Vitamin D3 Supplementation Increases Long-Chain Ceramide Levels in Overweight/Obese African Americans: A Post-Hoc Analysis of a Randomized Controlled Trial

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Randomization and Treatments
2.3. Measurements and Laboratory Assessments
2.4. LC-MS/MS Analysis
2.5. Statistical Analysis
3. Results
3.1. General Characteristics
3.2. Effects of Vitamin D3 Supplementation on Sphingolipid Metabolites
3.3. Associations of the Changes in Sphingolipids with the Changes in BMI and A1c
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Hernandez-Corbacho, M.J.; Salama, M.F.; Canals, D.; Senkal, C.E.; Obeid, L.M. Sphingolipids in mitochondria. Biochim. Biophys. Acta 2017, 1862, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Thudichum, J. A Treatise on the Chemical Constitution of the Brain; Bailliere, Tindall and Cox: London, UK, 1962; p. 262. [Google Scholar]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Galadari, A.; Thayyullathil, F. Role of ceramide in diabetes mellitus: Evidence and mechanisms. Lipids Health Dis. 2013, 12, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayyaz, S.; Japtok, L.; Kleuser, B. Divergent role of sphingosine 1-phosphate on insulin resistance. Cell Physiol. Biochem. 2014, 34, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Maceyka, M.; Spiegel, S. Sphingolipid metabolites in inflammatory disease. Nature 2014, 510, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogretmen, B. Sphingolipid metabolism in cancer signalling and therapy. Nature reviews. Cancer 2018, 18, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Stiban, J.; Perera, M. Very long chain ceramides interfere with C16-ceramide-induced channel formation: A plausible mechanism for regulating the initiation of intrinsic apoptosis. Biochim. Biophys. Acta 2015, 1848, 561–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Zhang, N. Ceramide: Therapeutic Potential in Combination Therapy for Cancer Treatment. Curr. Drug Metab. 2015, 17, 37–51. [Google Scholar] [CrossRef]
- Galadari, S.; Rahman, A.; Pallichankandy, S.; Thayyullathil, F. Tumor suppressive functions of ceramide: Evidence and mechanisms. Apoptosis 2015, 20, 689–711. [Google Scholar] [CrossRef]
- Mencarelli, C.; Martinez-Martinez, P. Ceramide function in the brain: When a slight tilt is enough. Cell Mol. Life Sci. 2013, 70, 181–203. [Google Scholar] [CrossRef] [Green Version]
- Upreti, V.; Maitri, V.; Dhull, P.; Handa, A.; Prakash, M.S.; Behl, A. Effect of oral vitamin D supplementation on glycemic control in patients with type 2 diabetes mellitus with coexisting hypovitaminosis D: A parellel group placebo controlled randomized controlled pilot study. Diabetes Metab. Syndr. 2018. [Google Scholar] [CrossRef]
