Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease


1
College of Marine Science and Biological Engineering, Qingdao University of Science and Technology, Qingdao 266042, China
2
Department of Oncology, Qingdao Central Hospital, Siliu South Road No.127, Qingdao 266042, China
*
Author to whom correspondence should be addressed.
Nutrients 2022, 14(9), 1977; https://doi.org/10.3390/nu14091977
Submission received: 23 April 2022
/
Revised: 1 May 2022
/
Accepted: 6 May 2022
/
Published: 9 May 2022
(This article belongs to the Section Prebiotics and Probiotics)
Abstract
:Intestinal microbiota has its role as an important component of human physiology. It produces metabolites that module key functions to establish a symbiotic crosstalk with their host. Among them, short chain fatty acids (SCFAs), produced by intestinal bacteria during the fermentation of partially and non-digestible polysaccharides, play key roles in regulating colon physiology and changing intestinal environment. Recent research has found that SCFAs not only influence the signal transduction pathway in the gut, but they also reach tissues and organs outside of the gut, through their circulation in the blood. Growing evidence highlights the importance of SCFAs level in influencing health maintenance and disease development. SCFAs are probably involved in the management of host health in a complicated (positive or negative) way. Here, we review the current understanding of SCFAs effects on host physiology and discuss the potential prevention and therapeutics of SCFAs in a variety of disorders. It provides a systematic theoretical basis for the study of mechanisms and precise intake level of SCFAs to promote human health.
1. Introduction
The human intestine harbors trillions of bacteria, fungi, archaea, and viruses, which are often collectively referred to as “microbiota”. The intestinal microbiota has been considered as a fundamental player orchestrating host pathology and physiology [1]. Imbalance of intestinal microbiota is closely associated with diseases ranging from localized gastroenterological disorders to neurologic, respiratory, metabolic, hepatic, and cardiovascular illnesses [2]. Intestinal microbiota influences our health in multiple ways. An increasing body of evidence verifies that microbial metabolites have important effects on host physiology. Short chain fatty acids (SCFAs) are produced by intestinal microbes during the fermentation of partially and non-digestible polysaccharides. As the key player in the interplay between diet, microbiota, and health, SCFAs confer some degree of protection/destruction through a variety of pathways in a wide range of disorders. The purpose of this paper is to present a state-of-the-art review of our understanding of the role played by SCFAs in human health and disease and outline, as a result, possible further avenues of development of the field (Figure 1 and Figure 2).
2. SCFAs—The What, Where, and How
SCFAs are important food ingredients with 2-6 carbon atom chains that are abundant in nature, mostly in the form of triglycerides in plant oils and milk [3]. Nevertheless, the main source of SCFAs in humans is microbial fermentation in the proximal intestines (jejunum, ileum, and proximal colon) and the distal intestines (descending colon, sigmoid and rectum) during the breakdown of undigested food fibers, as well as peptides and proteins [4,5,6]. Acetate, propionate, and butyrate account for more than 95% of SCFAs at an estimated ratio of about 3:1:1 in the gut [7,8]. The proportion can vary depending on diet, microbiota, host genotype, and site of fermentation [9]. These SCFAs range from 20–140 mM in the gut, with very high concentrations in the proximal colon (70–140 mM) and relatively low concentrations in the distal colon (20–70 mM) and in the distal ileum (20–40 mM) (Table 1) [7,10]. Propionate and butyrate are released in the distal intestines and are higher than in the proximal intestines. Acetate is released only in the distal intestines [6]. It could be ascribed to several reasons. One explanation is that it could metabolize a relatively higher proportion of SCFAs in the proximal intestinal mucosa [11]. Another explanation is that the composition and activity of intestinal microbiota differ from one bowel to the next, resulting in variable SCFAs production [12]. The capacity of intestinal epithelium cells (IECs) to absorb and transport SCFAs differs between the proximal and the distal intestines. It also suggests that SCFAs have a function in regulating health in the colon epithelium and immune cells on a local level [13]. Furthermore, the majority of SCFAs are utilized within the gut; however, a small proportion of SCFAs reaches the liver where can be used as substrates for energy-producing and metabolized to generate glucose.
High throughput sequencing technology has made great advances in characterization of microbes responsible for SCFAs production. SCFAs are mainly mediated by specific intestinal microbes from food (Table 2). Acetate producing pathways are widely distributed among microbial groups. For example, acetate can be produced by enteric bacteria and acetogens (Blautia hydrogenotrophica) via the acetyl-CoA and Wood-Ljungdahl pathways, respectively. However, pathways for propionate and butyrate are substrate specific and highly conserved. For example, propionate is mainly generated by two pathways: Firmicutes lactate pathway and Bacteroidetes succinate pathway [14]. Propionate production is dominated by relatively few bacterial genera. Akkermansia municiphilla has been identified as key propionate producing mucin degrading species. Butyrate is produced by Firmicutes, such as Faecalibacterium prausnitzii, Eubacterium rectale, Eubacterium hallii, and Ruminococcus bromii [15]. Ruminococcus bromii may ferment resistant starch to produce butyrate. Recently, there is emerging evidence that diet-driven changes in microbial diversity led to variations in SCFAs; however, these changes into long-term effects on host health need more intervention studies. In addition, changes in intestinal microbiota with patients have been linked with changed intestinal microbiota and a loss of SCFAs producing organisms. Therefore, it highlights that diet and intestinal microbiota have profound effects on SCFAs [16].
A fraction of SCFAs can directly across the epithelial barrier; however, most exists require specialized transporter for their uptake. Therefore, the majority of SCFAs across the mucosa need active transport delivered by two receptors: the monocarboxylate transporter 1 (MCT-1) and the sodium-coupled monocarboxylate transporter 1 (SMCT-1). MCT-1 and SMCT-1 are highly expressed on colonocytes and along the overall gastrointestinal tract. Acetate, propionate, and butyrate can be transported from the gut into lacteal lymphatic system and/or hepatic portal circulation, with total concentrations at about 79 μM, 148 μM, and 375 μM in peripheral, portal, and hepatic blood, respectively [7] (Table 1). Propionate and butyrate, metabolized by hepatocytes, have the concentrations at 1-15 μM in the systemic circulation, with acetate appearing between 100-200 μM [17] (Table 1). Acetate is either released into the peripheral venous system or remains in the liver [18]. As a result, only acetate could be detected in peripheral blood (Table 1). Butyrate, in particular, has more beneficial effects than other SCFAs, but it is difficult to deliver into the colon due to short half-life and low compliance rate with rectal administration [19]. It has the potential to locate and deliver butyrate-producing intestinal bacteria for solving this problem and improving our health.

{kind=link}
{kind=link}
Table 2.
