In Vivo Anti-Inflammation Potential of Aster koraiensis Extract for Dry Eye Syndrome by the Protection of Ocular Surface



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation of A. koraiensis Extracts
2.2. Antioxidative Activities of AKE
2.3. Total Polyphenol and Flavonoid Contents of AKE
2.4. Animal Experiment and Induction of DE Model
2.5. Histology
2.6. Western Blot Analysis
2.7. Real-Time PCR (RT-qPCR)
2.8. ARPE-19 Cell Culture
2.9. Intracellular Calcium Release
2.10. VEGF-α Secretion
2.11. Statistical Analysis
3. Results
3.1. Antioxidative Effects and Polyphenol and Flavonoid Contents of AKE
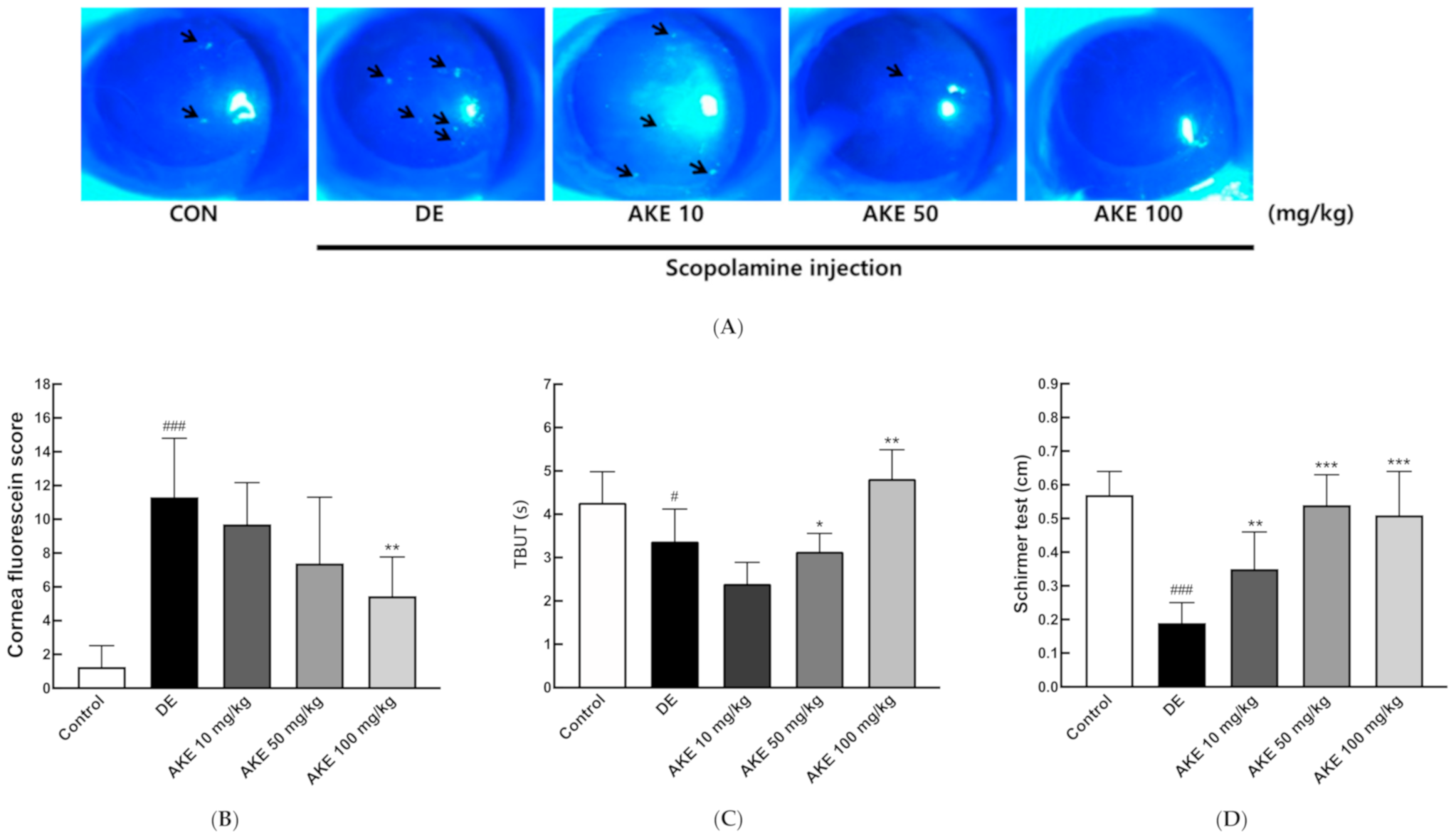
3.2. Effects of Aster koraiensis Ethanol Extracts on Eye Damage and Tear Production
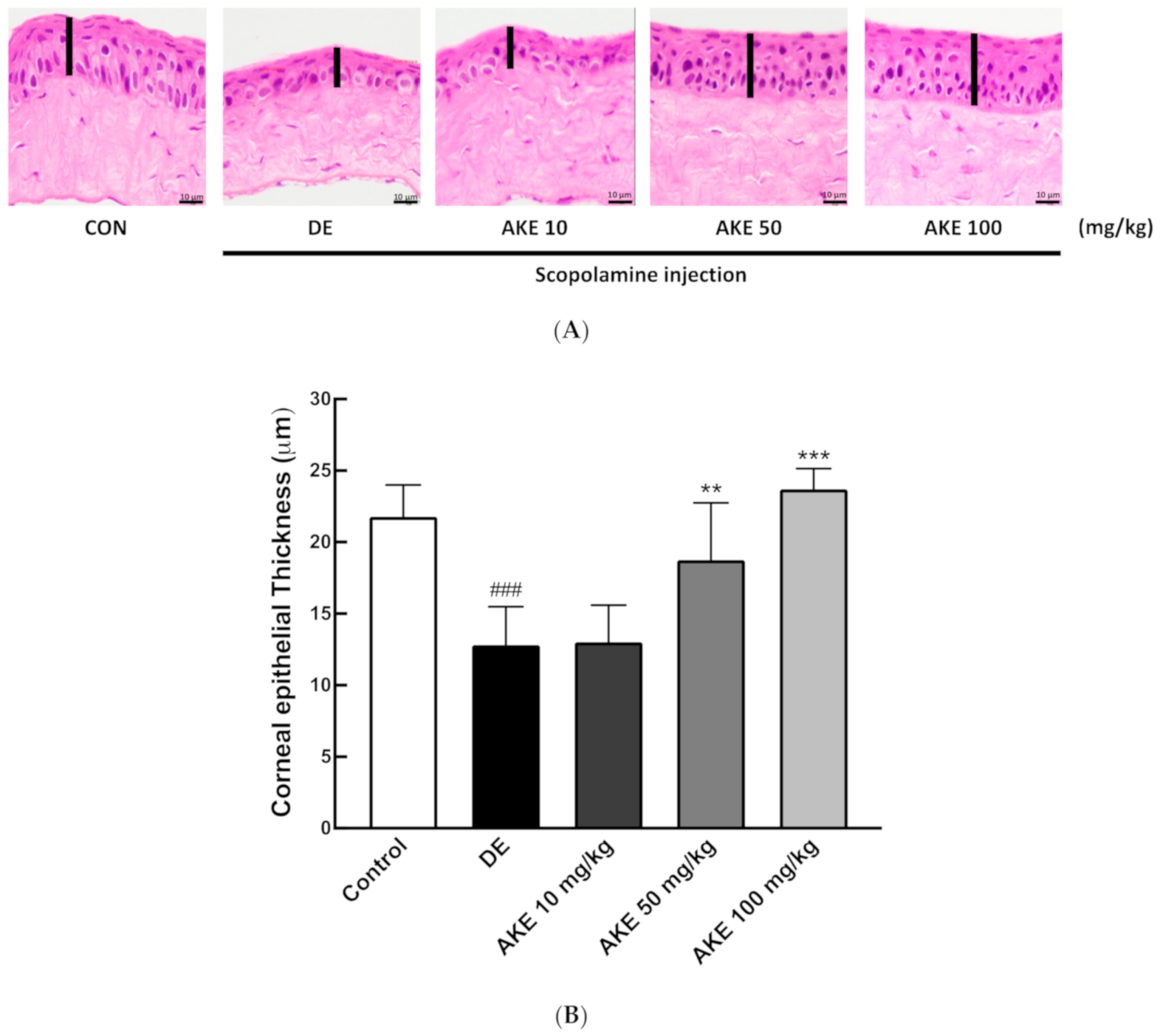
3.3. Histological Alterations of the Corneal Epithelial and Lacrimal Glands Following AKE Treatment in the DE Mouse Model
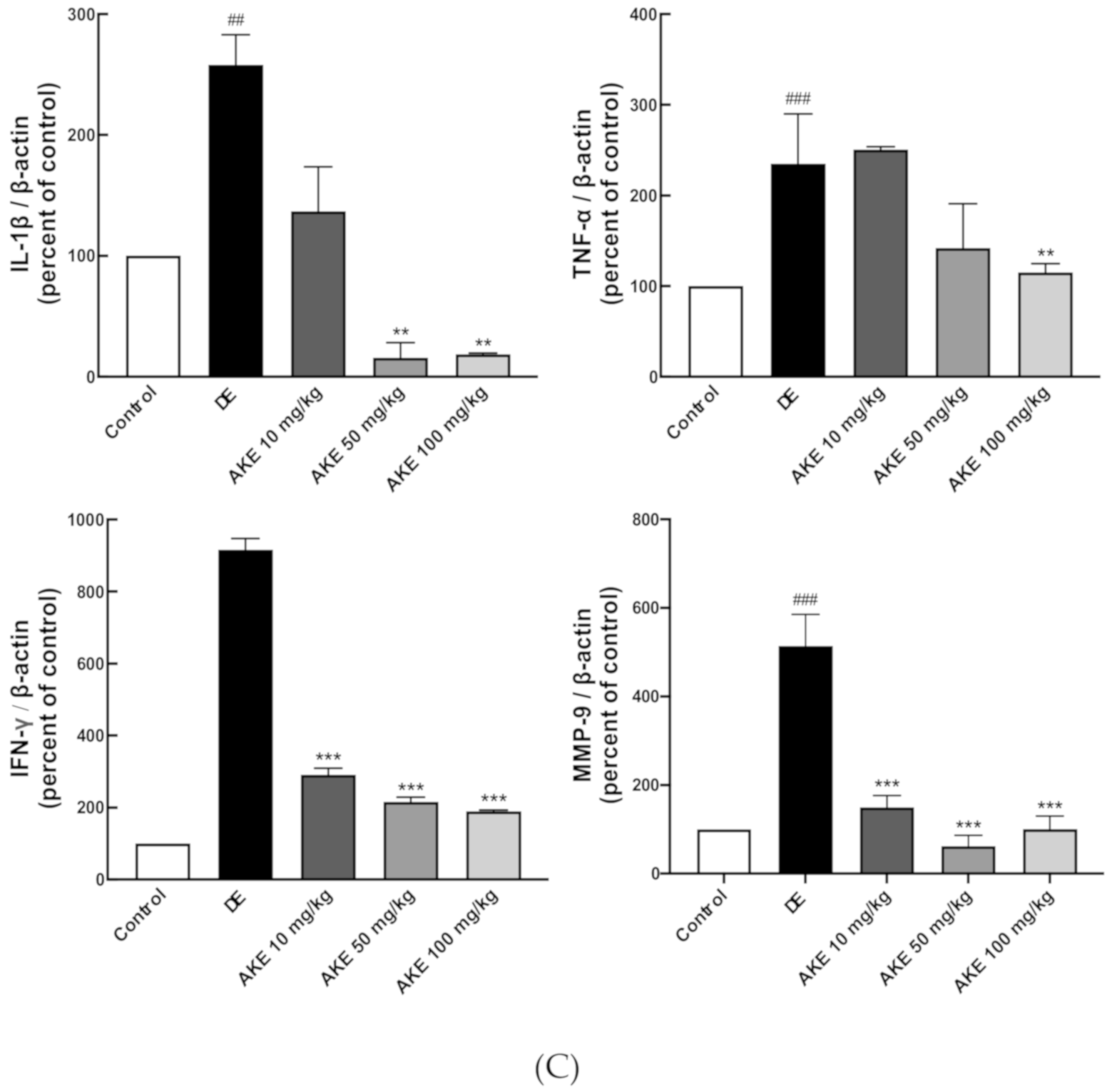
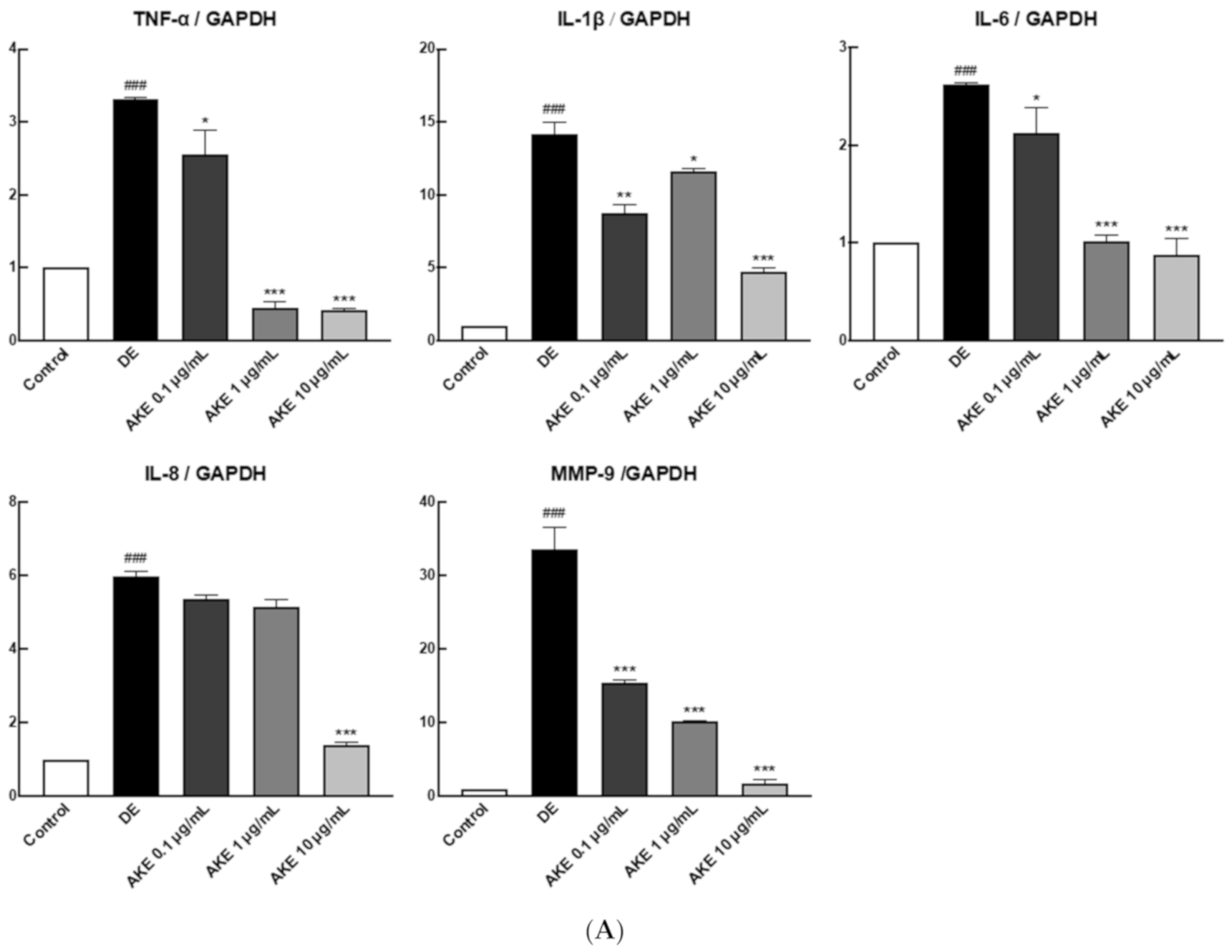
3.4. AKE Suppressed Immune Responses in the Corneal Epithelium and Lacrimal Glands of DE Model Mice
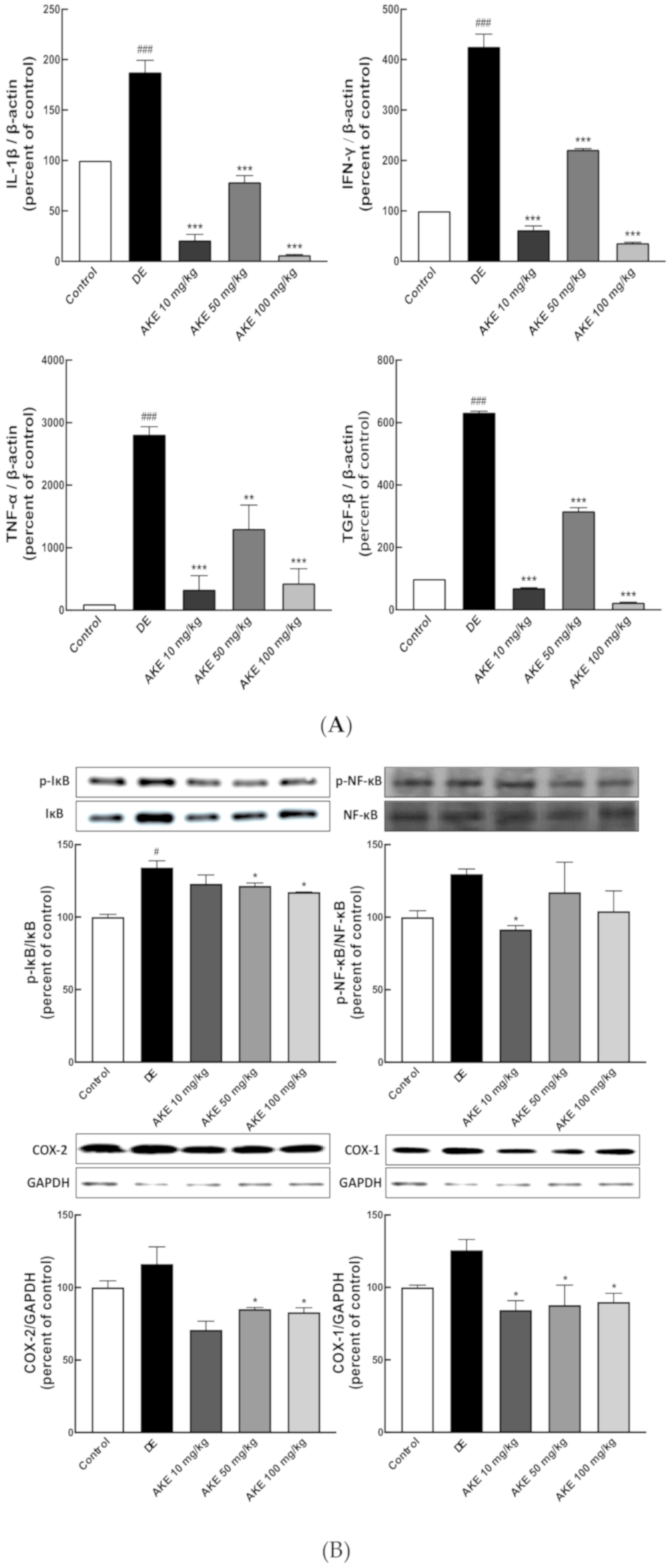
3.5. AKE Inhibited Inflammatory Response in Lacrimal Gland
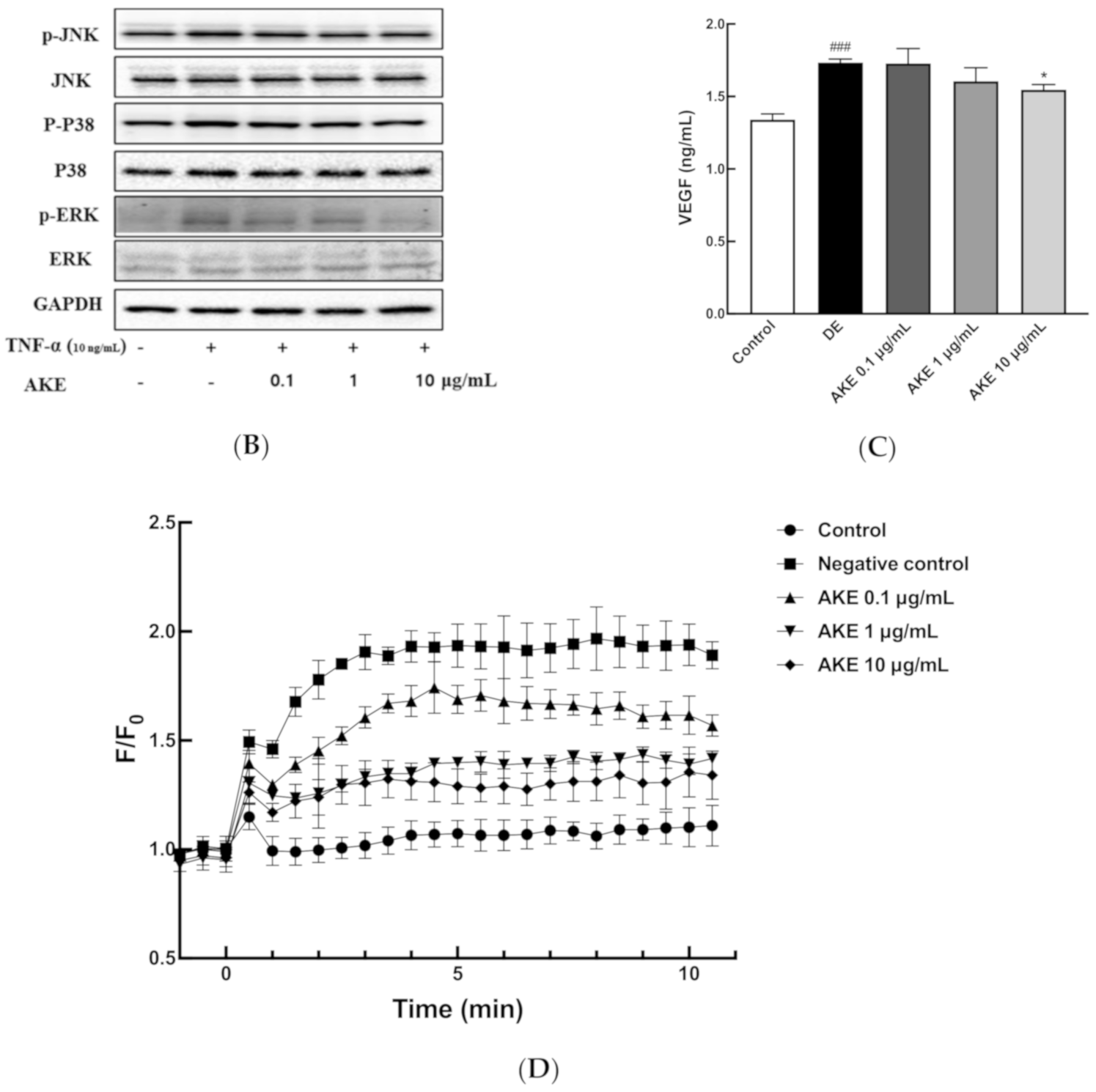
3.6. AKE Inhibited Inflammatory Reaction and ER Stress on ARPE-19 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Phadatare, S.P.; Momin, M.; Nighojkar, P.; Askarkar, S.; Singh, K.K. A comprehensive review on dry eye disease: Diagnosis, medical management, recent developments, and future challenges. Adv. Pharm. 2015, 2015, 704946. [Google Scholar] [CrossRef]
- Paulsen, A.J.; Cruickshanks, K.J.; Fischer, M.E.; Huang, G.-H.; Klein, B.E.; Klein, R.; Dalton, D.S. Dry eye in the beaver dam offspring study: Prevalence, risk factors, and health-related quality of life. Am. J. Ophthalmol. 2014, 157, 799–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, U.; Willcox, M.; Stapleton, F. Osmolality and tear film dynamics. Clin. Exp. Optom. 2012, 95, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qu, Y.; He, X.; Ou, S.; Bu, J.; Jia, C.; Wang, J.; Wu, H.; Liu, Z.; Li, W. Dry eye management: Targeting the ocular surface microenvironment. Int. J. Mol. Sci. 2017, 18, 1398. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.; Garrett, Q.; Flanagan, J.; Chakrabarti, S.; Papas, E. Genetic factors and molecular mechanisms in dry eye disease. Ocul. Surf. 2018, 16, 206–217. [Google Scholar] [CrossRef]
- Liu, K.C.; Huynh, K.; Grubbs, J.; Davis, R.M. Autoimmunity in the pathogenesis and treatment of keratoconjunctivitis sicca. Curr. Allergy Asthma Rep. 2014, 14, 403. [Google Scholar] [CrossRef]
