Potential of Moringa oleifera to Improve Glucose Control for the Prevention of Diabetes and Related Metabolic Alterations: A Systematic Review of Animal and Human Studies

 , ,
, ,
Abstract
:1. Introduction
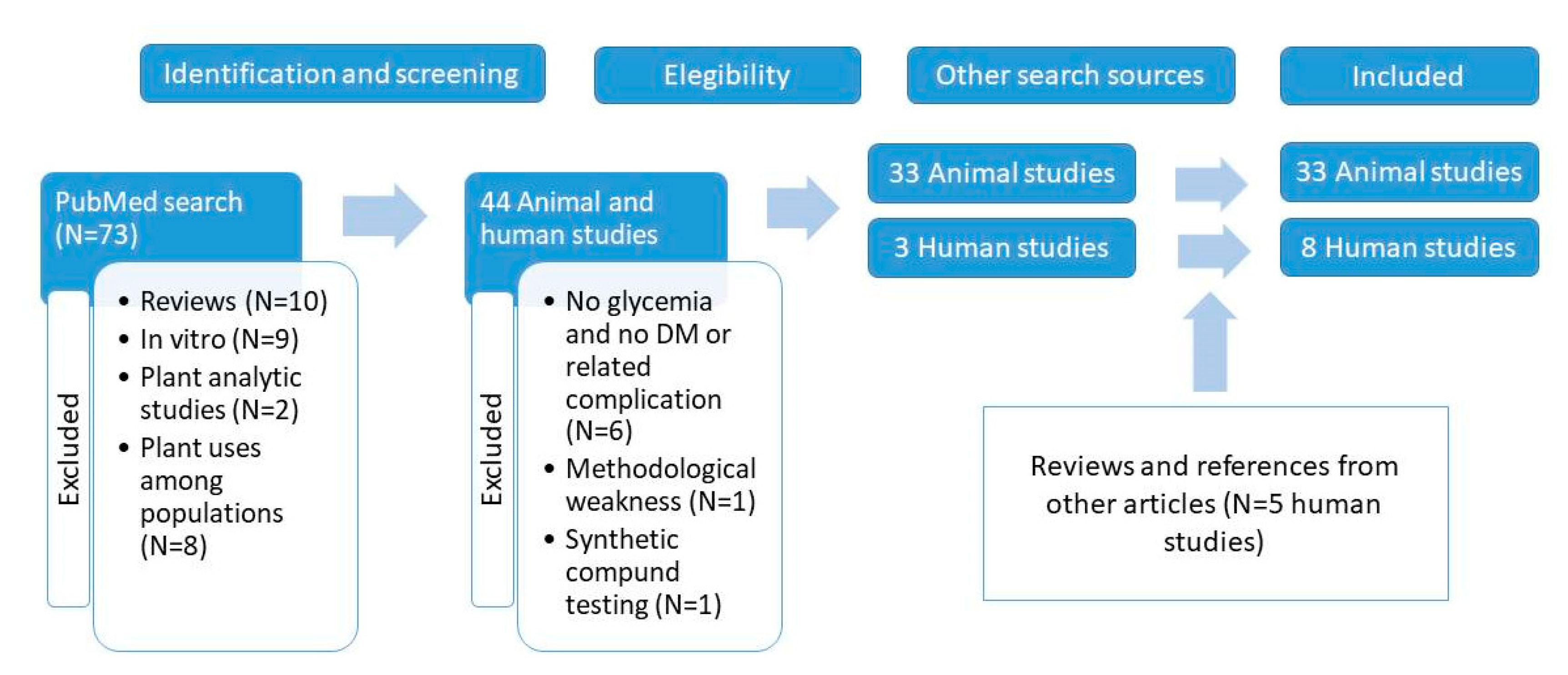
2. Materials and Methods
3. Scientific Evidences of M. oleifera’s Effect on Glucose Control in Animal Models
3.1. Experiments with Raw M. oleifera Leaves or Seeds
3.2. Experiments with M. oleifera Aqueous Extract
3.2.1. Acute Effects
3.2.2. Long-Term Effects
3.3. Experiments with M. oleifera Methanolic or Ethanolic Extracts
3.3.1. Acute Effects
3.3.2. Long-Term Effects Measured through Glucose Oral Challenge
3.3.3. Long-Term Effects Measured through Fasting Blood Glucose
4. Scientific Evidences of M. oleifera Effect on Glucose Control in Human Studies
5. Dose Comparison between Animal and Human Studies
6. Scientific Evidences of M. oleifera Effect on Metabolic Alterations Related to Diabetes
7. Final Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Gandji, K.; Chadare, F.; Idohou, R.; Salako, V.; Assogbadjo, A.; Kakaï, R.G. Status and utilisation of Moringa oleifera Lam: A review. Afr. Crop. Sci. J. 2018, 26, 137–156. [Google Scholar] [CrossRef] [Green Version]
- Fejer, J.; Kron, I.; Pellizzeri, V.; Pľuchtová, M.; Eliašová, A.; Campone, L.; Gervasi, T.; Bartolomeo, G.; Cicero, N.; Babejová, A.; et al. First Report on Evaluation of Basic Nutritional and Antioxidant Properties of Moringa Oleifera Lam. from Caribbean Island of Saint Lucia. Plants 2019, 8, 537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez, A.; Sánchez, T.; Armengol, N.; Reyes, F. Características y potencialidades de Moringa oleifera, Lamark: Una alternativa para la alimentación animal. Pastos Forrajes 2010, 33, 1. [Google Scholar]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Cultivation, Genetic, Ethnopharmacology, Phytochemistry and Pharmacology of Moringa oleifera Leaves: An Overview. Int. J. Mol. Sci. 2015, 16, 12791–12835. [Google Scholar] [CrossRef] [PubMed]
- Aluko, O.; Brai, M.R.; Adelore, A.O. Evaluation of sensory attributes of snack from maize-moringa seed flour blends. Int. J. Nut. Food Eng. 2013, 7, 944–946. [Google Scholar]
- Ogunsina, B.S.; Radha, C.; Indrani, D. Quality characteristics of bread and cookies enriched with debittered Moringa oleifera seed flour. Int. J. Food Sci. Nutr. 2010, 62, 185–194. [Google Scholar] [CrossRef]
- Anwar, F.; Latif, S.; Ashraf, M.; Gilani, A.H. Moringa oleifera: A food plant with multiple medicinal uses. Phytother. Res. 2006, 21, 17–25. [Google Scholar] [CrossRef]
- Ogunsina, B.S.; Indira, T.N.; Bhatnagar, A.S.; Radha, C.; Debnath, S.; Gopala Krishna, A.G. Quality characteristics and stability of Moringa oleifera seed oil of Indian origin. J. Food Sci. Technol. 2014, 51, 503–510. [Google Scholar] [CrossRef] [Green Version]
- FDA. Food and Drug Administration Agency Response Letter 2001. G.R.A Notice. 000069; FDA: Washington, DC, USA, 2001.
