Steamed Ginger May Enhance Insulin Secretion through KATP Channel Closure in Pancreatic β-Cells Potentially by Increasing 1-Dehydro-6-Gingerdione Content

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. Ethics Statement
2.4. Extraction of Steamed Ginger Extract (GG03)
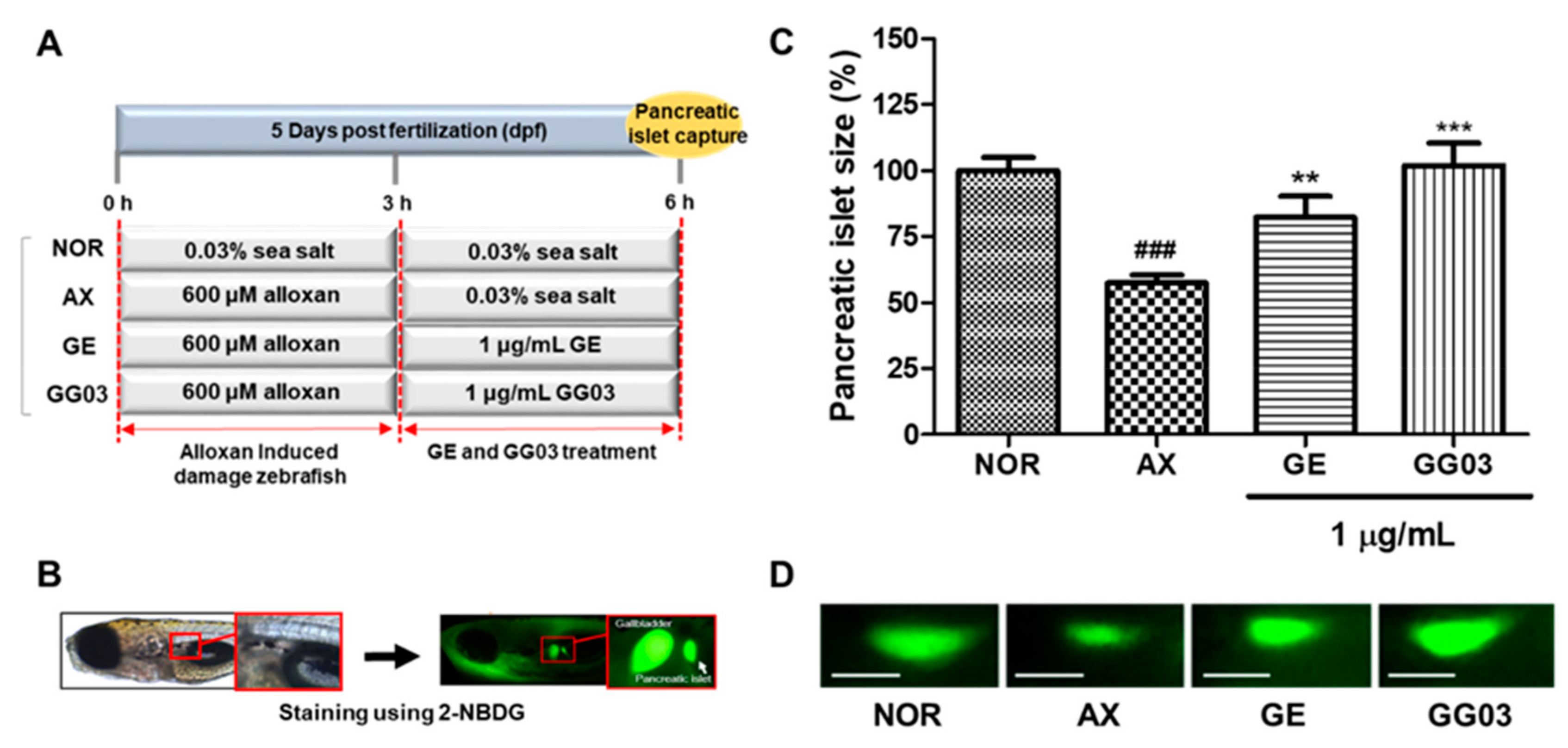
2.5. GE, GG03 and GD Efficacy in Treating Alloxan-Induced Pancreatic Islet Damage in Zebrafish
2.6. The 50% Effective Concentration (EC50) of GE and GG03
2.7. The 50% Lethal Concentration (LC50) Values of GE and GG03
2.8. Therapeutic Index (TI)
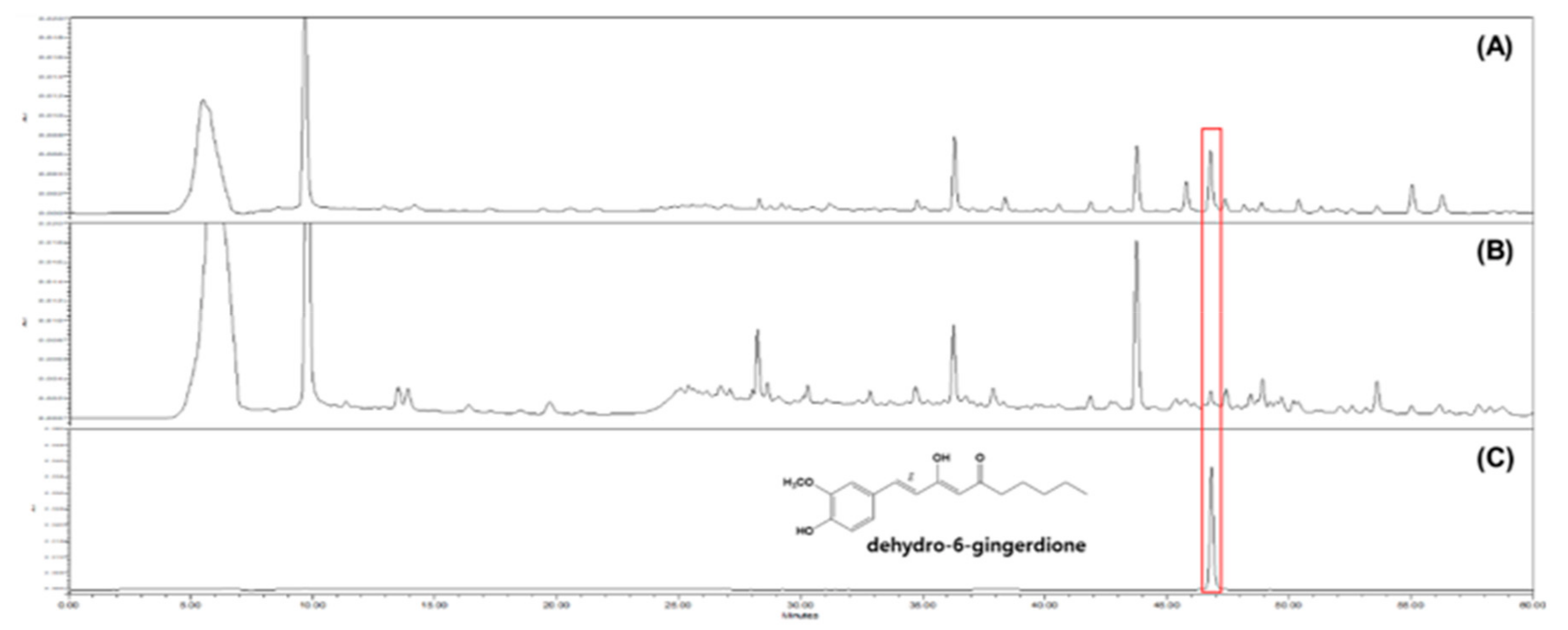
2.9. Quantitative Analysis of GD in GE and GG03
2.10. Protein Tyrosine Phosphatase 1B (PTP1B) Inhibitory Activity
2.11. Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Activity
2.12. α-Glucosidase Inhibitory Activity
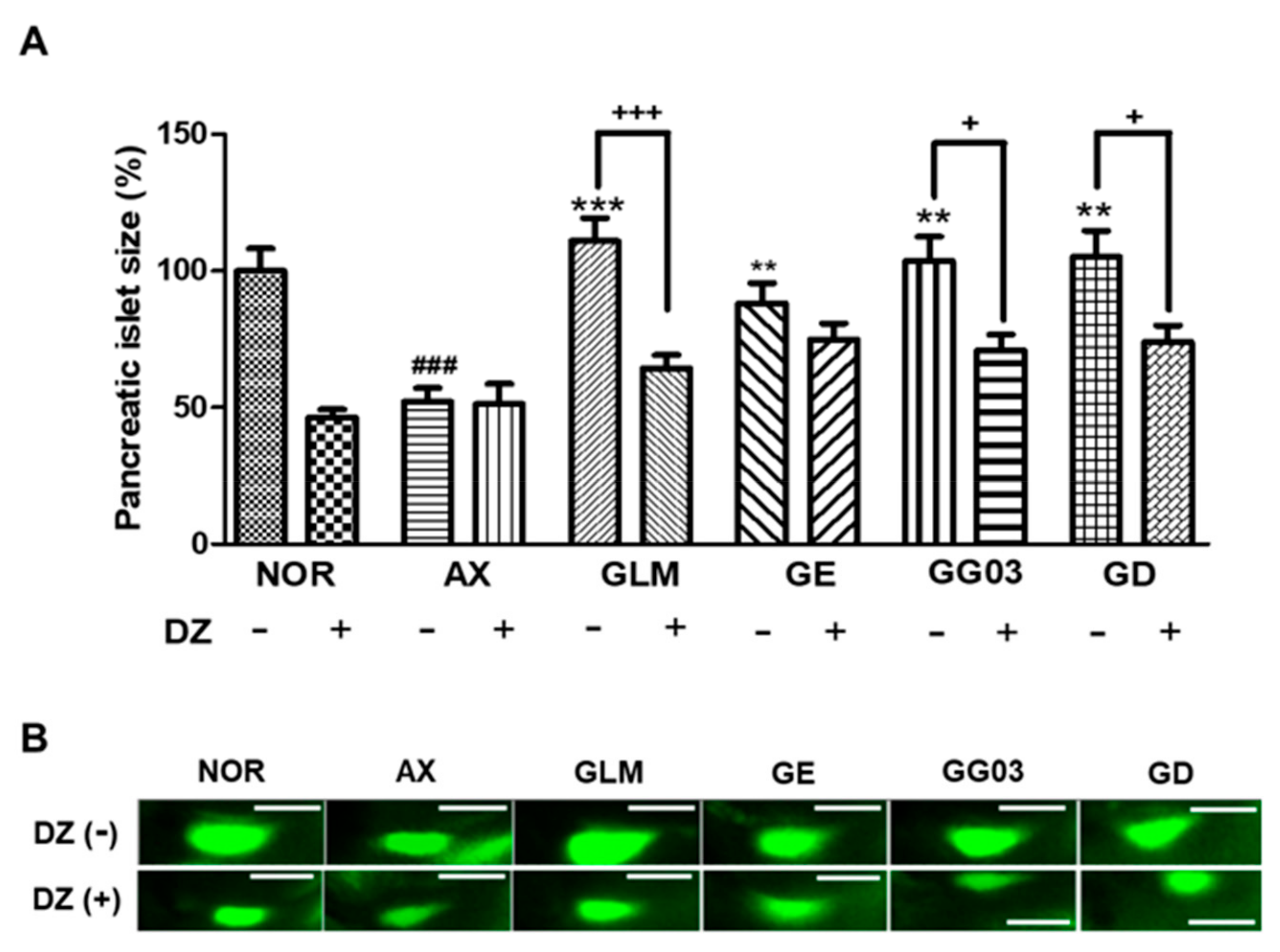
2.13. Action of diazoxide on Alloxan-Induced Diabetic Zebrafish
2.14. Induction of DM
2.15. Treatment with GG03
2.16. Blood Glucose Level Measurement
2.17. Measurement of Biomarkers
2.18. Statistical Analysis
3. Results
3.1. Efficacy of GE and GG03 in Alloxan-Induced Diabetic Zebrafish
3.2. EC50 values of GE and GG03
3.3. LC50 values of GE and GG03
3.4. Therapeutic Index (TI)
3.5. HPLC Analysis of GE and GG03
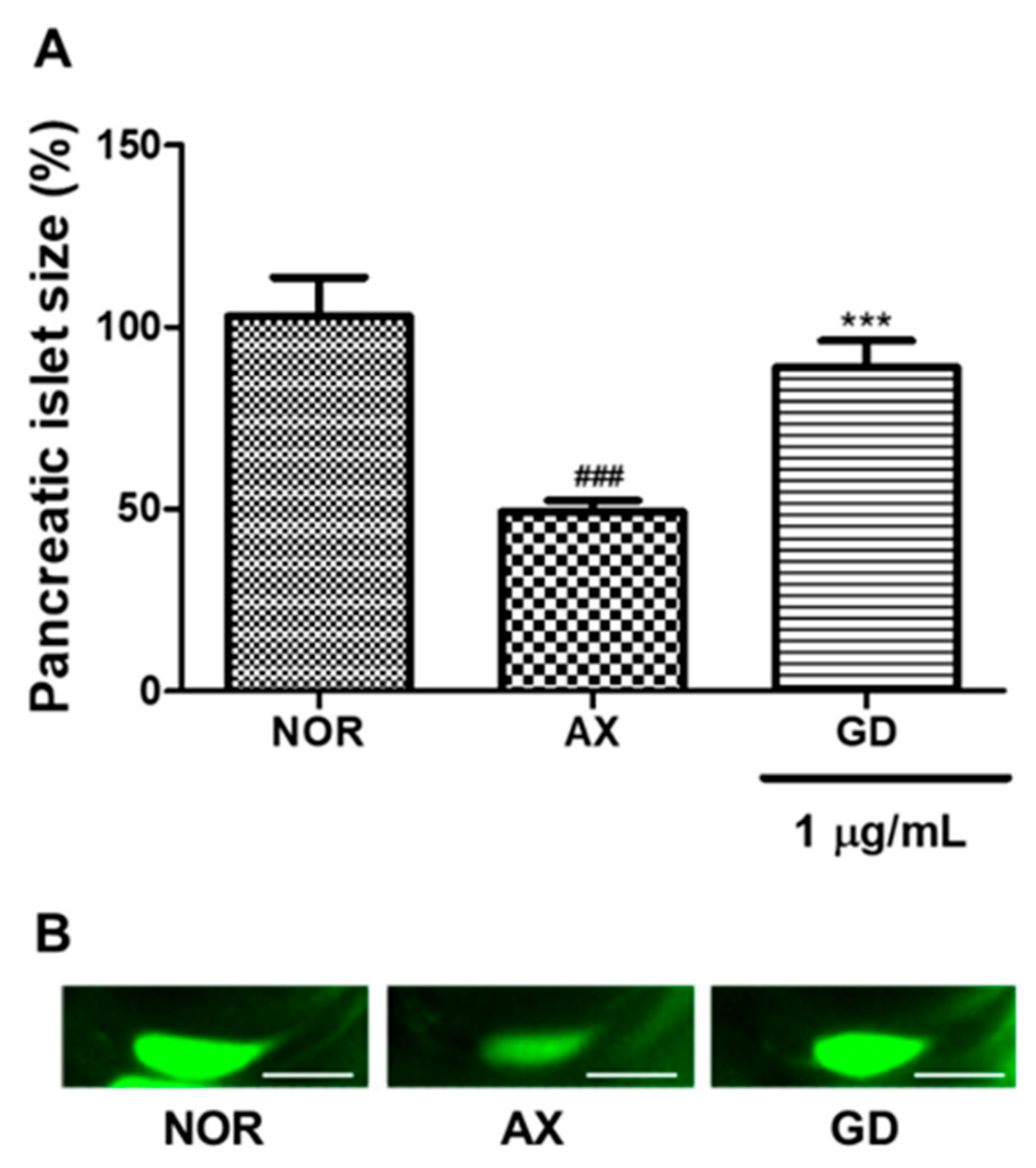
3.6. Efficacy of GD in Alloxan-Induced Diabetic Zebrafish
3.7. Mechanism of Anti-Diabetes Action
3.8. Action on Insulin Secretion Through KATP Channels Modulation
3.9. Body Weight and Blood Glucose Level
3.10. Pancreas Weight
3.11. Measurement of Biomarkers
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Attokaran, M. Ginger: Zingiber officinale R (Zingiberaceae). Nat. Food Flavors Color. 2017, 57, 209–214. [Google Scholar] [CrossRef]