The production, absorption, and transport receptors of major SCFAs.
| SCFAs | Related Microbes | Absorption Site | G protein-Coupled Receptors |
|---|---|---|---|
| Acetate | Enteric bacteria and acetogens (Blautia hydrogenotrophica) [14] | Liver or peripheral venous system | GPR41, GPR43 [20,21] |
| Propionate | Firmicutes and Bacteroidetes [14] | Hepatocytes | GPR41, GPR43 [20,21] |
| Butyrate | Faecalibacterium prausnitzii, Eubacterium rectale, Eubacterium hallii, Ruminococcus bromii [15] | Colonocytes | GPR109A, GPR41, GPR43 [20,21,22] |
3. SCFAs-Signaling Mechanisms
SCFAs have effects on biological responses based on two major signaling mechanisms (Figure 1). The first is that histone deacetylases (HDACs) influence gene expression by the direct inhibition, which is characteristic of propionate and butyrate. The second for SCFAs effects involves the signaling through G protein-coupled receptors (GPCRs), including GPR109A, GPR41, and GPR43 [23]. GPR43 expression has been identified along the overall gastrointestinal tract, including those cells of the immune system. In the immune system, GPR43 and GPR109A are expressed on neutrophils, macrophages, and dendritic cells (DCs), suggesting SCFAs’ roles in immune responses [24,25]. GPR43 with an anti-inflammatory role was also found in colitis and arthritis models [26]. GPR41 and GRP109A expressions activated by SCFAs were confirmed to have beneficial effects on body weight [27]. GPR43 and GPR41, sharing 43% amino acid identity, could bind to acetate, propionate, and butyrate [20] (Table 2). Among these SCFAs, butyrate is the most potent agonist for GPR109A, which is only expressed in humans, and propionate was more selective for GPR41 and GPR43 [22] (Table 2). There receptors could transmit signaling based on the different cell types and ligands. Therefore, we consider that it is valuable to develop certain drugs that target these receptors or their signaling pathways to influence SCFAs for disease treatment.
4. SCFAs-Host Physiology
Usually, SCFAs may reduce proliferation and migration of immune cells, lower cytokine levels, and induce apoptosis to inhibit inflammation. However, marked changes of SCFAs concentrations in blood or tissues could cause inflammatory, immunological, and metabolic diseases [28]. Appropriate concentrations of SCFAs help to maintain normal metabolism in the prevention and treatment of disease [29]. Notably, due to insufficient daily dietary fiber intake and low abundance of butyrate-producing intestinal microbiota, SCFAs concentrations are probably less than optimal in older people [30]. Therefore, SCFAs are identified as important metabolic biomarkers of disease-related changes [31]. SCFAs not only play a role in intestinal homeostasis, but also are transported across the gut to enter the blood circulation and influence the function and metabolism of peripheral tissues. Recent research suggests that SCFAs have impacts in the disease etiology, correlating systemic effects with diet and intestinal microbiota. The current knowledge and therapeutic interest of SCFAs on some organs and tissues in and beyond the gut are outlined in the following text (Figure 2). Examples of SCFAs involved in various diseases in vivo and in vitro studies were listed in Table 3, which were obtained by searching for “(short chain fatty acid [Title/Abstract]) AND (Inflammatory bowel disease/cancer/obesity/diabetes/kidney/hypertension [Title/Abstract])” on PubMed from 2000 to 2020.
4.1. Inflammatory Bowel Disease
Intestinal dysbiosis is identified in individuals with inflammatory bowel disease (IBD), with a decreased number of SCFAs-producing bacteria and a lower concentration of SCFAs in feces [54]. It has been reported that European children are susceptible to develop IBD than African children, due to decreased microbial diversity and a lack of SCFAs production [55]. Patients with ulcerative colitis have the intestinal dysbiosis, characterized by a reduction in butyrate-producing bacteria Eaecalibacterium prausnitizii and Roseburia hominis [56]. The administration of probiotic bacteria producing SCFAs and SCFAs supplements have been shown to reduce the pro-inflammatory microenvironment in animal models of IBD and human colitis [57]. SCFAs protect against IBD-related intestinal inflammation by immune regulation and intestinal epithelial barrier maintenance. SCFAs could bind to GPR43 and GPR109A, which are important for regulation of intestinal immunity, to trigger the development of regulatory T cell (Treg) [37]. SCFAs could protect mice against colitis by modulating colonic Treg number and activity in a GPR43-dependent manner [58], because GPR43-deficient (Gpr43(−/−)) mice showed unresolving or exacerbated inflammation in models of colitis [26]. SCFAs could change pro-inflammatory cytokine production in IECs via enhancing nuclear factor-kappa B (NF-κB) activation in toll-like receptor (TLR) ligand-responses [59]. SCFAs could suppress intestinal stem cell proliferation, regulate tight junction protein expression, and promote crypt IECs differentiation, all of which can influence IBD by playing a direct anti-inflammatory role in IECs [32,33]. SCFAs, particularly butyrate, could enhance interleukin-22 (IL-22) production in CD4+T cells and ILCs to alleviate colitis in mice by activating GPR41 and inhibiting HDACs [34]. Although it has not been made a standard protocol due to patients’ compliance and partial restricted indications, the administration of SCFAs or prebiotics that facilitate SCFAs production is increasingly used as an adjuvant therapy to improve the efficacy of conventional treatment such as corticosteroid and 5-aminosalicylic acid therapy.
4.2. Cancer
SCFAs have the ability to directly influence colonocyte proliferation and differentiation to help to prevent colonic cancer [60]. Patients with colorectal cancer have lower level of butyrate-producing bacteria and SCFAs [61]. SCFAs influence colon cancer partially in a GRCR-dependent way. The expression of GPR43 has been shown to be decreased in many colon cancer patients. Butyrate and propionate could inhibit colon cancer cell growth and cause apoptosis by activating the GPR43 pathway [36]. Inflammation-induced colon carcinogenesis is accelerated when GPR109A level is low. Butyrate with GPR109A could stimulate colonic DCs and macrophages to boost Treg number by raising the production of IL-10 and Aldh1a [37]. The mice of Niacr1−/−, encoding GPR109A, are inclined to suffer from development of colon cancer [37]. Niacin, a pharmacological GPR109A agonist, suppresses colon cancer in a GPR109A-dependent manner [37]. Butyrate acts as a potent HDAC inhibitor in colon cancer cells, enabling cell cycle arrest, differentiation, and apoptosis at physiological concentrations [62].
Recently, a substantial impact of SCFAs in affecting immunotherapy in patients with non-small cell lung cancer (NSCLC) has been considered to be a promising strategy for personalized management by identifying early progressor and long responder patients [63]. According to preliminary findings, propionate and butyrate were shown to be significantly related with long-term beneficial effects in NSCLC patients treated with nivolumab in second-line treatment [63]. However, another research reported that high blood butyrate and propionate levels restrained the higher proportion of Treg (CD80/CD86/CD4) induced by anti-CTLA-4 in melanoma mice and patients with ipilimumab therapy [64].
4.3. Obesity
Obesity and overweight are complex health concerns of multifactorial etiology that have a high risk of causing other chronic diseases, such as type 2 diabetes (T2D), insulin resistance, fatty liver disease, cardiovascular disease, and others [65]. Since the 1940s, the role of SCFAs in adiposity has been studied in human, as well as in vitro and in vivo animals [66,67]. Changes in the level of SCFAs produced by intestinal microbiota have contributed to the development of obesity. Several animal studies have indicated that SCFAs administration could reverse or reduce weight and adiposity growth [68,69]. In obese mice, treatment with sodium butyrate leads to weight loss by boosting energy expenditure and fat oxidation [42]. In the presence of continuous food intake and physical activity, oral administration of acetate, propionate, and butyrate to mice fed a high-fat diet results in body weight reduction and insulin sensitivity augment [39]. According to a study involving Mexican children, excess weight and obesity were shown to have lower concentrations of fecal propionate and butyrate than normal children [40]. In contrast, obesity individuals have been linked to higher fecal concentrations of SCFAs, particularly propionate, than lean individuals [8,41]. Another study found that overweight and obese people in the Netherlands had higher fecal SCFAs concentrations than lean ones, indicating that they had a higher microbial energy harvest [41]. These findings are debatable. One reason is that SCFAs produced by intestinal microbial fermentation account for 5-10% of daily energy intake [70]. Another reason is probably that fluctuations in SCFA concentrations in the feces are caused by a general dysbiosis in intestinal microbial community, leading to increased or decreased production or mucosal absorption.
