
Anti-Inflammatory and Immune Modulatory Effects of Synbio-Glucan in an Atopic Dermatitis Mouse Model

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Reagents
2.3. Induction of AD
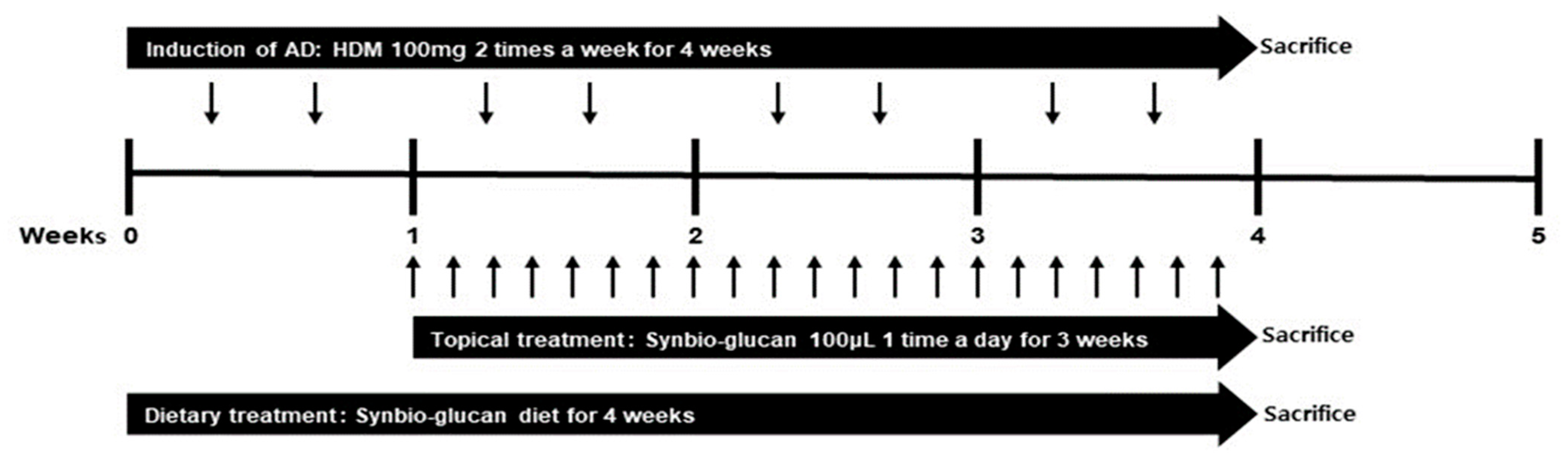
2.4. Treatment of Synbio-Glucan in the NC/Nga Mice
2.5. Serum IgE Concentration Assay
2.6. Serum Cytokine Antibody Assay
2.7. Scoring of Skin Lesions
2.8. Histological Analysis
2.9. Statistical Anlysis
3. Results
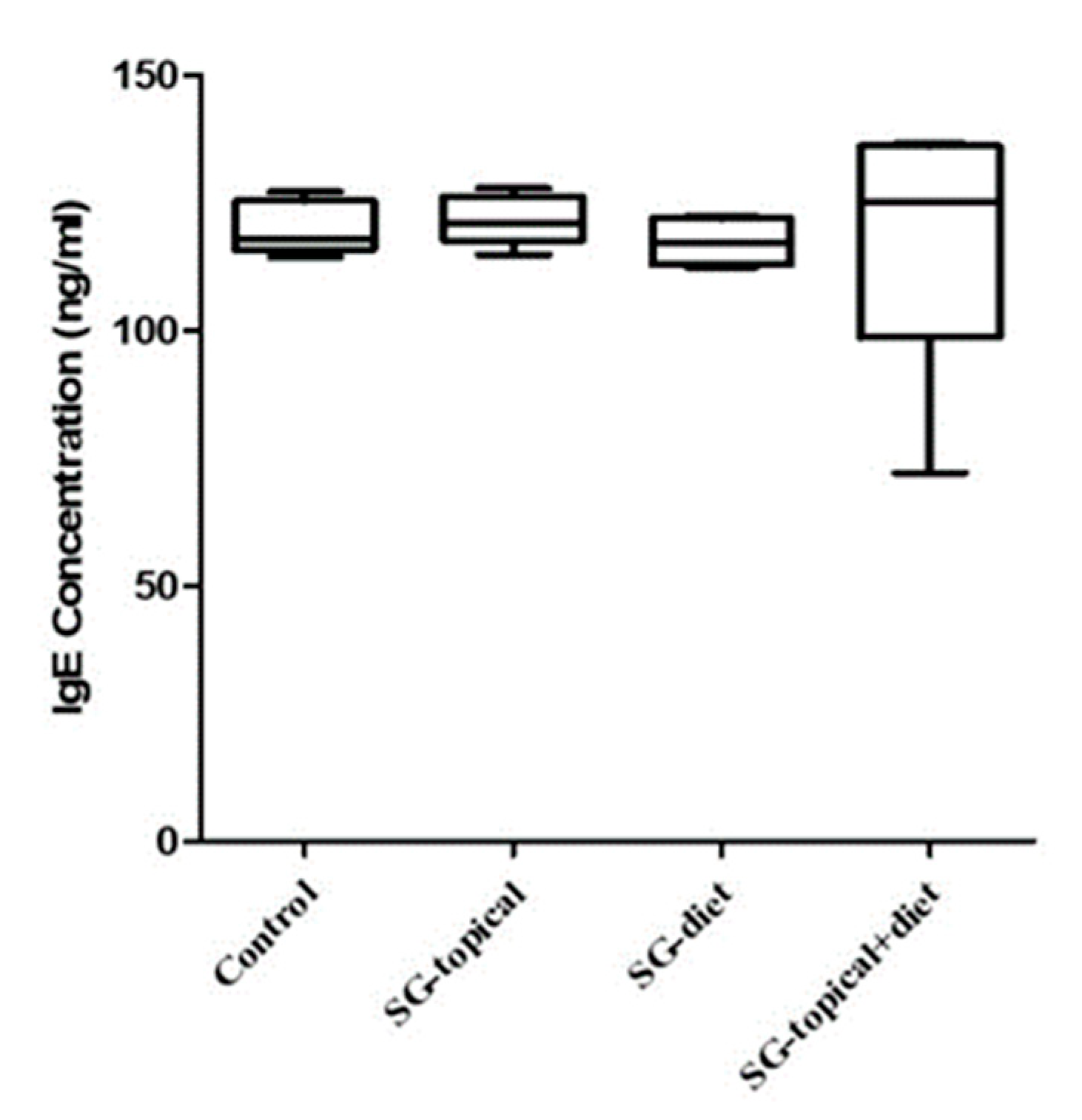
3.1. Comparison of the Serum IgE Concentration between Groups
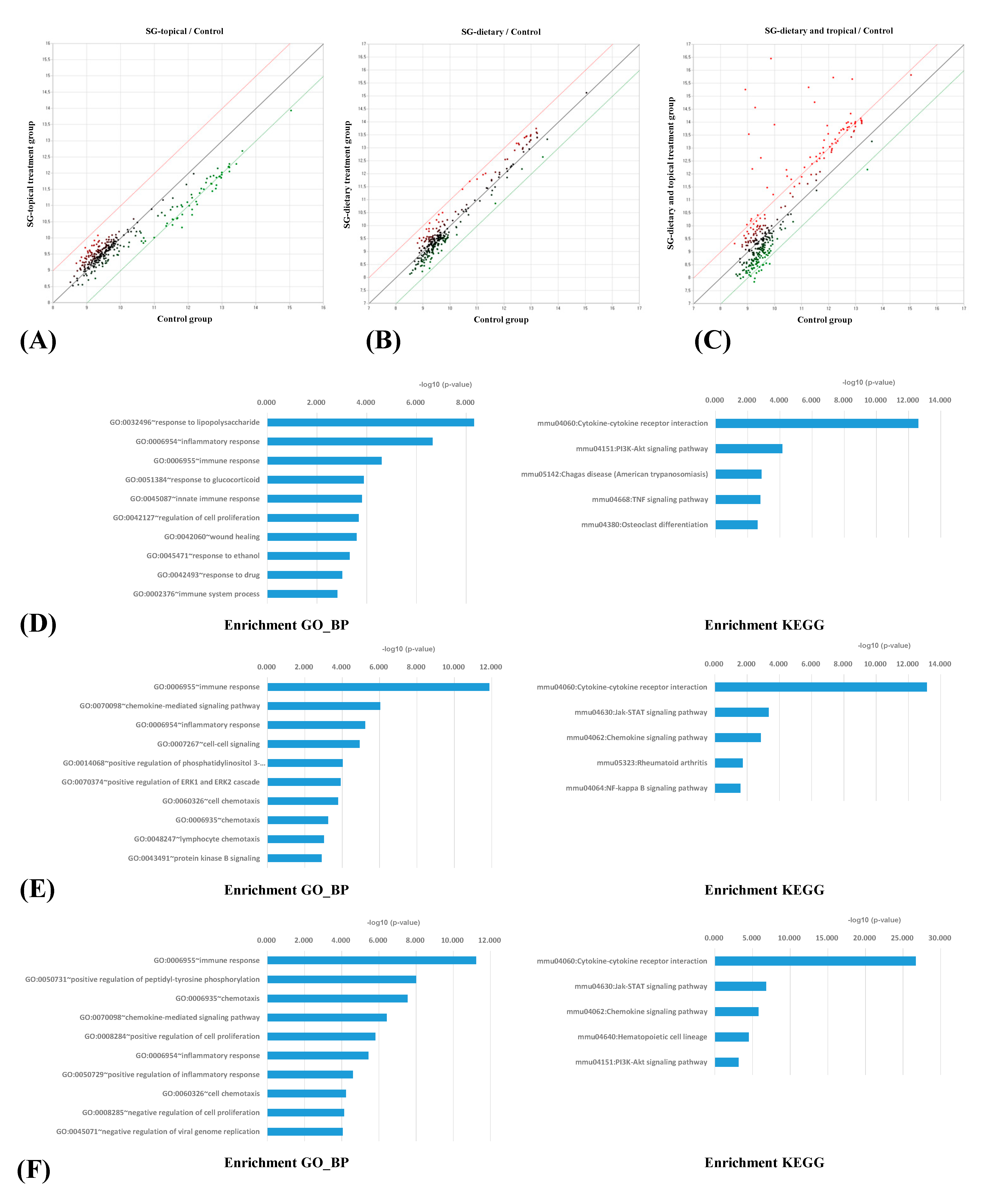
3.2. Comparison of the Serum Cytokine Antibody Arrays between Groups
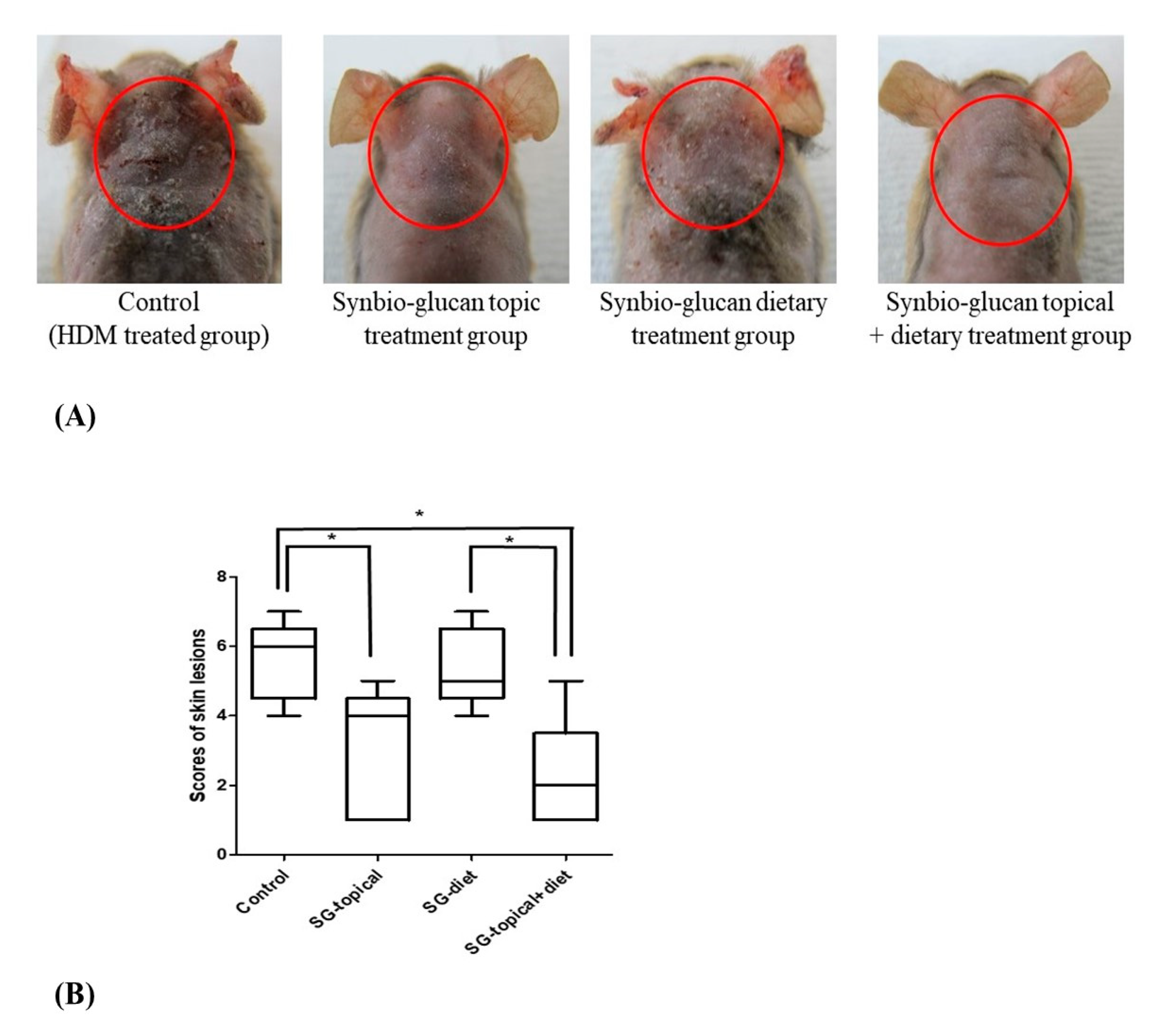
3.3. Comparison of Skin Lesion Scores between Groups
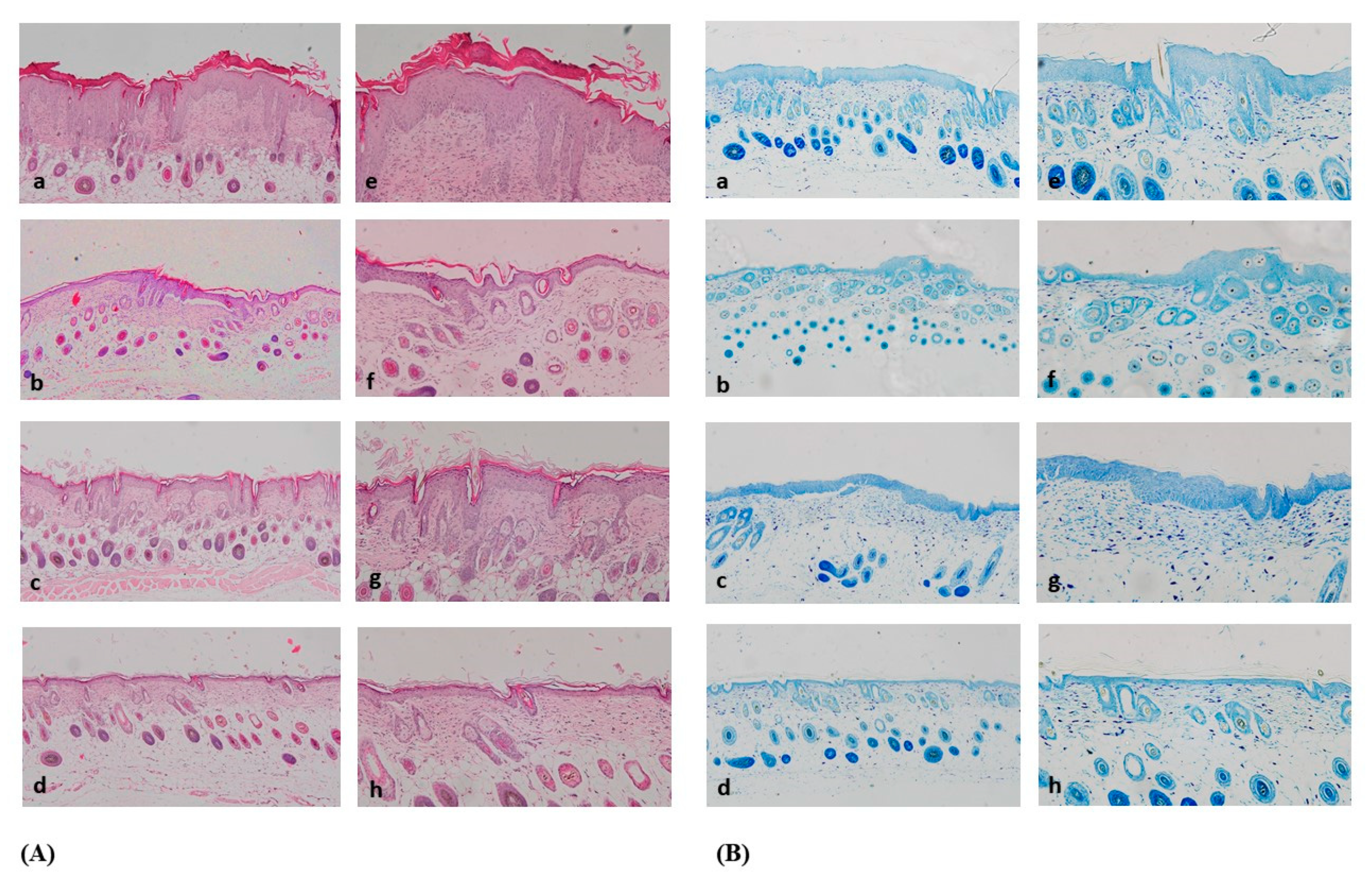
3.4. Comparison of Histological Results between the Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, C.; Park, K.Y.; Ahn, S.; Kim, D.H.; Li, K.; Kim, D.W.; Kim, M.B.; Jo, S.J.; Yim, H.W.; Seo, S.J. Economic Impact of Atopic Dermatitis in Korean Patients. Ann. Dermatol. 2015, 27, 298–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gittler, J.K.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis results in intrinsic barrier and immune abnormalities: Implications for contact dermatitis. J. Allergy Clin. Immunol. 2013, 131, 300–313. [Google Scholar] [CrossRef] [Green Version]
- Nettis, E.; Ortoncelli, M.; Pellacani, G.; Foti, C.; Di Leo, E.; Patruno, C.; Rongioletti, F.; Argenziano, G.; Ferrucci, S.M.; Macchia, L.; et al. A Multicenter Study on the Prevalence of Clinical Patterns and Clinical Phenotypes in Adult Atopic Dermatitis. J. Investig. Allergol. Clin. Immunol. 2020, 30, 448–450. [Google Scholar] [CrossRef] [PubMed]
- Wollenberg, A.; Barbarot, S.; Bieber, T.; Christen-Zaech, S.; Deleuran, M.; Fink-Wagner, A.; Gieler, U.; Girolomoni, G.; Lau, S.; Muraro, A.; et al. Consensus-based European guidelines for treatment of atopic eczema (atopic dermatitis) in adults and children: Part I. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 657–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dattola, A.; Bennardo, L.; Silvestri, M.; Nistico, S.P. What’s new in the treatment of atopic dermatitis? Dermatol. Ther. 2019, 32, e12787. [Google Scholar] [CrossRef]
