
Identification of Inhibitory Activities of Dietary Flavonoids against URAT1, a Renal Urate Re-Absorber: In Vitro Screening and Fractional Approach Focused on Rooibos Leaves

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Resources
2.2. Preparation of Plant Ethanolic Extracts
2.3. Cell Culture
2.4. Urate Transport Assay Using URAT1-Expressing 293A Cells
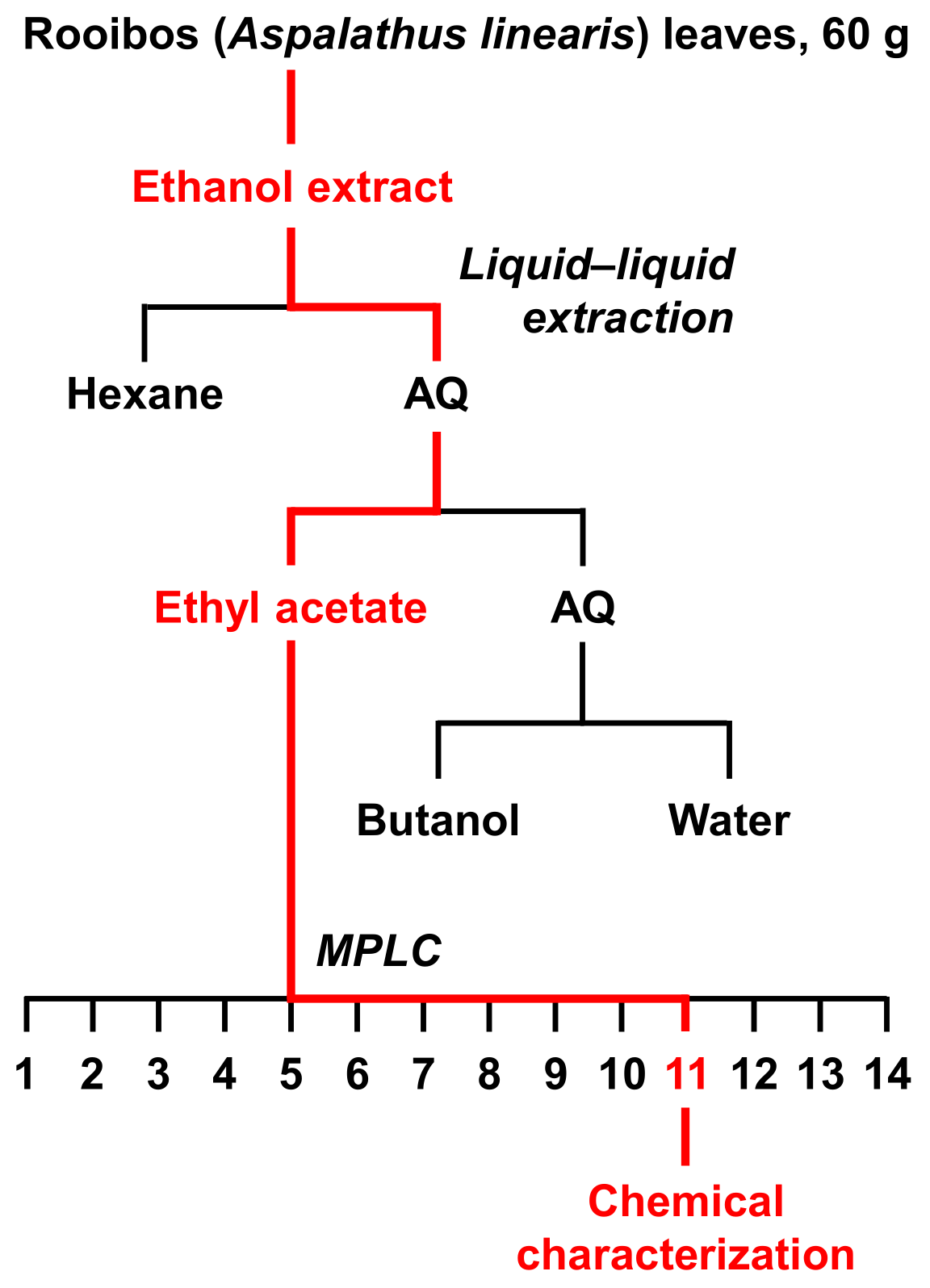
2.5. Fractionation of Rooibos Tea Leaves Extract
2.6. Chemical Characterization
2.7. Statistical Analysis
3. Results
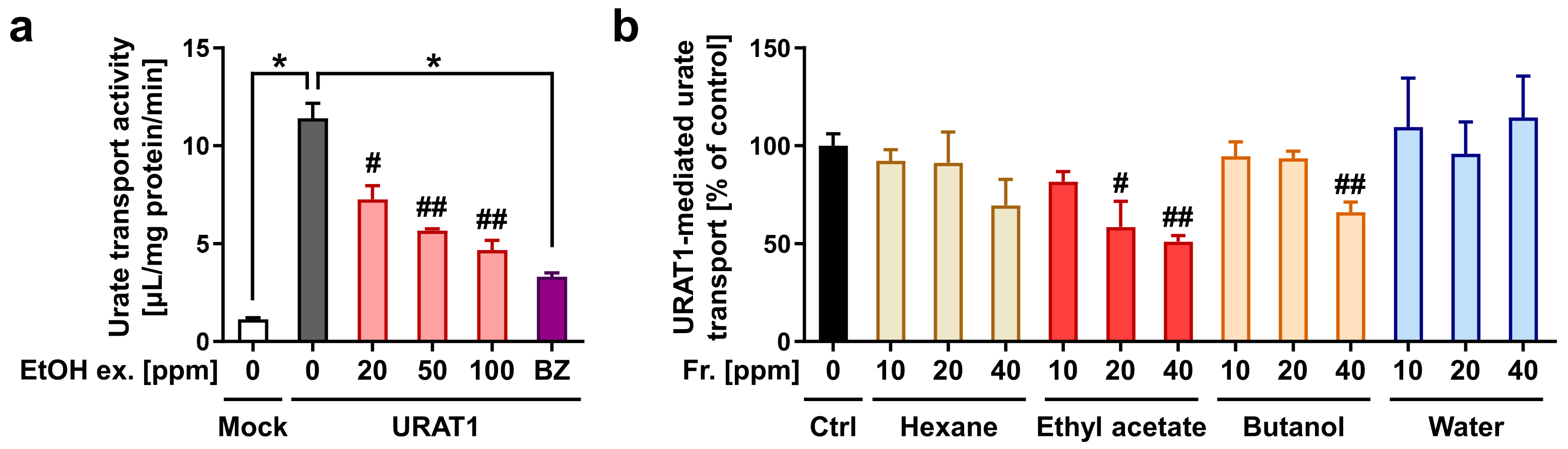
3.1. Screening the URAT1-Inhibitory Activities of Plant Extracts
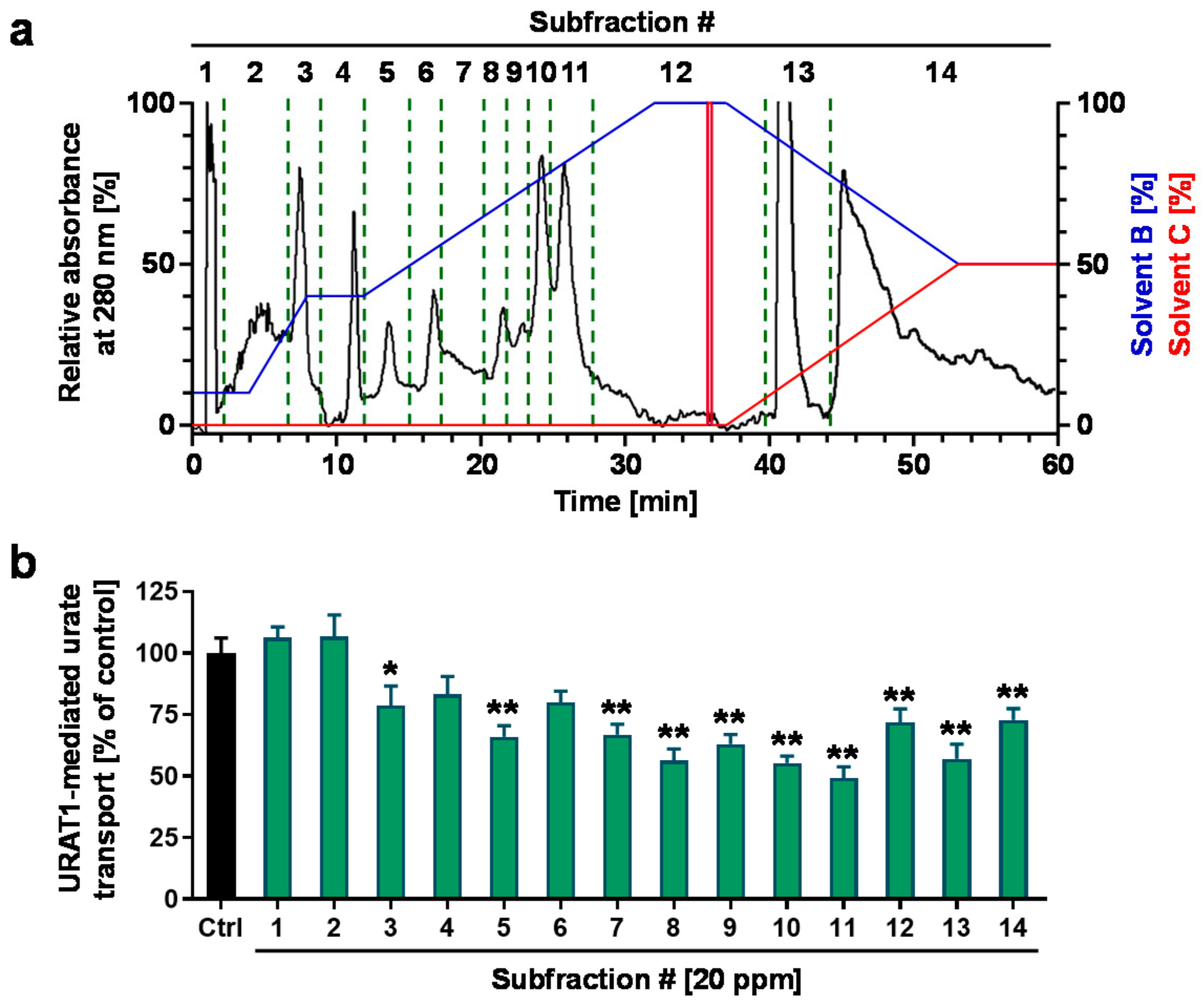
3.2. Fractionation and Isolation of the Aspalathus linearis (Rooibos Leaves) Extract
3.3. Structural Characterization of the Putative URAT1 Inhibitor Derived from Rooibos Leaves
3.4. Identification of Quercetin as an Active Ingredient with URAT1-Inhibitory Activity
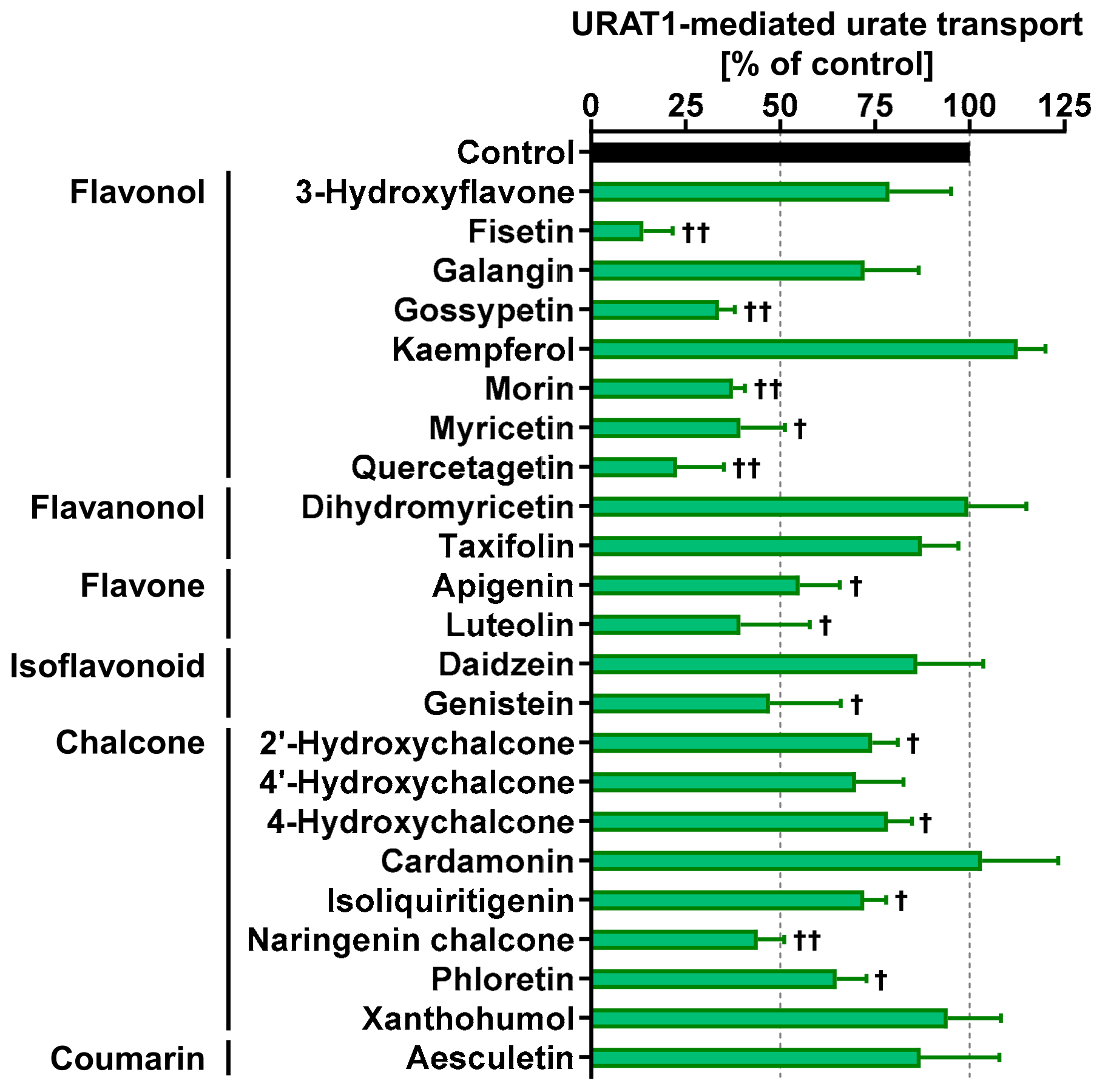
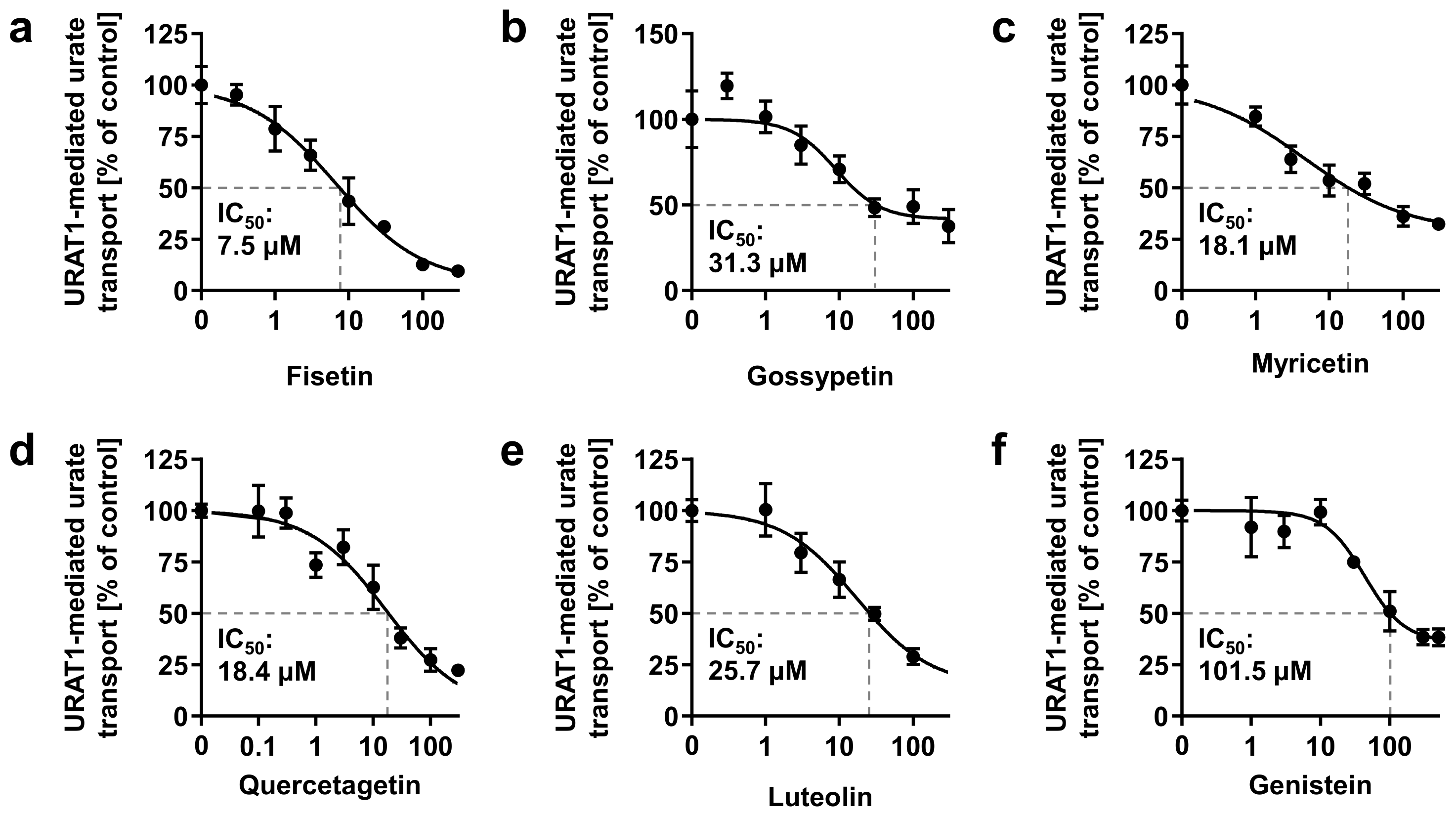
3.5. URAT1-Inhibitory Activities of Various Dietary Flavonoids
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Descriptions in This Study | Common Names | Academic Names | Details of Material * |
|---|---|---|---|
| Abelmoschus esculentus (beniokura) | Beniokura | Abelmoschus esculentus | Fresh |
| Agaricus bisporus | Common mushroom | Agaricus bisporus | Fresh |
| Allium cepa | Onion | Allium cepa | Fresh |
| Allium oschaninii | Shallot | Allium oschaninii | Fresh |
| Allium sativum | Garlic | Allium sativum | Fresh |
| Allium sativum (sprout) | Garlic shoots | Allium sativum | Fresh sprout |
| Allium tuberosum | Chinese chive | Allium tuberosum | Fresh |
| Ananas comosus (coat) | Pineapple | Ananas comosus | Fresh coat |
| Apium graveolens | Celery | Apium graveolens | Fresh |
| Apium graveolens (salad celery) | Salad celery | Apium graveolens | Fresh |
| Arachis hypogaea (beans) | Peanut | Arachis hypogaea | Fresh beans |
| Arachis hypogaea (shell) | Peanut | Arachis hypogaea | Fresh shell |
| Aralia cordata | Udo | Aralia cordata | Fresh |
| Aralia elata (sprout) | Fatsia sprouts | Aralia elata | Fresh sprout |
| Arctium lappa | Edible burdock | Arctium lappa | Fresh root |
| Arctium lappa (burdock tea) | Burdock root tea | Arctium lappa | Dried root for tea |
| Aspalathus linearis | Rooibos tea leaves | Aspalathus linearis | Dried leaves for tea |
| Asparagus officinalis (grass roots) | Asparagus | Asparagus officinalis | Fresh grass roots |
| Asparagus spp. | Asparagus | Asparagus spp. | Fresh stalk |
| Auricularia auricula-judae | Jew’s ear fungus | Auricularia auricula-judae | Fresh |
| Barley (Hordeum vulgare) Miso | Barley Miso | Hordeum vulgare# | Japanese traditional fermented product |
| Basella alba | Indian spinach | Basella alba | Fresh |
| Benincasa hispida (coat, placenta, seed) | Winter melon | Benincasa hispida | Fresh coat, placenta and seeds |
| Benincasa hispida (meat) | Winter melon | Benincasa hispida | Fresh meat |
| Brassica chinensis | Green pak choi | Brassica rapa var. chinensis | Fresh |
| Brassica oleracea (broccoli, anthotaxy) | Broccoli | Brassica oleracea var. italica | Fresh anthotaxy |
| Brassica oleracea (broccoli, sprout) | Broccoli | Brassica oleracea var. italica | Fresh sprout |
| Brassica oleracea (broccoli, stem) | Broccoli | Brassica oleracea var. italica | Fresh stem |
| Brassica oleracea (kohlrabi, peel) | German turnip or turnip cabbage | Brassica oleracea var. gongylodes | Fresh peel |
| Brassica oleracea (red cabbage, sprout) | Red cabbage | Brassica oleracea var. capitata F. rubra | Fresh sprout |
| Brassica oleracea (romanesco broccoli, stem) | Romanesco broccoli | Brassica oleracea var. botrytis | Fresh stem |
| Brassica oleracea (soft kale) | Soft kale | Brassica oleracea var. acephala | Fresh stems and leaves |