4.4. Diabetes
SCFAs exhibit the correlative beneficial effects in glucose homeostasis. SCFAs have the capacity to enhance glucose-stimulated insulin secretion and control glucose based on the SCFAs-GPR axis [45,71]. Propionate could activate ileal mucosal GPR43 to reduce hepatic glucose production in healthy rats in vivo [44]. Propionate potentiated glucose-stimulated insulin release in vivo and in vitro studies [45]. Marino et al. found that non-obese diabetic (NOD) model mice, which were given a combination of acetate and butyrate by drinking water, had suffered less from type 1 diabetes (T1D) [43]. Moreover, acetate and butyrate improved intestinal barrier function and increased the number of Bacteroides species in NOD model mice, which helped to inhibit T1D [43]. Zhao et al. suggested that T2D could be a consequence of deficiency in SCFAs produced by carbohydrate fermentation in the gut [72]. However, researchers also discovered that mice exposed to more than 150 mM SCFAs developed urethritis and hydronephrosis, as well as alterations in lymphocytes [73]. The causes need further investigation.
4.5. Kidney Diseases
Kidney diseases are partly developed by inflammatory processes, which have effects on the physiological response to renal infection and injury and contribute to the development of possibly irreversible kidney damage [74,75]. Here, acute kidney injury (ACI) and chronic kidney diseases (CDK) are the key topics. Growing evidence has highlighted that SCFAs play a favorable role in kidney diseases in both experimental animals and patients. Treating with acetate or acetate-producing bacteria could alleviate ACI in the ischemia-reperfusion injury model [46]. The cause could be the function of SCFAs in inhibiting inflammatory, reactive oxygen species, apoptosis, and chromatin modification [46]. Long-term oral administration of sodium butyrate attenuated ACI via improving the activity of renal antioxidant enzymes in an animal experimental model [47]. Increased dietary fiber significantly increased intestinal epithelial tight junctions, reduced oxidative stress and inflammation, and slowed severe renal dysfunction in CDK rats [76]. Furthermore, providing butyrate post-treatment to juvenile diabetic rats showed that SCFAs reduced renal histological alterations, DNA damage, and apoptosis in addition to lowering creatinine, plasma glucose, and urea [48]. However, there are still a number of unfavorable findings between SCFAs and CDK. A study found that oral dosages of SCFAs higher than physiological levels in mice produced Th1 and Th17 cells to induce inflammation and trigger kidney hydronephrosis [73]. Mice on a high-fiber diet had higher gut butyrate levels but were more susceptible to E. coli infection [49]. The underlying mechanism is still not clear. Furthermore, individuals with kidney diseases had the reduced bacteria that possessed SCFAs-forming enzymes compared to control groups [77]. Reduced dietary fiber intake was strongly associated with a reduction in the population of butyrate-forming bacteria in individuals with end-stage renal disease, and following dietary changes, the pathology ameliorated [77]. The effects of SCFAs on kidney diseases are mainly by decreasing inflammation and increasing antioxidant activity. However, SCFAs have only indirect favorable effects on remission of kidney diseases in human. These studies are either preliminary or controversial. More research is needed to fully comprehend the direct effects on kidney diseases, as well as better understand these mechanisms and their implications.
4.6. Hypertension
Hypertension is involved in a metabolic disorder. By activating T cells and macrophages with angiotensin II (AngII), hypertension could cause organ damage by infiltrating target organs such as the vasculature and the heart [78]. The organ damage caused by hypertension depends on the pro- and anti-inflammatory immune cells, as well as the hemodynamic load [52]. Intestinal microbiota and their metabolites SCFAs were supposed to be associated with hypertension [79]. Lactobacillus was decreased and Th17 cells were increased in a hypertensive mice model fed a high salt diet. After Lactobacillus supplementation, the number of Th17 cells and systolic blood pressure were both reduced [80]. SCFAs influence hypertension via interacting with GPCR receptors in the vascular tissues [51]. Propionate supplement caused the reduction of blood pressure in the Olfr78 gene knockout mice model, probably due to the GPR41 receptor activated in the vascular endothelium [51]. Bartolomaeus et al. discovered that propionate intervention could protect from hypertensive damage by mitigating systemic inflammation based on the reduction of Treg in wild-type NMRI and apolipoprotein E knockout-deficient mice model [52]. Butyrate supplement was reported to ameliorate endothelium-dependent vasodilation in a hypertension mouse model induced by chronic angiotensin II infusion [53]. Ganesh et al. identified that acetate was a key player in a hypertensive rat model of obstructive sleep apnea [50]. Clinical evidence supported the idea that microbially mediated SCFAs had anti-hypertensive properties, and that a high intake of fruits and vegetables lowered the incidence of hypertension [81].
4.7. Liver Diseases
The alleviation effects of SCFAs on liver diseases have been well established in both animal and human investigations. In animal studies, sucralose intake in maternal mice reduced SCFA-producing bacteria and diminished cecal butyrate production in offspring, causing hepatic steatosis to exacerbate in adulthood [82]. Both sodium acetate and sodium butyrate supplementation protected against nicotine-induced excess hepatic steatosis and western-style diet-induced non-alcoholic steatohepatitis (NASH) [83,84]. In human investigations, SCFAs were shown to be lower in non-obese non-alcoholic fatty liver disease (NAFLD) patients than in non-obese healthy people [85].
SCFAs contribute to prevent the development of NAFLD through a variety of mechanisms. The first is that SCFAs have key effects on fatty acid metabolism and visceral adipose tissue (VAT), both of which play a vital role in the development of NAFLD. Excess accumulation of VAT could increase the release of free fatty acids (FFAs) into liver [86]. The FFAs are considered as a key role in the development of NAFLD because they induce hepatic TNF expression via activating NF-κB [87]. In addition, VAT causes an imbalance of pro-inflammatory and anti-inflammatory adipokines, resulting in systematic inflammation, including the liver inflammation. In human multipotent adipose tissue-derived stem adipocytes, acetate could attenuate hormone-sensitive lipase (HSL) phosphorylation in a Gi-coupled way [88]. In rabbits, acetate could retrain lipid accumulation, increasing lipolysis and fatty acid oxidation while retraining fatty acid synthesis [89]. The second is that SCFAs could increase energy harvest, improve nutrient absorption, and enhance hepatic lipogenesis by regulating intestinal mobility. The primary mechanism is that activation of GPR41 and GPR43 promotes secretion of 5-hydroxytryptamine, peptide-YY, and glucagon-like peptide-1, which could impede intestinal transit and reduce gastric emptying, food intake, and intestinal motility [90]. The third is that SCFAs could protect the intestinal barrier. For example, acetate, propionate, and butyrate could stimulate intestinal Nod-like receptor family pyrin domain containing 3 (NLRP3) inflammasome to increase IL-18 secretion further to ameliorate intestinal barrier integrity [91]. The fourth is that SCFAs could come into the liver directly via the portal vein, inhibiting inflammation and hepatic steatosis. Acetate, propionate, and butyrate could relieve hepatic steatosis in the liver by activating AMP-activated protein kinase, expressing fatty acid oxidation gene, and blocking macrophage proinflammatory activation [92]. SCFAs also have a role in the epigenetic regulation of NAFLD development. Propionate, acetate, and butyrate as inhibitors of histone deacetylases, play a key role in NAFLD and prevent gene transcription by reducing histone-bound acetyl groups [93,94].