- Eichenfield, L.F.; McCollum, A.; Msika, P. The benefits of sunflower oleodistillate (SOD) in pediatric dermatology. Pediatr. Dermatol. 2009, 26, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Danby, S.G.; AlEnezi, T.; Sultan, A.; Lavender, T.; Chittock, J.; Brown, K.; Cork, M.J. Effect of olive and sunflower seed oil on the adult skin barrier: Implications for neonatal skin care. Pediatr. Dermatol. 2013, 30, 42–50. [Google Scholar] [CrossRef]
- Sato, M.; Yamamoto-Hanada, K.; Yang, L.; Irahara, M.; Ishikawa, F.; Iwama-Mitsui, M.; Saito-Abe, M.; Miyaji, Y.; Inagaki, S.; Fukuie, T.; et al. Complementary and alternative medicine and atopic dermatitis in children. J. Dermatol. Sci. 2020, 97, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Delaney, B.; Nicolosi, R.J.; Wilson, T.A.; Carlson, T.; Frazer, S.; Zheng, G.H.; Hess, R.; Ostergren, K.; Haworth, J.; Knutson, N. Beta-glucan fractions from barley and oats are similarly antiatherogenic in hypercholesterolemic Syrian golden hamsters. J. Nutr. 2003, 133, 468–475. [Google Scholar] [CrossRef]
- Chan, G.C.; Chan, W.K.; Sze, D.M. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baldassano, S.; Accardi, G.; Vasto, S. Beta-glucans and cancer: The influence of inflammation and gut peptide. Eur. J. Med. Chem. 2017, 142, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Ho, H.V.; Sievenpiper, J.L.; Zurbau, A.; Blanco Mejia, S.; Jovanovski, E.; Au-Yeung, F.; Jenkins, A.L.; Vuksan, V. The effect of oat beta-glucan on LDL-cholesterol, non-HDL-cholesterol and apoB for CVD risk reduction: A systematic review and meta-analysis of randomised-controlled trials. Br. J. Nutr. 2016, 116, 1369–1382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jesenak, M.; Banovcin, P.; Rennerova, Z.; Majtan, J. beta-Glucans in the treatment and prevention of allergic diseases. Allergol. Immunopathol. 