- Baudouin, C.; Irkeç, M.; Messmer, E.M.; Benítez-del-Castillo, J.M.; Bonini, S.; Figueiredo, F.C.; Geerling, G.; Labetoulle, M.; Lemp, M.; Rolando, M. Clinical impact of inflammation in dry eye disease: Proceedings of the ODISSEY group meeting. Acta Ophthalmol. 2018, 96, 111–119. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, X.; Yang, L.; Li, M.; Li, B.; Wang, W.; Sheng, M. Decreased PPAR-γ expression in the conjunctiva and increased expression of TNF-α and IL-1β in the conjunctiva and tear fluid of dry eye mice. Mol. Med. Rep. 2014, 9, 2015–2023. [Google Scholar] [CrossRef]
- VanDerMeid, K.R.; Su, S.P.; Ward, K.W.; Zhang, J.-Z. Correlation of tear inflammatory cytokines and matrix metalloproteinases with four dry eye diagnostic tests. Investig. Ophthalmol. Vis. Sci. 2012, 53, 1512–1518. [Google Scholar] [CrossRef]
- Kunert, K.S.; Tisdale, A.S.; Stern, M.E.; Smith, J.; Gipson, I.K. Analysis of topical cyclosporine treatment of patients with dry eye syndrome: Effect on conjunctival lymphocytes. Arch. Ophthalmol. 2000, 118, 1489–1496. [Google Scholar] [CrossRef]
- Rolando, M.; Stern, M.E.; Calonge, M. Modern perspectives on dry eye disease. Eur. Ophthalmic Rev. 2017, 11, 2–6. [Google Scholar]
- Stern, M.E.; Schaumburg, C.S.; Pflugfelder, S.C. Dry eye as a mucosal autoimmune disease. Int. Rev. Immunol. 2013, 32, 19–41. [Google Scholar] [CrossRef]
- Wei, Y.; Asbell, P.A. The core mechanism of dry eye disease (DED) is inflammation. Eye Contact Lens 2014, 40, 248. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Bailly-Maitre, B.; Reed, J.C. Endoplasmic reticulum stress: Cell life and death decisions. J. Clin. Investig. 2005, 115, 2656–2664. [Google Scholar] [CrossRef] [Green Version]
- Bahar, E.; Kim, H.; Yoon, H. ER stress-mediated signaling: Action potential and Ca2+ as key players. Int. J. Mol. Sci. 2016, 17, 1558. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Endoplasmic reticulum stress and the inflammatory basis of metabolic disease. Cell 2010, 140, 900–917. [Google Scholar] [CrossRef] [Green Version]
- Seo, Y.; Ji, Y.; Lee, S.; Shim, J.; Noh, H.; Yeo, A.; Park, C.; Park, M.; Chang, E.; Lee, H. Activation of HIF-1 α (hypoxia inducible factor-1 α) prevents dry eye-induced acinar cell death in the lacrimal gland. Cell Death Dis. 2014, 5, e1309. [Google Scholar] [CrossRef] [Green Version]
- Barabino, S.; Chen, Y.; Chauhan, S.; Dana, R. Ocular surface immunity: Homeostatic mechanisms and their disruption in dry eye disease. Prog. Retin. Eye Res. 2012, 31, 271–285. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Yiu, S.C. Dry eye disease: A review of diagnostic approaches and treatments. Saudi J. Ophthalmol. 2014, 28, 173–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Messmer, E.M. The pathophysiology, diagnosis, and treatment of dry eye disease. Dtsch. Ärztebl. Int. 2015, 112, 71. [Google Scholar] [CrossRef] [Green Version]
