
Evaluation of the Cholesterol-Lowering Mechanism of Enterococcus faecium Strain 132 and Lactobacillus paracasei Strain 201 in Hypercholesterolemia Rats

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cholesterol-Lowering Activity In Vitro
2.2. Animal and Study Design
2.3. Serum Biochemical Index Analysis
2.4. Liver and Faeces TC, TBA and TG Contents
2.5. Determination of Related Inflammatory Factors in Liver and Epididymal Fat
2.6. Histopathological Examination
2.7. Real-Time Fluorescence Quantitative PCR (RT-qPCR)
2.8. Intestinal Microbial Diversity
2.9. Determination of Short Chain Fatty Acids (SCFAs)
2.10. Statistical Analysis
3. Results
3.1. Cholesterol-Lowering Ability of Selected Strains
3.2. Effects of Screened Strains on Serum Parameters in of Hypercholesterolemic Rats
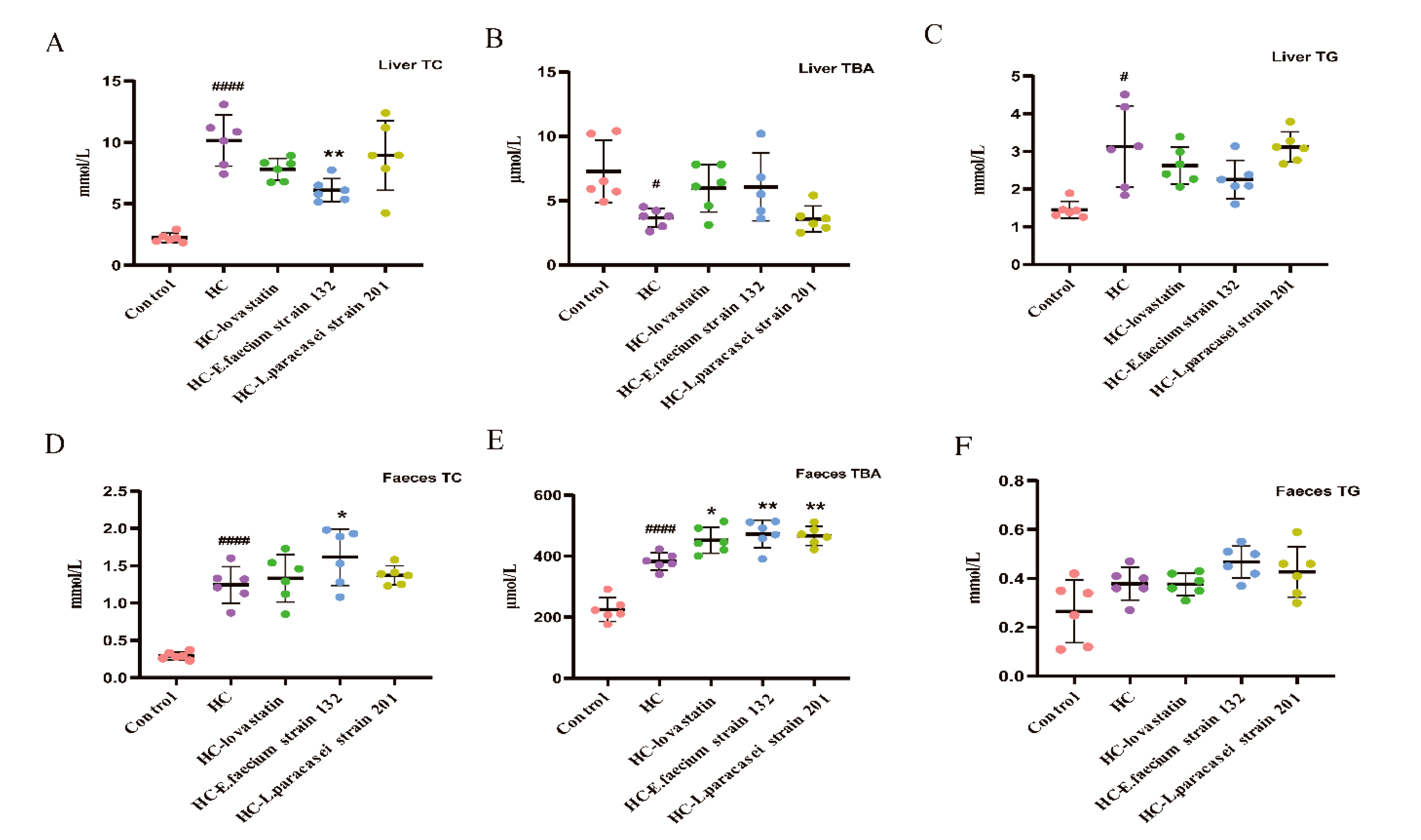
3.3. Liver and Faecal TC, TBA and TG Levels in Hypercholesterolemic Rats
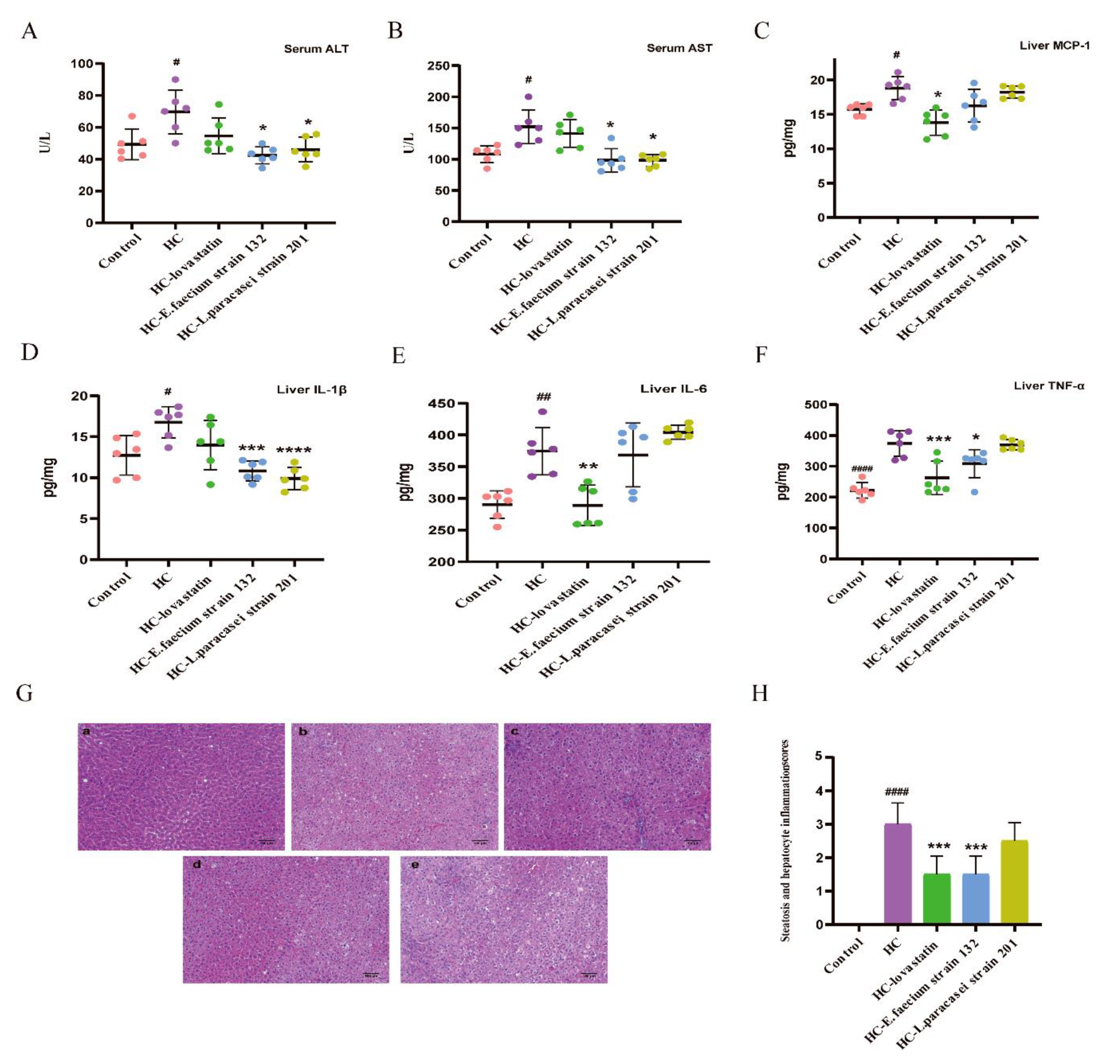
3.4. Screening Strains Improved Liver Injury in Hypercholesterolemia Rats
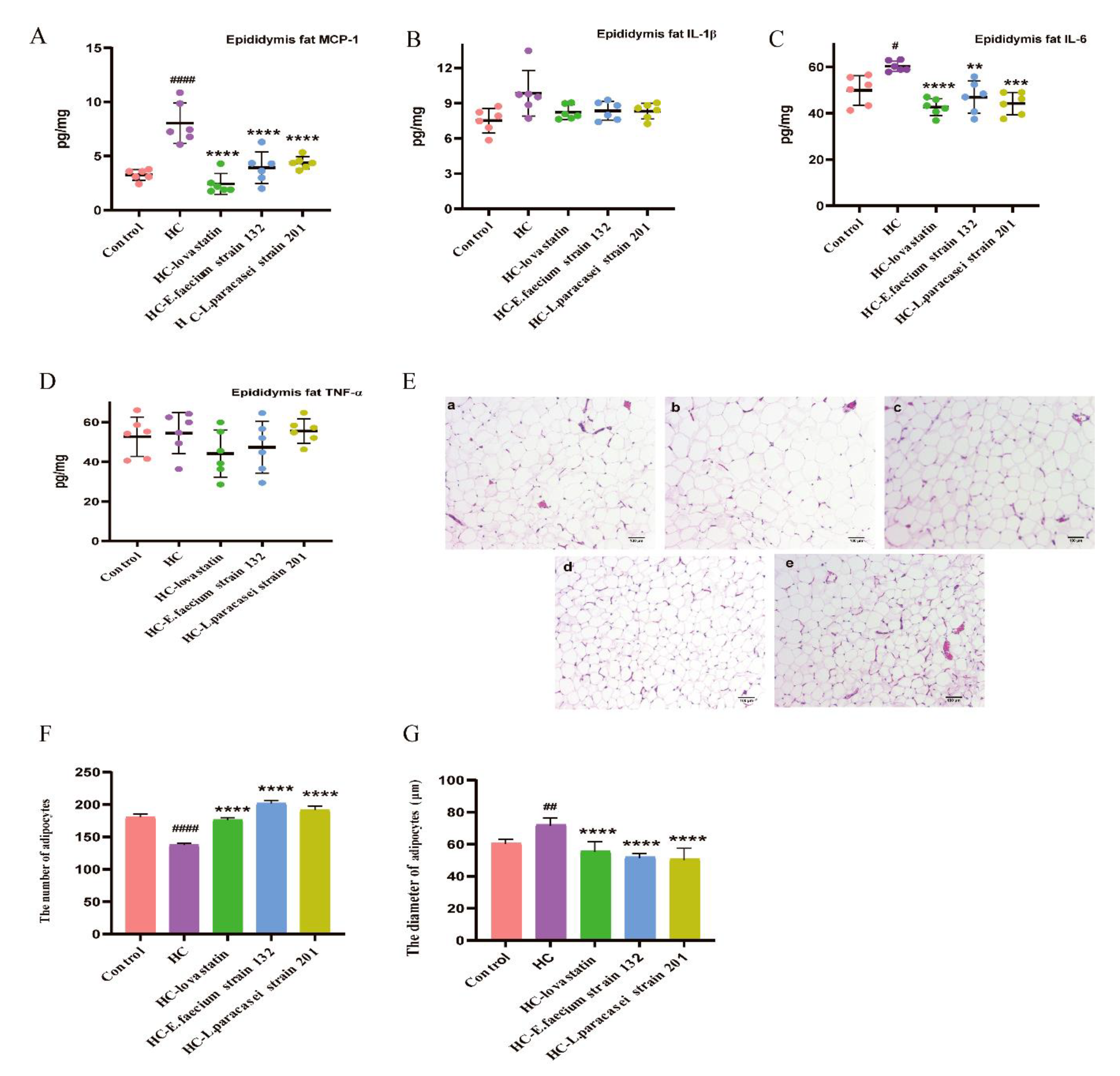
3.5. Screened Strains Improved the Inflammation of Epididymis Fat and Fatty Hypertrophy in Hypercholesterolemic Rats
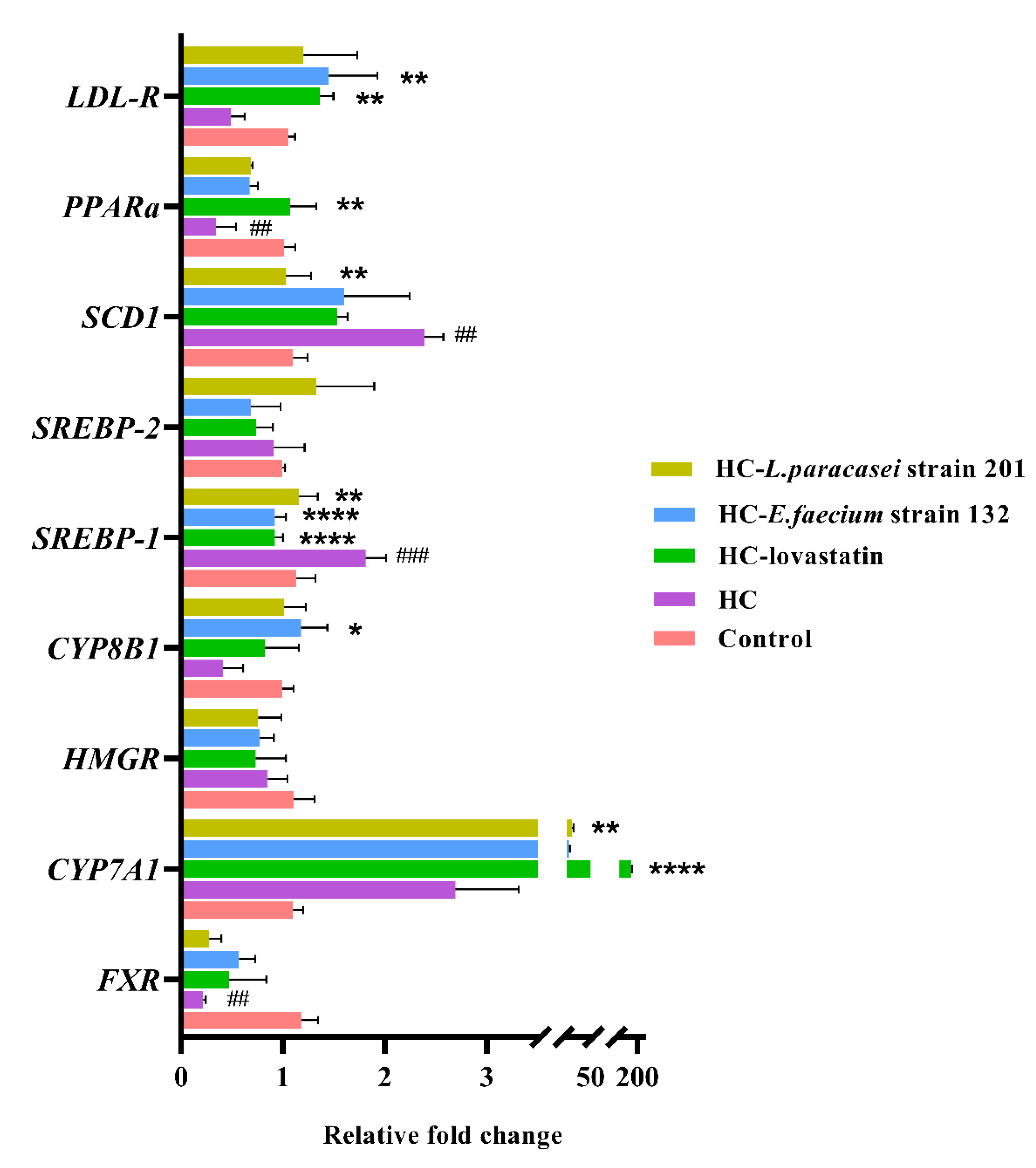
3.6. mRNA Expression Levels of Genes Related to Lipid Metabolism
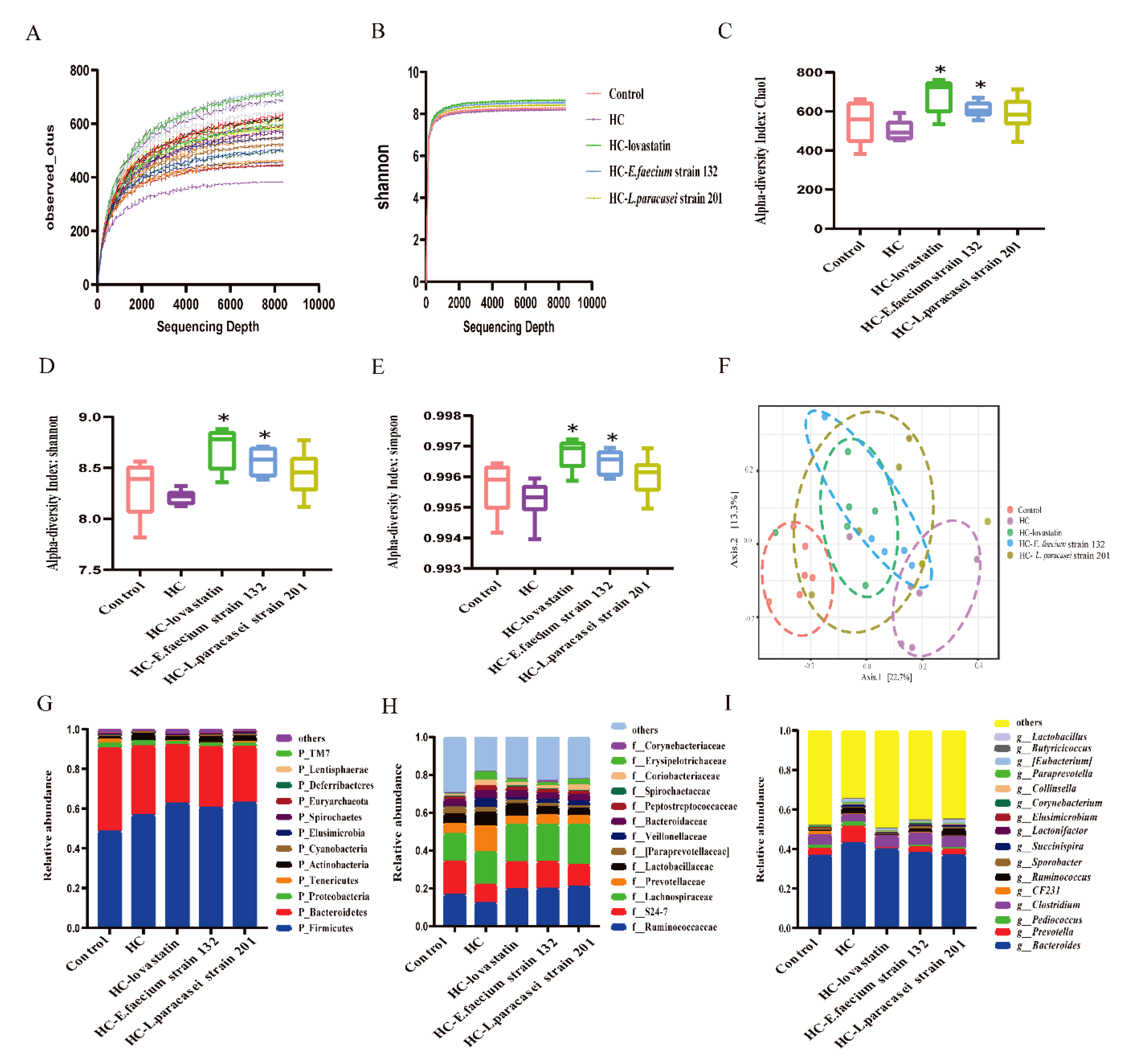
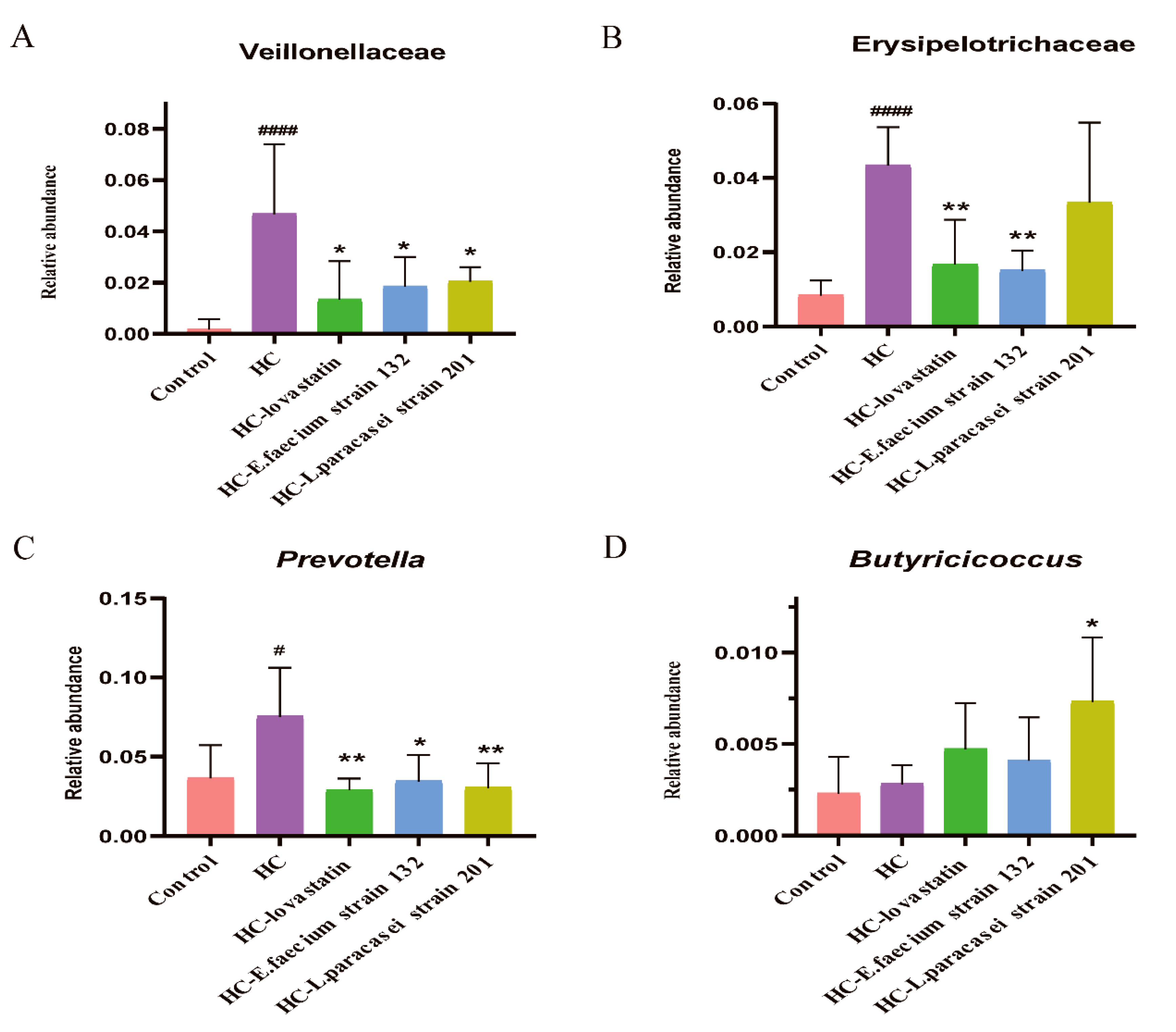
3.7. Gut Microbiota Modulation by Selected Strains
3.8. SCFA Content in Faeces in Hypercholesterolaemic Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benjamin, E.J.; Muntner, P.; Alonso, A.; Bittencourt, M.S.; Callaway, C.W.; Carson, A.P.; Chamberlain, A.M.; Chang, A.R.; Cheng, S.; Das, S.R.; et al. Heart Disease and Stroke Statistics-2019 Update: A Report From the American Heart Association. Circulation 2019, 139, e56–e528. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Nagpal, R.; Kumar, R.; Hemalatha, R.; Verma, V.; Kumar, A.; Chakraborty, C.; Singh, B.; Marotta, F.; Jain, S.; et al. Cholesterol-lowering probiotics as potential biotherapeutics for metabolic diseases. Exp. Diabetes Res. 2012, 2012, 902917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimwe, N.; Daliri, E.B.; Lee, B.H.; Fang, F.; Du, G. The perspective on cholesterol-lowering mechanisms of probiotics. Mol. Nutr. Food Res. 2015, 59, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [Green Version]
- Feingold, K.R. Cholesterol Lowering Drugs. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Grossman, A., Hershman, J.M., Hofland, J., et al., Eds.; Endotext [Internet]: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Sultan, S.; Hynes, N. The Ugly Side of Statins. Systemic Appraisal of the Contemporary Un-Known Unknowns. Open J. Endocr. Metab. Dis. 2013, 3, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Sanders, M.E. Probiotics: Definition, sources, selection, and uses. Clin. Infect. Dis. 2008, 46 (Suppl. S2), S58–S61; discussion S144–S151. [Google Scholar] [CrossRef] [Green Version]
- Napoli, C.; Crudele, V.; Soricelli, A.; Al-Omran, M.; Vitale, N.; Infante, T.; Mancini, F.P. Primary prevention of atherosclerosis: A clinical challenge for the reversal of epigenetic mechanisms? Circulation 2012, 125, 2363–2373. [Google Scholar] [CrossRef] [Green Version]
- Hlivak, P.; Odraska, J.; Ferencik, M.; Ebringer, L.; Jahnova, E.; Mikes, Z. One-year application of probiotic strain Enterococcus faecium M-74 decreases serum cholesterol levels. Bratisl. Lek. Listy 2005, 106, 67–72. [Google Scholar]
- Zhang, F.; Qiu, L.; Xu, X.; Liu, Z.; Zhan, H.; Tao, X.; Shah, N.P.; Wei, H. Bene fi cial effects of probiotic cholesterol-lowering strain of Enterococcus faecium WEFA23 from infants on diet-induced metabolic syndrome in rats. J. Dairy Sci. 2017, 100, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.L.; Martoni, C.J.; Prakash, S. Cholesterol lowering and inhibition of sterol absorption by Lactobacillus reuteri NCIMB 30242: A randomized controlled trial. Eur. J. Clin. Nutr. 2012, 66, 