- Grzanna, R.; Lindmark, L.; Frondoza, C.G. Ginger—an herbal medicinal product with broad anti-inflammatory actions. J. Med. Food. 2005, 8, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.-S.; Lee, W.-C.; Lin, Y.-E.; Ho, C.T.; Lu, K.H.; Lin, S.H.; Panyod, S.; Chu, Y.L.; Sheen, L.Y. Ginger Essential Oil Ameliorates Hepatic Injury and Lipid Accumulation in High Fat Diet-Induced Nonalcoholic Fatty Liver Disease. J. Agr. Food. Chem. 2016, 64, 2062–2071. [Google Scholar] [CrossRef] [PubMed]
- Daily, W.; Zhang, X.; Kim, D.S.; Park, S. Efficacy of ginger for alleviating the symptoms of primary dysmenorrhea: A systematic review and meta—analysis of randomized clinical trials. Pain Med. 2015, 16, 2243–2255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bryer, E. A literature review of the effectiveness of ginger in alleviating mild-to-moderate nausea and vomiting of pregnancy. JMWH. 2005, 50, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Panahi, Y.; Saadat, A.; Sahebkar, A.; Hashemian, F.; Taghikhani, M.; Abolhasani, E. Effect of ginger on acute and delayed chemotherapy-induced nausea and vomiting: A pilot, randomized, open-label clinical trial. Integr. Cancer Ther. 2012, 11, 204–211. [Google Scholar] [CrossRef]
- Samad, M.B.; Mohsin, M.N.A.B.; Razu, B.A.; Hossain, M.T.; Mahzabeen, S.; Unnoor, N.; Muna, I.A.; Akhter, F.; Kabir, A.U.; Hannan, J.M.A. [6]-Gingerol, from Zingiber officinale, potentiates GLP-1 mediated glucose-stimulated insulin secretion pathway in pancreatic β-cells and increases RAB8/RAB10-regulated membrane presentation of GLUT4 transporters in skeletal muscle to improve hyperglycemia in Lepr db/db type 2 diabetic mice. BMC Complement. Altern. Med. 2017, 17, 395. [Google Scholar]
