TNF-α and IL-6, in mouse microglia BV2 cells to identify the anti-neuroinflammatory effects of VRSE. The secretion of cytokines, which was greatly increased when LPS alone was treated, was significantly decreased in a concentration-dependent manner by VRSE pretreatment, as shown in Figure 4B,C.
3.5. VRSE Ameliorate the Transcriptional Activity of NF-κB and Phosphorylation of MAPK in LPS-Stimulated BV2 Microglia
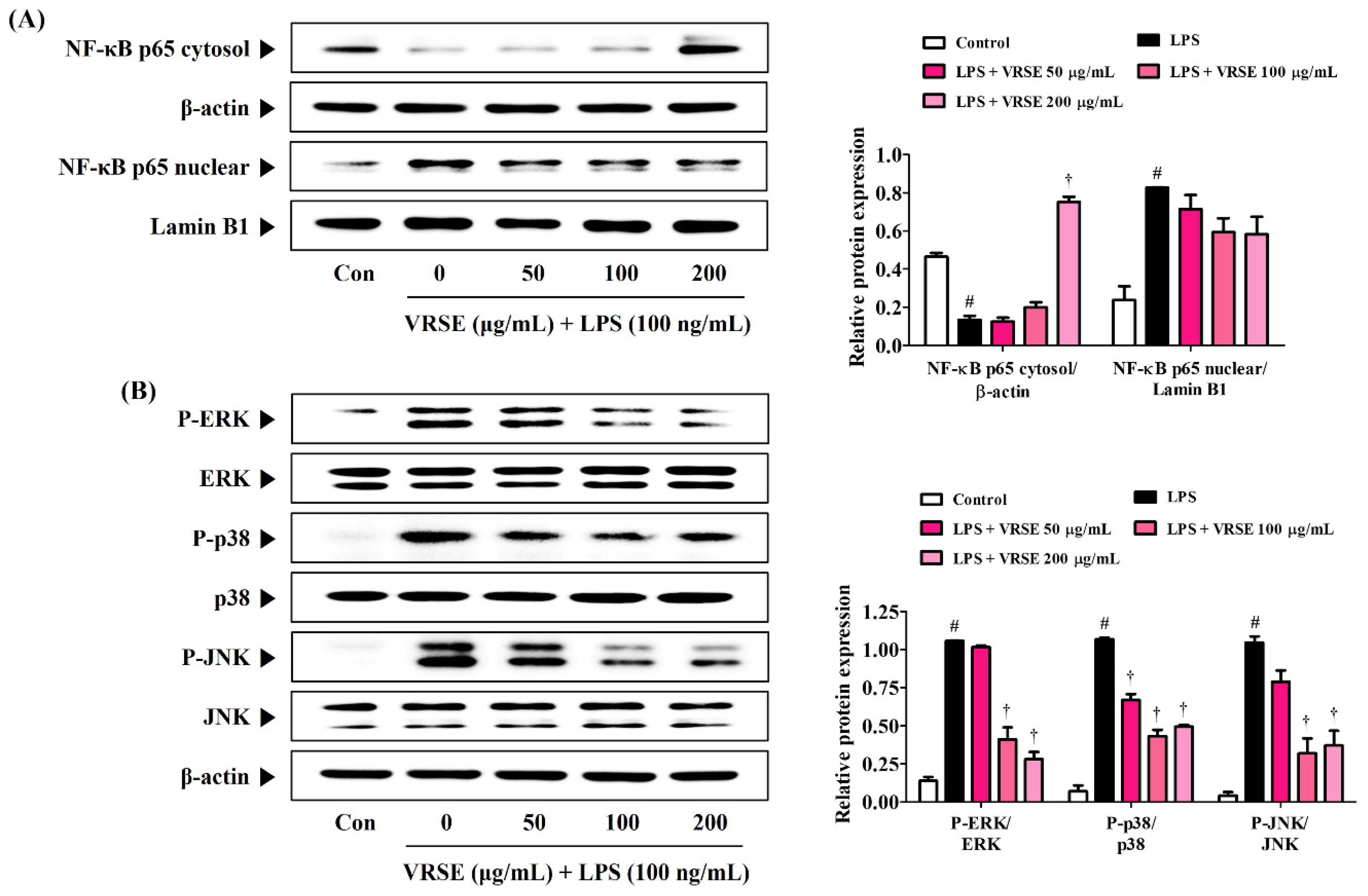
The transcription factor, NF-κB, is an essential regulator that is closely related to inflammatory responses. Therefore, we determined the inhibition effect of VRSE in LPS-induced p65 translocation into the nucleus via Western blot analysis. Figure 5A shows that VRSE effectively blocks LPS-induced translocation of NF-κB subunit p65 to the nucleus in a concentration-dependent manner. Additionally, MAPK, ERK, p38, and JNK are important immune response regulators and have a crucial role in regulating NF-κB activation. Thus, we assessed the effects of VRSE on the phosphorylation of MAPK. Our data revealed that VRSE pretreatment statistically significantly diminished the phosphorylation of ERK, p38, and JNK (Figure 5B).
3.6. Effects of VRSE on Nrf-2-Mediated Antioxidant Enzyme Expression in H2O2-Exposed HT22 Hippocampal Cells
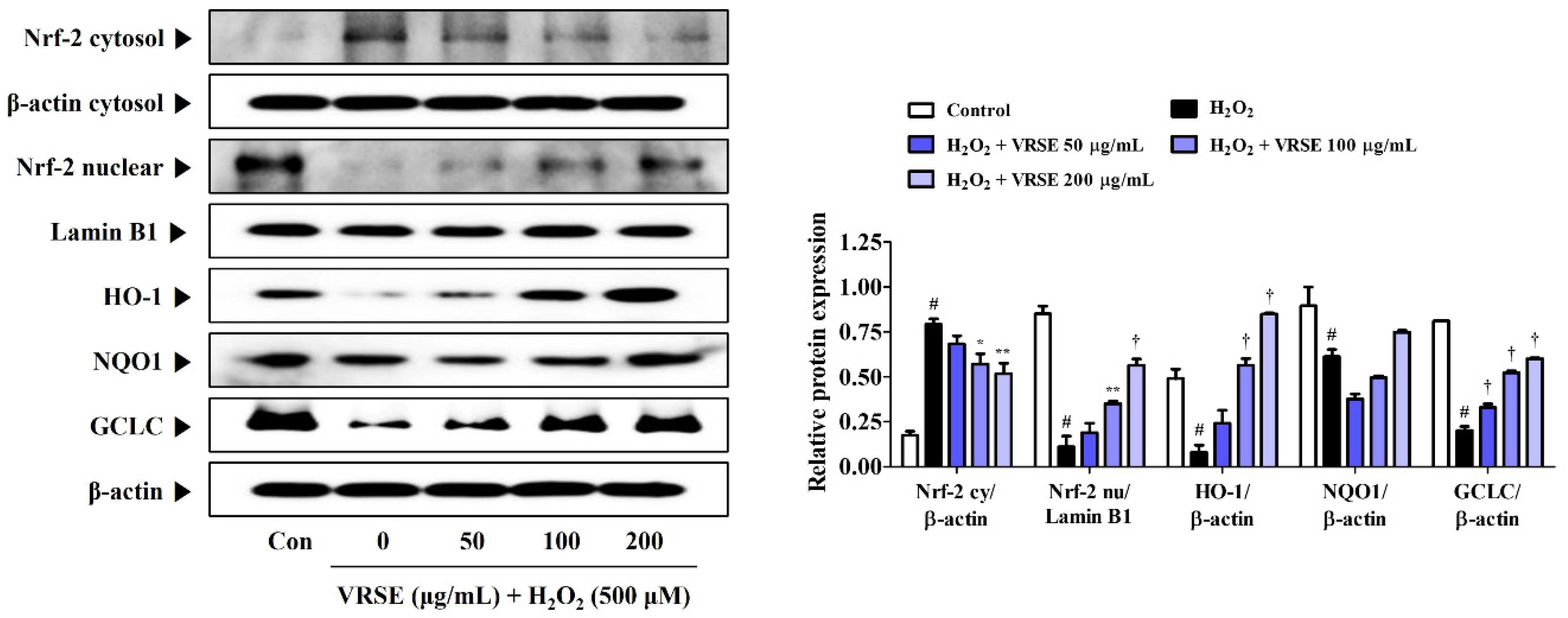
The protective action of antioxidant enzymes by Nrf-2 activation suppresses oxidative stress in several models of neurodegenerative diseases, thereby exhibiting neuroprotective effects [17,18]. Therefore, we examined the induction effect of VRSE pretreatment in antioxidant enzyme expression, including HO-1, NQO-1, and GCLC, through Nrf-2 translocation to the HT22 cell nucleus. Figure 6 shows that VRSE completely increased the nuclear Nrf-2 translocation in a concentration-dependent manner. Furthermore, VRSE pretreatment strongly increased the antioxidant enzyme expression, including HO-1, NQO-1, and GCLC, in H2O2-exposed HT22 cells (Figure 6). Therefore, the antioxidant effects of VRSE may be due to Nrf-2-mediated antioxidant enzyme activation.