- Fahey, J.W.; Wade, K.; Stephenson, K.K.; Shi, Y.; Liu, H.; Panjwani, A.A.; Warrick, C.R.; Olson, M.E. A Strategy to Deliver Precise Oral Doses of the Glucosinolates or Isothiocyanates from Moringa oleifera Leaves for Use in Clinical Studies. Nutrients 2019, 11, 1547. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Sivanesan, I.; Keum, Y.-S. Phytochemicals of Moringa oleifera: A review of their nutritional, therapeutic and industrial significance. 3 Biotech 2016, 6, 203. [Google Scholar] [CrossRef] [Green Version]
- Leone, A.; Bertoli, S.; Di Lello, S.; Bassoli, A.; Ravasenghi, S.; Borgonovo, G.; Forlani, F.; Battezzati, A. Effect of Moringa oleifera Leaf Powder on Postprandial Blood Glucose Response: In Vivo Study on Saharawi People Living in Refugee Camps. Nutrients 2018, 10, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.E.; Sankaran, R.P.; Fahey, J.W.; Grusak, M.A.; Odee, D.; Nouman, W. Leaf Protein and Mineral Concentrations across the “Miracle Tree” Genus Moringa. PLoS ONE 2016, 11, e0159782. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Chen, X. Current Status and Potential of Moringa oleifera Leaf as an Alternative Protein Source for Animal Feeds. Front. Vet. Sci. 2020, 7, 53. [Google Scholar] [CrossRef] [PubMed]
- Rébufa, C.; Pany, I.; Bombarda, I. NIR spectroscopy for the quality control of Moringa oleifera (Lam.) leaf powders: Prediction of minerals, protein and moisture contents. Food Chem. 2018, 261, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Busani, M.; Patrick, J.M.; Arnold, H.; Voster, M.; Moyo, B.; Masika, P.; Hugo, A.; Muchenje, V. Nutritional characterization of Moringa (Moringa oleifera Lam.) leaves. Afr. J. Biotechnol. 2011, 10, 12925–12933. [Google Scholar] [CrossRef] [Green Version]
- Stohs, S.J.; Hartman, M.J. Review of the Safety and Efficacy of Moringa oleifera. Phytother. Res. 2015, 29, 796–804. [Google Scholar] [CrossRef]
- Witt, K.A. The Nutrient Content of Moringa oleifera Leaves. Echo note Nº1. 2013. Available online: https://miracletrees.org/moringa-doc/nutrient-content-of-moringa-oleifera-leaves.pdf (accessed on 22 June 2020).
- Kumssa, D.B.; Joy, E.J.; Young, S.D.; Odee, D.W.; Broadley, M.R.; Broadley, M.R. Variation in the mineral element concentration of Moringa oleifera Lam. and M. stenopetala (Bak. f.) Cuf.: Role in human nutrition. PLoS ONE 2017, 12, e0175503. [Google Scholar] [CrossRef] [Green Version]
- Luetragoon, T.; Sranujit, R.P.; Noysang, C.; Thongsri, Y.; Potup, P.; Suphrom, N.; Nuengchamnong, N.; Usuwanthim, K. Bioactive Compounds in Moringa oleifera Lam. Leaves Inhibit the Pro-Inflammatory Mediators in Lipopolysaccharide-Induced Human Monocyte-Derived Macrophages. Molecules 2020, 25, 191. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Bhattacharya, A.; Tiwari, P.; Sahu, P.K. A review of the phytochemical and pharmacological characteristics of Moringa oleifera. J. Pharm. Bioallied Sci. 2018, 10, 181–191. [Google Scholar] [CrossRef]
- Aekthammarat, D.; Pannangpetch, P.; Tangsucharit, P. Moringa oleifera leaf extract lowers high blood pressure by alleviating vascular dysfunction and decreasing oxidative stress in L-NAME hypertensive rats. Phytomedicine 2019, 54, 9–16. [Google Scholar] [CrossRef]
- Arulselvan, P.; Tan, W.S.; Gothai, S.; Muniandy, K.; Fakurazi, S.; Esa, N.M.; Alarfaj, A.A.; Kumar, S.S. Anti-Inflammatory Potential of Ethyl Acetate Fraction of Moringa oleifera in Downregulating the NF-κB Signaling Pathway in Lipopolysaccharide-Stimulated Macrophages. Molecules 2016, 21, 1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kou, X.; Li, B.; Olayanju, J.B.; Drake, J.M.; Chena, N. Nutraceutical or Pharmacological Potential of Moringa oleifera Lam. Nutrients 2018, 10, 343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linares Rivero, C.; Quiñones-Gálvez, J.; Pérez Martínez, A.; Carvajal Ortiz, C.; Rivas Paneca, M.; Cid Valdéz, G.A.; Pérez Gómez, L.; La Rosa González, S.; Capdesuñer Ruiz, Y.K. Obtaining phenolic extracts from the leaf of Moringa oleifera Lam by using different extraction methods. Veg. Biotechnol. 2018, 18, 47–56. [Google Scholar]
- Dou, Z.; Chen, C.; Fu, X. Bioaccessibility, antioxidant activity and modulation effect on gut microbiota of bioactive compounds from Moringa oleifera Lam. leaves during digestion and fermentation in vitro. Food Funct. 2019, 10, 5070–5079. [Google Scholar] [CrossRef] [PubMed]
- Vergara-Jimenez, M.; AlMatrafi, M.M.; Fernandez, M.L. Bioactive Components in Moringa Oleifera Leaves Protect against Chronic Disease. Antioxidants 2017, 6, 91. [Google Scholar] [CrossRef] [Green Version]
- Waterman, C.; Rojas-Silva, P.; Tumer, T.B.; Kuhn, P.; Richard, A.J.; Wicks, S.; Stephens, J.M.; Wang, Z.; Mynatt, R.; Cefalu, W.; et al. Isothiocyanate-rich Moringa oleifera extract reduces weight gain, insulin resistance, and hepatic gluconeogenesis in mice. Mol. Nutr. Food Res. 2015, 59, 1013–1024. [Google Scholar] [CrossRef] [Green Version]
- Devisetti, R.; Sreerama, Y.N.; Bhattacharya, S. Processing effects on bioactive components and functional properties of moringa leaves: Development of a snack and quality evaluation. J. Food Sci. Technol. 2015, 53, 649–657. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [Green Version]
- Tabák, A.G.; Herder, C.; Rathmann, W.; Brunner, E.J.; Kivimaki, M. Prediabetes: A high-risk state for diabetes development. Lancet 2012, 379, 2279–2290. [Google Scholar] [CrossRef] [Green Version]
- Larkin, A.T.; Hoffman, C.; Stevens, A.; Douglas, A.; Bloomgarden, Z. Determinants of adherence to diabetes treatment. J. Diabetes 2015, 7, 864–871. [Google Scholar] [CrossRef]
- Coskun, O.; Kanter, M.; Korkmaz, A.; Oter, S. Quercetin, a flavonoid antioxidant, prevents and protects streptozotocin-induced oxidative stress and β-cell damage in rat pancreas. Pharmacol. Res. 2005, 51, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of .ALPHA.-Glucosidase and .ALPHA.-Amylase by Flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboh, G.; Agunloye, O.; Adefegha, S.A.; Akinyemi, A.; Ademiluyi, A.O. Caffeic and chlorogenic acids inhibit key enzymes linked to type 2 diabetes (in vitro): A comparative study. J. Basic Clin. Physiol. Pharmacol. 2015, 26. [Google Scholar] [CrossRef] [PubMed]