- Wei, C.K.; Tsai, Y.H.; Korinek, M.; Hung, P.H.; El-Shazly, M.; Cheng, Y.B.; Wu, Y.C.; Hsieh, T.J.; Chang, F.R. 6-paradol and 6-shogaol, the pungent compounds of ginger, promote glucose utilization in adipocytes and myotubes, and 6-paradol reduces blood glucose in high-fat diet-fed mice. Int. J. Mol. Sci. 2017, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Chen, H.; Song, Z.; Wang, X.; Sun, Z. Effects of ginger (Zingiber officinale Roscoe) on type 2 diabetes mellitus and components of the metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Evid. Based Complement. Alternat. Med. 2018, 2018, 5692962. [Google Scholar] [CrossRef] [Green Version]
- Yi, J.K.; Ryoo, Z.Y.; Ha, J.J.; Oh, D.Y.; Kim, M.O.; Kim, S.H. Beneficial effects of 6-shogaol on hyperglycemia, islet morphology and apoptosis in some tissues of streptozotocin-induced diabetic mice. Diabetol. Metab. Syndr. 2019, 11, 15. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.L.; Liu, Q.; Peng, Y.B.; Qi, L.W.; Li, P. Steamed ginger (Zingiber officinale): Changed chemical profile and increased anticancer potential. Food Chem. 2011, 129, 1785–1792. [Google Scholar] [CrossRef]
- Cavaghan, M.K.; Ehrmann, D.A.; Polonsky, K.S. Interactions between insulin resistance and insulin secretion in the development of glucose intolerance. J. Clin. Invest. 2000, 106, 329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Belle, T.L.; Coppieters, K.T.; Von Herrath, M.G. Type 1 diabetes: Etiology, immunology, and therapeutic strategies. Phys. Rev. 2011, 91, 79–118. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Hebrok, M. Intercellular signals regulating pancreas development and function. Genes Dev. 2001, 15, 111–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rorsman, P.; Renström, E. Insulin granule dynamics in pancreatic beta cells. Diabetologia 2003, 46, 1029–1045. [Google Scholar] [CrossRef]