3.7. VRSE Enhances Mature BDNF Expression via TrkB/Akt/CREB Pathway Activation
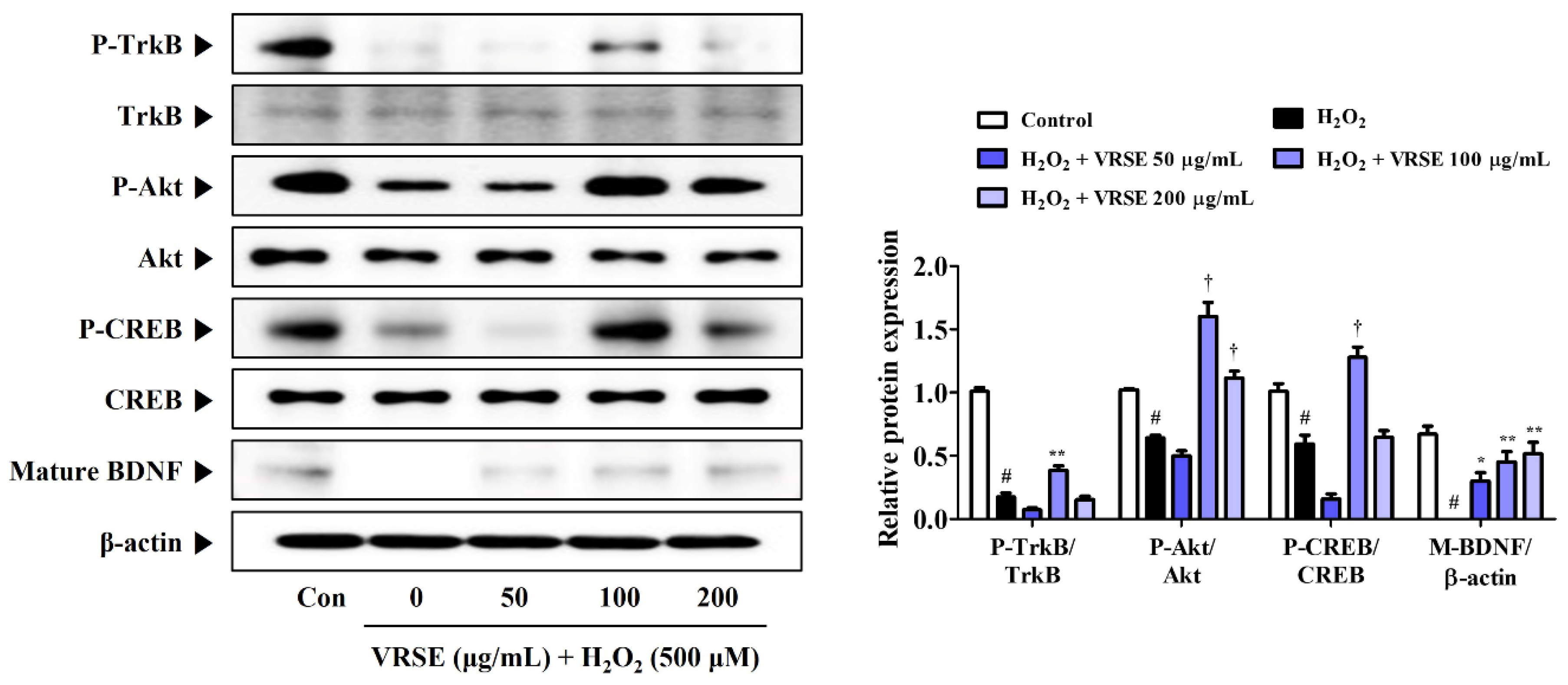
TrkB/Akt/CREB/BDNF pathway activation is involved in neuronal growth and survival. Hence, we determined the effects of VRSE on BDNF formation and change of TrkB/Akt/CREB pathway activation. Figure 7 shows that VRSE effectively improved the BDNF expression, as well as P-TrkB, P-Akt, and P-CREB, only at a 100 μg/mL concentration, compared to cells treated with H2O2 alone. Conversely, VRSE pretreatment at low and 200 mg/mL concentrations showed little or no effect. Therefore, VRSE had a significant effect on TrkB/Akt/CREB/BDNF activation to some extent but was not concentration-dependent. Therefore, the neuroprotective effects of VRSE are only partially related to TrkB/Akt/CREB/BDNF signaling pathway activation.