1234–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, M.; Darimont, C.; Drapeau, V.; Emady-Azar, S.; Lepage, M.; Rezzonico, E.; Ngom-Bru, C.; Berger, B.; Philippe, L.; Ammon-Zuffrey, C.; et al. Effect of Lactobacillus rhamnosus CGMCC1.3724 supplementation on weight loss and maintenance in obese men and women. Br. J. Nutr. 2014, 111, 1507–1519. [Google Scholar] [CrossRef] [Green Version]
- Bernini, L.J.; Simao, A.N.C.; de Souza, A.H.B.; Alfieri, D.F.; Segura, L.G.; Costa, G.N.; Dichi, I. Effect of Bifidobacterium lactis HN019 on inflammatory markers and oxidative stress in subjects with and without the metabolic syndrome. Brit. J. Nutr. 2018, 120, 645–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lan, T.; Haywood, J.; Dawson, P.A. Inhibition of ileal apical but not basolateral bile acid transport reduces atherosclerosis in apoE−/− mice. Atherosclerosis 2013, 229, 374–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesaro, C.; Tiso, A.; Del Prete, A.; Cariello, R.; Tuccillo, C.; Cotticelli, G.; Del Vecchio Blanco, C.; Loguercio, C. Gut microbiota and probiotics in chronic liver diseases. Dig. Liver Dis. 2011, 43, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, F.; Dashty, M. Role of Adipose Tissue in Metabolic System Disorders. J. Diabetes Metab. 2013, 1. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Chung, H.J.; Lee, I.A.; D’Souza, R.; Kim, H.J.; Hong, S.T. Elucidation of the anti-hyperammonemic mechanism of Lactobacillus amylovorus JBD401 by comparative genomic analysis. BMC Genomics 2018, 19, 292. [Google Scholar] [CrossRef] [Green Version]
- Falcinelli, S.; Rodiles, A.; Hatef, A.; Picchietti, S.; Cossignani, L.; Merrifield, D.L.; Unniappan, S.; Carnevali, O. Influence of Probiotics Administration on Gut Microbiota Core: A Review on the Effects on Appetite Control, Glucose, and Lipid Metabolism. J. Clin. Gastroenterol. 2018, 52, S50–S56. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Q.; Wang, J.; Li, P.; Cheng, L.; An, Y.; Duan, Y.; Dai, H.; Wang, T.; Zhao, B. Relationship between hyperlipidemia and the gut microbiome of rats, characterized using high-throughput sequencing. J. Tradit. Chin. Med. Sci. 2020, 7, 154–161. [Google Scholar] [CrossRef]