- Downie, L.E.; Keller, P.R. A pragmatic approach to the management of dry eye disease: Evidence into practice. Optom. Vis. Sci. 2015, 92, 957–966. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.-N.; Wu, T.-Y.; Chau, C.-F. Natural dietary and herbal products in anti-obesity treatment. Molecules 2016, 21, 1351. [Google Scholar] [CrossRef]
- Granato, D.; Barba, F.J.; Bursać Kovačević, D.; Lorenzo, J.M.; Cruz, A.G.; Putnik, P. Functional foods: Product development, technological trends, efficacy testing, and safety. Annu. Rev. Food Sci. Technol. 2020, 11, 93–118. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Toalá, J.; Hernández-Mendoza, A.; González-Córdova, A.; Vallejo-Cordoba, B.; Liceaga, A. Potential role of natural bioactive peptides for development of cosmeceutical skin products. Peptides 2019, 122, 170170. [Google Scholar]
- Hooshmand, S.; Arjmandi, B.H. Dried plum, an emerging functional food that may effectively improve bone health. Ageing Res. Rev. 2009, 8, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-H.; Sim, Y.-B.; Kim, S.-M.; Kang, Y.-J.; Lee, J.-K.; Lim, S.-S.; Kim, J.-K.; Suh, H.-W. Antinociceptive profiles and mechanisms of orally administered Aster Koraiensis extract in the mouse. J. Med. Plants Res. 2011, 5, 6267–6272. [Google Scholar]
- Ahn, D. Illustrated Book of Korean Medicinal Herbs; Kyohaksa: Seoul, Korea, 1998; Volume 497, pp. 23–115. [Google Scholar]
- Ko, J.; Lee, K. Effect of plant growth regulators on growth and flowering of potted Lychnis cognata, Aster koraiensis and Campanula takesimana. RDA J. Agric. Sci. Korea Repub. 1996, 38, 627–632. [Google Scholar]
- Lee, J.; Lee, Y.M.; Lee, B.W.; Kim, J.-H.; Kim, J.S. Chemical constituents from the aerial parts of Aster koraiensis with protein glycation and aldose reductase inhibitory activities. J. Nat. Prod. 2012, 75, 267–270. [Google Scholar] [CrossRef]
- Sohn, E.; Kim, J.; Kim, C.-S.; Kim, Y.S.; Jang, D.S.; Kim, J.S. Extract of the aerial parts of Aster koraiensis reduced development of diabetic nephropathy via anti-apoptosis of podocytes in streptozotocin-induced diabetic rats. Biochem. Biophys. Res. Commun. 2010, 391, 733–738. [Google Scholar] [CrossRef]