- Li, X.; Liu, Y.; Zheng, Y.; Wang, P.; Zhang, Y. The Effect of Vitamin D Supplementation on Glycemic Control in Type 2 Diabetes Patients: A Systematic Review and Meta-Analysis. Nutrients 2018, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Chavez, J.A.; Siddique, M.M.; Wang, S.T.; Ching, J.; Shayman, J.A.; Summers, S.A. Ceramides and glucosylceramides are independent antagonists of insulin signaling. J. Biol. Chem. 2014, 289, 723–734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cinar, R.; Godlewski, G.; Liu, J.; Tam, J.; Jourdan, T.; Mukhopadhyay, B.; Harvey-White, J.; Kunos, G. Hepatic cannabinoid-1 receptors mediate diet-induced insulin resistance by increasing de novo synthesis of long-chain ceramides. Hepatology 2014, 59, 143–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, A.; Grammatikos, G.; Trautmann, S.; Schreiber, Y.; Thomas, D.; Bruns, F.; Pfeilschifter, J.; Badenhoop, K.; Penna-Martinez, M. Vitamin D Supplementation Enhances C18(dihydro)ceramide Levels in Type 2 Diabetes Patients. Int. J. Mol. Sci. 2017, 18, 1532. [Google Scholar] [CrossRef]
- Turpin, S.M.; Nicholls, H.T.; Willmes, D.M.; Mourier, A.; Brodesser, S.; Wunderlich, C.M.; Mauer, J.; Xu, E.; Hammerschmidt, P.; Bronneke, H.S.; et al. Obesity-induced CerS6-dependent C16:0 ceramide production promotes weight gain and glucose intolerance. Cell Metab. 2014, 20, 678–686. [Google Scholar] [CrossRef] [Green Version]
- Raichur, S.; Wang, S.T.; Chan, P.W.; Li, Y.; Ching, J.; Chaurasia, B.; Dogra, S.; Ohman, M.K.; Takeda, K.; Sugii, S.; et al. CerS2 haploinsufficiency inhibits beta-oxidation and confers susceptibility to diet-induced steatohepatitis and insulin resistance. Cell Metab. 2014, 20, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Bergman, B.C.; Brozinick, J.T.; Strauss, A.; Bacon, S.; Kerege, A.; Bui, H.H.; Sanders, P.; Siddall, P.; Kuo, M.S.; Perreault, L. Serum sphingolipids: Relationships to insulin sensitivity and changes with exercise in humans. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E398–E408. [Google Scholar] [CrossRef] [Green Version]
- Kasumov, T.; Solomon, T.P.; Hwang, C.; Huang, H.; Haus, J.M.; Zhang, R.; Kirwan, J.P. Improved insulin sensitivity after exercise training is linked to reduced plasma C14:0 ceramide in obesity and type 2 diabetes. Obesity 2015, 23, 1414–1421. [Google Scholar] [CrossRef]