- Sun, J.; Kato, I. Gut microbiota, inflammation and colorectal cancer. Genes Dis. 2016, 3, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2014, 7, 17–44. [Google Scholar] [CrossRef]
- Knip, M.; Siljander, H. The role of the intestinal microbiota in type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2016, 12, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Wu, B.H.; Chu, Y.L.; Chang, W.C.; Wu, M.C. Effects of Tempeh Fermentation with Lactobacillus plantarum and Rhizopus oligosporus on Streptozotocin-Induced Type II Diabetes Mellitus in Rats. Nutrients 2018, 10, 1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Guo, Y.; Chen, H.; Wei, H.; Wan, C. Potential of Lactobacillus plantarum ZDY2013 and Bifidobacterium bifidum WBIN03 in relieving colitis by gut microbiota, immune, and anti-oxidative stress. Can J. Microbiol. 2018, 64, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Carvajal-Zarrabal, O.; Waliszewski, S.M.; Barradas-Dermitz, D.M.; Orta-Flores, Z.; Hayward-Jones, P.M.; Nolasco-Hipolito, C.; Angulo-Guerrero, O.; Sanchez-Ricano, R.; Infanzon, R.M.; Trujillo, P.R. The consumption of Hibiscus sabdariffa dried calyx ethanolic extract reduced lipid profile in rats. Plant Foods Hum. Nutr. 2005, 60, 153–159. [Google Scholar] [CrossRef]
- Wang, B.; Kong, Q.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. A High-Fat Diet Increases Gut Microbiota Biodiversity and Energy Expenditure Due to Nutrient Difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef]
- Lepercq, P.; Relano, P.; Cayuela, C.; Juste, C. Bifidobacterium animalis strain DN-173 010 hydrolyses bile salts in the gastrointestinal tract of pigs. Scand. J. Gastroenterol. 2004, 39, 1266–1271. [Google Scholar] [CrossRef]
- Kajiura, K.; Ohkusa, T.; Okayasu, I. Relationship between fecal bile acids and the occurrence of colorectal neoplasia in experimental murine ulcerative colitis. Digestion 1998, 59, 69–72. [Google Scholar] [CrossRef]