2014, 42, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.S.; Lee, S.H.; Kim, J.A.; Yu, D.Y.; Hong, Y.H.; Kim, J.Y.; Lim, J.M.; Lee, S.S.; Yun, C.H.; Choi, I.S.; et al. Effect of oral administration of beta-glucans derived from Aureobasidium pullulans SM-2001 in model mice and rat with atopic dermatitis-like phenotypes. Food Sci. Biotechnol. 2018, 27, 1185–1192. [Google Scholar] [CrossRef] [PubMed]
- Jesenak, M.; Urbancek, S.; Majtan, J.; Banovcin, P.; Hercogova, J. Beta-Glucan-based cream (containing pleuran isolated from pleurotus ostreatus) in supportive treatment of mild-to-moderate atopic dermatitis. J. Dermatol. Treat. 2016, 27, 351–354. [Google Scholar] [CrossRef]
- Krzych-Falta, E.; Furmanczyk, K.; Tomaszewska, A.; Olejniczak, D.; Samolinski, B.; Samolinska-Zawisza, U. Probiotics: Myths or facts about their role in allergy prevention. Adv. Clin. Exp. Med. 2018, 27, 119–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.Q.; Hu, H.J.; Liu, C.Y.; Zhang, Q.; Shakya, S.; Li, Z.Y. Probiotics for Prevention of Atopy and Food Hypersensitivity in Early Childhood: A PRISMA-Compliant Systematic Review and Meta-Analysis of Randomized Controlled Trials. Medicine 2016, 95, e2562. [Google Scholar] [CrossRef]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ogawa, H.; Ra, C. NC/Nga mice: A mouse model for atopic dermatitis. Int. Arch. Allergy Immunol. 1999, 120 (Suppl. 1), 70–75. [Google Scholar] [CrossRef]
- Takakura, M.; Takeshita, F.; Aihara, M.; Xin, K.Q.; Ichino, M.; Okuda, K.; Ikezawa, Z. Hyperproduction of IFN-gamma by CpG oligodeoxynucleotide-induced exacerbation of atopic dermatitis-like skin lesion in some NC/Nga mice. J. Investig. Dermatol. 2005, 125, 1156–1162. [Google Scholar] [CrossRef]
- Volman, J.J.; Ramakers, J.D.; Plat, J. Dietary modulation of immune function by beta-glucans. Physiol. Behav. 2008, 94, 276–284. [Google Scholar] [CrossRef]
- Brennan, C.S.; Cleary, L.J. The potential use of cereal (1 -> 3,1 -> 4)-beta-d-glucans as functional food ingredients. J. Cereal Sci. 2005, 42, 1–13. [Google Scholar] [CrossRef]