- Rhodes, C.J.; White, M.F. Molecular insights into insulin action and secretion. Eur. J. Clin. Invest. 2002, 32, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Hill, D.E.; Chernoff, J. Direct binding of the proline-rich region of protein tyrosine phosphatase 1B to the Src homology 3 domain of p130(Cas). J. Biol. Chem. 1996, 271, 31290–31295. [Google Scholar] [CrossRef] [Green Version]
- Al-Masri, I.M.; Mohammad, M.K.; Tahaa, M.O. Inhibition of dipeptidyl peptidase IV (DPP IV) is one of the mechanisms explaining the hypoglycemic effect of berberine. J. Enzyme Inhib. Med. Chem. 2009, 24, 1061–1066. [Google Scholar] [CrossRef]
- Dewi, R.T.; Iskandar, Y.M.; Hanafi, M.; Kardono, L.B.S.; Angelina, M.; Dewijanti, I.D.; Banjarnahor, S.D. Inhibitory effect of Koji Aspergillus terreus on a-glucosidase activity and postprandial hyperglycemia. Pak. J. Biol. Sci. 2007, 10, 3131–3135. [Google Scholar]
- Song, C.H.; Seo, Y.C.; Choi, W.Y.; Lee, C.G.; Kim, D.U.; Chung, J.Y.; Chung, H.C.; Park, D.S.; Ma, C.J.; Lee, H.Y. Enhancement of antioxidative activity of Codonopsis lanceolata by stepwise steaming process. Korean J. Crop Sci. 2012, 20, 238–244. [Google Scholar] [CrossRef]
- Desgraz, R.; Bonal, C.; Herrera, P.L. β-Cell regeneration: The pancreatic intrinsic faculty. Trends Endocrinol. Metab. 2011, 22, 34–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nam, Y.H.; Hong, B.N.; Rodriguez, I.; Ji, M.G.; Kim, K.; Kim, U.J.; Kang, T.H. Synergistic potentials of coffee on injured pancreatic islets and insulin action via KATP channel-blocking in zebrafish. J. Agric. Food Chem. 2015, 63, 5612–5621. [Google Scholar] [CrossRef] [PubMed]
- Muller, P.Y.; Milton, M.N. The determination and interpretation of the therapeutic index in drug development. Nat. Rev. Drug Discov. 2012, 11, 751–761. [Google Scholar] [CrossRef] [PubMed]