- Junatas, K.L.; Tonar, Z.; Kubikova, T.; Liska, V.; Palek, R.; Mik, P.; Kralickova, M.; Witter, K. Stereological analysis of size and density of hepatocytes in the porcine liver. J. Anat. 2017, 230, 575–588. [Google Scholar] [CrossRef] [Green Version]
- Kleiner, D.E.; Brunt, E.M.; Van Natta, M.; Behling, C.; Contos, M.J.; Cummings, O.W.; Ferrell, L.D.; Liu, Y.C.; Torbenson, M.S.; Unalp-Arida, A.; et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology 2005, 41, 1313–1321. [Google Scholar] [CrossRef]
- Wu, Y.; Li, X.; Tan, F.; Zhou, X.; Mu, J.; Zhao, X. Lactobacillus fermentum CQPC07 attenuates obesity, inflammation and dyslipidemia by modulating the antioxidant capacity and lipid metabolism in high-fat diet induced obese mice. J. Inflamm. 2021, 18, 5. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Buttarazzi, I.; Kolida, S.; Quercia, S.; Baldini, J.; Swann, J.R.; Brigidi, P.; Gibson, G.R. An in vivo assessment of the cholesterol-lowering efficacy of Lactobacillus plantarum ECGC 13110402 in normal to mildly hypercholesterolaemic adults. PLoS ONE 2017, 12, e0187964. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, M.; Houten, S.M.; Wang, L.; Moschetta, A.; Mangelsdorf, D.J.; Heyman, R.A.; Moore, D.D.; Auwerx, J. Bile acids lower triglyceride levels via a pathway involving FXR, SHP, and SREBP-1c. J. Clin. Investig. 2004, 113, 1408–1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.Y.; Cho, Y.K.; Kang, M.S.; Yoo, T.W.; Park, J.H.; Kim, H.J.; Park, D.I.; Sohn, C.I.; Jeon, W.K.; Kim, B.I.; et al. The association between increased alanine aminotransferase activity and metabolic factors in nonalcoholic fatty liver disease. Metab. Clin. Exp. 2006, 55, 1604–1609. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Yang, L.; Tian, L.; Li, W.; Yang, L.; Li, L. Macrophage Migration Inhibitor Factor Upregulates MCP-1 Expression in an Autocrine Manner in Hepatocytes during Acute Mouse Liver Injury. Sci. Rep. 2016, 6, 27665. [Google Scholar] [CrossRef] [Green Version]