4.8. Immune System Diseases
SCFAs are involved in immune system diseases by modulating the production of immune mediators, cytokines, and chemokines, as well as regulating the differentiation, recruitment, and activation of immune cells, such as neutrophil, macrophages, DCs, and T lymphocytes [95]. Butyrate had direct salutary effects on target tissues of graft-versus-host disease (GVHD). Supplementing with butyrate could help to improve junctional integrity of IECs, reduce apoptosis, and mitigate the severity of GVHD [96,97]. Recently, enteral nutrition has been shown to have a higher advantage in terms of GVHD protection than parenteral nutrition because it favors a prompt recovery of the SCFAs-producing intestinal microbiota [98]. Propionate and butyrate could ameliorate bone destruction in rheumatoid arthritis, raise systemic bone density, modify the metabolic state of pre-osteoclasts, protect against postmenopausal bone loss, and mitigate arthritis [99]. Butyrate may increase Treg level in colon lamina propria and induce IL-10 secretion in DCs and macrophages to regulate immune functions, which helps to prevent the progression of multiple sclerosis [29]. Furthermore, SCFAs can also shape the immunological environment and influence the allergic inflammation. High levels of propionate in the blood protected against allergic inflammation in lungs by enhancing generation of macrophage and DC precursors depending on GPR41 [100].
4.9. Neurological/Psychiatric Diseases
SCFAs have the potential contributions in determining the etiology and treatment of stress-related disorders [101]. SCFAs supplementation in mice improved sociability, anxiety and depressive-like behaviors, cognition, and stress response [102]. Butyrate has been reported to ameliorate cognitive impairments in mid-adult high-fat-diet-induced obese mice model [103]. Propionate reduced anticipatory reward responses to high-energy food in the human striatum [104]. However, in a previous study, propionate-treated rats showed a series of autism spectrum disorders (ASD)-linked neurochemical changes, which was found to contribute directly or indirectly to acquired mitochondrial dysfunction in breaking carnitine-dependent pathways, similar to the finding in ASD patients [105]. Altogether, these findings suggest that SCFAs have a complicated regulatory mechanism in a variety of diseases.
5. Conclusions
A continually emerging body of evidence supports the role of SCFAs by microbial fermentation in the colon as key mediators potentially aiding disease prevention, recovery, and slowing progression in animal models and preclinical trials. However, there are still some pathogenic/disease-causing consequences in response to diverse diseases due to the differences in SCFAs level, thus, an optimal health-promoting level of SCFAs is still an open question. Indeed, the variability in intestinal microbiota of different individuals exerts the overarching effects in SCFAs level. Additionally, considering that the amount and the variety of dietary partially and non-digestible polysaccharides ingested is difficult to determine whether it is a direct effect of SCFAs or intestinal microbiota that influence host homoeostasis. Therefore, future work remains to be done to comprehensively explore the mechanisms of SCFAs alone or with other metabolites in physiology and pathophysiology of the gut, as well as other tissues and organs in host. Furthermore, more SCFAs-related dietary strategies and pharmacological drugs might develop new possibilities for precision prevention and therapy of various diseases in the future.
Author Contributions
Conceptualization, J.C.; resources, J.C.; writing—original draft preparation, J.C., R.Z. and P.Z.; writing—review and editing, J.C., R.Z. and P.Z.; funding acquisition, J.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by CNS-ZD Tizhi and Health Fund, grant number “CNS-ZD2020-21” and Key Research and Development Project of Hubei Province, grant number “2020BCA081-03“.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The assistance of the staff is gratefully appreciated.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ACI | acute kidney injury |
| AngII | angiotensin II |
| ASD | autism spectrum disorders |
| CDK | chronic kidney disease |
| DCs | dendritic cells |
| GPCR | G-protein-coupled cell surface receptors |
| GVHD | graft-versus-host disease |
| HDAC | histone deacetylase |
| HSL | hormone-sensitive lipase |
| IBD | inflammatory bowel disease |
| IECs | intestinal epithelial cells |
| MCT-1 | monocarboxylate transporter 1 |
| NAFLD | non-obese non-alcoholic fatty liver disease |
| NF-κB | nuclear factor-kappa B |
| NLRP3 | Nod-like receptor family pyrin domain containing 3 |
| NOD | non-obese diabetic |
| NSCLC | non-small cell lung cancer |
| SCFAs | short chain fatty acids |
| SMCT-1 | sodium-coupled monocarboxylate transporter |
| T1D | type 1 diabetes |
| T2D | type 2 diabetes |
| TLR | toll-like receptor |
| Treg | regulatory T cell |
References
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-Induced Changes in the Intestinal Microbiota and Disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidelin, K.N. Fatty acid composition of adipose tissue in humans. Implications for the dietary fat-serum cholesterol-CHD issue. Prog. Lipid Res. 1995, 34, 199–217. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Backhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Windey, K.; De Preter, V.; Verbeke, K. Relevance of protein fermentation to gut health. Mol. Nutr. Food Res. 2012, 56, 184–196. [Google Scholar] [CrossRef]
- Neis, E.P.; van Eijk, H.M.; Lenaerts, K.; Olde Damink, S.W.; Blaak, E.E.; Dejong, C.H.; Rensen, S.S. Distal versus proximal intestinal short-chain fatty acid release in man. Gut 2019, 68, 764–765. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, J.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.; Comelli, E.M. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef]