| Brassica oleracea (stick senor) | Stick senor | Brassica oleracea var. italica | Fresh |
| Brassica oleracea (wild cabbage, flower) | Romanesco broccoli | Brassica oleracea var. botrytis | Fresh flower |
| Brassica rapa (ayameyuki-kabu) | Ayameyuki-kabu | Brassica rapa | Fresh leaves |
| Brassica rapa (ayameyuki-kabu, meat) | Ayameyuki-kabu | Brassica rapa | Fresh meat |
| Brassica rapa (nabana) | Chinese colza | Brassica rapa L. var. nippo-oleifera | Fresh leaves |
| Brassica rapa (nabana, flower) | Chinese colza | Brassica rapa L. var. nippo-oleifera | Fresh flower |
| Brassica rapa (red) | Red potherb mustard | Brassica rapa var. laciniifolia | Fresh |
| Brassica rapa (santo-sai) | Santo-sai | Brassica rapa L. var. pekinensis | Fresh |
| Capsicum annuum (redpepper) | Chili pepper | Capsicum annuum L. | Fresh |
| Capsicum annuum (sweet pepper) | Sweet pepper | Capsicum annuum L. ‘grossum’ | Fresh |
| Capsicum annuum (red) | Red bell pepper | Capsicum annuum L. ‘grossum’ | Fresh |
| Capsicum annuum (shishitou) | Shishitou | Capsicum annuum L. | Fresh |
| Capsicum annuum (yellow) | Yellow bell pepper | Capsicum annuum L. ‘grossum’ | Fresh |
| Capsicum frutescens | Shima pepper | Capsicum frutescens L. | Fresh |
| Carica papaya (immature, meat) | Green papaya | Carica papaya L. | Fresh meat |
| Carica papaya (immature, peel, placenta, seed) | Green papaya | Carica papaya L. | Fresh peel, placenta and seed |
| Caulerpa lentillifera | Sea grape | Caulerpa lentillifera | Fresh |
| Citrus aurantiifolia (peel) | Lime | Citrus aurantiifolia | Peel |
| Citrus depressa (peel) | Shikuwasa | Citrus depressa | Peel |
| Citrus junos (peel) | Yuzu | Citrus junos | Peel |
| Citrus maxima (peel) | Pomelo | Citrus maxima | Peel |
| Citrus maxima (placenta) | Pomelo | Citrus maxima | Inner white and soft tissue layer |
| Citrus natsudaidai (peel) | Suruga elegant | Citrus natsudaidai | Peel |
| Citrus paradisi (peel) | Grapefruit | Citrus paradisi | Peel |
| Citrus reticulata (peel) | Ponkan | Citrus reticulata | Peel |
| Citrus sinensis (blood orange, peel) | Blood orange | Citrus sinensis | Peel |
| Citrus sinensis (navel) | Navel | Citrus sinensis | Peel |
| Citrus sphaerocarpa (peel) | Kabosu | Citrus sphaerocarpa | Peel |
| Citrus sudachi (peel) | Sudachi | Citrus sudachi | Peel |
| Citrus tangelo (peel) | Mineola orange (tangelo) | Citrus tangelo | Peel |
| Cocos nucifera (young) | Young coconut | Cocos nucifera | Fresh |
| Colocasia esculenta | Eddoe | Colocasia esculenta L. schott | Fresh |
| Coriandrum sativum | Coriander | Coriandrum sativum | Fresh leaves |
| Coriandrum sativum (leaves) | Coriander | Coriandrum sativum | Fresh leaves |
| Cucumis melo (coat) | Melon | Cucumis melo | Fresh coat |
| Cucurbita (meat) | Squash | Cucurbita | Fresh meat, without seeds |
| Cucurbita (peel) | Squash | Cucurbita | Fresh peel |
| Cucurbita pepo (yellow, peel) | Zucchini | Cucurbita pepo | Fresh peel |
| Curcuma longa | Turmeric | Curcuma longa L. | Dried powder |
| Cyperus esculentus (powder) | Yellow nutsedge | Cyperus esculentus | Milled powder of stem |
| Daucus carota | Carrot | Daucus carota subsp. sativus | Fresh |
| Daucus carota (purple carrot) | Purple carrot | Daucus carota subsp. sativus | Fresh |
| Dioscorea japonica | Japanese yam | Dioscorea japonica | Fresh |
| Diospyros kaki (shibugaki, meat) | Kaki persimmon | Diospyros kaki | Fresh |
| Diospyros kaki (shibugaki, peel) | Kaki persimmon | Diospyros kaki | Fresh |
| Eriobotrya japonica (peel) | Loquat | Eriobotrya japonica | Fresh |
| Eutrema japonicum | Japanese horseradish | Eutrema japonicum | Fresh root |
| Eutrema japonicum (stem) | Japanese horseradish | Eutrema japonicum | Fresh stem |
| Fagopyrum tataricum | Tartary buckwheat | Fagopyrum tataricum | Dried seed |
| Ficus carica | Fig tree | Ficus carica | Fresh fruit |