- Biorklund, M.; van Rees, A.; Mensink, R.P.; Onning, G. Changes in serum lipids and postprandial glucose and insulin concentrations after consumption of beverages with beta-glucans from oats or barley: A randomised dose-controlled trial. Eur. J. Clin. Nutr. 2005, 59, 1272–1281. [Google Scholar] [CrossRef] [Green Version]
- Bae, I.Y.; Lee, S.; Kim, S.M.; Lee, H.G. Effect of partially hydrolyzed oat beta-glucan on the weight gain and lipid profile of mice. Food Hydrocoll. 2009, 23, 2016–2021. [Google Scholar] [CrossRef]
- Sugiyama, A.; Hata, S.; Suzuki, K.; Yoshida, E.; Nakano, R.; Mitra, S.; Arashida, R.; Asayama, Y.; Yabuta, Y.; Takeuchi, T. Oral administration of paramylon, a beta-1,3-d;-glucan isolated from Euglena gracilis Z inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Vet. Med. Sci. 2010, 72, 755–763. [Google Scholar] [CrossRef] [Green Version]
- Meydani, S.N.; Ha, W.K. Immunologic effects of yogurt. Am. J. Clin. Nutr. 2000, 71, 861–872. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.; Kim, H.R.; Kim, N.R.; Jeong, B.J.; Lee, J.S.; Jang, S.; Chung, D.K. Oral administration of Lactobacillus plantarum lysates attenuates the development of atopic dermatitis lesions in mouse models. J. Microbiol. 2015, 53, 47–52. [Google Scholar] [CrossRef]
- Kim, I.S.; Lee, S.H.; Kwon, Y.M.; Adhikari, B.; Kim, J.A.; Yu, D.Y.; Kim, G.I.; Lim, J.M.; Kim, S.H.; Lee, S.S.; et al. Oral Administration of beta-Glucan and Lactobacillus plantarum Alleviates Atopic Dermatitis-Like Symptoms. J. Microbiol. Biotechnol. 2019, 29, 1693–1706. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, H.; Jiang, L.; Guo, H.; Luo, X.; Ren, F. Bifidobacterium longum BBMN68-specific modulated dendritic cells alleviate allergic responses to bovine beta-lactoglobulin in mice. J. Appl. Microbiol. 2015, 119, 1127–1137. [Google Scholar] [CrossRef]