- Oda, Y.; Uchida, Y.; Moradian, S.; Crumrine, D.; Elias, P.M.; Bikle, D.D. Vitamin D receptor and coactivators SRC2 and 3 regulate epidermis-specific sphingolipid production and permeability barrier formation. J. Investig. Dermatol. 2009, 129, 1367–1378. [Google Scholar] [CrossRef] [Green Version]
- Bhagatwala, J.; Zhu, H.; Parikh, S.J.; Guo, D.H.; Kotak, I.; Huang, Y.; Havens, R.; Pham, M.; Afari, E.; Kim, S.; et al. Dose and time responses of vitamin D biomarkers to monthly vitamin D3 supplementation in overweight/obese African Americans with suboptimal vitamin d status: A placebo controlled randomized clinical trial. BMC Obes. 2015, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Vanni, N.; Fruscione, F.; Ferlazzo, E.; Striano, P.; Robbiano, A.; Traverso, M.; Sander, T.; Falace, A.; Gazzerro, E.; Bramanti, P.; et al. Impairment of ceramide synthesis causes a novel progressive myoclonus epilepsy. Ann. Neurol. 2014, 76, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Taki, T. An approach to glycobiology from glycolipidomics: Ganglioside molecular scanning in the brains of patients with Alzheimer’s disease by TLC-blot/matrix assisted laser desorption/ionization-time of flight MS. Biol. Pharm. Bull. 2012, 35, 1642–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, J.M.; Gupta, S. Vitamin D and neurocognitive disorder due to Alzheimer’s disease: A review of the literature. Annals of clinical psychiatry. Off. J. Am. Acad. Clin. Psychiatr. 2015, 27, 206–212. [Google Scholar]
- Gezen-Ak, D.; Yilmazer, S.; Dursun, E. Why vitamin D in Alzheimer’s disease? The hypothesis. J. Alzheimers Dis. 2014, 40, 257–269. [Google Scholar] [CrossRef] [PubMed]
- Karakis, I.; Pase, M.P.; Beiser, A.; Booth, S.L.; Jacques, P.F.; Rogers, G.; DeCarli, C.; Vasan, R.S.; Wang, T.J.; Himali, J.J.; et al. Association of Serum Vitamin D with the Risk of Incident Dementia and Subclinical Indices of Brain Aging: The Framingham Heart Study. J. Alzheimers Dis. 2016, 51, 451–461. [Google Scholar] [CrossRef] [Green Version]
- Lerner, P.P.; Sharony, L.; Miodownik, C. Association between mental disorders, cognitive disturbances and vitamin D serum level: Current state. Clin. Nutr. ESPEN 2018, 23, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, M.; Jiang, X.C. Sphingomyelin and its role in cellular signaling. Adv. Exp. Med. Biol. 2013, 991, 1–14. [Google Scholar] [CrossRef]
- Ramprasath, V.R.; Jones, P.J.; Buckley, D.D.; Woollett, L.A.; Heubi, J.E. Effect of dietary sphingomyelin on absorption and fractional synthetic rate of cholesterol and serum lipid profile in humans. Lipids Health Dis. 2013, 12, 125. [Google Scholar] [CrossRef] [Green Version]