- Tamrakar, A.K.; Maurya, C.K.; Rai, A.K. PTP1B inhibitors for type 2 diabetes treatment: A patent review (2011–2014). Expert Opin. Ther. Pat. 2014, 24, 1101–1115. [Google Scholar] [CrossRef]
- Thornberry, N.A.; Gallwitz, B. Mechanism of action of inhibitors of dipeptidyl-peptidase-4 (DPP-4). Best Pract. Res. Clin. Endocrinol. Metab. 2009, 23, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Moelands, S.V.; Lucassen, P.L.; Akkermans, R.P.; De Grauw, W.J.; Van de Laar, F.A. Alpha-glucosidase inhibitors for prevention or delay of type 2 diabetes mellitus and its associated complications in people at increased risk of developing type 2 diabetes mellitus. Cochrane Database Syst. Rev. 2018, 12, CD005061. [Google Scholar] [CrossRef]
- Ashcroft, F.M. ATP-sensitive K+ channels and disease: From molecule to malady. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E880–E889. [Google Scholar] [CrossRef] [Green Version]
- Henquin, J.C. Triggering and amplifying pathways of regulation of insulin secretion by glucose. Diabetes 2000, 49, 1751–1760. [Google Scholar] [CrossRef] [Green Version]
- Tabachnick, I.I.; Gulbenkian, A.; Seidman, F. The effect of a benzothiadiazine, diazoxide, on carbohydrate metabolism. Diabetes 1964, 13, 408–418. [Google Scholar] [CrossRef]
- Nam, Y.H.; Le, H.T.; Rodriguez, I.; Kim, E.Y.; Kim, K.; Jeong, S.Y.; Woo, S.H.; Lee, Y.R.; Castañeda, R.; Hong, J.; et al. Enhanced antidiabetic efficacy and safety of compound K⁄ β-cyclodextrin inclusion complex in zebrafish. J. Ginseng. Res. 2017, 41, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Rani, M.P.; Krishna, M.S.; Padmakumari, K.P.; Raghu, K.G.; Sundaresan, A. Zingiber officinale extract exhibits antidiabetic potential via modulating