- Yoshida, Y.; Seki, T.; Matsunaka, H.; Watanabe, T.; Shindo, M.; Yamada, N.; Yamamoto, O. Clinical Effects of Probiotic Bifidobacterium breve Supplementation in Adult Patients with Atopic Dermatitis. Yonago Acta Med. 2010, 53, 37–45. [Google Scholar]
- Duchaine, C.; Israel-Assayag, E.; Fournier, M.; Cormier, Y. Proinflammatory effect of Pediococcus pentosaceus, a bacterium used as hay preservative. Eur. Respir. J. 1996, 9, 2508–2512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.S.; Eom, J.E.; Shin, D.U.; Yeon, S.H.; Lim, S.I.; Lee, S.Y. Preventive Effects of a Probiotic Mixture in an Ovalbumin-Induced Food Allergy Model. J. Microbiol. Biotechnol. 2018, 28, 65–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vestergaard, C.; Yoneyama, H.; Murai, M.; Nakamura, K.; Tamaki, K.; Terashima, Y.; Imai, T.; Yoshie, O.; Irimura, T.; Mizutani, H.; et al. Overproduction of Th2-specific chemokines in NC/Nga mice exhibiting atopic dermatitis-like lesions. J. Clin. Investig. 1999, 104, 1097–1105. [Google Scholar] [CrossRef]
- Spergel, J.M.; Mizoguchi, E.; Oettgen, H.; Bhan, A.K.; Geha, R.S. Roles of TH1 and TH2 cytokines in a murine model of allergic dermatitis. J. Clin. Investig. 1999, 103, 1103–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollenberg, A.; Rawer, H.C.; Schauber, J. Innate immunity in atopic dermatitis. Clin. Rev. Allergy Immunol. 2011, 41, 272–281. [Google Scholar] [CrossRef]
- Ciprandi, G.; De Amici, M.; Giunta, V.; Marseglia, A.; Marseglia, G. Serum interleukin-9 levels are associated with clinical severity in children with atopic dermatitis. Pediatr. Dermatol. 2013, 30, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Xue, H.B.; Guan, X.H.; Shu, C.M.; Zhang, J.H.; Yu, J. Possible pathogenic role of T helper type 9 cells and interleukin (IL)-9 in atopic dermatitis. Clin. Exp. Immunol. 2014, 175, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Holck, P.; Sletmoen, M.; Stokke, B.T.; Permin, H.; Norn, S. Potentiation of histamine release by Microfungal (1-->3)- and (1-->6)-beta-d-glucans. Basic Clin. Pharmacol. Toxicol. 2007, 101, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium. The Gene Ontology (GO) project in 2006. Nucleic Acids Res. 2006, 34, D322–D326. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rank | Antibody Name | Fold-Change | Gene Symbol | Swiss-Prot Entry |