- Vesper, H.; Schmelz, E.M.; Nikolova-Karakashian, M.N.; Dillehay, D.L.; Lynch, D.V.; Merrill, A.H., Jr. Sphingolipids in food and the emerging importance of sphingolipids to nutrition. J. Nutr. 1999, 129, 1239–1250. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hosozawa, M.; Kudo, N.; Yoshikawa, N.; Hisata, K.; Shoji, H.; Shinohara, K.; Shimizu, T. The pilot study: Sphingomyelin-fortified milk has a positive association with the neurobehavioural development of very low birth weight infants during infancy, randomized control trial. Brain Dev. 2013, 35, 45–52. [Google Scholar] [CrossRef]
- Magrassi, L.; Adorni, L.; Montorfano, G.; Rapelli, S.; Butti, G.; Berra, B.; Milanesi, G. Vitamin D metabolites activate the sphingomyelin pathway and induce death of glioblastoma cells. Acta Neurochir. 1998, 140, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Bektas, M.; Orfanos, C.E.; Geilen, C.C. Different vitamin D analogues induce sphingomyelin hydrolysis and apoptosis in the human keratinocyte cell line HaCaT. Cell Mol. Biol. 2000, 46, 111–119. [Google Scholar] [PubMed]
- Liu, R.; Xu, Y.; Farach-Carson, M.C.; Vogel, J.J.; Karin, N.J. 1,25 dihydroxyvitamin D (3) activates sphingomyelin turnover in ROS17/2.8 osteosarcoma cells without sphingolipid-induced changes in cytosolic Ca (2+). Biochem. Biophys. Res. Commun. 2000, 273, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Pollock, N.; Stallmann-Jorgensen, I.S.; Gutin, B.; Lan, L.; Chen, T.C.; Keeton, D.; Petty, K.; Holick, M.F.; Zhu, H. Low 25-hydroxyvitamin D levels in adolescents: Race, season, adiposity, physical activity, and fitness. Pediatrics 2010, 125, 1104–1111. [Google Scholar] [CrossRef] [Green Version]
- Belenchia, A.M.; Tosh, A.K.; Hillman, L.S.; Peterson, C.A. Correcting vitamin D insufficiency improves insulin sensitivity in obese adolescents: A randomized controlled trial. Am. J. Clin. Nutr. 2013, 97, 774–781. [Google Scholar] [CrossRef] [Green Version]


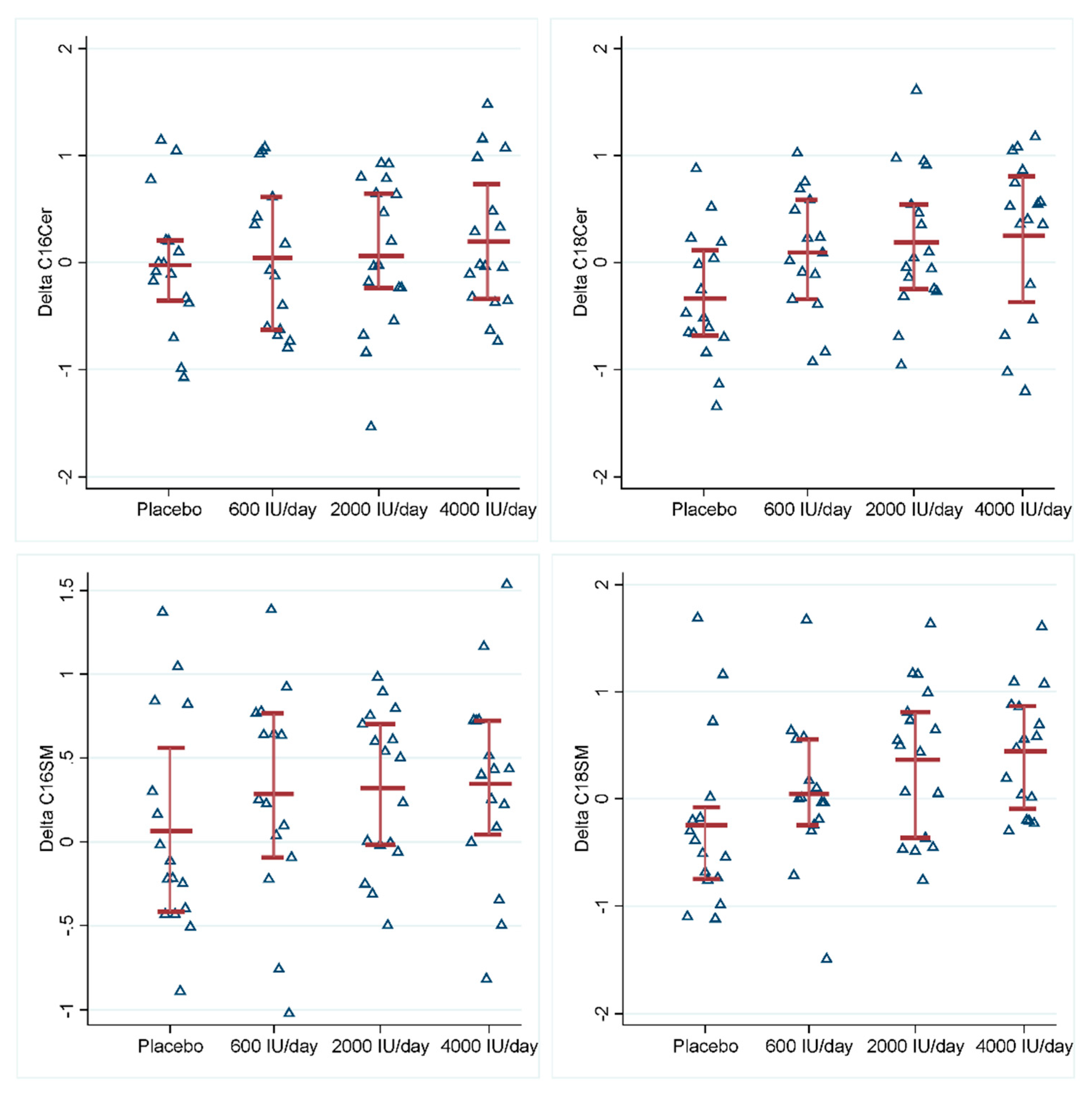{kind=link}
| Characteristics | Total | Groups | ||||
|---|---|---|---|---|---|---|
| Placebo (N = 16) | 600 IU/day (N = 15) | 2000 IU/day (N = 17) | 4000 IU/day (N = 16) | p-Values | ||
| Age (year) | 26.0 ± 9.4 | 27.9 ± 10.4 | 26.3 ± 9.8 | 24.5 ± 8.5 | 25.5 ± 9.6 | 0.833 |
| Male (N) | 11 (17) | 4 (25) | 2 (13) | 3 (18) | 2 (13) | 0.843 |
| BMI (kg/m2) | 35.7 ± 7.2 | 36.2 ± 8.1 | 34.6 ± 5.6 | 37.1 ± 7.7 | 34.8 ± 7.3 | 0.636 |
| Obese (N) | 51 (80) | 12 (75) | 12 (80) | 14 (82) | 13 (81) | 0.974 |
| A1c (%) | 5.4 ± 0.4 | 5.4 ± 0.5 | 5.4 ± 0.5 | 5.3 ± 0.5 | 5.5 ± 0.3 | 0.422 |
| 25(OH)D (nmol/L) | 36.9 ± 11.3 | 39.7 ± 14.7 | 35.0 ± 7.7 | 39.8 ± 10.7 | 32.9 ± 10.3 | 0.276 |
| Metabolites. | 600 IU/day | 2000 IU/day | 4000 IU/day | |||