- Hamer, H.M.; Jonkers, D.; Venema, K.; Vanhoutvin, S.; Troost, F.J.; Brummer, R.J. Review article: The role of butyrate on colonic function. Aliment. Pharmacol. Ther. 2008, 27, 104–119. [Google Scholar] [CrossRef]
- Wong, J.M.; de Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef]
- Topping, D.L.; Clifton, P.M. Short-chain fatty acids and human colonic function: Roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 2001, 81, 1031–1064. [Google Scholar] [CrossRef] [PubMed]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Berg, M.P.; Kiers, E.T.; Driessen, G.; Van Der Heijden, M.; Kooi, B.W.; Kuenen, F.; Liefting, M.; Verhoef, H.A.; Ellers, J. Adapt or disperse: Understanding species persistence in a changing world. Glob. Chang. Biol. 2010, 16, 587–598. [Google Scholar] [CrossRef]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Louis, P.; Young, P.; Holtrop, G.; Flint, H.J. Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 2010, 12, 304–314. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef] [Green Version]
- Meesters, R.J.; van Eijk, H.M.; ten Have, G.A.; de Graaf, A.A.; Venema, K.; van Rossum, B.E.; Deutz, N.E. Application of liquid chromatography-mass spectrometry to measure the concentrations and study the synthesis of short chain fatty acids following stable isotope infusions. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 854, 57–62. [Google Scholar] [CrossRef]
- Pomare, E.W.; Branch, W.J.; Cummings, J.H. Carbohydrate fermentation in the human colon and its relation to acetate concentrations in venous blood. J. Clin. Investig. 1985, 75, 1448–1454. [Google Scholar] [CrossRef] [Green Version]
- Van Immerseel, F.; Ducatelle, R.; De Vos, M.; Boon, N.; Van De Wiele, T.; Verbeke, K.; Rutgeerts, P.; Sas, B.; Louis, P.; Flint, H.J. Butyric acid-producing anaerobic bacteria as a novel probiotic treatment approach for inflammatory bowel disease. J. Med. Microbiol. 2010, 59, 141–143. [Google Scholar] [CrossRef] [Green Version]
- Stoddart, L.A.; Smith, N.J.; Jenkins, L.; Brown, A.J.; Milligan, G. Conserved polar residues in transmembrane domains V, VI, and VII of free fatty acid receptor 2 and free fatty acid receptor 3 are required for the binding and function of short chain fatty acids. J. Biol. Chem. 2008, 283, 32913–32924. [Google Scholar] [CrossRef] [Green Version]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.Y.; Lannoy, V.; Decobecq, M.E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sivaprakasam, S.; Prasad, P.D.; Singh, N. Benefits of short-chain fatty acids and their receptors in inflammation and carcinogenesis. Pharmacol. Ther. 2016, 164, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaki, S.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the short-chain fatty acid receptor, GPR43, in the human colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Wanders, D.; Graff, E.C.; Judd, R.L. Effects of high fat diet on GPR109A and GPR81 gene expression. Biochem. Biophys. Res. Commun. 2012, 425, 278–283. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Kimura, I.; Inoue, D.; Maeda, T.; Hara, T.; Ichimura, A.; Miyauchi, S.; Kobayashi, M.; Hirasawa, A.; Tsujimoto, G. Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc. Natl. Acad. Sci. USA 2011, 108, 8030–8035. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.; Berk, M.; Carvalho, A.; Caso, J.R.; Sanz, Y.; Walder, K.; Maes, M. The Role of the Microbial Metabolites Including Tryptophan Catabolites and Short Chain Fatty Acids in the Pathophysiology of Immune-Inflammatory and Neuroimmune Disease. Mol. Neurobiol. 2017, 54, 4432–4451. [Google Scholar] [CrossRef]
- Haase, S.; Haghikia, A.; Wilck, N.; Müller, D.N.; Linker, R.A. Impacts of microbiome metabolites on immune regulation and autoimmunity. Immunology 2018, 154, 230–238. [Google Scholar] [CrossRef] [Green Version]
- Hippe, B.; Zwielehner, J.; Liszt, K.; Lassl, C.; Unger, F.; Haslberger, A.G. Quantification of butyryl CoA:acetate CoA-transferase genes reveals different butyrate production capacity in individuals according to diet and age. FEMS Microbiol. Lett. 2011, 316, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.; Duan, Y.; Yang, L.; Schnabl, B. Small metabolites, possible big changes: A microbiota-centered view of non-alcoholic fatty liver disease. Gut 2019, 68, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The Colonic Crypt Protects Stem Cells from Microbiota-Derived Metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless, A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Cell Host Microbe 2015, 17, 662–671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Yu, T.; Huang, X.; Bilotta, A.J.; Xu, L.; Lu, Y.; Sun, J.; Pan, F.; Zhou, J.; Zhang, W.; et al. Intestinal microbiota-derived short-chain fatty acids regulation of immune cell IL-22 production and gut immunity. Nat. Commun. 2020, 11, 4457. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W.; Sommer, H.; Kirchner, T.; Paganelli, G.M.; Bartram, P.; Christl, S.; Richter, F.; Dusel, G.; Kasper, H. Effect of butyrate enemas on the colonic mucosa in distal ulcerative colitis. Gastroenterology 1992, 103, 51–56. [Google Scholar] [CrossRef]
- Tang, Y.; Chen, Y.; Jiang, H.; Robbins, G.T.; Nie, D. G-protein-coupled receptor for short-chain fatty acids suppresses colon cancer. Int. J. Cancer 2011, 128, 847–856. [Google Scholar] [CrossRef]
- Singh, N.; Gurav, A.; Sivaprakasam, S.; Brady, E.; Padia, R.; Shi, H.; Thangaraju, M.; Prasad, P.D.; Manicassamy, S.; Munn, D.H.; et al. Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity 2014, 40, 128–139. [Google Scholar] [CrossRef] [Green Version]
- McNabney, S.M.; Henagan, T.M. Short Chain Fatty Acids in the Colon and Peripheral Tissues: A Focus on Butyrate, Colon Cancer, Obesity and Insulin Resistance. Nutrients 2017, 9, 1348. [Google Scholar] [CrossRef] [Green Version]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch from Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef] [Green Version]