| Flammulina velutipes | Enoki mushroom | Flammulina velutipes | Fresh |
| Fortunella (peel) | Kumquat | Fortunella | Peel |
| Fragaria ananassa | Strawberry | Fragaria ananassa | Fresh |
| Ginkgo biloba (seed) | Ginkgo | Ginkgo biloba | Fresh |
| Glebionis coronaria | Crown daisy | Glebionis coronaria | Fresh |
| Glycine max | Soybeans (yellow soybean) | Glycine max | Dried product |
| Glycine max (hidenmame) | Soybeans (green soybean) | Glycine max | Dried product |
| Glycine max (immature) | Immature soybeans | Glycine max | Fresh |
| Glycine max (immature, shuck) | Immature soybeans | Glycine max | Fresh shuck |
| Glycine max × Bacillus subtilis | Natto | Glycine max | Commercially available Japanese traditional fermented product |
| Grifola frondosa | Hen-of-the-woods | Grifola frondosa | Fresh |
| Hibiscus rosa-sinensis | Chinese hibiscus | Hibiscus rosa-sinensis | Fresh |
| Hosta sieboldiana | Hosta | Hosta sieboldiana | Fresh young leaves |
| Houttuynia cordata | Fish mint | Houttuynia cordata | Dried leaves and stem |
| Humulus lupulus (cone) | Hop | Humulus lupulus | Frozen hop cone |
| Hylocereus undatus (peel) | Dragon fruit | Hylocereus undatus | Fresh peel |
| Hypsizygus marmoreus | Shimeji mushroom | Hypsizygus marmoreus | Fresh |
| Ilex paraguariensis (roasted) | Yerba mate tea leaves | Ilex paraguariensis | Dried and roasted leaves for tea |
| Illicium verum | Star anise | Illicium verum | Dried fruit |
| Ipomoea aquatica | Water morning glory | Ipomoea aquatica | Fresh |
| Jasminum sambac | Jasmine tea leaves | Jasminum sambac | Dried leaves for tea |
| Lactuca sativa | Stem lettuce | Lactuca sativa L. var. crispa | Fresh |
| Laminaria longissima (tororomekonbu) | Tororomekonbu | Laminaria longissima | Dried product |
| Laurus nobilis (leaves) | Laurel | Laurus nobilis | Fresh leaves |
| Lentinula edodes | Shiitake mushroom | Lentinula edodes | Fresh |
| Lycopersicum esculentum (yellow) | Cherry tomato | Solanum lycopersicum L. | Fresh |
| Matricaria recutita | Chamomile | Matricaria recutita | Dried herb product |
| Matteuccia struthiopteris (young) | Ostrich fern | Matteuccia struthiopteris | Fresh young leaves |
| Mesembryanthemum crystallinum | Common ice plant | Mesembryanthemum crystallinum | Fresh |
| Momordica charantia (coat) | Bitter melon | Momordica charantia | Fresh coat |
| Musa spp. (peel) | Banana | Musa spp. | Fresh peel |
| Musa spp. (peel, Ecuador) | Banana | Musa spp. | Fresh peel |
| Nasturtium officinale | Watercress | Nasturtium officinale | Fresh |
| Nelumbo nucifera | Lotus root | Nelumbo nucifera | Fresh root |
| Ocimum basilicum | Basil | Ocimum basilicum | Fresh |
| Ocimum basilicum (purple) | Purple basil | Ocimum basilicum | Fresh |
| Oryza sativa (black) | Brack rice | Oryza sativa | Fresh |
| Perilla frutescens | Perilla | Perilla frutescens | Fresh |
| Persea americana (coat) | Avocado | Persea americana | Fresh coat |
| Persea americana (seed) | Avocado | Persea americana | Fresh seed |
| Petasites japonicus | Giant butterbur | Petasites japonicus | Fresh |
| Petroselinum crispum (leaves) | Parsley | Petroselinum crispum | Fresh leaves |
| Phaseolus vulgaris | Common bean | Phaseolus vulgaris | Fresh |
| Phaseolus vulgaris (Moroccan kidney beans) | Moroccan kidney beans | Phaseolus vulgaris | Fresh |
| Pholiota microspora | Butterscotch mushroom | Pholiota microspora | Fresh |
| Phyllostachys pubescens (young, dried) | Bamboo shoot | Phyllostachys pubescens | Dried young stem |
| Pisum sativum | Pea | Pisum sativum | Fresh |
| Pisum sativum (shelled) | Shelled pea | Pisum sativum | Fresh |
| Pisum sativum (shuck-edible) | Shuck-edible pea | Pisum sativum | Fresh |
| Pisum sativum (young leaves) | Pea young leaves | Pisum sativum | Fresh |
| Pleurotus cornucopiae | Golden oyster mushroom | Pleurotus cornucopiae var. citrinopileatus | Fresh |