3.8. Identification and Quantitative Analysis of the Chemical Constituents of VRSE
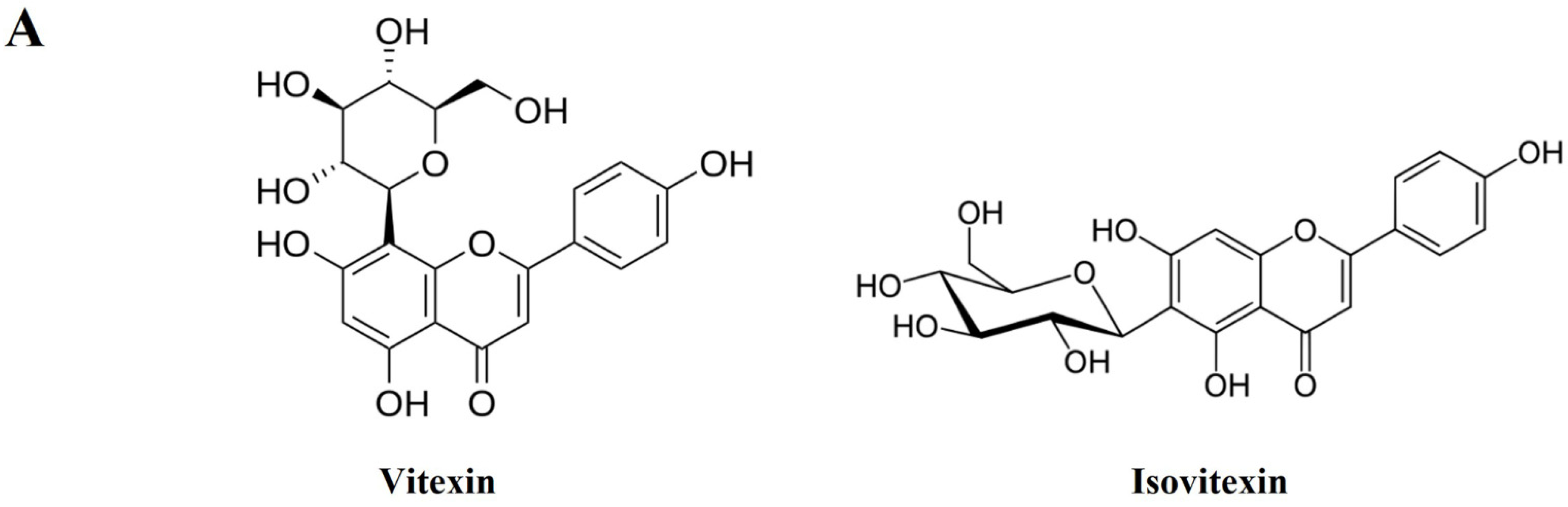
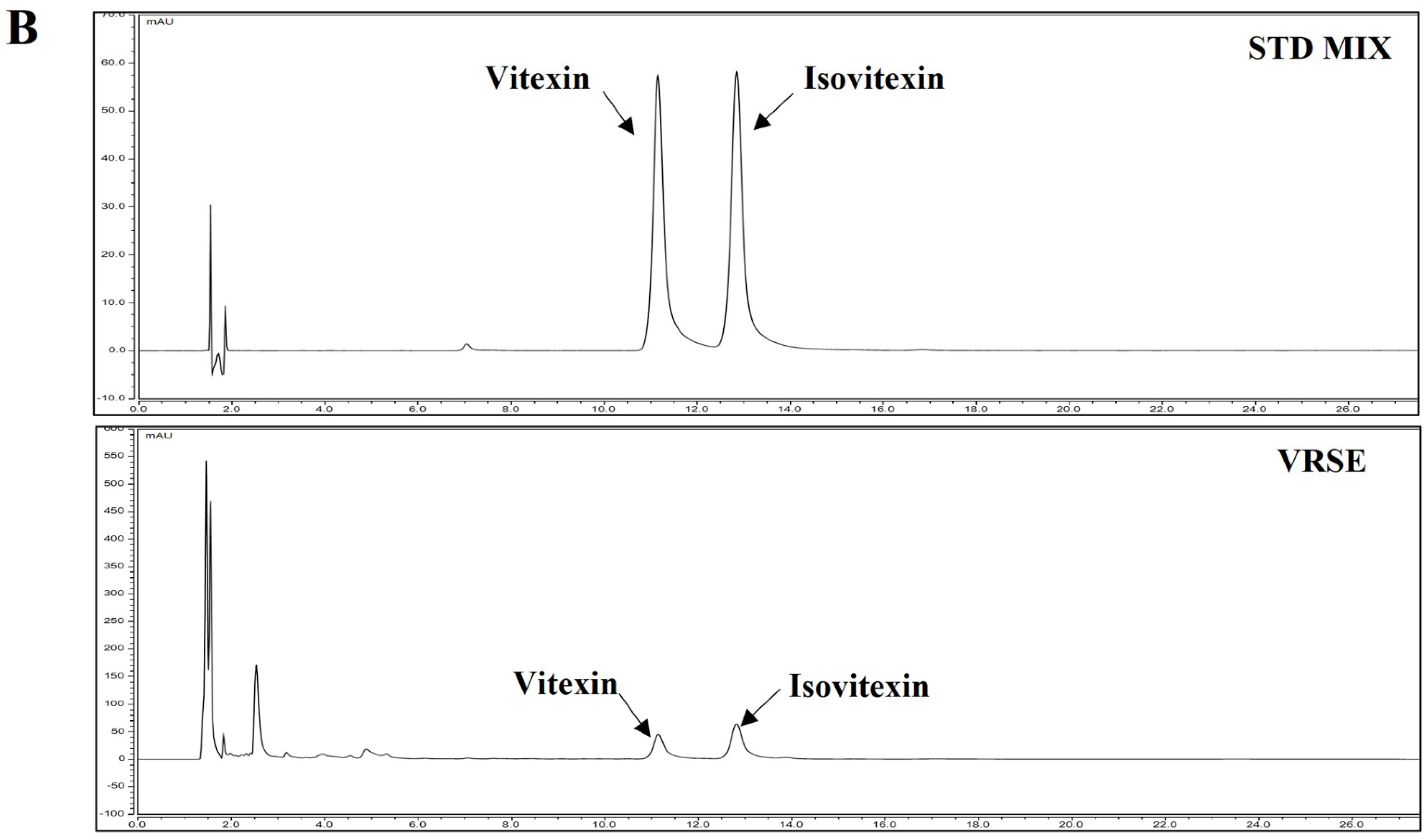
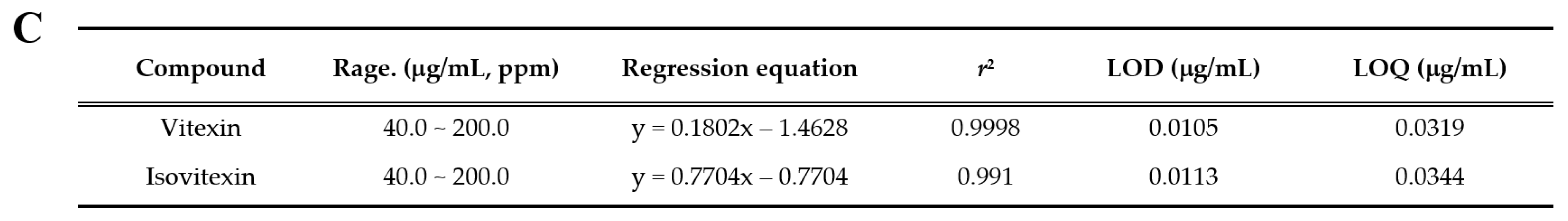
HPLC analysis was performed to determine the compounds contained in VRSE and their quantification. Two compounds of vitexin (tR 11.140 min) and isovitexin (tR 12.817 min) were detected under the established HPLC condition (Figure 8). The calibration curves of the two marker compounds were y = 0.1808x−1.4628 and y = 0.1672x−0.7704, with determination coefficients of 0.9998 and 0.9991 at injected concentration ranges of 40.0–200.0 μg/mL, respectively. These results showed the good linearity of the calibration curve of two marker compounds at the tested concentration range. The mean area of VRSE was calculated for each compound calibration curve equation. The contents of vitexin and isovitexin were 1243 μg/g and 1635 μg/g, respectively. Thus, we confirmed the contents of two marker compounds in the VRSE using the developed HPLC-UV analysis.