- Bajt, M.L.; Farhood, A.; Jaeschke, H. Effects of CXC chemokines on neutrophil activation and sequestration in hepatic vasculature. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 281, G1188–G1195. [Google Scholar] [CrossRef]
- Yu, R.; Kim, C.S.; Kang, J.H. Inflammatory Components of Adipose Tissue as Target for Treatment of Metabolic Syndrome. Forum Nutr. 2009, 61, 95–103. [Google Scholar] [CrossRef]
- Dulloo, A.G.; Montani, J.P. Body composition, inflammation and thermogenesis in pathways to obesity and the metabolic syndrome: An overview. Obes. Rev. 2012, 13 (Suppl. 2), 1–5. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.S.; Lim, W.C.; Lee, S.J.; Lee, S.H.; Lee, J.H.; Cho, H.Y. Antiobesity Effect of Garlic Extract Fermented by Lactobacillus plantarum BL2 in Diet-Induced Obese Mice. J. Med. Food 2016, 19, 823–829. [Google Scholar] [CrossRef]
- Goulet, O. Potential role of the intestinal microbiota in programming health and disease. Nutr. Rev. 2015, 73, 32–40. [Google Scholar] [CrossRef]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K.; et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef]
- Alard, J.; Lehrter, V.; Rhimi, M.; Mangin, I.; Peucelle, V.; Abraham, A.L.; Mariadassou, M.; Maguin, E.; Waligora-Dupriet, A.J.; Pot, B.; et al. Beneficial metabolic effects of selected probiotics on diet-induced obesity and insulin resistance in mice are associated with improvement of dysbiotic gut microbiota. Environ. Microbiol. 2016, 18, 1484–1497. [Google Scholar] [CrossRef]
- Martinez, I.; Perdicaro, D.J.; Brown, A.W.; Hammons, S.; Carden, T.J.; Carr, T.P.; Eskridge, K.M.; Walter, J. Diet-induced alterations of host cholesterol metabolism are likely to affect the gut microbiota composition in hamsters. Appl. Environ. Microbiol. 2013, 79, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Perez, O.; Cruz-Ramon, V.; Chinchilla-Lopez, P.; Mendez-Sanchez, N. The Role of the Gut Microbiota in Bile Acid Metabolism. Ann. Hepatol. 