| Pleurotus eryngii | King trumpet mushroom | Pleurotus eryngii | Fresh |
| Pleurotus ostreatus | Oyster mushroom | Pleurotus ostreatus | Fresh |
| Prunus domestica (extract) | Prune extract | Prunus domestica | Product of prune pulp extract ‡ |
| Prunus domestica (meat) | Prune | Prunus domestica | Product of prune pulp without seed |
| Prunus tomentosa (peel) | Cherry | Prunus tomentosa | Fresh peel |
| Psidium guajava (Chinese) | Guava tea leaves | Psidium guajava | Dried leaves for tea cultivated in China |
| Psidium guajava (Japanese) | Guava tea leaves | Psidium guajava | Dried leaves for tea cultivated in Japan |
| Psophocarpus tetragonolobus | Winged bean | Psophocarpus tetragonolobus | Fresh |
| Pteridium aquilinum | Western bracken fern | Pteridium aquilinum | Fresh |
| Pyrus communis (peel) | Pear | Pyrus communis | Fresh peel |
| Raphanus sativus (leaves) | Radish | Raphanus sativus L. var. sativus | Fresh leaves |
| Raphanus sativus (meat) | Radish | Raphanus sativus L. var. sativus | Fresh meat |
| Raphanus sativus (radish sprout) | Radish sprout | Raphanus sativus | Fresh |
| Rice (Oryza sativa) Miso | Rice Miso | Oryza sativa # | Japanese traditional fermented product |
| Rosmarinus officinalis (raw) | Rosemary | Rosmarinus officinalis | Fresh |
| Sechium edule (meat) | Chayote | Sechium edule | Fresh meat |
| Sechium edule (peel, placenta) | Chayote | Sechium edule | Fresh peel and placenta |
| Sesamum indicum | Sesame | Sesamum indicum | Dried seeds |
| Siranuhi, (Citrus unshiu × C. sinensis) × C. reticulata (peel) | Siranuhi | (Citrus unshiu × C. sinensis) × C. reticulata | Fresh peel |
| Smallanthus sonchifolius | Yacón tea | Smallanthus sonchifolius | Dried tea powder |
| Smallanthus sonchifolius (meat) | Yacón | Smallanthus sonchifolius | Fresh meat |
| Smallanthus sonchifolius (peel) | Yacón | Smallanthus sonchifolius | Fresh peel |
| Solanum melongena (peel) | Aubergine | Solanum melongena | Fresh peel |
| Vitis labruscana (peel) | Delaware grapes | Vitis labruscana | Fresh peel |
| Zanthoxylum bungeanum | Sichuan pepper | Zanthoxylum bungeanum | Dried powder |
| Zea mays (baby corn) | Baby corn | Zea mays | Fresh |
| Zea mays (kiritani) | Kiritani | Zea mays | Fresh |
| Zingiber mioga | Myoga | Zingiber mioga | Fresh |
| Zingiber officinale | Ginger | Zingiber officinale | Fresh |
| Descriptions in This Study | % * | Descriptions in This Study | % * | Descriptions in This Study | % * |
|---|---|---|---|---|---|
| Abelmoschus esculentus (beniokura) | 57.3 | Citrus natsudaidai (peel) | 74.5 | Matricaria recutita | 126.1 |
| Agaricus bisporus | 38.1 | Citrus paradisi (peel) | 92.4 | Matteuccia struthiopteris (young) | 116.6 |
| Allium cepa | 32.9 | Citrus reticulata (peel) | 50.2 | Mesembryanthemum crystallinum | 97.4 |
| Allium oschaninii | 59.5 | Citrus sinensis (blood orange, peel) | 65.2 | Momordica charantia (coat) | 97.8 |
| Allium sativum | 92.1 | Citrus sinensis (navel) | 81.4 | Musa spp. (peel) | 62.2 |
| Allium sativum (sprout) | 117.9 | Citrus sphaerocarpa (peel) | 119.1 | Musa spp. (peel, Ecuador) | 94.8 |
| Allium tuberosum | 77.7 | Citrus sudachi (peel) | 59.3 | Nasturtium officinale | 107.7 |
| Ananas comosus (coat) | 108.1 | Citrus tangelo (peel) | 34.7 | Nelumbo nucifera | 41.4 |
| Apium graveolens | 68.7 | Cocos nucifera (young) | 100.2 | Ocimum basilicum | 100.0 |
| Apium graveolens (salad celery) | 35.8 | Colocasia esculenta | 58.5 | Ocimum basilicum (purple) | 79.4 |
| Arachis hypogaea (beans) | 38.0 | Coriandrum sativum | 130.2 | Oryza sativa (black) | 67.8 |
| Arachis hypogaea (shell) | 24.3 | Coriandrum sativum (leaves) | 20.8 | Perilla frutescens | 81.6 |
| Aralia cordata | 43.9 | Cucumis melo (coat) | 67.3 | Persea americana (coat) | 106.7 |
| Aralia elata (sprout) | 86.4 | Cucurbita (meat) | 91.5 | Persea americana (seed) | 63.8 |