- Hyun, S.-W.; Kim, J.; Jo, K.; Kim, J.S.; Kim, C.-S. Aster koraiensis extract improves impaired skin wound healing during hyperglycemia. Integr. Med. Res. 2018, 7, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Jo, K.; Lee, I.-S.; Kim, C.-S.; Kim, J.S. The extract of Aster Koraiensis prevents retinal Pericyte apoptosis in diabetic rats and its active compound, chlorogenic acid inhibits AGE formation and AGE/RAGE interaction. Nutrients 2016, 8, 585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Lee, Y.M.; Jung, W.; Park, S.-B.; Kim, C.-S.; Kim, J.S. Aster koraiensis extract and chlorogenic acid inhibit retinal angiogenesis in a mouse model of oxygen-induced retinopathy. Evid. Based Complementary Altern. Med. 2018, 2018, 4916497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serpen, A.; Gökmen, V.; Fogliano, V. Total antioxidant capacities of raw and cooked meats. Meat Sci. 2012, 90, 60–65. [Google Scholar] [CrossRef]
- Thaipong, K.; Boonprakob, U.; Crosby, K.; Cisneros-Zevallos, L.; Byrne, D.H. Comparison of ABTS, DPPH, FRAP, and ORAC assays for estimating antioxidant activity from guava fruit extracts. J. Food Compos. Anal. 2006, 19, 669–675. [Google Scholar] [CrossRef]
- Benzie, I.F.; Strain, J. [2] Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 15–27. [Google Scholar]
- Oki, T.; Nagai, S.; Yoshinaga, M.; Nishiba, Y.; Suda, I. Contribution of β-carotene to radical scavenging capacity varies among orange-fleshed sweet potato cultivars. Food Sci. Technol. Res. 2006, 12, 156–160. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Alves, R.C.; Costa, A.S.; Jerez, M.; Casal, S.; Sineiro, J.; Nunez, M.J.; Oliveira, B. Antiradical activity, phenolics profile, and hydroxymethylfurfural in espresso coffee: Influence of technological factors. J. Agric. Food Chem. 2010, 58, 12221–12229. [Google Scholar] [CrossRef] [PubMed]
- Pourmorad, F.; Hosseinimehr, S.; Shahabimajd, N. Antioxidant activity, phenol and flavonoid contents of some selected Iranian medicinal plants. Afr. J. Biotechnol. 2006, 5, 1142–1145. [Google Scholar]
- Marinova, D.; Ribarova, F.; Atanassova, M. Total phenolics and total flavonoids in Bulgarian fruits and vegetables. J. Univ. Chem. Technol. Metallurgy 2005, 40, 255–260. [Google Scholar]
- Kasote, D.M.; Katyare, S.S.; Hegde, M.V.; Bae, H. Significance of antioxidant potential of plants and its relevance to therapeutic applications. Int. J. Biol. Sci. 2015, 11, 982. [Google Scholar] [CrossRef] [Green Version]
- El Jemli, M.; Kamal, R.; Marmouzi, I.; Zerrouki, A.; Cherrah, Y.; Alaoui, K. Radical-scavenging activity and ferric reducing ability of Juniperus thurifera (L.), J. oxycedrus (L.), J. phoenicea (L.) and Tetraclinis articulata (L.). Adv. Pharmacol. Sci. 2016, 2016, 6392656. [Google Scholar]
- Javadi, M.-A.; Feizi, S. Dry eye syndrome. J. Ophthalmic Vis. Res. 2011, 6, 192. [Google Scholar]