4. Discussion
Oxidative stress and excessive inflammatory responses pose the CNS to susceptible damage and various neurodegenerative disorders, such as Alzheimer’s and Parkinson’s diseases [1,2]. Thus, an agent with both antioxidative and anti-inflammatory effects was proposed as a promising strategy for several neurodegenerative disease prevention and treatment [33]. VRS, the most important edible legume crop, has long been used as food and traditional medicine in East Asia. VRS has been traditionally used as a medicinal herb for its detoxification activities, mentality recuperation, heat stroke alleviation, and gastrointestinal upset regulation. However, the neuroprotective and anti-neuroinflammatory effects and detailed molecular mechanisms of VRS remained unknown. Therefore, the current study explored the therapeutic potential and underlying mechanism of VRSE in H2O2-exposed HT22 hippocampal cells and LPS-stimulated BV2 microglial cells.
This study revealed that VRSE pretreatment protected HT22 cells from H2O2-induced cytotoxicity, as evidenced by high cell viability (Figure 1). Additionally, VRSE significantly reduced the intracellular ROS level caused by H2O2 treatment (Figure 2). Moreover, H2O2-induced oxidative stress resulted in cell death as determined using Western blot analysis. Our results indicate that VRSE pretreatment remarkably inhibited the mitochondrial apoptotic proteins, including BAX, AIF, cleaved-caspase-3, and cleaved-PARP, as well as significantly increased the anti-apoptotic protein expression, including Bcl-2 and PARP (Figure 3). These results suggested that VRSE protected neuronal cell death by suppressing the excessive ROS accumulation through H2O2 treatment. One possible mechanism concerning the neuroprotective action of VRSE may be correlated with TrkB/CREB/BDNF pathway. BDNF is a member of the neurotrophin family of growth factors that influences cell proliferation, differentiation, neuronal growth, and synaptic plasticity by binding to the Trk receptor [10,11,12]. Thus, BDNF is an important factor for oxidative stress-treated neuronal cell survival. The TrkB activation by BDNF in the signaling pathway contributes to the phosphorylation and activation of CREB in neuronal cells [13]. Our present data revealed that VRSE not only enhanced the BDNF level but also elevated the P-TrkB and P-CREB activation (Figure 7). Consequently, these results indicated that VRSE induces BDNF expression, and then the released BDNF activates TrkB/CREB pathway in HT22 cells. Another possible mechanism for the neuroprotection of VRSE may be related to antioxidant enzyme regulation. Nrf-2 is a transcription factor that initiates an endogenous antioxidant response element and activates several antioxidant enzyme productions, including NQO1, GCLC, and HO-1 [14,15,16,34]. Under oxidative stress stimulation, Nrf-2 dissociates with KEAP1, and then Nrf-2 is combined into the ARE region on DNA after it translocates into the nucleus and activates the antioxidant enzyme secretion in neuronal cells [35,36]. Therefore, our experimental results revealed that VRSE treatment dramatically improved the nuclear Nrf-2 translocation and antioxidant enzyme expressions, such as GCLC, NQO1, and HO-1, in H2O2-treated HT22 cells (Figure 6). Therefore, VRSE exerts neuroprotective effects by promoting the antioxidant enzyme expression through Nrf-2 activation in oxidative stress-induced neuronal cells.
The pathogenesis of various neurodegenerative diseases is closely related to the neuroinflammatory process, which can be triggered by various biological stimuli, such as glial reactions and oxidative stress [19]. Microglia are major resident immune cells and play an important role in mediating inflammatory responses in the CNS. Microglia-mediated neuroinflammation is considered a major source of ROS and reactive nitrogen species in the brain [37]. Excessively activated microglia by LPS stimulation can lead to neuronal cell death and generate pro-inflammatory factors, including TNF-α, IL-6, and IL-1β. Our study results revealed that VRSE significantly decreased the TNF-α and IL-6 secretion in LPS-activated BV2 microglial cells (Figure 4). Additionally, we evaluated the MAPK phosphorylation and NF-κB signaling pathway activation to investigate the potential mechanisms of anti-neuroinflammatory effects in LPS-stimulated BV2 cells. NF-κB, as a key transcription factor, plays an essential role in immune and inflammatory responses. Inactivated p65 of NF-κB is located in the cytoplasm in unstimulated cells bound to inhibit NF-κB alpha (IκBα). Microglial cell stimulation by inflammatory stimulants, such as LPS, significantly enhances the phosphorylation and degradation of IκBα, which then frees NF-κB to translocate into the nucleus to promote pro-inflammatory factors [37]. Hence, we decided to measure the p65 expression level in cytoplasmic and nuclear extracts. Western blot analysis revealed that p65 translocation into the nucleus strongly increased by LPS stimulation, whereas VRSE treatment notably inhibited nuclear NF-κB p65 migration in microglia BV2 cells. The MAPK pathway plays a significant role in the LPS-treated expression of several endogenous inflammatory factors, as well as NF-κB activation [38,39]. Therefore, we evaluated the effect of VRSE on LPS-stimulated phosphorylation of MAPK signaling and revealed that VRSE remarkably inhibited the phosphorylation of ERK, p38, and JNK in a concentration-dependent manner. These results demonstrated that the anti-neuroinflammatory effect of VRSE can inhibit several pro-inflammatory factors in the microglial BV2 cells by blocking NF-κB and MAPK signaling pathways. Therefore, the targeting inhibition of the overproduction of inflammatory factors is a valuable clinical approach for neurodegenerative disease treatment.
We performed phytochemical analyses using HPLC to investigate the relationships between the VRSE physiological activities and its components. Our result identified vitexin and isovitexin as the main VRSE component. Previous studies revealed that vitexin exerts neuroprotective effects against isoflurane-induced neurotoxicity by targeting the TRPV1 and NR2B signaling pathways in rats and PC12 cells [40]. Additionally, vitexin attenuates LPS-induced lung inflammation by activating the Nrf-2 pathway [41]. Further, isovitexin has neuroprotective activity against glutamate-induced cell damage in mice hippocampal slices [42]. Moreover, isovitexin exerts anti-inflammatory and antioxidant activities on LPS-induced lung injury by inhibiting MAPK and NF-κB and activating HO-1/Nrf-2 pathways [43]. Therefore, the neuroprotective and anti-neuroinflammatory effects of VRSE can reflect the presence of vitexin and isovitexin.
5. Conclusions
In conclusion, this study revealed that VRSE alleviated H2O2-induced cytotoxicity in HT22 cells via ROS accumulation and apoptosis inhibition. Additionally, VRSE exhibited neuroprotective effects through TrkB/CREB/BDNF pathway and Nrf-2/antioxidant enzyme activation. Further, we proved that VRSE ameliorated neuroinflammatory reactions in LPS-stimulated microglia BV2 cells through NF-κB and MAPK inhibitions. Moreover, the main components of VRSE, including vitexin and isovitexin, may be closely related to its neuroprotective and anti-neuroinflammatory effects. Therefore, VRSE may be a potential candidate for preventing and treating neurodegenerative diseases related to neurotoxicity, oxidative stress, and neuroinflammation.