- Bin Azad, S.; Ansari, P.; Azam, S.; Hossain, S.M.; Shahid, M.I.-B.; Hasan, M.; Hannan, J. Anti-hyperglycaemic activity of Moringa oleifera is partly mediated by carbohydrase inhibition and glucose-fibre binding. Biosci. Rep. 2017, 37. [Google Scholar] [CrossRef] [Green Version]
- Hanhineva, K.; Törrönen, R.; Bondia-Pons, I.; Pekkinen, J.; Kolehmainen, M.; Mykkänen, H.; Poutanen, K. Impact of Dietary Polyphenols on Carbohydrate Metabolism. Int. J. Mol. Sci. 2010, 11, 1365–1402. [Google Scholar] [CrossRef]
- Ader, P.; Blöck, M.; Pietzsch, S.; Wolffram, S. Interaction of quercetin glucosides with the intestinal sodium/glucose co-transporter (SGLT-1). Cancer Lett. 2001, 162, 175–180. [Google Scholar] [CrossRef]
- Ndong, M.; Uehara, M.; Katsumata, S.-I.; Suzuki, K. Effects of Oral Administration of Moringa oleifera Lam on Glucose Tolerance in Goto-Kakizaki and Wistar Rats. J. Clin. Biochem. Nutr. 2007, 40, 229–233. [Google Scholar] [CrossRef] [Green Version]
- Villarruel-Lopez, A.; La Mora, D.A.L.-D.; Vázquez-Paulino, O.D.; Puebla-Mora, A.G.; Torres-Vitela, M.R.; Guerrero-Quiroz, L.A.; Nuño, K. Effect of Moringa oleifera consumption on diabetic rats. BMC Complement. Altern. Med. 2018, 18, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oboh, G.; Oyeleye, S.I.; Akintemi, O.A.; Olasehinde, T.A. Moringa oleifera supplemented diet modulates nootropic-related biomolecules in the brain of STZ-induced diabetic rats treated with acarbose. Metab. Brain Dis. 2018, 33, 457–466. [Google Scholar] [CrossRef]
- Al-Malki, A.L.; El Rabey, H.A. The Antidiabetic Effect of Low Doses of Moringa oleifera Lam. Seeds on Streptozotocin Induced Diabetes and Diabetic Nephropathy in Male Rats. Biomed. Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Khan, W.; Parveen, R.; Chester, K.; Parveen, S.; Ahmad, S. Hypoglycemic Potential of Aqueous Extract of Moringa oleifera Leaf and In Vivo GC-MS Metabolomics. Front. Pharmacol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, L.; Rai, P.K.; Kumar, A.; Mehta, S.; Watal, G. Effect of Moringa oleifera Lam. leaves aqueous extract therapy on hyperglycemic rats. J. Ethnopharmacol. 2009, 123, 392–396. [Google Scholar] [CrossRef] [PubMed]
- Fombang, E.N.; Saa, R.W. Antihyperglycemic Activity of Moringa oleifera Lam Leaf Functional Tea in Rat Models and Human Subjects. Food Nutr. Sci. 2016, 7, 1021–1032. [Google Scholar] [CrossRef] [Green Version]
- Eldaim, M.A.A.; Elrasoul, A.S.A.; Elaziz, S.A.A.; Shaban, A. An aqueous extract from Moringa oleifera leaves ameliorates hepatotoxicity in alloxan-induced diabetic rats. Biochem. Cell Biol. 2017, 95, 524–530. [Google Scholar] [CrossRef] [PubMed]
- El Latif, A.A.; Bialy, B.E.S.E.; Mahboub, H.D.; Eldaim, M.A. Moringa oleifera leaf extract ameliorates alloxan-induced diabetes in rats by regeneration of β cells and reduction of pyruvate carboxylase expression. Biochem. Cell Biol. 2014, 92, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Adepoju-Bello, A.A.; Jolayemi, O.M.; Ehianeta, T.S.; Ayoola, G.A. Preliminary phytochemical screening, antioxidant and antihyperglycaemic activity of Moringa oleifera leaf extracts. Pak. J. Pharm. Sci. 2017, 30, 2217–2222. [Google Scholar]
- Tuorkey, M.J. Effects of Moringa oleifera aqueous leaf extract in alloxan induced diabetic mice. Interv. Med. Appl. Sci. 2016, 8, 109–117. [Google Scholar] [CrossRef]
- Yassa, H.D.; Tohamy, A.F. Extract of Moringa oleifera leaves ameliorates streptozotocin-induced Diabetes mellitus in adult rats. Acta Histochem. 2014, 116, 844–854. [Google Scholar] [CrossRef]
- Gupta, S.K.; Kumar, B.; Srinivasan, B.; Nag, T.C.; Srivastava, S.; Saxena, R.; Aggarwal, A. Retinoprotective Effects of Moringa oleifera via Antioxidant, Anti-Inflammatory, and Anti-Angiogenic Mechanisms in Streptozotocin-Induced Diabetic Rats. J. Ocul. Pharmacol. Ther. 2013, 29, 419–426. [Google Scholar] [CrossRef]
- Mohamed, M.A.; Ahmed, M.A.; El Sayed, R.A. Molecular effects of Moringa leaf extract on insulin resistance and reproductive function in hyperinsulinemic male rats. J. Diabetes Metab. Disord. 2019, 18, 487–494. [Google Scholar] [CrossRef]
- Paula, P.C.; Sousa, D.O.B.; Oliveira, J.T.A.; Carvalho, A.F.U.; Alves, B.G.T.; Pereira, M.L.; Farias, D.F.; Viana, M.P.; Santos, F.A.; Morais, T.C.; et al. A Protein Isolate from Moringa oleifera Leaves Has Hypoglycemic and Antioxidant Effects in Alloxan-Induced Diabetic Mice. Molecules 2017, 22, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, H.; Kim, B.; Park, H.; Lee, K.; Kim, H.-H.; Sim, H.-C.; Do, H.-J.; Hyun, C.-K.; Do, M.-S. Fermented Moringa oleifera Decreases Hepatic Adiposity and Ameliorates Glucose Intolerance in High-Fat Diet-Induced Obese Mice. J. Med. Food 2017, 20, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Olayaki, L.A.; Irekpita, J.E.; Yakubu, M.T.; Ojo, O. Methanolic extract of Moringa oleifera leaves improves glucose tolerance, glycogen synthesis and lipid metabolism in alloxan-induced diabetic rats. J. Basic Clin. Physiol. Pharmacol. 2015, 26. [Google Scholar] [CrossRef] [PubMed]
- Jaja-Chimedza, A.; Zhang, L.; Wolff, K.; Graf, B.L.; Kühn, P.; Moskal, K.; Carmouche, R.; Newman, S.; Salbaum, J.M.; Raskin, I. A dietary isothiocyanate-enriched moringa (Moringa oleifera) seed extract improves glucose tolerance in a high-fat-diet mouse model and modulates the gut microbiome. J. Funct. Foods 2018, 47, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Omodanisi, E.I.; Aboua, Y.G.; Oguntibeju, O.O. Assessment of the Anti-Hyperglycaemic, Anti-Inflammatory and Antioxidant Activities of the Methanol Extract of Moringa Oleifera in Diabetes-Induced Nephrotoxic Male Wistar Rats. Molecules 2017, 22, 439. [Google Scholar] [CrossRef]
- Omodanisi, E.I.; Aboua, Y.G.; Chegou, N.N.; Oguntibeju, O.O. Hepatoprotective, Antihyperlipidemic and Anti-inflammatory Activity of Moringa oleifera in Diabetic-induced Damage in Male Wistar Rats. Pharmacogn. Res. 2017, 9, 182–187. [Google Scholar]
- Aju, B.; Rajalakshmi, R.; Mini, S. Protective role of Moringa oleifera leaf extract on cardiac antioxidant status and lipid peroxidation in streptozotocin induced diabetic rats. Heliyon 2019, 5, e02935. [Google Scholar] [CrossRef]
- Gupta, R.; Mathur, M.; Bajaj, V.K.; Katariya, P.; Yadav, S.; Kamal, R.; Gupta, R.S. Evaluation of antidiabetic and antioxidant activity of Moringa oleifera in experimental diabetes. J. Diabetes 2012, 4, 164–171. [Google Scholar] [CrossRef]