- Dienes, L.; Kiss, H.J.; Perényi, K.; Nagy, Z.Z.; Acosta, M.C.; Gallar, J.; Kovács, I. Corneal sensitivity and dry eye symptoms in patients with keratoconus. PLoS ONE 2015, 10, e0141621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conrady, C.D.; Joos, Z.P.; Patel, B.C. The lacrimal gland and its role in dry eye. J. Ophthalmol. 2016, 2016, 7542929. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, S.; Ali, M.J.; Vemuganti, G.K. Human lacrimal gland regeneration: Perspectives and review of literature. Saudi J. Ophthalmol. 2014, 28, 12–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khwaja, A. Akt is more than just a Bad kinase. Nature 1999, 401, 33–34. [Google Scholar] [CrossRef] [PubMed]
- Viatour, P.; Merville, M.P.; Bours, V.; Chariot, A. Phosphorylation of NF-kappaB and IkappaB proteins: Implications in cancer and inflammation. Trends Biochem. Sci. 2005, 30, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Chávez, G.; Hernández-Ramírez, E.; Osorio-Paz, I.; Hernández-Espinosa, C.; Salceda, R. Potential role of endoplasmic reticulum stress in pathogenesis of diabetic retinopathy. Neurochem. Res. 2016, 41, 1098–1106. [Google Scholar] [CrossRef]
- Bose, T.; Diedrichs-Möhring, M.; Wildner, G. Dry eye disease and uveitis: A closer look at immune mechanisms in animal models of two ocular autoimmune diseases. Autoimmun. Rev. 2016, 15, 1181–1192. [Google Scholar] [CrossRef]
- An, E.; Gordish-Dressman, H.; Hathout, Y. Effect of TNF-α on human ARPE-19-secreted proteins. Mol. Vis. 2008, 14, 2292. [Google Scholar]
- Salminen, A.; Kauppinen, A.; Hyttinen, J.M.; Toropainen, E.; Kaarniranta, K. Endoplasmic reticulum stress in age-related macular degeneration: Trigger for neovascularization. Mol. Med. 2010, 16, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, J.J.; Yu, Q.; Wang, M.; Zhang, S.X. Endoplasmic reticulum stress is implicated in retinal inflammation and diabetic retinopathy. FEBS Lett. 2009, 583, 1521–1527. [Google Scholar] [CrossRef] [Green Version]
- Krzystolik, M.G.; Afshari, M.A.; Adamis, A.P.; Gaudreault, J.; Gragoudas, E.S.; Michaud, N.A.; Li, W.; Connolly, E.; O’Neill, C.A.; Miller, J.W. Prevention of experimental choroidal neovascularization with intravitreal anti–vascular endothelial growth factor antibody fragment. Arch. Ophthalmol. 2002, 120, 338–346. [Google Scholar] [CrossRef]
- Krebs, J.; Agellon, L.B.; Michalak, M. Ca2+ homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef]
- Pflugfelder, S.C.; de Paiva, C.S. The pathophysiology of dry eye disease: What we know and future directions for research. Ophthalmology 2017, 124, S4–S13. [Google Scholar] [CrossRef] [PubMed]