Author Contributions
Conceptualization, J.Y.M.; Investigation, Y.H.J., Y.-C.O. and T.I.K.; Methodology, Y.H.J. and Y.-C.O.; Validation, Y.H.J., Y.-C.O. and T.I.K.; Writing—original draft, Y.H.J.; Writing—review and editing, Y.-C.O. and J.Y.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Korea Institute of Oriental Medicine (KIOM), provided by the Ministry of Science and ICT, Republic of Korea, grants number KSN2022230.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are contained within the article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Swerdlow, R.H.; Burns, J.M.; Khan, S.M. The Alzheimer’s Disease Mitochondrial Cascade Hypothesis. J. Alzheimers Dis. 2010, 20, S265–S279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, G.E.; Zhang, H. Abnormalities in oxidative processes in non-neuronal tissues from patients with Alzheimer’s disease. J. Alzheimers Dis. 2001, 3, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Ryter, S.W.; Kim, H.P.; Hoetzel, A.; Park, J.W.; Nakahira, K.; Wang, X.; Choi, A.M.K. Mechanisms of cell death in oxidative stress. Antioxid. Redox Signal. 2007, 9, 49–89. [Google Scholar] [CrossRef] [PubMed]
- Storr, S.J.; Woolston, C.M.; Zhang, Y.M.; Martin, S.G. Redox Environment, Free Radical, and Oxidative DNA Damage. Antioxid. Redox Signal. 2013, 18, 2399–2408. [Google Scholar] [CrossRef] [PubMed]
- Baillet, A.; Chanteperdrix, V.; Trocme, C.; Casez, P.; Garrel, C.; Besson, G. The Role of Oxidative Stress in Amyotrophic Lateral Sclerosis and Parkinson’s Disease. Neurochem. Res. 2010, 35, 1530–1537. [Google Scholar] [CrossRef]
- A Ma, Z. The role of peroxidation of mitochondrial membrane phospholipids in pancreatic β-cell failure. Curr. Diabetes Rev. 2012, 8, 69–75. [Google Scholar] [CrossRef]
- Walker, D.G.; Lue, L.F.; Beach, T.G. Gene expression profiling of amyloid beta peptide-stimulated human post-mortem brain microglia. Neurobiol. Aging 2001, 22, 957–966. [Google Scholar] [CrossRef]
- Glezer, I.; Simard, A.R.; Rivest, S. Neuroprotective role of the innate immune system by microglia. Neuroscience 2007, 147, 867–883. [Google Scholar] [CrossRef]
- Rossler, O.G.; Giehl, K.M.; Thiel, G. Neuroprotection of immortalized hippocampal neurones by brain-derived neurotrophic factor and raf-1 protein kinase: Role of extracellular signal-regulated protein kinase and phosphatidylinositol 3-kinase. J. Neurochem. 2004, 88, 1240–1252. [Google Scholar] [CrossRef]
- Baek, S.Y.; Kim, M.R. Neuroprotective Effect of Carotenoid-Rich Enteromorpha prolifera Extract via TrkB/Akt Pathway against Oxidative Stress in Hippocampal Neuronal Cells. Mar. Drugs 2020, 18, 372. [Google Scholar] [CrossRef]
- Johnson, J.A.; Johnson, D.A.; Kraft, A.D.; Calkins, M.J.; Jakel, R.J.; Vargas, M.R.; Chen, P.C. The Nrf2-ARE pathway: An indicator and modulator of oxidative stress in neurodegeneration. Ann. N. Y. Acad. Sci. 2008, 1147, 61–69. [Google Scholar] [CrossRef]
- Shin, S.K.; Yoo, J.M.; Li, F.Y.; Baek, S.Y.; Kim, M.R. Mulberry fruit improves memory in scopolamine-treated mice: Role of cholinergic function, antioxidant system, and TrkB/Akt signaling. Nutr. Neurosci. 2021, 24, 940–950. [Google Scholar] [CrossRef]
- Ji, J.F.; Ji, S.J.; Sun, R.; Li, K.; Zhang, Y.; Zhang, L.Y.; Tian, Y. Forced running exercise attenuates hippocampal neurogenesis impairment and the neurocognitive deficits induced by whole-brain irradiation via the BDNF-mediated pathway. Biochem. Biophys. Res. Commun. 2014, 443, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Gong, J.; Yoshida, T.; Eberhart, C.G.; Xu, Z.; Kombairaju, P.; Sporn, M.B.; Handa, J.T.; Duh, E.J. Nrf2 has a protective role against neuronal and capillary degeneration in retinal ischemia-reperfusion injury. Free Radic. Biol. Med. 2011, 51, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.G.; Zhang, G.D.; Shi, P.Q.; Du, B.S. Expression and antioxidation of Nrf2/ARE pathway in traumatic brain injury. Asian Pac. J. Trop. Med. 2013, 6, 305–310. [Google Scholar] [CrossRef]
- Trinh, K.; Andrews, L.; Krause, J.; Hanak, T.; Lee, D.; Gelb, M.; Pallanck, L. Decaffeinated coffee and nicotine-free tobacco provide neuroprotection in Drosophila models of Parkinson’s disease through an NRF2-dependent mechanism. J. Neurosci. 2010, 30, 5525–5532. [Google Scholar] [CrossRef] [Green Version]
- Houghton, C.A.; Fassett, R.G.; Coombes, J.S. Sulforaphane and other nutrigenomic Nrf2 activators: Can the Clinician’s expectation be matched by the reality? Oxid. Med. Cell. Longev. 2016, 2016, 7857186. [Google Scholar] [CrossRef] [Green Version]
- Crunkhorn, S. Deal watch: Abbott boosts investment in NRF2 activators for reducing oxidative stress. Nat. Rev. Drug Discov. 2012, 11, 96. [Google Scholar] [CrossRef]
- Niranjan, R. The role of inflammatory and oxidative stress mechanisms in the pathogenesis of Parkinson’s disease: Focus on astrocytes. Mol. Neurobiol. 2014, 49, 28–38. [Google Scholar] [CrossRef]
- Gilad, E.; Wong, H.R.; Zingarelli, B.; Virág, L.; O’Connor, M.; Salzman, A.L.; Szabó, C. Melatonin inhibits expression of the inducible isoform of nitric oxide synthase in murine macrophages: Role of inhibition of NFkappaB activation. FASEB J. 1998, 12, 685–693. [Google Scholar] [CrossRef]
- Zucoloto, A.Z.; Manchope, M.F.; Staurengo-Ferrari, L.; Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Saraiva, A.L.L.; Cecílio, N.T.; Alves-Filho, J.C.; Cunha, T.M.; Menezes, G.B.; et al. Probucol attenuates lipopolysaccharide-induced leukocyte recruitment and inflammatory hyperalgesia: Effect on NF-кB activation and cytokine production. Eur. J. Pharmacol. 2017, 809, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Chang, L.; Karin, M. Mammalian MAP kinase signalling cascades. Nature 2001, 410, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Kim, D.C.; Yoon, C.S.; Ko, W.M.; Lee, S.J.; Sohn, J.H.; Jang, J.H.; Ahn, J.S.; Kim, Y.C.; Oh, H. Anti-neuroinflammatory effects of citreohybridonol involving TLR4-MyD88-mediated inhibition of NF-small ka, CyrillicB and MAPK signaling pathways in lipopolysaccharide-stimulated BV2 cells. Neurochem. Int. 2016, 95, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Gan, R.Y.; Lui, W.Y.; Wu, K.; Chan, C.L.; Dai, S.H.; Sui, Z.Q.; Corke, H. Bioactive compounds and bioactivities of germinated edible seeds and sprouts: An updated review. Trends Food Sci. Technol. 2017, 59, 1–14. [Google Scholar] [CrossRef]