|---|---|---|---|---|
| Downregulated | ||||
| 1 | IFN-beta | 0.354 | Ifnb1 | P01575 |
| 2 | GDF-8 | 0.377 | Mstn | O08689 |
| 3 | Common gamma Chain/IL-2 R gamma | 0.408 | Il2rg | P34902 |
| 4 | Endostatin | 0.412 | Col18a1 | P39061 |
| 5 | IGFBP-3 | 0.423 | Igfbp3 | P47878 |
| 6 | SPARC | 0.424 | Sparc | P07214 |
| 7 | WISP-1/CCN4 | 0.432 | Wisp1 | O54775 |
| 8 | TLR2 | 0.440 | Tlr2 | Q9QUN7 |
| 9 | SLPI | 0.450 | Slpi | P97430 |
| 10 | MIP2 | 0.453 | Cxcl2 | P10889 |
| 11 | VEGF-B | 0.456 | Vegfb | P49766 |
| 12 | CCL28 | 0.458 | Ccl28 | Q9JIL2 |
| 13 | ICAM-1 | 0.460 | Icam1 | P13597 |
| 14 | Fas/TNFRSF6 | 0.461 | Fas | P25446 |
| 15 | CXCR6 | 0.465 | Cxcr6 | Q9EQ16 |
| 16 | IL-1 RI | 0.475 | Il1r1 | P13504 |
| 17 | IGFBP-1 | 0.475 | Igfbp1 | P47876 |
| 18 | b FGF | 0.482 | Fgf2 | P15655 |
| 19 | Prolactin | 0.483 | Prl | P06879 |
| 20 | M-CSF | 0.485 | Csf1 | P07141 |
| 21 | TGF-beta RII | 0.485 | Tgfbr2 | Q62312 |
| 22 | CRP | 0.488 | Crp | P14847 |
| 23 | Lymphotoxin beta R/TNFRSF3 | 0.488 | Ltbr | P50284 |
| 24 | Frizzled-6 | 0.496 | Fzd6 | Q61089 |
| 25 | IL-27 | 0.499 | Il27 | Q8K3I6 |
| 26 | IL-23 R | 0.499 | Il23r | Q8K4B4 |
| 27 | TCCR/WSX-1 | 0.499 | Il27ra | O70394 |
| Rank | Antibody Name | Fold-Change | Gene Symbol | Swiss-Prot Entry |
|---|---|---|---|---|
| Upregulated | ||||
| 1 | IL-9 | 83.568 | Il9 | P15247 |
| 2 | Dtk | 62.380 | Tyro3 | P55144 |
| 3 | FGF R3 | 24.767 | Fgfr3 | Q61851 |
| 4 | GFR alpha-4/GDNF R alpha-4 | 16.304 | Gfra4 | Q9JJT2 |
| 5 | Thymus Chemokine-1 | 14.052 | Ppbp, | Q9EQI5 |
| 6 | TRAIL/TNFSF10 | 9.431 | Tnfsf10 | P50592 |
| 7 | Follistatin-like 1 | 8.738 | Fstl1 | Q62356 |
| 8 | VE-Cadherin | 6.714 | Cdh5 | P55284 |
| 9 | BLC | 6.328 | Cxcl13 | O55038 |
| 10 | ICAM-2/CD102 | 5.753 | Icam2 | P35330 |
| 11 | IL-22 | 5.632 | Il22 | Q9JJY9 |
| 12 | IL-10 R alpha | 5.608 | Il10ra | Q61727 |