|---|---|---|---|---|---|---|
| β (95% CI) | p | β (95% CI) | p | β (95% CI) | p | |
| C16Cer | 0.06 (−0.40, 0.53) | 0.791 | 0.09 (−0.36, 0.54) | 0.702 | 0.22 (−0.23, 0.54) | 0.336 |
| C18Cer | 0.44 (0.00, 0.87) | 0.049 | 0.52 (0.10, 0.94) | 0.016 | 0.58 (0.15, 1.00) | 0.008 |
| C16dhCer | −0.06 (−0.57, 0.46) | 0.830 | −0.22 (−0.71, 0.28) | 0.369 | −0.00 (−0.51, 0.50) | 0.988 |
| C18dhCer | 0.28 (−0.33, 0.90) | 0.363 | 0.54 (−0.05, 1.15) | 0.071 | 0.39 (−0.21, 1.00) | 0.200 |
| Sphingosine | −0.09 (−0.88, 0.71) | 0.833 | −0.27 (−1.03, 0.50) | 0.499 | −0.35 (−1.13, 0.43) | 0.382 |
| S1P | −0.17 (−0.86, 0.53) | 0.638 | 0.26 (−0.42, 0.93) | 0.460 | −0.42 (−1.10, 0.27) | 0.232 |
| C16SM | 0.21 (−0.19, 0.62) | 0.299 | 0.26 (−0.13, 0.65) | 0.198 | 0.28 (−0.11, 0.68) | 0.160 |
| C18SM | 0.30 (−0.18, 0.76) | 0.222 | 0.61 (0.15, 1.06) | 0.009 | 0.68 (0.22, 1.15) | 0.004 |
| Metabolites | 25(OH)D | |
|---|---|---|
| β (95% CI) | p | |
| C16Cer | 0.13 (−0.31, 0.57) | 0.570 |
| C18Cer | 0.44 (0.02, 0.86) | 0.041 |
| C16dhCer | −0.01 (−0.51, 0.50) | 0.973 |
| C18dhCer | 0.39 (−0.16, 0.93) | 0.161 |
| Sphingosine | 0.08 (−0.68, 0.83) | 0.841 |
| S1P | −0.10 (−0.77, 0.57) | 0.758 |
| C16SM | 0.25 (−0.12, 0.61) | 0.179 |
| C18SM | 0.47 (0.05, 0.90) | 0.030 |
| Metabolites | BMI | A1c | ||
|---|---|---|---|---|
| β | p | β | p | |
| C16Cer | 0.65 | 0.068 | −0.04 | 0.419 |
| C18Cer | 0.97 | 0.006 | 0.05 | 0.302 |
| C16dhCer | 0.57 | 0.076 | 0.05 | 0.284 |
| C18dhCer | 0.58 | 0.033 | 0.11 | 0.003 |
| Sphingosine | 0.01 | 0.953 | 0.08 | 0.037 |
| S1P | −0.04 | 0.882 | 0.09 | 0.020 |
| C16SM | 0.31 | 0.414 | −0.02 | 0.594 |
| C18SM | 0.52 | 0.116 | 0.03 | 0.468 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, L.; Dong, Y.; Bhagatwala, J.; Raed, A.; Huang, Y.; Zhu, H. Vitamin D3 Supplementation Increases Long-Chain Ceramide Levels in Overweight/Obese African Americans: A Post-Hoc Analysis of a Randomized Controlled Trial. Nutrients 2020, 12, 981. https://doi.org/10.3390/nu12040981
Chen L, Dong Y, Bhagatwala J, Raed A, Huang Y, Zhu H. Vitamin D3 Supplementation Increases Long-Chain Ceramide Levels in Overweight/Obese African Americans: A Post-Hoc Analysis of a Randomized Controlled Trial. Nutrients. 2020; 12(4):981. https://doi.org/10.3390/nu12040981
Chicago/Turabian StyleChen, Li, Yanbin Dong, Jigar Bhagatwala, Anas Raed, Ying Huang, and Haidong Zhu. 2020. "Vitamin D3 Supplementation Increases Long-Chain Ceramide Levels in Overweight/Obese African Americans: A Post-Hoc Analysis of a Randomized Controlled Trial" Nutrients 12, no. 4: 981. https://doi.org/10.3390/nu12040981