- Liyanage, R.; Kiramage, C.; Visvanathan, R.; Jayathilake, C.; Weththasinghe, P.; Bangamuwage, R.; Chaminda Jayawardana, B.; Vidanarachchi, J. Hypolipidemic and hypoglycemic potential of raw, boiled, and sprouted mung beans (Vigna radiata L. Wilczek) in rats. J. Food Biochem. 2018, 42, e12457. [Google Scholar] [CrossRef]
- Ali, N.M.; Mohd, Y.H.; Yeap, S.K.; Ho, W.Y.; Beh, B.K.; Long, K.; Koh, S.P.; Abdullah, M.P.; Alitheen, N.B. Anti-inflammatory and antinociceptive activities of untreated, germinated, and fermented mung bean aqueous extract. Evid. Based Complement. Altern. Med. 2014, 2014, 350507. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Srivastava, N.; Bhagyawant, S.S. Vicilin—A major storage protein of mungbean exhibits antioxidative potential, antiproliferative effects and ace inhibitory activity. PLoS ONE 2018, 13, e0191265. [Google Scholar] [CrossRef]
- Chai, W.M.; Wei, Q.M.; Deng, W.L.; Zheng, Y.L.; Chen, X.Y.; Huang, Q.; Ou-Yang, C.; Peng, Y.Y. Anti-melanogenesis properties of condensed tannins from Vigna angularis seeds with potent antioxidant and DNA damage protection activities. Food Funct. 2019, 10, 99–111. [Google Scholar] [CrossRef]
- Xie, J.; Du, M.; Shen, M.; Wu, T.; Lin, L. Physico-chemical properties, antioxidant activities and angiotensin-i converting enzyme inhibitory of protein hydrolysates from mung bean (Vigna radiate). Food Chem. 2019, 270, 243–250. [Google Scholar] [CrossRef]
- Lopes, L.A.R.; Martins, M.D.C.C.E.; Farias, L.M.; Brito, A.K.D.S.; Lima, G.M.; Carvalho, V.B.L.; Pereira, C.F.C.; Conde Júnior, A.M.; Saldanha, T.; Arêas, J.A.G.; et al. Cholesterol-lowering and liver-protective effects of cooked and germinated mung beans (Vigna radiata L.). Nutrients 2018, 10, 821. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Kim, T.I.; Oh, Y.C.; Ma, J.Y. Selaginella tamariscina Inhibits Glutamate-Induced Autophagic Cell Death by Activating the PI3K/AKT/mTOR Signaling Pathways. Int. J. Mol. Sci. 2022, 23, 11445. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Oh, Y.C.; Cho, W.K.; Yim, N.H.; Ma, J.Y. Anti-inflammatory effect of Rhapontici Radix ethanol extract via inhibition of NF-κB and MAPK and induction of HO-1 in macrophages. Mediat. Inflamm. 2016, 2016, 7216912. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.; Sagara, Y.; Liu, Y.; Maher, P.; Schubert, D. The regulation of reactive oxygen species production during programmed cell death. J. Cell Biol. 1998, 141, 1423–1432. [Google Scholar] [CrossRef] [Green Version]
- Seiler, M.; Schneider, H.; Forster, S.; Roth, E.K.; Wirth, C.; Culmsee, N.; Plesnila, E.; Kremmer, O.; Radmark, W.; Wurst, G.W.; et al. Glutathione peroxidase 4 senses and translates oxidative stress into 12/15-lipoxygenase dependent- and AIF-mediated cell death. Cell Metab. 2008, 8, 237–248. [Google Scholar] [CrossRef] [Green Version]
- Schipper, H.M.; Bennett, D.A.; Liberman, A.; Bienias, J.L.; Schneider, J.A.; Kelly, J.; Arvanitakis, Z. Glial heme oxygenase-1 expression in Alzheimer disease and mild cognitive impairment. Neurobiol. Aging 2006, 27, 252–261. [Google Scholar] [CrossRef]
- Wang, Y.; Santa–Cruz, K.; DeCarli, C.; Johnson, J.A. NAD (P) H: Quinone oxidoreductase activity is increased in hippocampal pyramidal neurons of patients with Alzheimer’s disease. Neurobiol. Aging 2000, 21, 525–531. [Google Scholar] [CrossRef]
- Brown, J.; Wang, H.; Hajishengallis, G.N.; Martin, M. TLR-signaling networks: An integration of adaptor molecules, kinases, and cross-talk. J. Dent. Res. 2011, 90, 417–427. [Google Scholar] [CrossRef]
- Kim, B.W.; Koppula, S.; Hong, S.S.; Jeon, S.B.; Kwon, J.H.; Hwang, B.Y.; Park, E.J.; Choi, D.K. Regulation of microglia activity by glaucocalyxin-A: Attenuation of lipopolysaccharide-stimulated neuroinflammation through NF-κB and p38 MAPK signaling pathways. PLoS ONE 2013, 8, e55792. [Google Scholar] [CrossRef]
- Kim, A.R.; Lee, M.S.; Choi, J.W.; Utsuki, T.; Kim, J.I.; Jang, B.C.; Kim, H.R. Phlorofucofuroeckol A suppresses expression of inducible nitric oxide synthase, cyclooxygenase-2, and proinflammatory cytokines via inhibition of nuclear factor-κB, c-Jun NH2-terminal kinases, and Akt in microglial cells. Inflammation 2013, 36, 259–271. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, B.; Shan, S.; Zhao, X. Neuroprotective effects of vitexin against isoflurane-induced neurotoxicity by targeting the TRPV1 and NR2B signaling pathways. Mol. Med. Rep. 2016, 14, 5607–5613. [Google Scholar] [CrossRef]
- Lu, Y.; Yu, T.; Liu, J.; Gu, L. Vitexin attenuates lipopolysaccharide-induced acute lung injury by controlling the Nrf2 pathway. PLoS ONE 2018, 13, e0196405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dos Santos, K.C.; Borges, T.V.; Olescowicz, G.; Ludka, F.K.; Santos, C.A.; Molz, S. Passiflora actinia hydroalcoholic extract and its major constituent, isovitexin, are neuroprotective against glutamate-induced cell damage in mice hippocampal slices. J. Pharm. Pharmacol. 2016, 68, 282–291. [Google Scholar] [CrossRef] [PubMed]
- Lv, H.; Yu, Z.; Zheng, Y.; Wang, L.; Qin, X.; Cheng, G.; Ci, X. Isovitexin Exerts Anti-Inflammatory and Anti-Oxidant Activities on Lipopolysaccharide-Induced Acute Lung Injury by Inhibiting MAPK and NF-κB and Activating HO-1/Nrf2 Pathways. Int. J. Biol. Sci. 2016, 12, 72–86. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on hydrogen peroxide (H2O2)-induced cytotoxicity in HT22 cells. (A) HT22 cells were incubated with VRSE concentrations of 10, 50, 100, or 200 μg/mL. (B,C) After VRSE pretreatment concentrations of 10, 50, 100, or 200 μg/mL, HT22 cells were stimulated with 500 μM of H2O2. (C) Images represent the three independent experiments at a magnification of 400×. Scale bar = 30 μm. Control cells were incubated with the vehicle alone. Data are presented as mean ± standard error of the mean of three independent experiments. Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, and † p < 0.001 (vs. H2O2).