| 13 | WIF-1 | 4.277 | Wif1 | Q9WUA1 |
| 14 | MIP-3 beta | 3.753 | Ccl19 | O70460 |
| 15 | MIP-1alpha | 3.608 | Ccl3 | P10855 |
| 16 | LIF | 3.496 | Lif | P09056 |
| 17 | VEGF-D | 3.197 | Figf | P97946 |
| 18 | RANTES | 3.117 | Ccl5 | P30882 |
| 19 | Decorin | 2.988 | Dcn | P28654 |
| 20 | P-Selectin | 2.687 | Selp | Q01102 |
| 21 | IL-13 | 2.309 | Il13 | P20109 |
| 22 | IL-1 Ra | 2.288 | Il1r1 | P13504 |
| 23 | IL-1 R4/ST2 | 2.245 | Il1rl1 | P14719 |
| 24 | PDGF-C | 2.180 | Pdgfc | Q8CI19 |
| 25 | CD27 Ligand/TNFSF7 | 2.173 | Cd70 | O55237 |
| 26 | ICK | 2.153 | Ick | Q9JKV2 |
| 27 | SDF-1 | 2.074 | Cxcl12 | P40224 |
| Rank | Antibody Name | Fold-Change | Gene Symbol | Swiss-Prot Entry |
|---|---|---|---|---|
| Upregulated | ||||
| 1 | Dtk | 95.560 | Tyro3 | P55144 |
| 2 | IL-9 | 80.615 | Il9 | P15247 |
| 3 | GFR alpha-4/GDNF R alpha-4 | 38.326 | Gfra4 | Q9JJT2 |
| 4 | FGF R3 | 22.346 | Fgfr3 | Q61851 |
| 5 | Follistatin-like 1 | 16.970 | Fstl1 | Q62356 |
| 6 | Thymus Chemokine-1 | 14.916 | Ppbp, | Q9EQI5 |
| 7 | ICAM-2/CD102 | 11.732 | Icam2 | P35330 |
| 8 | VE-Cadherin | 9.709 | Cdh5 | P55284 |
| 9 | TRAIL/TNFSF10 | 8.673 | Tnfsf10 | P50592 |
| 10 | Decorin | 8.051 | Dcn | P28654 |
| 11 | IL-1 Ra | 6.894 | Il1r1 | P13504 |
| 12 | ICK | 3.760 | Ick | Q9JKV2 |
| 13 | Frizzled-7 | 3.344 | Fzd7 | Q61090 |
| 14 | GDF-5 | 3.263 | Gdf5 | P43027 |
| 15 | IL-1 R4/ST2 | 2.961 | Il1rl1 | P14719 |
| 16 | Common gamma Chain/IL-2 R gamma | 2.836 | Il2rg | P34902 |
| 17 | CXCR6 | 2.769 | Cxcr6 | Q9EQ16 |
| 18 | Lungkine | 2.718 | Cxcl15 | Q9WVL7 |
| 19 | VEGFC | 2.509 | Vegfc | P97953 |
| 20 | Glut2 | 2.441 | Slc2a2 | P14246 |
| 21 | Endostatin | 2.422 | Col18a1 | P39061 |
| 22 | RANTES | 2.364 | Ccl5 | P30882 |
| 23 | CTACK | 2.329 | Ccl27 | Q9Z1X0 |
| 24 | LIF | 2.310 | Lif | P09056 |
| 25 | IL-28/IFN-lambda | 2.298 | Il28b | Q8CGK6 |
| 26 | TCA-3 | 2.284 | Ccl1 | P10146 |
| 27 | IGFBP-2 | 2.246 | Igfbp2 | P47877 |
| 28 | IL-17 R | 2.232 | Il17ra | Q60943 |
| 29 | Eotaxin-2 | 2.216 | Ccl24 | Q9JKC0 |