| Arctium lappa | 59.9 | Cucurbita (peel) | 66.5 | Petasites japonicus | 64.3 |
| Arctium lappa (burdock tea) | 97.4 | Cucurbita pepo (yellow, peel) | 37.8 | Petroselinum crispum (leaves) | 75.7 |
| Aspalathus linearis | 29.0 | Curcuma longa | 83.7 | Phaseolus vulgaris | 132.6 |
| Asparagus officinalis (grass roots) | 48.1 | Cyperus esculentus (powder) | 99.2 | Phaseolus vulgaris (Moroccan kidney beans) | 140.3 |
| Asparagus spp. | 77.0 | Daucus carota | 55.1 | Pholiota microspora | 55.4 |
| Auricularia auricula-judae | 31.4 | Daucus carota (purple carrot) | 52.0 | Phyllostachys pubescens (young, dried) | 47.4 |
| Barley (Hordeum vulgare) Miso | 105.5 | Dioscorea japonica | 44.8 | Pisum sativum | 62.2 |
| Basella alba | 63.5 | Diospyros kaki (shibugaki, meat) | 112.0 | Pisum sativum (shelled) | 107.8 |
| Benincasa hispida (coat, placenta, seed) | 50.3 | Diospyros kaki (shibugaki, peel) | 51.7 | Pisum sativum (shuck-edible) | 50.2 |
| Benincasa hispida (meat) | 25.2 | Eriobotrya japonica (peel) | 109.7 | Pisum sativum (young leaves) | 49.3 |
| Brassica chinensis | 93.8 | Eutrema japonicum | 73.7 | Pleurotus cornucopiae | 90.7 |
| Brassica oleracea (broccoli, anthotaxy) | 22.7 | Eutrema japonicum (stem) | 102.2 | Pleurotus eryngii | 49.9 |
| Brassica oleracea (broccoli, sprout) | 61.4 | Fagopyrum tataricum | 36.5 | Pleurotus ostreatus | 73.1 |
| Brassica oleracea (broccoli, stem) | 68.5 | Ficus carica | 79.1 | Prunus domestica (extract) | 84.4 |
| Brassica oleracea (kohlrabi, peel) | 73.4 | Flammulina velutipes | 39.7 | Prunus domestica (meat) | 96.3 |
| Brassica oleracea (red cabbage, sprout) | 106.2 | Fortunella (peel) | 62.5 | Prunus tomentosa (peel) | 49.5 |
| Brassica oleracea (romanesco broccoli, stem) | 109.8 | Fragaria ananassa | 52.0 | Psidium guajava (Chinese) | 73.4 |
| Brassica oleracea (soft kale) | 94.4 | Ginkgo biloba (seed) | 40.2 | Psidium guajava (Japanese) | 52.5 |
| Brassica oleracea (stick senor) | 51.2 | Glebionis coronaria | 17.9 | Psophocarpus tetragonolobus | 90.7 |
| Brassica oleracea (wild cabbage, flower) | 60.0 | Glycine max | 66.4 | Pteridium aquilinum | 192.2 |
| Brassica rapa (ayameyuki-kabu) | 108.2 | Glycine max (hidenmame) | 104.0 | Pyrus communis (peel) | 58.8 |
| Brassica rapa (ayameyuki-kabu, meat) | 85.2 | Glycine max (immature) | 64.6 | Raphanus sativus (leaves) | 70.8 |
| Brassica rapa (nabana) | 86.1 | Glycine max (immature, shuck) | 96.1 | Raphanus sativus (meat) | 64.7 |
| Brassica rapa (nabana, flower) | 64.2 | Glycine max × Bacillus subtilis | 60.6 | Raphanus sativus (radish sprout) | 79.6 |
| Brassica rapa (red) | 52.6 | Grifola frondosa | 69.5 | Rice (Oryza sativa) Miso | 58.6 |
| Brassica rapa (santo-sai) | 72.2 | Hibiscus rosa-sinensis | 97.5 | Rosmarinus officinalis (raw) | 13.6 |
| Capsicum annuum (redpepper) | 83.0 | Hosta sieboldiana | 45.2 | Sechium edule (meat) | 130.0 |
| Capsicum annuum (sweet pepper) | 107.8 | Houttuynia cordata | 84.0 | Sechium edule (peel, placenta) | 52.0 |
| Capsicum annuum (red) | 80.3 | Humulus lupulus (cone) | 78.9 | Sesamum indicum | 158.6 |
| Capsicum annuum (shishitou) | 73.0 | Hylocereus undatus (peel) | 120.0 | Siranuhi, (Citrus unshiu × C. sinensis) × C. reticulata (peel) | 52.7 |
| Capsicum annuum (yellow) | 81.4 | Hypsizygus marmoreus | 58.1 | Smallanthus sonchifolius | 35.4 |
| Capsicum frutescens | 58.8 | Ilex paraguariensis (roasted) | 69.1 | Smallanthus sonchifolius (meat) | 74.3 |
| Carica papaya (immature, meat) | 78.5 | Illicium verum | 24.6 | Smallanthus sonchifolius (peel) | 110.5 |
| Carica papaya (immature, peel, placenta, seed) | 94.3 | Ipomoea aquatica | 75.5 | Solanum melongena (peel) | 133.8 |
| Caulerpa lentillifera | 65.5 | Jasminum sambac | 66.5 | Vitis labruscana (peel) | 150.9 |
| Citrus aurantiifolia (peel) | 185.5 | Lactuca sativa | 91.6 | Zanthoxylum bungeanum | 57.2 |