2017, 16 (Suppl. 1), S21–S26. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Qi, C.; Zhu, H.; Yu, R.; Xie, C.; Peng, Y.; Yin, S.W.; Fan, J.; Zhao, S.; Sun, J. Lactobacillus reuteri improves gut barrier function and affects diurnal variation of the gut microbiota in mice fed a high-fat diet. Food Funct. 2019, 10, 4705–4715. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Oligonucleotide Sequence (5′-3′) |
|---|---|
| CYP7A1 F | GCCTTCCTATTCACTTGTTC |
| CYP7A1 R | GTGGAGAGCGTGTCATTG |
| CYP8B1 F | CCTCTTCCACTTCTGCTAC |
| CYP8B1 R | GTCCTGCTCCTTGTCCTT |
| HMGR F | GGTGCGAAGTTCCTTAGTGAT |
| HMGR R | ATGAGGGTTTCCAGTTTGTAGG |
| SREBP-2 F | CAGCAGCAGACGGTGATGA |
| SREBP-2 R | TGGTTGCGGCATTCTGGTAT |
| SREBP-1 F | ACACAGACAAACTGCCCATC |
| SREBP-1 R | TCATTGATAGAGGAACGGTAGC |
| PPARα F | CTTCACGATGCTGTCCTCCT |
| PPARα R | GATGTCGCAGAATGGCTTCC |
| LDL_R F | TGTGGCAGTAGTGAGTGT |
| LDL_R R | GTTCTCCTCGTCCGACTT |
| FXR F | TCCTCCTCGTCCTATTATTCCA |
| FXR R | GCATTCGCCTGAGTTCATAGA |
| SCD1 F | TCCTACACGACCACCACTAC |
| SCD1 R | GGCACCTTCTTCATCTTCTCT |
| Parameters | Groups | ||||
|---|---|---|---|---|---|
| Control | HC | HC-Lovastatin | HC-E. faecium Strain 132 | HC-L. paracasei Strain 201 | |
| Weigh gained (g/6 weeks) | 155.02 ± 32.60 | 184.31 ± 26.32 ### | 147.03 ± 30.68 *** | 156.91 ± 20.57 ** | 165.14 ± 22.65 ** |
| Cardiac index | 0.29 ± 0.03 | 0.30 ± 0.03 | 0.29 ± 0.04 | 0.31 ± 0.03 | 0.30 ± 0.02 |
| Liver index | 2.50 ± 0.35 | 3.79 ± 0.60 ### | 3.11 ± 0.25 ** | 3.10 ± 0.51 * | 2.95 ± 0.46 ** |
| Spleen index | 0.19 ± 0.02 | 0.21 ± 0.09 | 0.18 ± 0.04 | 0.19 ± 0.04 | 0.22 ± 0.09 |
| Kidney index | 0.60 ± 0.06 | 0.57 ± 0.07 | 0.54 ± 0.08 | 0.56 ± 0.05 | 0.55 ± 0.08 |
| Epididymal fat index | 0.99 ± 0.18 | 1.41 ± 0.29 ## | 0.98 ± 0.19 * | 1.08 ± 0.12 * | 0.99 ± 0.14 * |
| TBA(μmol/L) | 13.98 ± 5.81 | 14.52 ± 5.99 | 13.51 ± 7.78 | 13.97 ± 3.92 | 14.47 ± 8.15 |
| TG (mmol/L) | 0.33 ± 0.11 | 0.63 ± 0.12 ### | 0.24 ± 0.09 **** | 0.38 ± 0.08 ** | 0.40 ± 0.04 ** |