- Sánchez-Muñoz, M.A.; Solana, M.A.V.; Almazán, M.I.C.; Flores-Herrera, O.; Esparza-Perusquía, M.; Olvera-Sanchez, S.; García-Arenas, G.; Avitia-Domínguez, C.; Tellez-Valencia, A.; Sierra-Campos, E. Streptozotocin-Induced Adaptive Modification of Mitochondrial Supercomplexes in Liver of Wistar Rats and the Protective Effect of Moringa oleifera Lam. Biochem. Res. Int. 2018, 2018, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tang, Y.; Choi, E.-J.; Han, W.C.; Oh, M.; Kim, J.; Hwang, J.-Y.; Park, P.-J.; Moon, S.-H.; Kim, Y.-S.; Kim, E.-K. Moringa oleifera from Cambodia Ameliorates Oxidative Stress, Hyperglycemia, and Kidney Dysfunction in Type 2 Diabetic Mice. J. Med. Food 2017, 20, 502–510. [Google Scholar] [CrossRef]
- Omabe, M.; Nwudele, C.; Omabe, K.N.; Okorocha, A.E. Anion Gap Toxicity in Alloxan Induced Type 2 Diabetic Rats Treated with Antidiabetic Noncytotoxic Bioactive Compounds of Ethanolic Extract of Moringa oleifera. J. Toxicol. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kar, A.; Choudhary, B.K.; Bandyopadhyay, N.G. Comparative evaluation of hypoglycaemic activity of some Indian medicinal plants in alloxan diabetic rats. J. Ethnopharmacol. 2003, 84, 105–108. [Google Scholar] [CrossRef]
- Olurishe, C.; Kwanashie, H.; Zezi, A.; Danjuma, N.; Mohammed, B. Chronic administration of ethanol leaf extract of Moringa oleifera Lam. (Moringaceae) may compromise glycaemic efficacy of Sitagliptin with no significant effect in retinopathy in a diabetic rat model. J. Ethnopharmacol. 2016, 194, 895–903. [Google Scholar] [CrossRef] [PubMed]
- Raafat, K.; Hdaib, F. Neuroprotective effects of Moringa oleifera: Bio-guided GC-MS identification of active compounds in diabetic neuropathic pain model. Chin. J. Integr. Med. 2017, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Xiao, J.; Weng, Z.; Lu, X.; Shen, X.; Wang, F. A phenolic glycoside from Moringa oleifera Lam. improves the carbohydrate and lipid metabolisms through AMPK in db/db mice. Food Chem. 2020, 311, 125948. [Google Scholar] [CrossRef]
- Wang, F.; Zhong, H.-H.; Chen, W.-K.; Liu, Q.-P.; Li, C.; Zheng, Y.; Peng, G.-P. Potential hypoglycaemic activity phenolic glycosides from Moringa oleifera seeds. Nat. Prod. Res. 2016, 31, 1869–1874. [Google Scholar] [CrossRef]
- Ahmad, J.; Khan, I.; Johnson, S.K.; Alam, I.; Din, Z.U. Effect of Incorporating Stevia and Moringa in Cookies on Postprandial Glycemia, Appetite, Palatability, and Gastrointestinal Well-Being. J. Am. Coll. Nutr. 2017, 37, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Taweerutchana, R.; Lumlerdkij, N.; Vannasaeng, S.; Akarasereenont, P.; Sriwijitkamol, A. Effect of Moringa oleifera Leaf Capsules on Glycemic Control in Therapy-Naïve Type 2 Diabetes Patients: A Randomized Placebo Controlled Study. Evid. Based Complement. Altern. Med. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, S.; Chawla, P.; Kochhar, A. Effect of supplementation of drumstick (Moringa oleifera) and amaranth (Amaranthus tricolor) leaves powder on antioxidant profile and oxidative status among postmenopausal women. J. Food Sci. Technol. 2012, 51, 3464–3469. [Google Scholar] [CrossRef] [Green Version]
- Kumari, D.J. Hypoglycemic effect of Moringa oleifera and Azadirachta indica in type-2 diabetes. Bioscan 2010, 5, 211–214. [Google Scholar]
- Giridhari, V.A.; Malathi, D.; Geetha, K. Anti diabetic property of drumstick (Moringa oleifera) leaf tablets. Int. J. Health Nutr. 2011, 2, 1–5. [Google Scholar]
- Anthanont, P.; Lumlerdkij, N.; Akarasereenont, P.; Vannasaeng, S.; Sriwijitkamol, A. Moringa oleifera Leaf Increases Insulin Secretion after Single Dose Administration: A Preliminary Study in Healthy Subjects. J. Med. Assoc. Thail. 2016, 99, 308–313. [Google Scholar]
- Muhammad, A.A.; Arulselvan, P.; Cheah, P.S.; Abas, F.; Fakurazi, S. Evaluation of wound healing properties of bioactive aqueous fraction from Moringa oleifera Lam on experimentally induced diabetic animal model. Drug Des. Dev. Ther. 2016, 10, 1715–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaiswal, D.; Rai, P.K.; Mehta, S.; Chatterji, S.; Shukla, S.; Rai, D.K.; Sharma, G.; Sharma, B.; Khair, S.; Watal, G. Role of Moringa oleifera in regulation of diabetes-induced oxidative stress. Asian Pac. J. Trop. Med. 2013, 6, 426–432. [Google Scholar] [CrossRef]
- Gheibi, S.; Kashfi, K.; Ghasemi, A. A practical guide for induction of type-2 diabetes in rat: Incorporating a high-fat diet and streptozotocin. Biomed. Pharmacother. 2017, 95, 605–613. [Google Scholar] [CrossRef]
- King, A.J.F. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Reed, M.; Meszaros, K.; Entes, L.; Claypool, M.; Pinkett, J.; Gadbois, T.; Reaven, G. A new rat model of type 2 diabetes: The fat-fed, streptozotocin-treated rat. Metabolism 2000, 49, 1390–1394. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [Green Version]
- Asiedu-Gyekye, I.J.; Frimpong-Manso, S.; Awortwe, C.; Antwi, D.A.; Nyarko, A.K. Micro- and Macroelemental Composition and Safety Evaluation of the Nutraceutical Moringa oleifera Leaves. J. Toxicol. 2014, 2014, 1–13. [Google Scholar] [CrossRef]
- Dhakad, A.K.; Ikram, M.; Sharma, S.; Khan, S.; Pandey, V.V.; Singh, A. Biological, nutritional, and therapeutic significance of Moringa oleifera Lam. Phytotherapy Res. 2019, 33, 2870–2903. [Google Scholar] [CrossRef]
- Falowo, A.B.; Mukumbo, F.E.; Idamokoro, E.M.; Lorenzo, J.M.; Afolayan, A.J.; Muchenje, V. Multi-functional application of Moringa oleifera Lam. in nutrition and animal food products: A review. Food Res. Int. 2018, 106, 317–334. [Google Scholar] [CrossRef] [PubMed]
- Ajibade, T.O.; Arowolo, R.; Olayemi, F. Phytochemical screening and toxicity studies on the methanol extract of the seeds of Moringa oleifera. J. Complement. Integr. Med. 2013, 10, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Davison, G.W.; George, L.; Jackson, S.K.; Young, I.S.; Davies, B.; Bailey, D.M.; Peters, J.R.; Ashton, T. Exercise, free radicals, and lipid peroxidation in type 1 diabetes mellitus. Free. Radic. Biol. Med. 2002, 33, 1543–1551. [Google Scholar] [CrossRef]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; De Bittencourt, P.I.H.; De Bittencourt, P.I.H. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Model | Treatment and Duration | Measurements | Evidences: MO Treated Animals vs. Untreated | Ref |
|---|---|---|---|---|
| Studies with raw MO | ||||