| Citrus depressa (peel) | 38.9 | Laminaria longissima (tororomekonbu) | 112.2 | Zea mays (baby corn) | 65.3 |
| Citrus junos (peel) | 61.6 | Laurus nobilis (leaves) | 33.6 | Zea mays (kiritani) | 78.1 |
| Citrus maxima (peel) | 64.7 | Lentinula edodes | 62.8 | Zingiber mioga | 28.0 |
| Citrus maxima (placenta) | 68.9 | Lycopersicum esculentum (yellow) | 60.6 | Zingiber officinale | 57.3 |

References
- Dalbeth, N.; Merriman, T.R.; Stamp, L.K. Gout. Lancet 2016, 388, 2039–2052. [Google Scholar] [CrossRef]
- Dalbeth, N.; Choi, H.K.; Joosten, L.A.B.; Khanna, P.P.; Matsuo, H.; Perez-Ruiz, F.; Stamp, L.K. Gout. Nat. Rev. Dis. Primers 2019, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.W.; Muzny, D.M.; Lee, C.C.; Caskey, C.T. Two independent mutational events in the loss of urate oxidase during hominoid evolution. J. Mol. Evol. 1992, 34, 78–84. [Google Scholar] [CrossRef] [PubMed]
- Bobulescu, I.A.; Moe, O.W. Renal transport of uric acid: Evolving concepts and uncertainties. Adv. Chronic Kidney Dis. 2012, 19, 358–371. [Google Scholar] [CrossRef] [Green Version]
- Hyndman, D.; Liu, S.; Miner, J.N. Urate Handling in the Human Body. Curr. Rheumatol. Rep. 2016, 18, 34. [Google Scholar] [CrossRef] [Green Version]
- Enomoto, A.; Kimura, H.; Chairoungdua, A.; Shigeta, Y.; Jutabha, P.; Cha, S.H.; Hosoyamada, M.; Takeda, M.; Sekine, T.; Igarashi, T.; et al. Molecular identification of a renal urate anion exchanger that regulates blood urate levels. Nature 2002, 417, 447–452. [Google Scholar] [CrossRef]
- Kawamura, Y.; Toyoda, Y.; Ohnishi, T.; Hisatomi, R.; Higashino, T.; Nakayama, A.; Shimizu, S.; Yanagi, M.; Kamimaki, I.; Fujimaru, R.; et al. Identification of a dysfunctional splicing mutation in the SLC22A12/URAT1 gene causing renal hypouricaemia type 1: A report on two families. Rheumatology 2020, 59, 3988–3990. [Google Scholar] [CrossRef]
- Nakayama, A.; Matsuo, H.; Ohtahara, A.; Ogino, K.; Hakoda, M.; Hamada, T.; Hosoyamada, M.; Yamaguchi, S.; Hisatome, I.; Ichida, K.; et al. Clinical practice guideline for renal hypouricemia (1st edition). Hum. Cell 2019, 32, 83–87. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, A.; Kawamura, Y.; Toyoda, Y.; Shimizu, S.; Kawaguchi, M.; Aoki, Y.; Takeuchi, K.; Okada, R.; Kubo, Y.; Imakiire, T.; et al. Genetic-epidemiological analysis of hypouricemia from 4,993 Japanese on nonfunctional variants of URAT1/SLC22A12 gene. Rheumatology 2021. [Google Scholar] [CrossRef]
- Miner, J.N.; Tan, P.K.; Hyndman, D.; Liu, S.; Iverson, C.; Nanavati, P.; Hagerty, D.T.; Manhard, K.; Shen, Z.; Girardet, J.L.; et al. Lesinurad, a novel, oral compound for gout, acts to decrease serum uric acid through inhibition of urate transporters in the kidney. Arthritis Res. Ther. 2016, 18, 214. [Google Scholar] [CrossRef] [Green Version]
- Toyoda, Y.; Takada, T.; Saito, H.; Hirata, H.; Ota-Kontani, A.; Kobayashi, N.; Tsuchiya, Y.; Suzuki, H. Inhibitory effect of Citrus flavonoids on the in vitro transport activity of human urate transporter 1 (URAT1/SLC22A12), a renal re-absorber of urate. NPJ Sci. Food 2020, 4, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tashiro, Y.; Sakai, R.; Hirose-Sugiura, T.; Kato, Y.; Matsuo, H.; Takada, T.; Suzuki, H.; Makino, T. Effects of Osthol Isolated from Cnidium monnieri Fruit on Urate Transporter 1. Molecules 2018, 23, 2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Fong, W.P.; Cheng, C.H. Morin (3,5,7,2′,4′-pentahydroxyflavone) exhibits potent inhibitory actions on urate transport by the human urate anion transporter (hURAT1) expressed in human embryonic kidney cells. Drug Metab. Dispos. Biol. Fate Chem. 2007, 35, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Saito, H.; Toyoda, Y.; Takada, T.; Hirata, H.; Ota-Kontani, A.; Miyata, H.; Kobayashi, N.; Tsuchiya, Y.; Suzuki, H. Omega-3 Polyunsaturated Fatty Acids Inhibit the Function of Human URAT1, a Renal Urate Re-Absorber. Nutrients 2020, 12, 1601. [Google Scholar] [CrossRef]