glucose uptake, protein glycation and inhibiting adipocyte differentiation: An in vitro study. J. Sci. Food Agr. 2012, 92, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.O.; Kim, N.; Lee, H.J.; Moon, J.W.; Lee, S.K.; Kim, S.J.; Kim, J.K.; Park, S.H.; Kim, H.S. [6]–Gingerol Affects Glucose Metabolism by Dual Regulation via the AMPKα2–Mediated AS160–Rab5 Pathway and AMPK–Mediated Insulin Sensitizing Effects. J. Cell Biochem. 2015, 116, 1401–1410. [Google Scholar] [CrossRef] [PubMed]
- Al-Amin, Z.M.; Thomson, M.; Al-Qattan, K.K.; Peltonen-Shalaby, R.; Ali, M. Anti-diabetic and hypolipidaemic properties of ginger (Zingiber officinale) in streptozotocin-induced diabetic rats. Br. J. Nutr. 2006, 96, 660–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al Hroob, A.M.; Abukhalil, M.H.; Alghonmeen, R.D.; Mahmoud, A.M. Ginger alleviates hyperglycemia-induced oxidative stress, inflammation and apoptosis and protects rats against diabetic nephropathy. Biomed Pharmacother. 2018, 106, 381–389. [Google Scholar] [CrossRef]
- Abdulrazaq, N.B.; Cho, M.M.; Win, N.N.; Zaman, R.; Rahman, M.T. Beneficial effects of ginger (Zingiber officinale) on carbohydrate metabolism in streptozotocin-induced diabetic rats. Br. J. Nutr. 2012, 108, 1194–1201. [Google Scholar] [CrossRef] [Green Version]
- Ojewole, J.A. Analgesic, antiinflammatory and hypoglycaemic effects of ethanol extract of Zingiber officinale (Roscoe) rhizomes (Zingiberaceae) in mice and rats. Phytother. Res. 2006, 20, 764–772. [Google Scholar] [CrossRef]
- Bhandari, U.; Kanojia, R.; Pillai, K.K. Effect of ethanolic extract of Zingiber officinale on dyslipidaemia in diabetic rats. J. Ethnopharmacol. 2005, 97, 227–230. [Google Scholar] [CrossRef]