| Alloxan-induced diabetic Sprague Dawley rats. Normal Sprague Dawley rats | MO dry leaf powder (50 mg/day, gavage) 8 wk. | Body weight, BG, lipid profile. Intestine histopathology, lactic acid bacteria and Enterobacteriaceae (culture). | ↓ BG. Prevented weight loss. No effect on lipid profile. No histopathology observations. No effect in BG in normal rats. ↓ Enterobacteria enumeration | [40] |
| STZ-diabetic male Wistar rats | Diets containing 2% and 4% MO leaves or MO seeds (± Acarbose, ACA) 2 wk. | BG (every 3 days), acetylcholinesterase (AChE), butyrylcholinesterase (BChE)] angiotensin-I converting enzyme (ACE), arginase, CAT, GST and GSH-Px activities, GSH and nitric oxide (NO) levels in brain | ↓ BG (all treated groups). The highest reduction occurred with 4% MO leaves + ACA. ↓ AChE, BChE and ACE activities and ↑ antioxidant molecules (both preventive of cognitive dysfunction) | [41] |
| Male spontaneously diabetic Goto-Kakizaki rats Normal male Wistar rats | MO leaf powder (200 mg/kg) Single dose (glucose-MO solution, oral admin.) | OGTT (BG at 10, 20, 30, 45, 60, 90 and 120 min and iAUC). Stomach, small intestine and caecum content weights. | ↓ BG at 20, 30, 45, and 60 min. ↓iAUC. ↑ stomach contents (⇒delayed gastric emptying). ↓ BG at 10, 30 and 45 min in normal rats. | [39] |
| STZ-induced diabetic adult male Wistar rats | MO seed powder (50 or 100 mg/kg) in the diet 4 wk. | FBG, HbA1c, lipid peroxidation, antioxidant enzymes, liver and renal function, IgG, IgA, serum and kidney IL-6 and kidney and pancreas histopathology | Prevented weight loss, ↓ FBG (35% and 45%, 50 and 100 dose, resp.) and ↓ HbA1C (13% and 22%). Improvement of all oxidative status parameters, Igs and IL6; all approaching the negative control. Restoration of the normal histology of both kidney and pancreas. Both doses are effective but, overall, the higher dose is more effective. | [42] |
| Studies with MO aqueous extract | ||||
| STZ-diabetic female Wistar rats HFD-induced diabetic C57BL/6 mice Normal rats and mice | 1) Aq MO, 100 mg/kg (in the diet) to STZ-induced rats 2) Aq MO, 200 mg/kg (in the diet) to HFD-fed mice. Either, 2 single doses (2 days) or 3 wk. | FBG, OGTT, lipid profile, liver marker enzymes. | ↓ FBG, ↓ 2 h glucose and AUC of glucose in OGTT. ↓ SGOT and SGPT (HFD- and STZ-induced animals), improved lipid profile (more significant in HFD-mice than STZ-diabetic rats) No differences in FBG and biochemical parameters in normal rats and mice | [43] |
| STZ-induced diabetic male Wistar rats (sub, mild and severely diabetic) Normal Wistar rats | Aq MO (100, 200 and 300 mg/kg, oral gavage) Either, single dose or 21 days | BG and OGTT in response to single doses. FBG, PPG, haemoglobin, total protein and weight gain after 21 days. BG and OGTT in response to single doses | ↓ BG in OGTT (at 3 h post oral glucose): maximum fall of 31.1% and 32.8% in sub-diabetic and mild-diabetic rats, respectively, occurred always with the 200 mg/kg dose. ↓ FBG (69.2%) and ↓ PPG (51.2%) in severely diabetic rats on a 21-day treatment. ↑ Hemoglobin (10.9%) and total protein (11.3%) ↓ BG in normal rats: maximum fall of 26.7% occurred 6 h after 200 mg/kg, single dose. ↓ BG in OGTT (at 3 h post oral glucose) in normal rats: maximum fall of 29.9% with 200 mg/kg dose | [44] |
| Adult male Wistar rats | MO tea (10, 20 and 30 mL/kg, gavage) Single dose | OGTT (4 g/kg b.w. of glucose, 30 min after MO tea) | ↓16, 18 and 6% total PPG ⇒ Lower doses are more efficient. | [45] |
| Alloxan-induced diabetic Wistar rats. Normal Wistar rats | Aq MO (250 mg/kg, oral admin.) 18 days | BG, hepatic lipid peroxidation and antioxidant enzyme activities, histoarchitecture of hepatic and pancreatic tissues, gene expression of glycogen synthase (GS), pyruvate carboxylase (PC) and caspase 3, and SOD and CAT activities. | ↓ BG, prevented organ changes and significantly restored all measures. Normalized the expression of apoptotic, gluconeogenic, and glycogenic genes in hepatic tissue No effect on BG in normal rats. ↑ liver GSH and GS expression. No other significant changes in normal rats. | [46] |
| Alloxan-induced diabetic female Wistar rats. Normal Wistar rats | Aq MO (250 mg/kg, oral admin.) 18 days | Body weight, BG, lipid profile, lipid peroxidation, histoarchitecture of hepatic and pancreatic tissues, expression of pyruvate kinase (PK), pyruvate carboxylase (PC), and fatty acid synthase (FAS) in liver. | ↓ BG ↑ Body weight, ↓ TG and MDA. Normalized the expression of enzymes of gluconeogenesis and fatty acid synthesis. Normalized histological structures. No effects in normal rats. | [47] |
| Alloxan-induced diabetic Wistar rats | MO extracts: Aq, Me MO and Et MO (50% water: 50% alcohol and 100% alcohol, dose range: 200–400 mg/kg, oral admin.) 24 days | FBG | All extracts and doses ↓FBG around 70–87%. Aq MO (300 mg/kg) reduction was 82%. All extracts showed body weight restoration capacity. | [48] |
| Alloxan-Induced diabetic albino mice Normal albino mice | Aq MO (100 mg/kg, oral gavage) 14 days | FBG, insulin, HOMA-IR, TAC, creatinine, blood urea nitrogen (BUN). Percentage CD44, CD69 and IFN-γ positive cells in PBMCs | ↓FBG, ↑(NS) insulin, ↓ HOMA-IR. ↓creatinine and BUN and ↑ TAC and IFN-γ. ↑ Insulin in normal rats and no effect on FBG and HOMA-IR. No effect in other parameters measured. | [49] |
| STZ-induced diabetic male Sprague-Dawley rats Normal Sprague-Dawley rats | Aq MO (200 mg/kg, oral gavage) 8 wk | FPG; GSH, lipid peroxidation, histopathology and morphometric analyses of pancreas | ↓ FPG (62%) ↓ MDA and ↑ GSH, normalization of histopathological and morphometric changes No effects in normal rats. | [50] |
| STZ-induced diabetic rats | Aq MO (100 mg/kg, oral gavage) 24 wk. |
Body weight, BG and HbA1C. TNF-α, IL-1β, VEGF, PKC-β, GSH, SOD, CAT in retinae. Retinal leakage and retinal vessel caliber (arteriolar and venular) and basement membrane thickness. | ↓ BG (33%), and HbA1C (40%). Preserved weight gain. ↑Antioxidant parameters, ↓ inflammatory and angiogenic parameters, ↓ all morphological and structural alterations of the retinae. | [51] |