- Toyoda, Y.; Kawamura, Y.; Nakayama, A.; Nakaoka, H.; Higashino, T.; Shimizu, S.; Ooyama, H.; Morimoto, K.; Uchida, N.; Shigesawa, R.; et al. Substantial anti-gout effect conferred by common and rare dysfunctional variants of URAT1/SLC22A12. Rheumatology 2021, 60, 5224–5232. [Google Scholar] [CrossRef]
- Saito, H.; Toyoda, Y.; Hirata, H.; Ota-Kontani, A.; Tsuchiya, Y.; Takada, T.; Suzuki, H. Soy Isoflavone Genistein Inhibits an Axillary Osmidrosis Risk Factor ABCC11: In Vitro Screening and Fractional Approach for ABCC11-Inhibitory Activities in Plant Extracts and Dietary Flavonoids. Nutrients 2020, 12, 2452. [Google Scholar] [CrossRef]
- Hirata, H.; Takazumi, K.; Segawa, S.; Okada, Y.; Kobayashi, N.; Shigyo, T.; Chiba, H. Xanthohumol, a prenylated chalcone from Humulus lupulus L., inhibits cholesteryl ester transfer protein. Food Chem. 2012, 134, 1432–1437. [Google Scholar] [CrossRef]
- Miyata, H.; Takada, T.; Toyoda, Y.; Matsuo, H.; Ichida, K.; Suzuki, H. Identification of Febuxostat as a New Strong ABCG2 Inhibitor: Potential Applications and Risks in Clinical Situations. Front Pharmacol. 2016, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- McKay, D.L.; Blumberg, J.B. A review of the bioactivity of South African herbal teas: Rooibos (Aspalathus linearis) and honeybush (Cyclopia intermedia). Phytother Res. 2007, 21, 1–16. [Google Scholar] [CrossRef]
- Shimamura, N.; Miyase, T.; Umehara, K.; Warashina, T.; Fujii, S. Phytoestrogens from Aspalathus linearis. Biol. Pharm. Bull. 2006, 29, 1271–1274. [Google Scholar] [CrossRef] [Green Version]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vauzour, D.; Vafeiadou, K.; Rodriguez-Mateos, A.; Rendeiro, C.; Spencer, J.P. The neuroprotective potential of flavonoids: A multiplicity of effects. Genes Nutr. 2008, 3, 115–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, D.; Garg, V.K.; Tuli, H.S.; Yerer, M.B.; Sak, K.; Sharma, A.K.; Kumar, M.; Aggarwal, V.; Sandhu, S.S. Fisetin and Quercetin: Promising Flavonoids with Chemopreventive Potential. Biomolecules 2019, 9, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundarraj, K.; Raghunath, A.; Perumal, E. A review on the chemotherapeutic potential of fisetin: In vitro evidences. Biomed. Pharmacother. 2018, 97, 928–940. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Shi, Y.; Williamson, G. Quercetin lowers plasma uric acid in pre-hyperuricaemic males: A randomised, double-blinded, placebo-controlled, cross-over trial. Br. J. Nutr. 2016, 115, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Day, A.J.; Bao, Y.; Morgan, M.R.A.; Williamson, G. Conjugation position of quercetin glucuronides and effect on biological activity. Free Radic. Biol. Med. 2000, 29, 1234–1243. [Google Scholar] [CrossRef]
- Joubert, E.; de Beer, D. Rooibos (Aspalathus linearis) beyond the farm gate: From herbal tea to potential phytopharmaceutical. S. Afr. J. Bot. 2011, 77, 869–886. [Google Scholar] [CrossRef]
- Kondo, M.; Hirano, Y.; Nishio, M.; Furuya, Y.; Nakamura, H.; Watanabe, T. Xanthine oxidase inhibitory activity and hypouricemic effect of aspalathin from unfermented rooibos. J. Food Sci. 2013, 78, H1935–H1939. [Google Scholar] [CrossRef]
- Jiang, L.L.; Gong, X.; Ji, M.Y.; Wang, C.C.; Wang, J.H.; Li, M.H. Bioactive Compounds from Plant-Based Functional Foods: A Promising Choice for the Prevention and Management of Hyperuricemia. Foods 2020, 9, 973. [Google Scholar] [CrossRef]








| REAGENT or RESOURCE | SOURCE | IDENTIFIER |
|---|---|---|
| Chemicals | ||
| Clear-sol II | Nacalai Tesque | Cat# 09136-83 |
| [8-14C]-Uric acid (53 mCi/mmol) | American Radiolabeled Chemicals | Cat# ARC0513 |
| Dimethyl Sulfoxide | Nacalai Tesque | Cat# 13445-74; CAS: 67-68-5 |
| Ethanol | FUJIFILM Wako Pure Chemical | 057-00451; CAS: 64-17-5 |