- Murugesan, S.; Ulloa-Martínez, M.; Martínez-Rojano, H.; Galván-Rodríguez, F.M.; Miranda-Brito, C.; Romano, M.C.; Piña-Escobedo, A.; Pizano-Zárate, M.L.; Hoyo-Vadillo, C.; García-Mena, J. Study of the diversity and short-chain fatty acids production by the bacterial community in overweight and obese Mexican children. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 1337–1346. [Google Scholar] [CrossRef]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in lean and overweight healthy subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariño, E.; Richards, J.L.; McLeod, K.H.; Stanley, D.; Yap, Y.A.; Knight, J.; McKenzie, C.; Kranich, J.; Oliveira, A.C.; Rossello, F.J.; et al. Gut microbial metabolites limit the frequency of autoimmune T cells and protect against type 1 diabetes. Nat. Immunol. 2017, 18, 552–562. [Google Scholar] [CrossRef] [PubMed]
- Zadeh-Tahmasebi, M.; Duca, F.A.; Rasmussen, B.A.; Bauer, P.V.; Cote, C.D.; Filippi, B.M.; Lam, T.K. Activation of Short and Long Chain Fatty Acid Sensing Machinery in the Ileum Lowers Glucose Production in Vivo. J. Biol. Chem. 2016, 291, 8816–8824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veprik, A.; Laufer, D.; Weiss, S.; Rubins, N.; Walker, M.D. GPR41 modulates insulin secretion and gene expression in pancreatic β-cells and modifies metabolic homeostasis in fed and fasting states. FASEB J. 2016, 30, 3860–3869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade-Oliveira, V.; Amano, M.T.; Correa-Costa, M.; Castoldi, A.; Felizardo, R.J.F.; de Almeida, D.C.; Bassi, E.J.; Moraes-Vieira, P.M.; Hiyane, M.I.; Rodas, A.C.D.; et al. Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion. J. Am. Soc. Nephrol. 2015, 26, 1877–1888. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, B.; Hong, X.; Zhang, X.; Kong, X. Histone deacetylase inhibitor, sodium butyrate, attenuates gentamicin-induced nephrotoxicity by increasing prohibitin protein expression in rats. Eur. J. Pharmacol. 2013, 707, 147–154. [Google Scholar] [CrossRef]
- Khan, S.; Jena, G. Sodium butyrate, a HDAC inhibitor ameliorates eNOS, iNOS and TGF-β1-induced fibrogenesis, apoptosis and DNA damage in the kidney of juvenile diabetic rats. Food Chem. Toxicol. 2014, 73, 127–139. [Google Scholar] [CrossRef]
- Zumbrun, S.D.; Melton-Celsa, A.R.; Smith, M.A.; Gilbreath, J.J.; Merrell, D.S.; O’Brien, A.D. Dietary choice affects Shiga toxin-producing Escherichia coli (STEC) O157:H7 colonization and disease. Proc. Natl. Acad. Sci. USA 2013, 110, E2126–E2133. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, B.P.; Nelson, J.W.; Eskew, J.R.; Ganesan, A.; Ajami, N.J.; Petrosino, J.F.; Bryan, R.M., Jr.; Durgan, D.J. Prebiotics, Probiotics, and Acetate Supplementation Prevent Hypertension in a Model of Obstructive Sleep Apnea. Hypertension 2018, 72, 1141–1150. [Google Scholar] [CrossRef]
- Natarajan, N.; Hori, D.; Flavahan, S.; Steppan, J.; Flavahan, N.A.; Berkowitz, D.E.; Pluznick, J.L. Microbial short chain fatty acid metabolites lower blood pressure via endothelial G protein-coupled receptor 41. Physiol. Genom. 2016, 48, 826–834. [Google Scholar] [CrossRef] [PubMed]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects from Hypertensive Cardiovascular Damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel. Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef] [Green Version]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [Green Version]
- Machiels, K.; Joossens, M.; Sabino, J.; De Preter, V.; Arijs, I.; Eeckhaut, V.; Ballet, V.; Claes, K.; Van Immerseel, F.; Verbeke, K.; et al. A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 2014, 63, 1275–1283. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Harmsen, H.J.M.; Faber, K.N.; Hermoso, M.A. Short Chain Fatty Acids (SCFAs)-Mediated Gut Epithelial and Immune Regulation and Its Relevance for Inflammatory Bowel Diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [Green Version]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.Y.; de Zoete, M.R.; van Putten, J.P.; Strijbis, K. Redirection of Epithelial Immune Responses by Short-Chain Fatty Acids through Inhibition of Histone Deacetylases. Front. Immunol. 2015, 6, 554. [Google Scholar] [CrossRef] [Green Version]
- Elamin, E.E.; Masclee, A.A.; Dekker, J.; Pieters, H.J.; Jonkers, D.M. Short-chain fatty acids activate AMP-activated protein kinase and ameliorate ethanol-induced intestinal barrier dysfunction in Caco-2 cell monolayers. J. Nutr. 2013, 143, 1872–1881. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Yu, Y.; Wang, Y.Z.; Wang, J.J.; Guan, R.; Sun, Y.; Shi, F.; Gao, J.; Fu, X.L. Role of SCFAs in gut microbiome and glycolysis for colorectal cancer therapy. J. Cell. Physiol. 2019, 234, 17023–17049. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Umar, S.; Rust, B.; Lazarova, D.; Bordonaro, M. Secondary Bile Acids and Short Chain Fatty Acids in the Colon: A Focus on Colonic Microbiome, Cell Proliferation, Inflammation, and Cancer. Int. J. Mol. Sci. 2019, 20, 1214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Botticelli, A.; Vernocchi, P.; Marini, F.; Quagliariello, A.; Cerbelli, B.; Reddel, S.; Del Chierico, F.; Di Pietro, F.; Giusti, R.; Tomassini, A.; et al. Gut metabolomics profiling of non-small cell lung cancer (NSCLC) patients under immunotherapy treatment. J. Transl. Med. 2020, 18, 49. [Google Scholar] [CrossRef] [PubMed]
- Coutzac, C.; Jouniaux, J.M.; Paci, A.; Schmidt, J.; Mallardo, D.; Seck, A.; Asvatourian, V.; Cassard, L.; Saulnier, P.; Lacroix, L.; et al. Systemic short chain fatty acids limit antitumor effect of CTLA-4 blockade in hosts with cancer. Nat. Commun. 2020, 11, 2168. [Google Scholar] [CrossRef]
- Waalen, J. The genetics of human obesity. Transl. Res. 2014, 164, 293–301. [Google Scholar] [CrossRef]
- Reshef, L.; Niv, J.; Shapiro, B. Effect of propionate on lipogenesis in adipose tissue. J. Lipid. Res. 1967, 8, 682–687. [Google Scholar] [CrossRef]