- Clayton, J.A. Dry eye. N. Engl. J. Med. 2018, 378, 2212–2223. [Google Scholar] [CrossRef]
- Kang, W.S.; Jung, E.; Kim, J. Aucuba japonica extract and aucubin prevent desiccating stress-induced corneal epithelial cell injury and improve tear secretion in a mouse model of dry eye disease. Molecules 2018, 23, 2599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, E.; Oliver, K.M.; Pearce, E.I.; Tomlinson, A.; Simmons, P.; Hagan, S. Effect of tear supplements on signs, symptoms and inflammatory markers in dry eye. Cytokine 2018, 105, 37–44. [Google Scholar] [CrossRef]
- Devecı, H.; Kobak, S. The efficacy of topical 0.05% cyclosporine A in patients with dry eye disease associated with Sjögren’s syndrome. Int. Ophthalmol. 2014, 34, 1043–1048. [Google Scholar] [CrossRef]
- McMonnies, C.W. Conjunctival tear layer temperature, evaporation, hyperosmolarity, inflammation, hyperemia, tissue damage, and symptoms: A review of an amplifying cascade. Curr. Eye Res. 2017, 42, 1574–1584. [Google Scholar] [CrossRef]
- Guzmán, M.; Keitelman, I.; Sabbione, F.; Trevani, A.S.; Giordano, M.N.; Galletti, J.G. Mucosal tolerance disruption favors disease progression in an extraorbital lacrimal gland excision model of murine dry eye. Exp. Eye Res. 2016, 151, 19–22. [Google Scholar] [CrossRef] [PubMed]
- De Paiva, C.S.; Corrales, R.M.; Villarreal, A.L.; Farley, W.J.; Li, D.-Q.; Stern, M.E.; Pflugfelder, S.C. Corticosteroid and doxycycline suppress MMP-9 and inflammatory cytokine expression, MAPK activation in the corneal epithelium in experimental dry eye. Exp. Eye Res. 2006, 83, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Aragona, P.; Aguennouz, M.H.; Rania, L.; Postorino, E.; Sommario, M.S.; Roszkowska, A.M.; De Pasquale, M.G.; Pisani, A.; Puzzolo, D. Matrix metalloproteinase 9 and transglutaminase 2 expression at the ocular surface in patients with different forms of dry eye disease. Ophthalmology 2015, 122, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Tornatore, L.; Thotakura, A.K.; Bennett, J.; Moretti, M.; Franzoso, G. The nuclear factor kappa B signaling pathway: Integrating metabolism with inflammation. Trends Cell Biol. 2012, 22, 557–566. [Google Scholar] [CrossRef]
- Ricciotti, E.; FitzGerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Role of chlorogenic acids in controlling oxidative and inflammatory stress conditions. Nutrients 2016, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Abdel Motaal, A.; Ezzat, S.M.; Tadros, M.G.; El-Askary, H.I. In vivo anti-inflammatory activity of caffeoylquinic acid derivatives from Solidago virgaurea in rats. Pharm. Biol. 2016, 54, 2864–2870. [Google Scholar] [CrossRef] [Green Version]