| VHFD-induced obese male C57BL/6J mice | Aq MO (5% MO in VHFD, 66 mg/d MIC) 12 wk. | OGTT at 4th, 8th and 12th wk., plasma insulin, leptin, resistin, IL-1β and TNFα, total cholesterol and triglycerides. Liver histology and gene expression: TNF-α, IL-1β, IL-6, G6Pase, PEPCK and GcK. Insulin signaling proteins (liver and muscle) and lipolysis-related gene expression and protein levels (adipose tissue and liver). | ↓BG and ↓ AUC of glucose at 8th and 12th wk (NS ↓ at wk. 4th). ↓ Weight gain, ↓ body fat accumulation, ↓ plasma insulin, leptin, resistin, cytokines and cholesterol, ↓ Hepatic G6Pase and TNF-α expression, improved insulin signaling (↑IRSs, protein kinases, PI3K, and GLUT4) and liver lipolytic protein levels. | [28] |
| High fructose diet-induced diabetic male Sprague Dawley rats | Aq MO (300 mg/kg, oral admin.) 4 wk. | FSG, Insulin, HOMA-IR, testosterone and FSH; MDA, SOD, CAT in liver; insulin receptor (IR), IRS-1, GLUT-4 & GLUT-5 and SOD, steroidogenic acute regulatory protein (StAR) and 3β-hydroxysteroid dehydrogenase (3β-HSD) expression in liver | No effect on FSG; ↑ insulin and ↓ HOMA-IR, ↓ MDA, ↑ SOD, CAT and testosterone. Improvement of the down-regulation of the insulin signaling pathway. Improved regulatory proteins of testicular function | [52] |
| Alloxan-Induced diabetic Unib:SW (Swiss) mice | Protein isolate of MO leaves (500 mg/kg) (i.p. and oral admin.) Single dose and 7 days | BG (5 h after single dose and 4 h after daily treatment at 3rd and 7th day), insulin (5 h after single dose), liver MAD, CAT and SOD. | i.p. admin.: ↓BG, ↓ liver MDA and ↑CAT. Did not change serum insulin. Oral admin.: No effect on BG (due to protein digestion). | [53] |
| Studies with MO methanolic or ethanolic extracts | ||||
| STZ-induced diabetic Long Evan rats | Et MO (0.5 g/kg, oral admin.) and Et MO with glucose (OGTT). Single dose, both | FBG, OGTT (2.5 g/kg oral glucose), insulin, intestinal glucose absorption by perfusion technique through the pylorus. | ↓ FBG at 90 min post Et MO and ↓ BG in OGTT. No change in insulin. ↓ Glucose absorption. | [36] |
| HFD-induced obese male C57BL/6J mice | Me MO (250 mg/kg, oral admin.), fermented (FM) and non-fermented (NFM). Fermentation starter: 3 LAB strains isolated from cabbage kimchi 10 wk. | Glucose tolerance (2 g/kg glucose, ip. injection) at 8th wk., hepatic lipid accumulation, expression of proteins and genes involved in glucose and lipid regulation | FM: ↓ AUC glucose, ↓ hepatic lipid accumulation. Upregulation of genes related to lipid metabolism. ↓ Oxidative stress and lipotoxicity in muscle. ↓ proinflammatory cytokine expression in muscle, and liver tissues. NFM: No effect on glycemic response. ↓ Hepatic lipid accumulation. Mixed effects on proinflammatory cytokine expression and levels in different tissues. | [54] |
| Alloxan-induced diabetic rats | Me MO (300 or 600 mg/kg, oral gavage) 6 wk. | Food intake, body weight, intraperitoneal glucose tolerance (IPGT, 30, 60 and 120 min post 2 g/kg glucose administration), serum glucose, insulin, and lipids, liver and muscle glycogen synthase activity, glycogen content, and glucose uptake. | Prevented weight loss, ↓BG and ↑Insulin, ↓BG at all-time points in IPGT. Improved lipid profile, increased glycogen synthase activity and glycogen content in muscle and liver and improved glucose uptake. | [55] |
| Obese C57Bl/6J male mice fed VHFD Normal C57Bl/6J male mice fed LFD | Et MO (seed, 47% MIC) in the diet. Average dose: 161 ± 19 mg MIC-1/kg in VHFD; 335 ± 23 mg MIC-1/kg in LFD. 12 wk. | Body weight, body composition, OGTT at week 2nd, 4th, 6th, 9th and 12th; liver lipids, IL-1β, IL-6, TNF-α, iNOS, NQO-1 gene expression, intestinal microbiota composition and load. | ↓ AUC glucose in the VHFD animals. Similar AUC glucose in LFD animals (except at week 9, treated < untreated). ↓ body weight, ↓ adiposity; ↓ iNOS expression, ↑ antioxidant NQO-1 expression (both diets); ↓ bacterial load, modulation of bacterial community (both diets) | [56] |
| STZ/HFD-induced diabetic male Wistar rats Normal Wistar rats | Me MO (250 mg/kg, gavage) 6 wk | FPG, kidney lipid peroxidation, CAT, GPx, SOD activities, GSH and inflammatory biomarkers. | ↓ FPG, ↓ kidney weight and relative kidney weight. ↓ Kidney MDA, ↑ CAT (and ↑ NS SOD), ↓ pathological observation in histology. ↓ FPG, ↑ SOD, ↓ GPx (and ↑ NS CAT) in normal rats. | [57] |
| STZ-induced diabetic female Wistar rats Normal Wistar rats | Me MO (250 mg/kg, gavage) 6 wk. | FSG, liver weight and enzymes, lipid profile, liver antioxidant capacity, inflammatory cytokine levels and histopathology. | ↓ FSG ↓ Liver weight, SGOT, ALP, LDL and cholesterol. ↓ IL-6, TNF-α, and MCP-1. Improvement of liver histological alterations Non-significant ↓ FSG in normal rats.↓ Liver enzymes and ↑ HDL in normal rats. | [58] |
| STZ-induced diabetic male Sprague Dawley rats. Normal male Sprague Dawley rats. | Me MO (300 mg/kg, oral gavage) 60 days | FBG (glucometer), FSG (ion exchange), HbA1C, Insulin, SOD, CAT, GPx, GR, GSH, TBARS. | ↓ FBG, FSG, and HbA1c and ↑ increased insulin. ↑Activities of antioxidant enzymes in the heart of diabetic rats. No effects in normal rats. | [59] |
|
STZ-induced diabetic Wistar rats Normal Wistar rats | Me MO (pods, 150 and 300 mg/kg, oral admin.) 3 wk. | BG, insulin, total protein and albumin and NO. Pancreatic lipid peroxidation, SOD, GSH, CAT and glycogen and histopathology. | ↓ BG, ↑ insulin, total protein, albumin and NO. ↓ MDA and ↑ antioxidant activities in pancreas and reversed the histoarchitectural damage. No changes in normal rats. | [60] |
| STZ-diabetic Wistar rats | Me MO (200 mg/kg, gavage) 3 wk. | FBG, weight, supercomplex formation, ATPase activity, ROS production, GSH and GR levels, lipid peroxidation and protein carbonylation of liver mitochondria | ↓ FBG: 86 ± 4.2 mg/dL vs. 229 ± 9.05 mg/dL. ↑ GSH and GR; ↓ lipid peroxidation and protein carbonylation of liver mitochondria. | [61] |
| db/db mice | Et MO (150 mg/kg) 5 wk. | FPG, lipid profile, kidney histology and expression of inflammation markers. | ↓ FPG (36%), TG and LDL. ↑ insulin. ↓ kidney histopathological damage. ↓ expression of kidney inflammatory markers | [62] |
| Alloxan-Induced diabetic Wistar rats | Et MO (200 mg/kg, twice daily, oral admin.) 5–6 days | FBG, electrolytes: potassium (K), sodium (Na), chloride (Cl−), bicarbonate and lactate dehydrogenase (LDH) enzyme. | ↓FBG. No effect on weight. No cytotoxicity. ↓ Bicarbonate, ↑ anion gap value (⇒acidosis, but lower that metformin). | [63] |
| Alloxan-induced diabetic Charles Foster strain male albino rats | Et MO (stem bark, 250 mg/kg, oral admin.) 1 wk. | FBG and urine sugar | ↓ FBG and nil urine sugar detected. | [64] |