- Wolever, T.M.; Brighenti, F.; Royall, D.; Jenkins, A.L.; Jenkins, D.J. Effect of rectal infusion of short chain fatty acids in human subjects. Am. J. Gastroenterol. 1989, 84, 1027–1033. [Google Scholar]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J., Jr.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef]
- Royall, D.; Wolever, T.M.; Jeejeebhoy, K.N. Clinical significance of colonic fermentation. Am. J. Gastroenterol. 1990, 85, 1307–1312. [Google Scholar]
- Priyadarshini, M.; Villa, S.R.; Fuller, M.; Wicksteed, B.; Mackay, C.R.; Alquier, T.; Poitout, V.; Mancebo, H.; Mirmira, R.G.; Gilchrist, A.; et al. An Acetate-Specific GPCR, FFAR2, Regulates Insulin Secretion. Mol. Endocrinol. 2015, 29, 1055–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.; Goergen, C.J.; HogenEsch, H.; Kim, C.H. Chronically Elevated Levels of Short-Chain Fatty Acids Induce T Cell-Mediated Ureteritis and Hydronephrosis. J. Immunol. 2016, 196, 2388–2400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, S.D.; Blakeman, T. Chronic kidney disease: Identification and management in primary care. Pragmatic Obs. Res. 2016, 7, 21–32. [Google Scholar] [CrossRef] [Green Version]
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540. [Google Scholar] [CrossRef] [Green Version]
- Vaziri, N.D.; Liu, S.-M.; Lau, W.L.; Khazaeli, M.; Nazertehrani, S.; Farzaneh, S.H.; Kieffer, D.A.; Adams, S.H.; Martin, R.J. High amylose resistant starch diet ameliorates oxidative stress, inflammation, and progression of chronic kidney disease. PLoS ONE 2014, 9, e114881. [Google Scholar] [CrossRef]
- Wong, J.; Piceno, Y.M.; DeSantis, T.Z.; Pahl, M.; Andersen, G.L.; Vaziri, N.D. Expansion of urease- and uricase-containing, indole- and p-cresol-forming and contraction of short-chain fatty acid-producing intestinal microbiota in ESRD. Am. J. Nephrol. 2014, 39, 230–237. [Google Scholar] [CrossRef] [Green Version]
- Rudemiller, N.P.; Crowley, S.D. Interactions Between the Immune and the Renin-Angiotensin Systems in Hypertension. Hypertension 2016, 68, 289–296. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B.; et al. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Wilck, N.; Matus, M.G.; Kearney, S.M.; Olesen, S.W.; Forslund, K.; Bartolomaeus, H.; Haase, S.; Mähler, A.; Balogh, A.; Markó, L.; et al. Salt-responsive gut commensal modulates T(H)17 axis and disease. Nature 2017, 551, 585–589. [Google Scholar] [CrossRef]
- Miura, K.; Greenland, P.; Stamler, J.; Liu, K.; Daviglus, M.L.; Nakagawa, H. Relation of vegetable, fruit, and meat intake to 7-year blood pressure change in middle-aged men: The Chicago Western Electric Study. Am. J. Epidemiol. 2004, 159, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Guo, Z.; Chen, D.; Li, L.; Song, X.; Liu, T.; Jin, G.; Li, Y.; Liu, Y.; Ajiguli, A.; et al. Maternal sucralose intake alters gut microbiota of offspring and exacerbates hepatic steatosis in adulthood. Gut Microbes 2020, 11, 1043–1063. [Google Scholar] [CrossRef] [PubMed]
- Dangana, E.O.; Omolekulo, T.E.; Areola, E.D.; Olaniyi, K.S.; Soladoye, A.O.; Olatunji, L.A. Sodium acetate protects against nicotine-induced excess hepatic lipid in male rats by suppressing xanthine oxidase activity. Chem. Biol. Interact. 2020, 316, 108929. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.J.; Sellmann, C.; Engstler, A.J.; Ziegenhardt, D.; Bergheim, I. Supplementation of sodium butyrate protects mice from the development of non-alcoholic steatohepatitis (NASH). Br. J. Nutr. 2015, 114, 1745–1755. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered Fecal Microbiota Correlates with Liver Biochemistry in Nonobese Patients with Non-alcoholic Fatty Liver Disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef]
- Ghosh, A.; Gao, L.; Thakur, A.; Siu, P.M.; Lai, C.W.K. Role of free fatty acids in endothelial dysfunction. J. Biomed. Sci. 2017, 24, 50. [Google Scholar] [CrossRef] [Green Version]
- Cordeiro, A.; Costa, R.; Andrade, N.; Silva, C.; Canabrava, N.; Pena, M.J.; Rodrigues, I.; Andrade, S.; Ramalho, A. Does adipose tissue inflammation drive the development of non-alcoholic fatty liver disease in obesity? Clin. Res. Hepatol. Gastroenterol. 2020, 44, 394–402. [Google Scholar] [CrossRef]
- Jocken, J.W.E.; González Hernández, M.A.; Hoebers, N.T.H.; van der Beek, C.M.; Essers, Y.P.G.; Blaak, E.E.; Canfora, E.E. Short-Chain Fatty Acids Differentially Affect Intracellular Lipolysis in a Human White Adipocyte Model. Front. Endocrinol. 2017, 8, 372. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Fu, C. Acetate Affects the Process of Lipid Metabolism in Rabbit Liver, Skeletal Muscle and Adipose Tissue. Animals 2019, 9, 799. [Google Scholar] [CrossRef] [Green Version]
- Samuel, B.S.; Shaito, A.; Motoike, T.; Rey, F.E.; Backhed, F.; Manchester, J.K.; Hammer, R.E.; Williams, S.C.; Crowley, J.; Yanagisawa, M.; et al. Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc. Natl. Acad. Sci. USA 2008, 105, 16767–16772. [Google Scholar] [CrossRef] [Green Version]
- Macia, L.; Tan, J.; Vieira, A.T.; Leach, K.; Stanley, D.; Luong, S.; Maruya, M.; Ian McKenzie, C.; Hijikata, A.; Wong, C.; et al. Metabolite-sensing receptors GPR43 and GPR109A facilitate dietary fibre-induced gut homeostasis through regulation of the inflammasome. Nat. Commun. 2015, 6, 6734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skelly, A.N.; Sato, Y. Mining the microbiota for microbial and metabolite-based immunotherapies. Nat. Rev. Immunol. 2019, 19, 305–323. [Google Scholar] [CrossRef] [PubMed]
- Campisano, S.; La Colla, A.; Echarte, S.M.; Chisari, A.N. Interplay between early-life malnutrition, epigenetic modulation of the immune function and liver diseases. Nutr. Res. Rev. 2019, 32, 128–145. [Google Scholar] [CrossRef] [PubMed]
- Kolodziejczyk, A.A.; Zheng, D.; Shibolet, O.; Elinav, E. The role of the microbiome in NAFLD and NASH. EMBO Mol. Med. 2019, 11, e9302. [Google Scholar] [CrossRef]
- Corrêa-Oliveira, R.; Fachi, J.L.; Vieira, A.; Sato, F.T.; Vinolo, M.A.R. Regulation of immune cell function by short-chain fatty acids. Clin. Transl. Immunol. 2016, 5, e73. [Google Scholar] [CrossRef]