- Shimizu, R.; Sakazaki, F.; Okuno, T.; Nakamuro, K.; Ueno, H. Difference in glucose intolerance between C57BL/6J and ICR strain mice with streptozotocin/nicotinamide-induced diabetes. Biomed. Res. 2012, 33, 63–66. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, K.; Kojima, R.; Ito, M. Strain differences in the diabetogenic activity of streptozotocin in mice. Biol. Pharm. Bull. 2006, 29, 1110–1119. [Google Scholar] [CrossRef] [Green Version]
- Fajardo, R.J.; Karim, L.; Calley, V.I.; Bouxsein, M.L. A review of rodent models of type 2 diabetic skeletal fragility. J. Bone Miner. Res. 2014, 29, 1025–1240. [Google Scholar] [CrossRef] [Green Version]
- Leung, W.; Ho, F.M.; Li, W.P.; Liang, Y.C. Vitis thunbergii var. Taiwaniana leaf extract reduces blood glucose levels in mice with streptozotocin-induced diabetes. Int. J. Pharmacol. 2017, 13, 457–464. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.Y.; Kim, E.; Seo, K.; Kang, Y.J.; Kim, J.Y.; Kim, C.H.; Song, H.T.; Saksida, L.M.; Lee, J.E. Assessment of cognitive impairment in a mouse model of high-fat diet-induced metabolic stress with touchscreen-based automated battery system. Exp. Neurobiol. 2018, 27, 277–286. [Google Scholar] [CrossRef]
- Ito, M.; Kondo, Y.; Nakatani, A.; Hayashi, K.; Naruse, A. Characterization of low dose streptozotocin-induced progressive diabetes in mice. Environ. Toxicol. Pharmacol. 2001, 9, 71–78. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | IC50 (μg/mL) |
|---|---|
| Suramin | 12.3 |
| GE | ND |
| GG03 | ND |
| GD | ND |
| Sample | IC50 (μg/mL) |
|---|---|
| Diprotin A | 17.5 |
| GE | ND |
| GG03 | ND |
| GD | ND |
| Sample | IC50 (μg/mL) |
|---|---|
| Acarbose | 1302.8 |
| GE | ND |
| GG03 | ND |
| GD | ND |
| Groups | NOR | DM | GG03 |
|---|---|---|---|
| Albumin (g/dL) | 3.26 ± 0.09 | 2.73 ± 0.35 ++ | 3.13 ± 0.5 |