- Newton, K.; Dixit, V.M. Signaling in innate immunity and inflammation. Cold Spring Harb. Perspect. Biol. 2012, 4, a006049. [Google Scholar] [CrossRef]
- Kyriakis, J.M.; Avruch, J. Mammalian MAPK signal transduction pathways activated by stress and inflammation: A 10-year update. Physiol. Rev. 2012, 2, 807–869. [Google Scholar] [CrossRef] [Green Version]
- Whitcup, S.M.; Nussenblatt, R.B.; Lightman, S.L.; Hollander, D.A. Inflammation in retinal disease. Int. J. Inflamm. 2013, 2013, 724648. [Google Scholar] [CrossRef] [PubMed]
- Sprenkle, N.T.; Sims, S.G.; Sánchez, C.L.; Meares, G.P. Endoplasmic reticulum stress and inflammation in the central nervous system. Mol. Neurodegener. 2017, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kroeger, H.; Chiang, W.C.; Felden, J.; Nguyen, A.; Lin, J.H. ER stress and unfolded protein response in ocular health and disease. FEBS J. 2019, 286, 399–412. [Google Scholar] [CrossRef]
- Zhang, S.X.; Sanders, E.; Fliesler, S.J.; Wang, J.J. Endoplasmic reticulum stress and the unfolded protein responses in retinal degeneration. Exp. Eye Res. 2014, 125, 30–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goyal, S.; Chauhan, S.K.; El Annan, J.; Nallasamy, N.; Zhang, Q.; Dana, R. Evidence of corneal lymphangiogenesis in dry eye disease: A potential link to adaptive immunity? Arch. Ophthalmol. 2010, 128, 819–824. [Google Scholar] [CrossRef] [Green Version]
- Titchenell, P.M.; Antonetti, D.A. Using the past to inform the future: Anti-VEGF therapy as a road map to develop novel therapies for diabetic retinopathy. Diabetes 2013, 62, 1808–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolar, M. Glycemic control and complications in type 2 diabetes mellitus. Am. J. Med. 2010, 123, S3–S11. [Google Scholar] [CrossRef]
- Shin, E.H.; Park, S.J. Component analysis and antioxidant activity of Aster koraiensis Nakai. J. Korean Soc. Food Sci. Nutr. 2014, 43, 74–79. [Google Scholar] [CrossRef]
- Ha, J.-H.; Shil, P.K.; Zhu, P.; Gu, L.; Li, Q.; Chung, S. Ocular inflammation and endoplasmic reticulum stress are attenuated by supplementation with grape polyphenols in human retinal pigmented epithelium cells and in C57BL/6 mice. J. Nutr. 2014, 144, 799–806. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Dilution Factor | Corporation | |
|---|---|---|---|
| Primary antibody | phospho-p65 | 1:1000 | Cell signaling |
| p65 | 1:1000 | Cell signaling | |
| phospho-ERK | 1:1000 | Cell signaling | |
| ERK | 1:1000 | Santa Cruz | |
| phospho-JNK | 1:1000 | Cell signaling | |
| JNK | 1:1000 | Santa Cruz | |
| phospho-p38 | 1:1000 | Cell signaling | |