| Methanol | FUJIFILM Wako Pure Chemical | 137-01823; CAS: 67-56-1 |
| n-Hexane | FUJIFILM Wako Pure Chemical | 085-00416; CAS: 110-54-3 |
| Ethyl acetate | FUJIFILM Wako Pure Chemical | 051-00356; CAS: 141-78-6 |
| n-Buthanol | FUJIFILM Wako Pure Chemical | 026-03326; CAS: 71-36-3 |
| Polyethelenimine “MAX” | Polysciences | Cat# 24765; CAS: 49553-93-7 |
| 2′-Hydroxychalcone | Tokyo Chemical Industry | Cat# H0385; CAS: 1214-47-7; Purity: >98% |
| 3-Hydroxyflavone | Tokyo Chemical Industry | Cat# H0379; CAS: 577-85-5; Purity: ≥98% |
| 4-Hydroxychalcone | Tokyo Chemical Industry | Cat# H0955; CAS: 20426-12-4; Purity: >96% |
| 4′-Hydroxychalcone | Tokyo Chemical Industry | Cat# H0945; CAS: 2657-25-2; Purity: >95% |
| Aesculetin | FUJIFILM Wako Pure Chemical | Cat# A15393; CAS: 305-01-1; Purity: N/A |
| Apigenin | FUJIFILM Wako Pure Chemical | Cat# 016-18911; CAS: 520-36-5; Purity: ≥95% |
| Cardamonin | R&D systems | Cat# 2509/10; CAS: 19309-14-9; Purity: ≥98% |
| Daidzein | FUJIFILM Wako Pure Chemical | Cat# 043-28071; CAS: 486-66-8; Purity: ≥98% |
| Dihydromyricetin | EXTRASYNTHESE | Cat# 1351-10 mg; CAS: 27200-12-0; Purity: ≥95% |
| Fisetin | LKT Labs | Cat# F3473; CAS: 528-48-3; Purity: ≥97% |
| Galangin | ChromaDex | Cat# ASB-00007030-010; CAS: 548-83-4; Purity: N/A |
| Genistein | FUJIFILM Wako Pure Chemical | Cat# 073-05531; CAS: 446-72-0; Purity: ≥98% |
| Gossypetin | ChromaDex | Cat# ASB-00007390-010; CAS: 489-35-0; Purity: N/A |
| Isoliquiritigenin | Tokyo Chemical Industry | Cat# I0822; CAS: 961-29-5; Purity: ≥97% |
| Kaempferol | FUJIFILM Wako Pure Chemical | Cat# 110-00451; CAS: 520-18-3; Purity: ≥95% |
| Luteolin | Cayman Chemical | Cat# 10004161; CAS: 491-70-3; Purity: ≥98% |
| Morin | Combi-Blocks | Cat# QC-0527; CAS: 480-16-0; Purity: ≥98% |
| Myricetin | FUJIFILM Wako Pure Chemical | Cat# 137-16791; CAS: 529-44-2; Purity: ≥98% |
| Naringenin chalcone | ChromaDex | Cat# ASB-00014207-005; CAS: 73692-50-9; Purity: N/A |
| Phloretin | FUJIFILM Wako Pure Chemical | Cat# 160-17781; CAS: 60-82-2; Purity: ≥98% |
| Quercetagetin | ChromaDex | Cat# ASB-00017020-005; CAS: 90-18-6; Purity: N/A |
| Quercetin | ChromaDex | Cat# ASB-00017030-010; CAS: 117-39-5: Purity: ≥97% |
| Taxifolin | EXTRASYNTHESE | Cat# 1036; CAS: 17654-26-1; Purity: N/A |
| Xanthohumol | TOKIWA PHYTOCHEMICAL | Cat# P2217; CAS: 569-83-5; Purity: ≥98% |
| Critical Commercial Assays | ||
| Pierce BCA Protein Assay Reagent A, B | Thermo Fisher Scientific | Cat# 23223, Cat# 23224 |
| PureLink HiPure Plasmid Filter Midiprep Kit | Thermo Fisher Scientific | Cat# K210015 |
| Recombinant DNA | ||
| The complete human URAT1 cDNA in pEGFP-C1 | Saito et al. 2020 [14] | NCBI Reference Sequence: NM_144585.3 |
| Experimental Models: Cell Lines | ||
| 293A | Invitrogen | R70507 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toyoda, Y.; Takada, T.; Saito, H.; Hirata, H.; Ota-Kontani, A.; Tsuchiya, Y.; Suzuki, H. Identification of Inhibitory Activities of Dietary Flavonoids against URAT1, a Renal Urate Re-Absorber: In Vitro Screening and Fractional Approach Focused on Rooibos Leaves. Nutrients 2022, 14, 575. https://doi.org/10.3390/nu14030575
Toyoda Y, Takada T, Saito H, Hirata H, Ota-Kontani A, Tsuchiya Y, Suzuki H. Identification of Inhibitory Activities of Dietary Flavonoids against URAT1, a Renal Urate Re-Absorber: In Vitro Screening and Fractional Approach Focused on Rooibos Leaves. Nutrients. 2022; 14(3):575. https://doi.org/10.3390/nu14030575
Chicago/Turabian StyleToyoda, Yu, Tappei Takada, Hiroki Saito, Hiroshi Hirata, Ami Ota-Kontani, Youichi Tsuchiya, and Hiroshi Suzuki. 2022. "Identification of Inhibitory Activities of Dietary Flavonoids against URAT1, a Renal Urate Re-Absorber: In Vitro Screening and Fractional Approach Focused on Rooibos Leaves" Nutrients 14, no. 3: 575. https://doi.org/10.3390/nu14030575