Figure 1.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on hydrogen peroxide (H2O2)-induced cytotoxicity in HT22 cells. (A) HT22 cells were incubated with VRSE concentrations of 10, 50, 100, or 200 μg/mL. (B,C) After VRSE pretreatment concentrations of 10, 50, 100, or 200 μg/mL, HT22 cells were stimulated with 500 μM of H2O2. (C) Images represent the three independent experiments at a magnification of 400×. Scale bar = 30 μm. Control cells were incubated with the vehicle alone. Data are presented as mean ± standard error of the mean of three independent experiments. Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, and † p < 0.001 (vs. H2O2).

Figure 2.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) against hydrogen peroxide (H2O2)-induced intracellular reactive oxygen species (ROS) production in HT22 cells. Cells were pretreated with VRSE concentrations of 10, 50, 100, or 200 μg/mL and then with 500 μM of H2O2. H2DCFDA (20 μM) is an oxidation-sensitive fluorescence dye used to assess ROS levels. The ROS expression was determined using a fluorescence microscope and a fluorescence microplate reader. Scale bar = 200 μm. Control cells were incubated with the vehicle alone. All experiments were repeated at least three times, and similar results were obtained. Data are presented as mean ± standard error of the mean. Con, control. Statistical significance was defined as # p < 0.05 (vs. control) and † p < 0.001 (vs. H2O2).
Figure 2.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) against hydrogen peroxide (H2O2)-induced intracellular reactive oxygen species (ROS) production in HT22 cells. Cells were pretreated with VRSE concentrations of 10, 50, 100, or 200 μg/mL and then with 500 μM of H2O2. H2DCFDA (20 μM) is an oxidation-sensitive fluorescence dye used to assess ROS levels. The ROS expression was determined using a fluorescence microscope and a fluorescence microplate reader. Scale bar = 200 μm. Control cells were incubated with the vehicle alone. All experiments were repeated at least three times, and similar results were obtained. Data are presented as mean ± standard error of the mean. Con, control. Statistical significance was defined as # p < 0.05 (vs. control) and † p < 0.001 (vs. H2O2).

Figure 3.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) against hydrogen peroxide (H2O2)-induced apoptosis in HT22 cells. Cells were pretreated with VRSE concentrations of 50, 100, or 200 μg/mL and then exposed to 500 μM of H2O2. The expression levels of Bcl-2-associated X, B-cell lymphoma 2, apoptosis-inducing factor, caspase-3, and poly (ADP-ribose) polymerase were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images represent three independent experiments. Data are presented as mean ± standard error of the mean. BAX, Bcl-2-associated X; Bcl-2, B-cell lymphoma 2; AIF, apoptosis-inducing factor; Cas, caspase; PARP, poly (ADP-ribose) polymerase; Con, control. Statistical significance was defined as # p < 0.05 (vs. control), ** p < 0.01, and † p < 0.001 (vs. H2O2).
Figure 3.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) against hydrogen peroxide (H2O2)-induced apoptosis in HT22 cells. Cells were pretreated with VRSE concentrations of 50, 100, or 200 μg/mL and then exposed to 500 μM of H2O2. The expression levels of Bcl-2-associated X, B-cell lymphoma 2, apoptosis-inducing factor, caspase-3, and poly (ADP-ribose) polymerase were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images represent three independent experiments. Data are presented as mean ± standard error of the mean. BAX, Bcl-2-associated X; Bcl-2, B-cell lymphoma 2; AIF, apoptosis-inducing factor; Cas, caspase; PARP, poly (ADP-ribose) polymerase; Con, control. Statistical significance was defined as # p < 0.05 (vs. control), ** p < 0.01, and † p < 0.001 (vs. H2O2).

Figure 4.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on (A) microglia viability and (B,C) inflammatory cytokine secretion in BV2 cells. (A) BV2 cells were incubated with VRSE concentrations of 10, 50, 100, or 200 μg/mL. (B,C) After VRSE pretreatment concentrations of 10, 50, 100, or 200 μg/mL, BV2 cells were stimulated with 100 ng/mL of lipopolysaccharide. Control cells were incubated with the vehicle alone. Data are presented as mean ± standard error of the mean of three independent experiments. Con, control; LPS, lipopolysaccharide; TNF, tumor necrosis factor; IL, interleukin. Statistical significance was defined as # p < 0.05 (vs. control), ** p < 0.01, and † p < 0.001 (vs. LPS).
Figure 4.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on (A) microglia viability and (B,C) inflammatory cytokine secretion in BV2 cells. (A) BV2 cells were incubated with VRSE concentrations of 10, 50, 100, or 200 μg/mL. (B,C) After VRSE pretreatment concentrations of 10, 50, 100, or 200 μg/mL, BV2 cells were stimulated with 100 ng/mL of lipopolysaccharide. Control cells were incubated with the vehicle alone. Data are presented as mean ± standard error of the mean of three independent experiments. Con, control; LPS, lipopolysaccharide; TNF, tumor necrosis factor; IL, interleukin. Statistical significance was defined as # p < 0.05 (vs. control), ** p < 0.01, and † p < 0.001 (vs. LPS).

Figure 5.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on (A) the nuclear translocation of nuclear factor-κB p65 and (B) phosphorylation of mitogen-activated protein kinases. After VRSE pretreatment concentrations of 50, 100, or 200 μg/mL, BV2 cells were stimulated with 100 ng/mL of lipopolysaccharide. Control cells were incubated with the vehicle alone. Blot images represent three independent experiments. Data are presented as mean ± standard error of the mean. NF, nuclear factor; Con, control; LPS, lipopolysaccharide; ERK, extracellular signal-regulated kinase; JNK, c-Jun NH2-terminal kinase. Statistical significance was defined as # p < 0.05 (vs. control) and † p < 0.001 (vs. LPS).
Figure 5.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on (A) the nuclear translocation of nuclear factor-κB p65 and (B) phosphorylation of mitogen-activated protein kinases. After VRSE pretreatment concentrations of 50, 100, or 200 μg/mL, BV2 cells were stimulated with 100 ng/mL of lipopolysaccharide. Control cells were incubated with the vehicle alone. Blot images represent three independent experiments. Data are presented as mean ± standard error of the mean. NF, nuclear factor; Con, control; LPS, lipopolysaccharide; ERK, extracellular signal-regulated kinase; JNK, c-Jun NH2-terminal kinase. Statistical significance was defined as # p < 0.05 (vs. control) and † p < 0.001 (vs. LPS).