| p38 | 1:1000 | Santa Cruz | |
| phospho-AMPK | 1:1000 | Cell signaling | |
| AMPK | 1:1000 | Santa Cruz | |
| phospho-IκB | 1:1000 | Cell signaling | |
| IκB | 1:1000 | Santa Cruz | |
| COX-1 | 1:1000 | Cell signaling | |
| COX-2 | 1:1000 | Cell signaling | |
| GAPDH | 1:2000 | Cell signaling | |
| Secondary antibody | Goat anti-mouse-HRP | 1:2000 | Santa Cruz |
| Goat anti-rabbit-HRP | 1:5000 | Thermo scientific |
| Transcript | Forward Primer | Reverse Primer | Annealing Temp. (°C) |
|---|---|---|---|
| IL-1β | TCATTGTGGCTGTGGAGAAG | GGTGTGCCGTCTTTCATTAC | 53.8 |
| TNF-α | CTCAGATCATCTTCTCAA | CAGAGCAATGACTCCAAA | 55.1 |
| TGF-β | GAAAGCCCTGTATTCCGTCTCCTT | CAACAATTCCTGGCGTTACCTTGG | 53.8 |
| IFN-γ | AGCGGCTGACTGAACTCAGATTGTA | GTCACAGTTTTCAGCTGTATAGGG | 61.2 |
| IL-6 | TTCCCTACTTCACAAGTC | GGTTTGCCGAGTAGATCT | 52.9 |
| IL-23 | CAAGCAGAACTGGCTGTTGTC | GCACCAGCGGGACATATGAA | 60.2 |
| MMP-9 | CACAACCGACGACGACGAGTTGTG | CTGTGGTGAGGCCGAATAG | 65.0 |
| β-Actin | TTGTTACCAACTGGGACGACATGG | GATCTTGATCTTCATGGTGCTAGG | 59.2 |
| GAPDH | ATGGTGAAGGTCGGTGTG | ACCAGTGGATGCAGGGAT- | 58.0 |
| 0.1 mg/mL | 0.5 mg/mL | 1 mg/mL | 5 mg/mL | 10 mg/mL | |
|---|---|---|---|---|---|
| DPPH radical scavenging activity (mg ascorbic acid/g) | 30.8 ± 2.55 b | 80.0 ± 3.05 b | 112.7 ± 3.02 a | 117.8 ± 0.58 a | 118.4 ± 0.55 a |
| FRAP (nM FeSO4/mL) | 0.2 ± 0.01 e | 1.0 ± 0.02 d | 2.0 ± 0.10 c | 7.8 ± 0.41 b | 9.7 ± 0.07 a |
| TEAC (g TE/mL) | 0.2 ± 0.01 e | 0.4 ± 0.02 d | 0.9 ± 0.10 c | 2.7 ± 0.77 b | 2.9 ± 0.01 a |
| 0.1 mg/mL | 0.5 mg/mL | 1 mg/mL | 5 mg/mL | 10 mg/mL | |
|---|---|---|---|---|---|
| Total polyphenol content (mg GAE/mL) | 0.2 ± 0.00 e | 0.6 ± 0.01 d | 0.9 ± 0.01 c | 1.2 ± 0.01 b | 2.6 ± 0.03 a |
| Total flavonoids content (mg QE/mL) | 2.2 ± 0.58 c | 5.8 ± 1.37 c | 15.5 ± 0.61 c | 32.7 ± 1.51 b | 336.9 ± 11.06 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hong, S.-C.; Ha, J.-H.; Lee, J.K.; Jung, S.H.; Kim, J.-C. In Vivo Anti-Inflammation Potential of Aster koraiensis Extract for Dry Eye Syndrome by the Protection of Ocular Surface. Nutrients 2020, 12, 3245. https://doi.org/10.3390/nu12113245
Hong S-C, Ha J-H, Lee JK, Jung SH, Kim J-C. In Vivo Anti-Inflammation Potential of Aster koraiensis Extract for Dry Eye Syndrome by the Protection of Ocular Surface. Nutrients. 2020; 12(11):3245. https://doi.org/10.3390/nu12113245
Chicago/Turabian StyleHong, Sung-Chul, Jung-Heun Ha, Jennifer K. Lee, Sang Hoon Jung, and Jin-Chul Kim. 2020. "In Vivo Anti-Inflammation Potential of Aster koraiensis Extract for Dry Eye Syndrome by the Protection of Ocular Surface" Nutrients 12, no. 11: 3245. https://doi.org/10.3390/nu12113245