| 30 | IL-31 | 2.192 | Il31 | Q6EAL8 |
| 31 | BLC | 2.174 | Cxcl13 | O55038 |
| 32 | IL-11 | 2.150 | Il11 | P47873 |
| 33 | HVEM/TNFRSF14 | 2.145 | Tnfrsf14 | NP_849262 |
| 34 | CCL28 | 2.135 | Ccl28 | Q9JIL2 |
| 35 | CRP | 2.127 | Crp | P14847 |
| 36 | FLRG(Follistatin) | 2.105 | Fstl3 | Q9EQC7 |
| 37 | beta-Catenin | 2.103 | Ctnnb1 | Q02248 |
| 38 | Soggy-1 | 2.091 | Dkkl1 | Q9QZL9 |
| 39 | GDF-8 | 2.064 | Mstn | O08689 |
| 40 | IGFBP-5 | 2.040 | Igfbp5 | Q07079 |
| 41 | LIX | 2.017 | Cxcl5 | P50228 |
| 42 | Frizzled-6 | 2.015 | Fzd6 | Q61089 |
| 43 | b FGF | 2.011 | Fgf2 | P15655 |
| 44 | IFN-beta | 2.007 | Ifnb1 | P01575 |
| 45 | CCL1/I-309/TCA-3 | 2.001 | Ccl1 | P10146 |
| Downregulated | ||||
| 1 | Activin A | 0.373 | Inhba | Q04998 |
| 2 | Gremlin | 0.403 | Grem1 | O70326 |
| 3 | IL-4 | 0.408 | Il4 | P07750 |
| 4 | SLPI | 0.416 | Slpi | P97430 |
| 5 | Angiopoietin-like 2 | 0.425 | Angptl2 | Q9R045 |
| 6 | Frizzled-1 | 0.446 | Fzd1 | O70421 |
| 7 | Growth Hormone R | 0.447 | Ghr | P16882 |
| 8 | IL-6 R | 0.456 | Il6ra | P22272 |
| 9 | ICAM-5 | 0.469 | Icam5 | Q60625 |
| 10 | IL-1 Rb | 0.473 | Il1r2 | P27931 |
| 11 | Axl | 0.475 | Axl | Q00993 |
| 12 | Flt-3 Ligand | 0.487 | Flt3l | P49772 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.-H.; Kang, M.S.; Kim, T.H.; Jeong, Y.; Ahn, J.-O.; Choi, J.H.; Chung, J.-Y. Anti-Inflammatory and Immune Modulatory Effects of Synbio-Glucan in an Atopic Dermatitis Mouse Model. Nutrients 2021, 13, 1090. https://doi.org/10.3390/nu13041090
Kim Y-H, Kang MS, Kim TH, Jeong Y, Ahn J-O, Choi JH, Chung J-Y. Anti-Inflammatory and Immune Modulatory Effects of Synbio-Glucan in an Atopic Dermatitis Mouse Model. Nutrients. 2021; 13(4):1090. https://doi.org/10.3390/nu13041090
Chicago/Turabian StyleKim, Yoon-Hwan, Min Soo Kang, Tae Hyeong Kim, Yunho Jeong, Jin-Ok Ahn, Jung Hoon Choi, and Jin-Young Chung. 2021. "Anti-Inflammatory and Immune Modulatory Effects of Synbio-Glucan in an Atopic Dermatitis Mouse Model" Nutrients 13, no. 4: 1090. https://doi.org/10.3390/nu13041090