- Mathewson, N.D.; Jenq, R.; Mathew, A.V.; Koenigsknecht, M.; Hanash, A.; Toubai, T.; Oravecz-Wilson, K.; Wu, S.R.; Sun, Y.; Rossi, C.; et al. Gut microbiome-derived metabolites modulate intestinal epithelial cell damage and mitigate graft-versus-host disease. Nat. Immunol. 2016, 17, 505–513. [Google Scholar] [CrossRef]
- Biagi, E.; Zama, D.; Rampelli, S.; Turroni, S.; Brigidi, P.; Consolandi, C.; Severgnini, M.; Picotti, E.; Gasperini, P.; Merli, P.; et al. Early gut microbiota signature of aGvHD in children given allogeneic hematopoietic cell transplantation for hematological disorders. BMC Med. Genom. 2019, 12, 49. [Google Scholar] [CrossRef]
- D’Amico, F.; Biagi, E.; Rampelli, S.; Fiori, J.; Zama, D.; Soverini, M.; Barone, M.; Leardini, D.; Muratore, E.; Prete, A.; et al. Enteral Nutrition in Pediatric Patients Undergoing Hematopoietic SCT Promotes the Recovery of Gut Microbiome Homeostasis. Nutrients 2019, 11, 2958. [Google Scholar] [CrossRef] [Green Version]
- Lucas, S.; Omata, Y.; Hofmann, J.; Böttcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Krönke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef] [Green Version]
- Trompette, A.; Gollwitzer, E.S.; Yadava, K.; Sichelstiel, A.K.; Sprenger, N.; Ngom-Bru, C.; Blanchard, C.; Junt, T.; Nicod, L.P.; Harris, N.L.; et al. Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat. Med. 2014, 20, 159–166. [Google Scholar] [CrossRef]
- Foster, J.A.; Rinaman, L.; Cryan, J.F. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol. Stress 2017, 7, 124–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: Microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef] [PubMed]
- Arnoldussen, I.A.C.; Wiesmann, M.; Pelgrim, C.E.; Wielemaker, E.M.; van Duyvenvoorde, W.; Amaral-Santos, P.L.; Verschuren, L.; Keijser, B.J.F.; Heerschap, A.; Kleemann, R.; et al. Butyrate restores HFD-induced adaptations in brain function and metabolism in mid-adult obese mice. Int. J. Obes. 2017, 41, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Byrne, C.S.; Chambers, E.S.; Alhabeeb, H.; Chhina, N.; Morrison, D.J.; Preston, T.; Tedford, C.; Fitzpatrick, J.; Irani, C.; Busza, A.; et al. Increased colonic propionate reduces anticipatory reward responses in the human striatum to high-energy foods. Am. J. Clin. Nutr. 2016, 104, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfabe, D.F. Short-chain fatty acid fermentation products of the gut microbiome: Implications in autism spectrum disorders. Microb. Ecol. Health Dis. 2012, 23, 3402. [Google Scholar] [CrossRef]
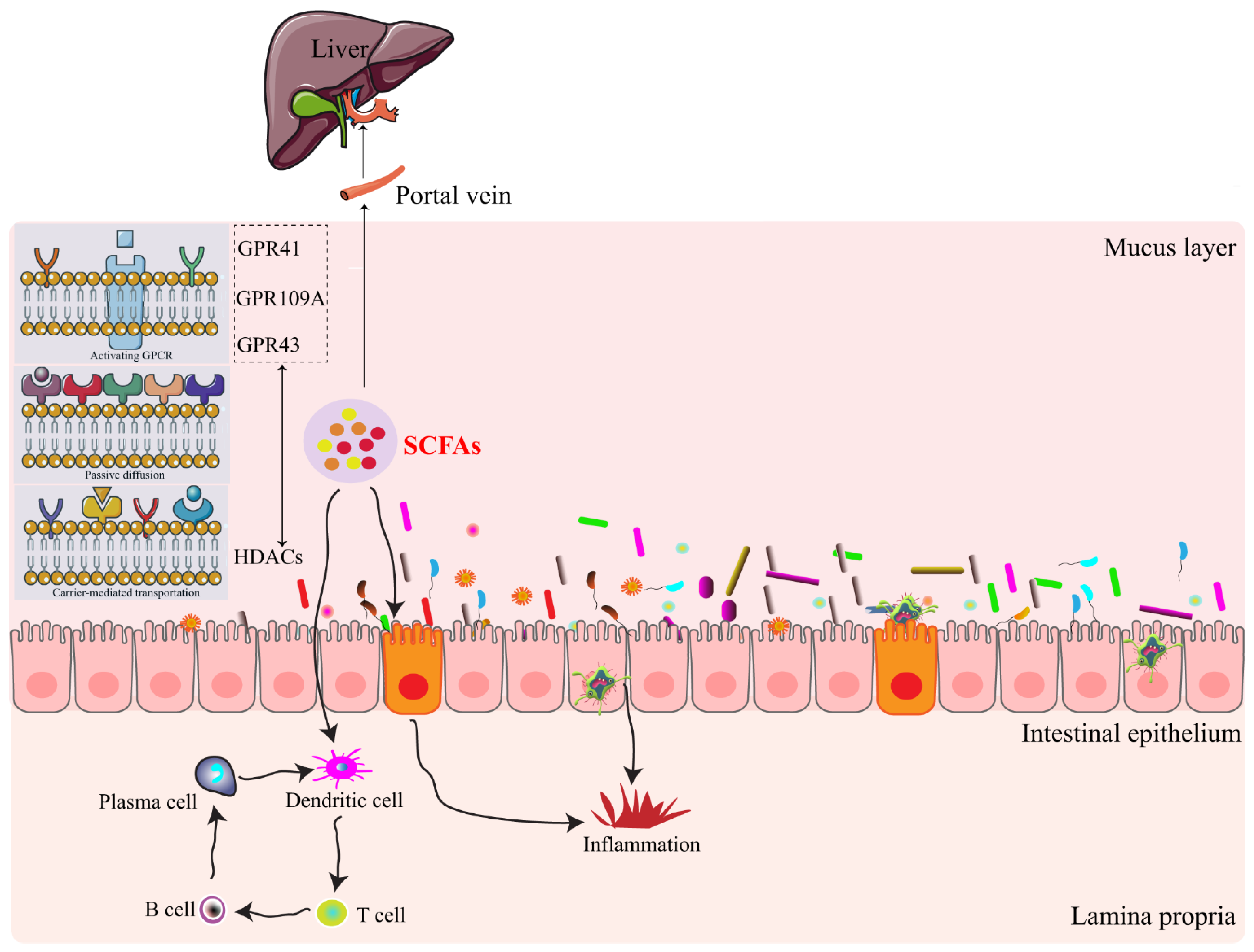
Figure 1.
Interactions of short chain fatty acids with intestinal epithelium based on histone deacetylases (HDACs) and G protein-coupled receptors (GPCRs). Intestinal microbiota-derived metabolites of SCFAs are absorbed in direct or indirect ways. They are then absorbed into the blood vessels and finally enter the liver through the portal vein, as well as interact with the intestinal immune system. GPR41, G-protein coupled receptor 41; GPR43, G-protein coupled receptor 43; GPR109A, G-protein coupled receptor 109A.
Figure 1.
Interactions of short chain fatty acids with intestinal epithelium based on histone deacetylases (HDACs) and G protein-coupled receptors (GPCRs). Intestinal microbiota-derived metabolites of SCFAs are absorbed in direct or indirect ways. They are then absorbed into the blood vessels and finally enter the liver through the portal vein, as well as interact with the intestinal immune system. GPR41, G-protein coupled receptor 41; GPR43, G-protein coupled receptor 43; GPR109A, G-protein coupled receptor 109A.

Figure 2.
Intestinal microbiota-derived metabolites SCFAs result in the alteration of intestinal barrier function and are closely linked with host health and disease.
Figure 2.
Intestinal microbiota-derived metabolites SCFAs result in the alteration of intestinal barrier function and are closely linked with host health and disease.

Table 1.
Gradient concentrations of SCFAs between gut lumen and periphery.
| SCFAs | Proximal Colon | Portal Vein | Hepatic Vein | Systemic | Peripheral | Cerebrospinal Fluid |
|---|---|---|---|---|---|---|
| Acetate | 42–84 mM | 375 μM | 148 μM | 100–200 μM | 79 μM | 35 μM |
| Propionate | 14–28 mM | - | - | 1–15 μM | - | - |
| Butyrate | 14–28 mM | - | - | 1–15 μM | - | - |
Table 3.
Examples of SCFAs involved in the diverse diseases in vivo and in vitro studies for recent 20 years (2000–2020).
Table 3.
Examples of SCFAs involved in the diverse diseases in vivo and in vitro studies for recent 20 years (2000–2020).
| Related Diseases | Relevant Short Chain Fatty Acids | Publications (2000–2020) |
|---|---|---|
| Inflammatory bowel disease | Butyrate [32,33,34,35] | 174 |
| Cancer | Propionate [36], Butyrate [37,38] | 413 |
| Obesity | Acetate [39], Propionate [8,39,40,41], Butyrate [39,40,42] | 493 |
| Diabetes | Acetate [43], Propionate [44,45], Butyrate [43] | 367 |
| Kidney diseases | Acetate [46]; Butyrate [47,48,49] | 108 |
| Hypertension | Acetate [50], Propionate [51,52], Butyrate [53] | 79 |
Number of results obtained by searching for “(short chain fatty acid [Title/Abstract]) AND (Inflammatory bowel disease/cancer/obesity/diabetes/kidney/hypertension[Title/Abstract])” on PubMed from 2000 to 2020.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Cong, J.; Zhou, P.; Zhang, R. Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease. Nutrients 2022, 14, 1977. https://doi.org/10.3390/nu14091977
AMA Style
Cong J, Zhou P, Zhang R. Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease. Nutrients. 2022; 14(9):1977. https://doi.org/10.3390/nu14091977
Chicago/Turabian StyleCong, Jing, Ping Zhou, and Ruiyan Zhang. 2022. "Intestinal Microbiota-Derived Short Chain Fatty Acids in Host Health and Disease" Nutrients 14, no. 9: 1977. https://doi.org/10.3390/nu14091977
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.