| Alloxan-Induced diabetic rats with/w-out sitagliptin treatment | Et MO (300 mg/kg, oral gavage) 42 days | FBG and other glycaemic control parameters, insulin, body weight, retinal microvasculature on lenticular opacity/morphology | No difference in FBG between day 42nd and day 1st. ↓ random BG (42nd vs. 1st day). Overall, less anti-hyperglicemic effect than sitagliptin alone. No changes in insulin secretion and body weight. No prevention or amelioration of retina lesions. | [65] |
| Alloxan-induced Swiss-Webster male mice | A) N-Hexane extract of MO seeds (40, 60 and 80 mg/kg, i.p.) B) β-sitosterol (BSL) fraction from hexane extract (18, 25 and 35 mg/kg, i.p.) Single dose (acute) and 8 days | BG (acute: 1, 2 and 6h; subchronic: 1st, 3rd, 5th, and 8th day, 6 h post daily injection); Insulin, HbA1C, CAT, lipid peroxidation (8 wk post-treatment end); Diabetic painful neuropathy measures (hot plate latency, tail flick latency and von Frey filaments test, 8 wk post-treatment end) | Acute: ↓ BG at 2 h and 6 h with all doses of MO and BSL. Subchronic: ↓ BG from day 1 to day 8 with MO and BSL (maximum ~ 70% reduction on day 8th). ↑ Insulin several fold, ↓ HbA1C, improved antioxidant markers, 8 wk. post-treatment end. ↑ body weight (both). Significantly improved thermal hyperalgesia and tactile allodynia (MO more powerful than BSL). | [66] |
| db/db mice | Niazirin (10 mg/kg and 20 mg/kg, extracted and concentrated from seeds. 95% purity. Oral gavage) 4 wk. | FBG and Insulin, HOMA-IR, OGTT. Plasma TNF-alpha, IL-10, LDL, HDL, TC, TG, NEFA levels; Liver glycogen, HK, PK, G6Pase and PEPCK activities; Liver histological analysis; Liver AMPK, p-AMPK, FAS, p-ACC, SREBP-1, PPAR-α, SirT1, FOXO1, HNF-4α, PGC-1α, PFKFB-3. | ↓ FBG and insulin with improved HOMA-IR (both doses). OGTT: lower BG at 90 and 120 min (both doses) Improved glucose uptake and glycogen storage in the liver through AMPK pathway. Improved lipid profile; reduced FA synthesis and induced FA oxidation through AMPK pathway. | [67] |
| STZ-induced diabetic ICR mice | 4 compounds isolated from MO seeds by macroporous resin adsorption and chromatography (20 mg/kg, i.v.) 2 wk. | BG | ↓ BG, 3 compounds: 1) N,N′-bis{4-[(α-l-rhamnosyloxy)benzyl]}thiourea, 2) niazirin A, 3) S-Methyl-N-{4-[(α-l-rhamnosyloxy)benzyl]}thiocar bamate No effect: 4-[(6-deoxy-α-l-mannopyranosyl)oxy]-benzaldehyde | [68] |
| Treatment and Duration | Study Design | Subjects | Measurements | ResultsCompared to Baseline | ResultsCompared to Control Group | Ref. |
|---|---|---|---|---|---|---|
| Meal containing MO leaf powder (20 g) C: Control meal. 2 single occasions | Randomized, placebo controlled, crossover, PP | 10 healthy adults (6 W/4M)Age: 42 ± 11 y BMI: No data 17 type 2 diabetic patients (9 W/8 M) Age: 62 ± 9 y BMI: 25.2 ± 4.3 kg/m2 | Fasting glucose (finger prick and glucometer) and postprandial glucose at 30, 60, 90, 120, 150, and 180 min from the beginning of the meal | -- | Healthy: ↔ glycemic response Diab: ↓ blood glucose from 60 to 180 min. ↓ increment form baseline at 90, 120 and 150 min. | [12] |
| Cookies containing MO leaf powder (5% w/w) C: Control cookies Isocaloric and containing 50 g available carbohydrates 2 single occasions | Randomized single-blinded, placebo controlled crossover, PP | 20 Healthy subjects (10 W/10 M) Age: 24.1 ± 1.33 y BMI: 22.0 ± 3.88 kg/m2 | Fasting and postprandial blood glucose (finger prick and glucometer) at 15, 30, 45, 60, 90 and 120 min. Appetite, hunger and palatability scales | -- | ↓ non-significantly iAUC of glucose (P = 0.077). ↓ blood glucose at 30 and 45 min ↓ Hunger ratings | [69] |
| MO leaf capsules (4 g/d) C: Placebo capsules. 4 weeks | Randomized, placebo controlled, parallel | 32 Therapy-naïve type 2 diabetics (15 W/17 M) Age: 50–60 y BMI: 27.5 kg/m2 | 9-point blood glucose (finger prick and glucometer) along 3 consecutive days.FPG and HBA1C levels.Creatinine and liver enzymes (ALT, AST) | ↔ Fasting plasma glucose and HbA1C ↔ Creatinine, ALT, AST. | ↔ Fasting plasma glucose and HbA1C ↔ mean daily BG, mean premeal, and mean postmeal BG. ↔ Creatinine, ALT and AST | [70] |
| MO leaf powder (7 g/d) in recipes in daily diet C: No supplementation 3 months | Randomized, controlled, parallel | 60 Healthy postmenopausal women (60 W/0 M) Age: 45–55 y BMI: No data | FBG, hemoglobin, ascorbic acid, retinol, glutathione peroxidase, superoxide dismutase and malondialdehyde | ↓ FBG↑ Blood haemoglobin ↑ Ascorbic acid, retinol, glutathione peroxidase and superoxide dismutase ↓ Malondialdehyde | ↓ FBG, ↑ Blood haemoglobin ↑ Ascorbic acid and superoxide dismutase ↓ Malondialdehyde | [71] |
| MO leaf tea (200 mL or 400 mL) C: Distilled water Single occasions | Randomized, controlled, parallel, PP | 15 Healthy subjects (0 W/15 M) Age: 20–29 y BMI:21.6 kg/m2 In 3 groups, N = 5 | OGTT (50 g glucose) 30 min after MO tea oral dose (finger prick and glucometer) | -- | ↓ glycemia (17% [200 mL] and 19% [400 mL] reduction). Higher reduction at 30 min with lowest amount (22.8 vs. 17.9%) | [45] |
| MO leaf powder (8 g/d) C: No supplementation 40 days | Randomized, controlled | 22 type 2 diabetics (8W/14 M) Age: 40–60 y BMI:18.5–35 kg/m2 | FBG, PPG, lipid profile (methods unspecified) | ↓FBG and PPG ↓ Total cholesterol, LDL and TAG | No statistical test performed | [72] |
| MO leaf tablet (2 units/day) C: No supplementation 90 days | Intervention controlled | 60 type 2 diabetics on sulfonylurea medication (gender unspecified) Age: 40–58 y BMI: 20–25 kg/m2 | HbA1c and PPG two hours after a meal | ↓HbA1 (7.4% reduction) ↓PPG | ↔ HbA1 and PPG | [73] |
| MO leaf powder capsules. Dosages: 0, 1, 2 and 4 g. C: 4 empty capsules. 4 single days separated by 2 wk. | Oral single dose study | 10 Healthy volunteers (5W/5M) Age: 29 ± 5 y BMI: 18.5–23 kg/m2 | Plasma glucose and insulin at intervals during 6 h after single dose of MO. Blood urea nitrogen, creatinine, AST and ALT at the first and forth visit | -- | 4 g: ↑ plasma insulin ↑ insulin AUC and ↑74% AUC of insulin/glucose ratio ↔ Plasma glucose, blood urea nitrogen, creatinine, AST and ALT | [74] |
| MO Material | Animal Model | Organ or Biological Sample | Evidences MO Treated vs. Untreated | Ref. |
|---|---|---|---|---|
| Gene expression of inflammation markers | ||||
| Me MO (FM) ME MO (NFM) | Obese (HFD) | Liver and muscle | ↓IL-6 and TNF-α. ↓IL-1β, only muscle ↓IL-6. ↓ (NS) TNF-α | [54] |