| Total proteins (g/dL) | 4.90 ± 0.19 | 4.61 ± 0.24 + | 4.91 ± 0.31 |
| Lactate dehydrogenase (U/L) | 266.40 ± 118.03 | 270.00 ± 130.66 | 325.14 ± 140.75 |
| Creatinine (g/dL) | 0.21 ± 0.04 | 0.25 ± 0.02 + | 0.28 ± 0.03 |
| Blood urea nitrogen (mg/dL) | 17.78 ± 3.67 | 17.18 ± 2.13 | 14.84 ± 2.73 |
| Groups | NOR | DM | GG03 |
|---|---|---|---|
| Glucose (mg/dL) | 240.80 ± 25.84 | 612.50 ± 107.64 +++ | 363.71 ± 141.09 *** |
| HDL (mg/dL) | 98.00 ± 13.19 | 78.50 ± 26.81 | 92.00 ± 23.61 |
| LDL (mg/dL) | 10.40 ± 3.05 | 10.75 ± 6.39 | 13.57 ± 3.41 |
| Total cholesterol (mg/dL) | 104.60 ± 16.38 | 91.50 ± 29.64 | 103.29 ± 23.10 |
| TG (mg/dL) | 106.40 ± 41.55 | 207.00 ± 68.36 | 156.86 ± 39.69 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, Y.H.; Hong, B.N.; Rodriguez, I.; Park, M.S.; Jeong, S.Y.; Lee, Y.-G.; Shim, J.H.; Yasmin, T.; Kim, N.W.; Koo, Y.T.; et al. Steamed Ginger May Enhance Insulin Secretion through KATP Channel Closure in Pancreatic β-Cells Potentially by Increasing 1-Dehydro-6-Gingerdione Content. Nutrients 2020, 12, 324. https://doi.org/10.3390/nu12020324
Nam YH, Hong BN, Rodriguez I, Park MS, Jeong SY, Lee Y-G, Shim JH, Yasmin T, Kim NW, Koo YT, et al. Steamed Ginger May Enhance Insulin Secretion through KATP Channel Closure in Pancreatic β-Cells Potentially by Increasing 1-Dehydro-6-Gingerdione Content. Nutrients. 2020; 12(2):324. https://doi.org/10.3390/nu12020324
Chicago/Turabian StyleNam, Youn Hee, Bin Na Hong, Isabel Rodriguez, Min Seon Park, Seo Yule Jeong, Yeong-Geun Lee, Ji Heon Shim, Tamanna Yasmin, Na Woo Kim, Young Tae Koo, and et al. 2020. "Steamed Ginger May Enhance Insulin Secretion through KATP Channel Closure in Pancreatic β-Cells Potentially by Increasing 1-Dehydro-6-Gingerdione Content" Nutrients 12, no. 2: 324. https://doi.org/10.3390/nu12020324