Figure 6.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on the activation of nuclear factor erythroid 2-related factor 2 (Nrf-2) and antioxidant enzymes in hydrogen peroxide (H2O2)-exposed HT22 cells. Cells were incubated with 500 μM of H2O2 with or without VRSE. The expression levels of Nrf-2, heme oxygenase-1, NAD(P)H quinone oxidoreductase 1, and glutamate-cysteine ligase catalytic subunit were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images were representative of the three independent experiments and data are presented as mean ± standard error of the mean. HO, heme oxygenase; NQO1, NAD(P)H quinone oxidoreductase 1; GCLC, glutamate-cysteine ligase catalytic subunit; Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, ** p < 0.01, and † p < 0.001 (vs. H2O2).
Figure 6.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on the activation of nuclear factor erythroid 2-related factor 2 (Nrf-2) and antioxidant enzymes in hydrogen peroxide (H2O2)-exposed HT22 cells. Cells were incubated with 500 μM of H2O2 with or without VRSE. The expression levels of Nrf-2, heme oxygenase-1, NAD(P)H quinone oxidoreductase 1, and glutamate-cysteine ligase catalytic subunit were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images were representative of the three independent experiments and data are presented as mean ± standard error of the mean. HO, heme oxygenase; NQO1, NAD(P)H quinone oxidoreductase 1; GCLC, glutamate-cysteine ligase catalytic subunit; Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, ** p < 0.01, and † p < 0.001 (vs. H2O2).

Figure 7.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on the phosphorylation of tyrosine receptor kinase B (TrkB), protein kinase B (Akt), and cAMP response element-binding protein (CREB), and the expression of mature brain-derived neurotrophic factor (BDNF) in hydrogen peroxide (H2O2)-exposed HT22 cells. Cells were pretreated with VRSE at concentrations of 50, 100, or 200 μg/mL and then exposed to H2O2 (500 μM). The expression levels of TrkB, Akt, CREB, and BDNF were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images are representative of the three independent experiments and data are expressed as mean ± standard error of the mean. Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, ** p < 0.01, and † p < 0.001 (vs. H2O2).
Figure 7.
Effects of Vignae Radiatae Semen ethanol extract (VRSE) on the phosphorylation of tyrosine receptor kinase B (TrkB), protein kinase B (Akt), and cAMP response element-binding protein (CREB), and the expression of mature brain-derived neurotrophic factor (BDNF) in hydrogen peroxide (H2O2)-exposed HT22 cells. Cells were pretreated with VRSE at concentrations of 50, 100, or 200 μg/mL and then exposed to H2O2 (500 μM). The expression levels of TrkB, Akt, CREB, and BDNF were determined via Western blot analysis. Control cells were incubated with the vehicle alone. Blot images are representative of the three independent experiments and data are expressed as mean ± standard error of the mean. Con, control. Statistical significance was defined as # p < 0.05 (vs. control), * p < 0.05, ** p < 0.01, and † p < 0.001 (vs. H2O2).

Figure 8.
HPLC chromatogram of Vignae Radiatae Semen ethanol extract (VRSE). (A) Chemical structure of vitexin and isovitexin. (B) HPLC chromatogram of standard solution and VRSE. (C) Regression equations, the limit of detection, and the limit of quantitation of standard compound. STD, standard; LOD, the limit of detection; LOQ, the limit of quantitation.
Figure 8.
HPLC chromatogram of Vignae Radiatae Semen ethanol extract (VRSE). (A) Chemical structure of vitexin and isovitexin. (B) HPLC chromatogram of standard solution and VRSE. (C) Regression equations, the limit of detection, and the limit of quantitation of standard compound. STD, standard; LOD, the limit of detection; LOQ, the limit of quantitation.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Primary and secondary antibodies use for Western blot analysis.
| Antibody | Corporation | Product No. | RRID | Dilution Rate |
|---|---|---|---|---|
| BAX | Cell Signaling | #2772 | AB_10695870 | 1:1000 |
| Bcl-2 | Cell Signaling | #3498 | AB_1903907 | 1:1000 |
| AIF | Cell Signaling | #4642 | AB_2224542 | 1:1000 |
| C-CAS-3 | Cell Signaling | #9664 | AB_2070042 | 1:1000 |
| CAS-3 | Cell Signaling | #9662 | AB_331439 | 1:1000 |
| C-PARP | Cell Signaling | #9548 | AB_2160592 | 1:1000 |
| PARP | Cell Signaling | #9532 | AB_659884 | 1:1000 |
| β-actin | Cell Signaling | #4970 | AB_2223172 | 1:1000 |
| NF-κB p65 | Cell Signaling | #8242 | AB_10859369 | 1:1000 |
| Lamin B1 | Cell Signaling | #13435 | AB_2737428 | 1:1000 |
| P-ERK | Cell Signaling | #4377 | AB_331775 | 1:1000 |
| ERK | Cell Signaling | #9102 | AB_330744 | 1:1000 |
| P-p38 | Cell Signaling | #9211 | AB_331641 | 1:1000 |
| P38 | Cell Signaling | #9212 | AB_330713 | 1:1000 |
| P-JNK | Cell Signaling | #9251 | AB_331659 | 1:1000 |
| JNK | Cell Signaling | #9252 | AB_2250373 | 1:1000 |
| Nrf-2 | Cell Signaling | #12721 | AB_2715528 | 1:1000 |
| HO-1 | Cell Signaling | #82206 | AB_2799989 | 1:1000 |
| NQO1 | Santa Cruz | #SC-32793 | AB_628036 | 1:1000 |
| GCLC | Thermo Fisher | #PA5-87854 | AB_2804457 | 1:1000 |
| P-TrkB | Cell Signaling | #4621 | AB_916186 | 1:1000 |
| TrkB | Cell Signaling | #4603 | AB_2155125 | 1:1000 |
| P-Akt | Cell Signaling | #4060 | AB_2315049 | 1:1000 |
| Akt | Cell Signaling | #4691 | AB_915783 | 1:1000 |
| P-CREB | Cell Signaling | #9191 | AB_331606 | 1:1000 |
| CREB | Cell Signaling | #9197 | AB_331277 | 1:1000 |
| BDNF | Cell Signaling | #47808 | AB_2894709 | 1:1000 |
| 2nd anti-mouse | Cell Signaling | #7076 | AB_330924 | 1:5000 |
| 2nd anti-rabbit | Cell Signaling | #7074 | AB_2099233 | 1:5000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jeong, Y.H.; Oh, Y.-C.; Kim, T.I.; Ma, J.Y. Neuroprotective and Anti-Neuroinflammatory Properties of Vignae Radiatae Semen in Neuronal HT22 and Microglial BV2 Cell Lines. Nutrients 2022, 14, 5265. https://doi.org/10.3390/nu14245265
AMA Style
Jeong YH, Oh Y-C, Kim TI, Ma JY. Neuroprotective and Anti-Neuroinflammatory Properties of Vignae Radiatae Semen in Neuronal HT22 and Microglial BV2 Cell Lines. Nutrients. 2022; 14(24):5265. https://doi.org/10.3390/nu14245265
Chicago/Turabian StyleJeong, Yun Hee, You-Chang Oh, Tae In Kim, and Jin Yeul Ma. 2022. "Neuroprotective and Anti-Neuroinflammatory Properties of Vignae Radiatae Semen in Neuronal HT22 and Microglial BV2 Cell Lines" Nutrients 14, no. 24: 5265. https://doi.org/10.3390/nu14245265
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.