| LDL-C(mmol/L) | 0.27 ± 0.04 | 0.93 ± 0.06 #### | 0.67 ± 0.22 ** | 0.71 ± 0.11 * | 0.69 ± 0.05 * |
| HDL-C(mmol/L) | 0.46 ± 0.19 | 0.26 ± 0.04 ## | 0.25 ± 0.07 | 0.29 ± 0.05 | 0.28 ± 0.05 |
| TC (mmol/L) | 1.42 ± 0.23 | 1.91 ± 0.70 # | 1.63 ± 0.53 | 1.71 ± 0.15 | 1.69 ± 0.31 |
| Short Chain Fatty Acid | Control | HC | HC-Lovastatin | HC-E. faecium Strain 132 | HC-L. paracasei Strain 201 |
|---|---|---|---|---|---|
| Acetic acid | 3.01 ± 0.36 | 2.42 ± 0.81 | 3.42 ± 0.04 * | 3.60 ± 0.12 * | 3.30 ± 0.19 |
| Propionic acid | 0.95 ± 0.10 | 0.81 ± 0.28 | 1.21 ± 0.05 * | 1.38 ± 0.12 ** | 1.17 ± 0.05 * |
| Isobutyric acid | 0.32 ± 0.01 | 0.37 ± 0.14 | 0.31 ± 0.01 | 0.31 ± 0.01 | 0.31 ± 0.00 |
| Butyric acid | 0.64 ± 0.02 | 0.57 ± 0.16 | 0.57 ± 0.01 | 0.60 ± 0.02 | 0.60 ± 0.02 |
| Isovaleric acid | 0.33 ± 0.00 | 0.33 ± 0.01 | 0.33 ± 0.00 | 0.32 ± 0.01 | 0.32 ± 0.00 |
| Valeric acid | 0.49 ± 0.00 | 0.49 ± 0.06 | 0.48 ± 0.00 | 0.48 ± 0.01 | 0.49 ± 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, L.; Xie, X.; Li, Y.; Wu, L.; Fan, C.; Liang, T.; Xi, Y.; Yang, S.; Li, H.; Zhang, J.; et al. Evaluation of the Cholesterol-Lowering Mechanism of Enterococcus faecium Strain 132 and Lactobacillus paracasei Strain 201 in Hypercholesterolemia Rats. Nutrients 2021, 13, 1982. https://doi.org/10.3390/nu13061982
Yang L, Xie X, Li Y, Wu L, Fan C, Liang T, Xi Y, Yang S, Li H, Zhang J, et al. Evaluation of the Cholesterol-Lowering Mechanism of Enterococcus faecium Strain 132 and Lactobacillus paracasei Strain 201 in Hypercholesterolemia Rats. Nutrients. 2021; 13(6):1982. https://doi.org/10.3390/nu13061982
Chicago/Turabian StyleYang, Lingshuang, Xinqiang Xie, Ying Li, Lei Wu, Congcong Fan, Tingting Liang, Yu Xi, Shuanghong Yang, Haixin Li, Jumei Zhang, and et al. 2021. "Evaluation of the Cholesterol-Lowering Mechanism of Enterococcus faecium Strain 132 and Lactobacillus paracasei Strain 201 in Hypercholesterolemia Rats" Nutrients 13, no. 6: 1982. https://doi.org/10.3390/nu13061982