| Et MO | Diabetic | Kidney | ↓TNF-α, IL-1β, IL-6, COX-2 and iNOS | [62] |
| Et MO (seed) | Obese (VHFD) Normal (LFD) | Liver and intestine | ↓iNOS in intestine of VHFD and LFD. ↓iNOS in liver of VHFD. No effect on IL-1β, IL-6 and TNF-α. ↑NQO-1 | [56] |
| Aq MO | Diabetic | skin wound tissues | ↓TNF-α, IL-1β, IL-6, COX-2 and iNOS ↑ VEGF. | [75] |
| Aq MO | Obese (VHFD) | Ileum and liver Adipose tissue | ↓TNF-α. ↓ (NS) IL-6 and IL-1β. ↑ Adiponectin | [28] |
| Inflammatory cytokines | ||||
| Me MO | Diabetic | Liver | ↓TNF- α, IL-6 and MCP-1 | [58] |
| Aq MO | Obese (VHFD) | Plasma | ↓TNF-α and IL-1β | [28] |
| MOP (seeds) | Diabetic | Blood and kidney | ↓IL-6 | [42] |
| Aq MO | Diabetic and normal | PBMC | ↑IFN-γ | [49] |
| Niazirin (seed) | Diabetic | Plasma | ↓TNF- α ↑ IL-10 | [67] |
| Oxidative status | ||||
| Me MO | Diabetic Normal | Liver | ↑(NS) ORAC ↑ORAC | [58] |
| Me MO | Diabetic | Kidney | ↓MDA | [57] |
| Me MO (pods) | Diabetic | Pancreas | ↓MDA | [60] |
| Me MO | Diabetic | Liver (mitochondria) | ↓MDA and ↓protein carbonilation | [61] |
| Aq MO | Diabetic | Serum | ↓MDA | [47] |
| Aq MO | Diabetic | Liver | ↓MDA | [46] |
| MOP (seed) | Diabetic | Serum and kidney | ↓MDA | [42] |
| MO leaf/ MO seed | Diabetic | Brain | ↓MDA | [41] |
| Aq MO | Diabetic | Brain, liver, kidney, pancreas and spleen | ↓MDA | [76] |
| Aq MO | Diabetic | Pancreas | ↓MDA | [50] |
| Aq MO | Diabetic | liver | ↓MDA | [52] |
| MO (protein isolate) | Diabetic | Liver | ↓MDA | [53] |
| Me MO | Diabetic | Heart | ↓MDA, HP and CD | [59] |
| Antioxidant enzyme activity | ||||
| Me MO | Diabetic and Normal | Kidney | ↑CAT (NS in normal) ↑SOD (NS in diabetic) ↓G-Px (NS in diabetic). No effect on GST | [57] |
| Me MO (pods) | Diabetic | Pancreas | ↑GSH, SOD and CAT | [60] |
| Aq MO | Diabetic | Pancreas | ↑GSH | [50] |
| Aq MO | Diabetic | liver | ↑SOD, CAT | [52] |
| Aq MO | Diabetic | Liver | ↑SOD, CAT, GSH | [46] |
| Aq MO | Diabetic | Plasma | ↑TAC | [49] |
| MOP (seed) | Diabetic | Serum and kidney | ↑CAT, SOD and GSH | [42] |
| MO leaf/ MO seed | Diabetic | Brain | ↑CAT, G-Px, GST, GSH | [41] |
| Aq MO | Diabetic | Brain, liver, kidney, pancreas and spleen. Liver and pancreas | ↑CAT, SOD ↑GST | [76] |
| MO (protein isolate) | Diabetic | Liver | ↑CAT No effect on SOD | [53] |
| Me MO | Diabetic | Heart | ↑CAT, SOD, G-Px, GR and GSH | [59] |
| Me MO | Diabetic | Liver (mitochondria) | ↑GSH and GR | [61] |
| Cholinergic dysfunction (associated to cognitive impairment as in diabetes encephalopathy) | ||||
| MO leaf/MO seed | Diabetic | Brain | ↓AChE, BChE and ACE | [41] |
| MO Material | Animal Model | Organ or Biological Sample | Evidences MO Treated vs. Untreated | Ref. |
|---|---|---|---|---|
| Accumulation of lipids in tissues | ||||
| Me MO (FM and NFM) | Obese (HFD) | Liver | ↓ Hepatic adiposity (H&E staining) | [54] |
| Et MO (seed) | Obese (VHFD) | Liver | ↓ Liver lipids (Folch’s method with modifications) | [56] |
| Aq MO | Obese (VHFD) | Liver | ↓ Liver lipids (H&E staining) | [28] |
| Niazirin (seed) | Diabetic | Liver | ↓ Hepatic lipid accumulation (H&E staining) | [67] |
| Circulating lipids | ||||
| Aq MO | Obese (VHFD) | Plasma | ↓Cholesterol | [28] |
| Et MO | Diabetic | Plasma | ↓TG and LDL-C | [62] |
| Aq MO | Diabetic | Serum | ↓TG. No effect on TC | [47] |
| Me MO | Diabetic | Serum | ↓ TG, total and LDL-C and ↑ HDL-C | [55] |
| Niazirin (seed) | Diabetic | Plasma | ↓ LDL-C, TG and NEFA and ↑ HDL-C. ↓TC (high dose only). | [67] |
| Aq MO | Diabetic (2 models: HFD and STZ) | Serum | ↓ TC, TG, VLDL-C, and LDL-C. ↑ HDL-C Smaller level of restoration of lipid profile in STZ- diabetic than in HFD-diabetic | [43] |
| Histopathology and organ functionality | ||||
| Et MO | Diabetic | Kidney | Restored histopathological damage in renal tissue | [62] |
| Aq MO | Diabetic | Serum | ↓ GOT and GPT enzyme. | [43] |
| Aq MO | Diabetic | Pancreas and liver | Prevented histoarchitectural changes | [46,47] |
| MOP (seed) | Diabetic | Kidney and pancreas | Restored the normal histology of kidney and pancreas | [42] |
| Niazirin (seed) | Diabetic | Liver | Restored NAFLD score and hepatocyte structure. | [67] |
| Me MO | Diabetic | Heart | Improved histopathology | [59] |
| Me MO | Diabetic | Liver | Improved histopathology, ↓ GOT and ALP | [58] |
| Me MO | Diabetic | Kidney | Improved histopathology | [57] |
| Me MO (pods) | Diabetic | Pancreas | MO reversed the histoarchitectural damage of islets cells | [66] |
| Gene expression of lipid metabolism and glucose metabolism | ||||
| Aq MO | Obese (VHFD) | Liver | ↓ Lipogenic proteins (FAS, SREBP1 and FSP27) and ↑ lipolytic ATGL. ↓ G6Pase | [28] |
| Fermented MO leaf | Obese (HFD) | Liver Muscle | ↓ ACC, FAS and SREBP-1⇒ Downregulated lipogenic genes. No effect on C/EBPα, PPAR-γ and LPL. ↑ CD36, ACOX1, ATGL, HSL⇒↑ Lipid uptake, oxidation and lipolysis. No effect on CPT1, PPARα. ↑ pAMPK/AMPK. ↓BIP and PDI (muscle) ⇒ ↓ endoplasmic reticulum stress | [54] |
| Aq MO | Diabetic | Liver | Normalized gene expression: ↑ GS, ↓PC and caspase 3 | [46] |
| Niazirin (seed) | Diabetic (db/db) | Liver | ↑ HK and PK enzymes (glycolytic) and ↓ G6Pase and PEPCK (gluconeogenic). ↑ PPAR-α, ↓ SREBP-1 and FAS expression. ↑ ratio P-ACC/ACC. | [67] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nova, E.; Redondo-Useros, N.; Martínez-García, R.M.; Gómez-Martínez, S.; Díaz-Prieto, L.E.; Marcos, A. Potential of Moringa oleifera to Improve Glucose Control for the Prevention of Diabetes and Related Metabolic Alterations: A Systematic Review of Animal and Human Studies. Nutrients 2020, 12, 2050. https://doi.org/10.3390/nu12072050
Nova E, Redondo-Useros N, Martínez-García RM, Gómez-Martínez S, Díaz-Prieto LE, Marcos A. Potential of Moringa oleifera to Improve Glucose Control for the Prevention of Diabetes and Related Metabolic Alterations: A Systematic Review of Animal and Human Studies. Nutrients. 2020; 12(7):2050. https://doi.org/10.3390/nu12072050
Chicago/Turabian StyleNova, Esther, Noemí Redondo-Useros, Rosa M. Martínez-García, Sonia Gómez-Martínez, Ligia E. Díaz-Prieto, and Ascensión Marcos. 2020. "Potential of Moringa oleifera to Improve Glucose Control for the Prevention of Diabetes and Related Metabolic Alterations: A Systematic Review of Animal and Human Studies" Nutrients 12, no. 7: 2050. https://doi.org/10.3390/nu12072050