Reduced Salivary Mucin Binding and Glycosylation in Older Adults Influences Taste in an In Vitro Cell Model

 and
and
Abstract
:1. Introduction
2. Subjects and Materials
2.1. Study Group
2.2. Saliva Collection
2.3. Analysis of Salivary Viscosity Using Differential Dynamic Microscopy (DDM)
2.4. Total Protein Concentration
2.5. Gel Electrophoresis
2.6. Western Blotting
2.7. Detection of Sialic Acid
2.8. Semi-Quantification of Immuno-Blots
2.9. Cell Culture
2.10. Mucin Muco-Adhesion Assay
2.11. Transfection and mRNA Analysis
2.11.1. Purification of Plasmid DNA
2.11.2. Primer Design
2.12. DNA Agarose Gel Electrophoresis
2.13. Determination of DNA/RNA Concentration
2.14. Transfection of TR146 and TR146/MUC1 Cells
2.15. RNA Extraction
2.16. Reverse Transcription
2.17. PCR Procedure
2.17.1. Polymerase Chain Reaction (PCR)
2.17.2. FLEX Station Intracellular Calcium Measurements
3. Statistics
4. Results
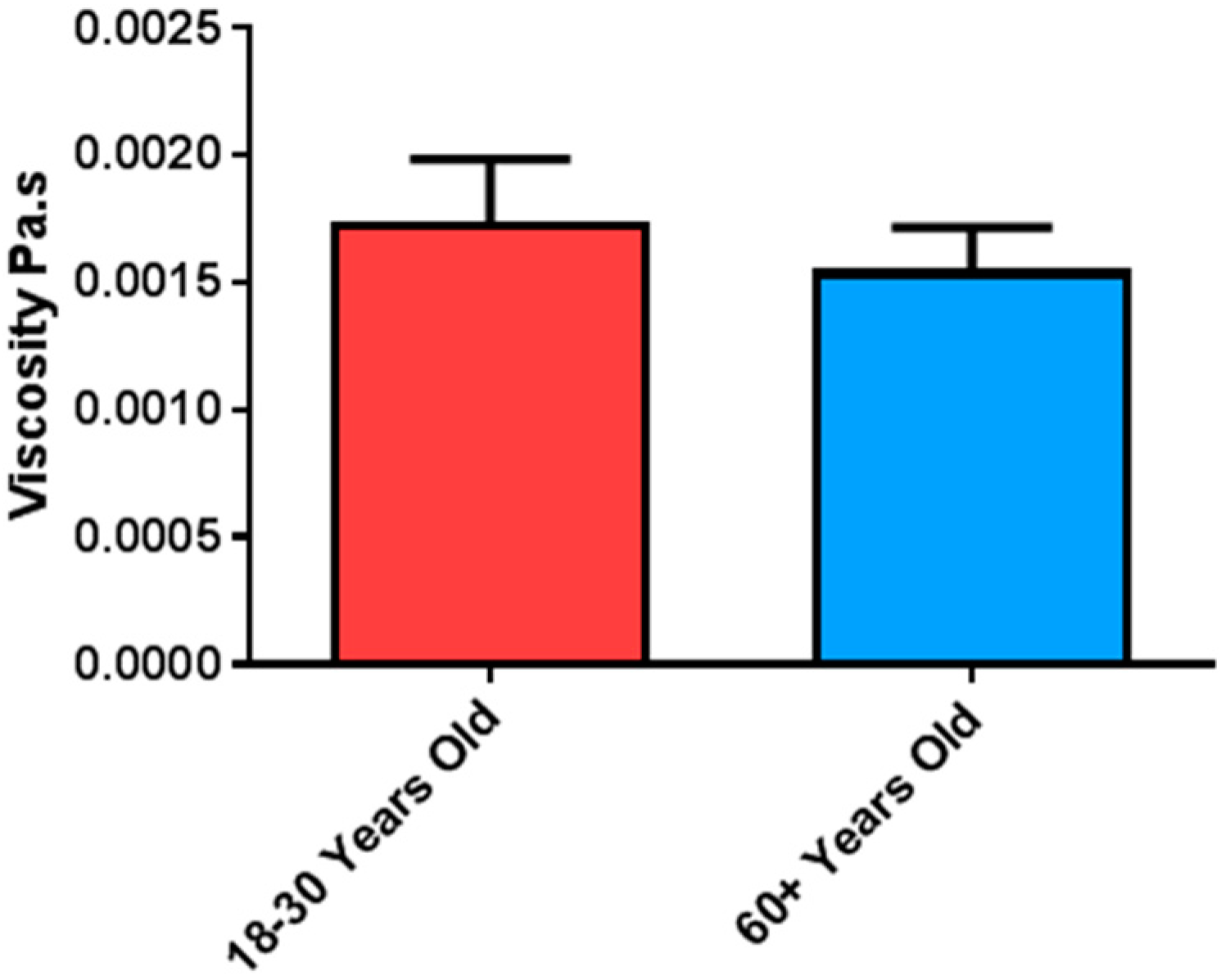
4.1. Comparative Viscosity of Saliva in Ageing
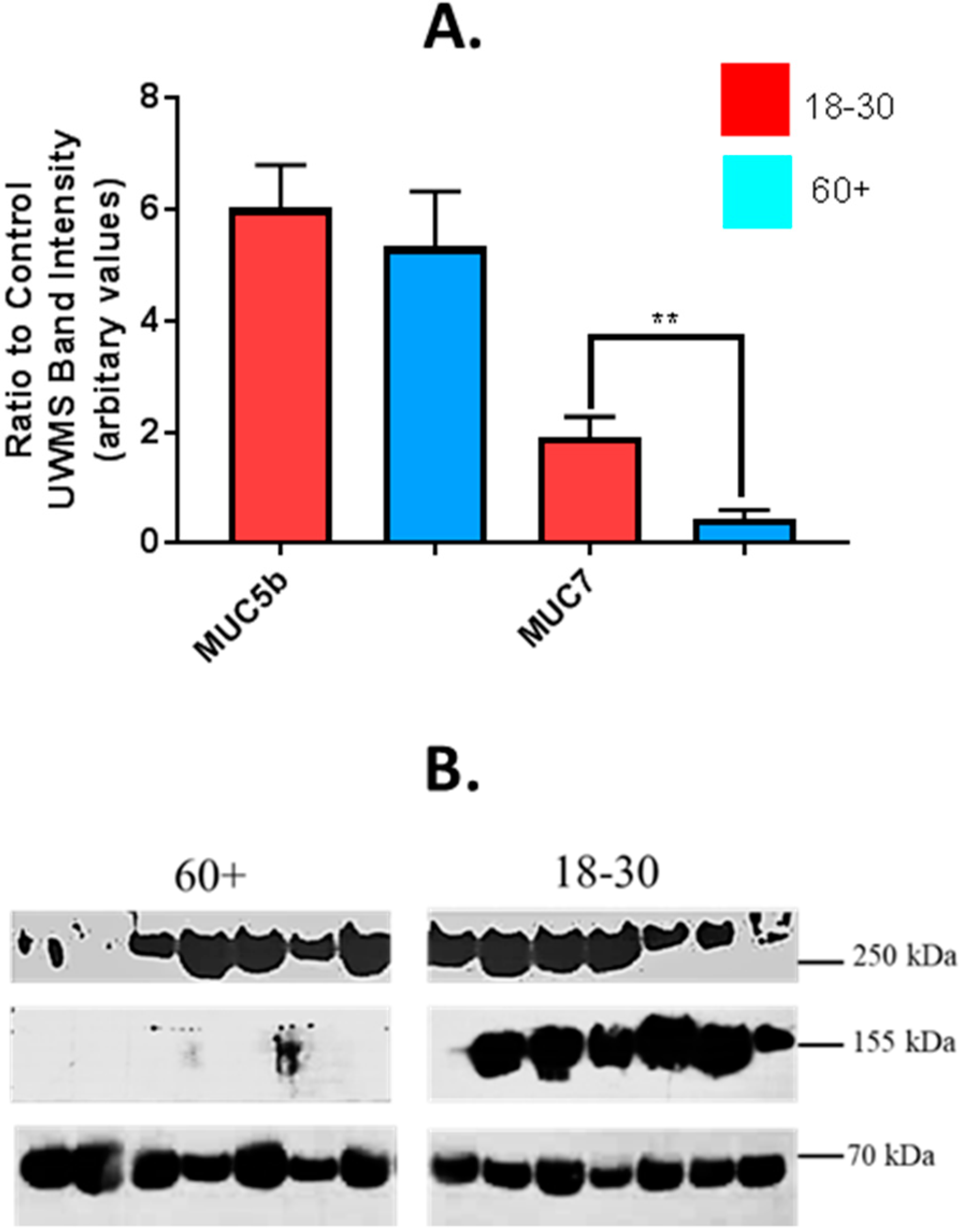
4.2. Analysis of MUC5b and MUC7 Expression in Saliva from Older and Younger Adults
4.3. Effect of Age on Sialylation of MUC5b and MUC7
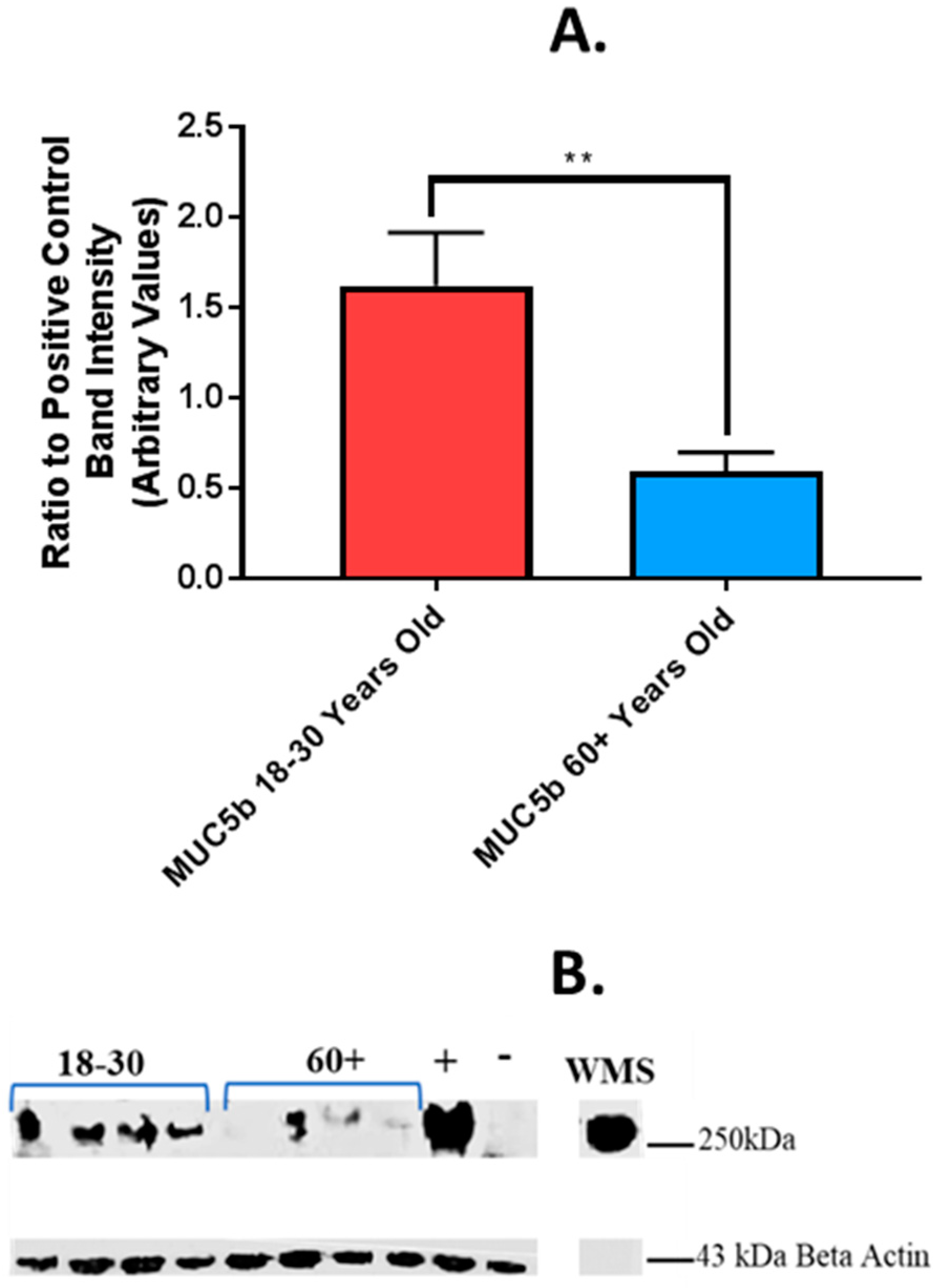
4.4. Effect of Age of Donor on Salivary MUC5b Binding in an In Vitro Model of the Oral Epithelium
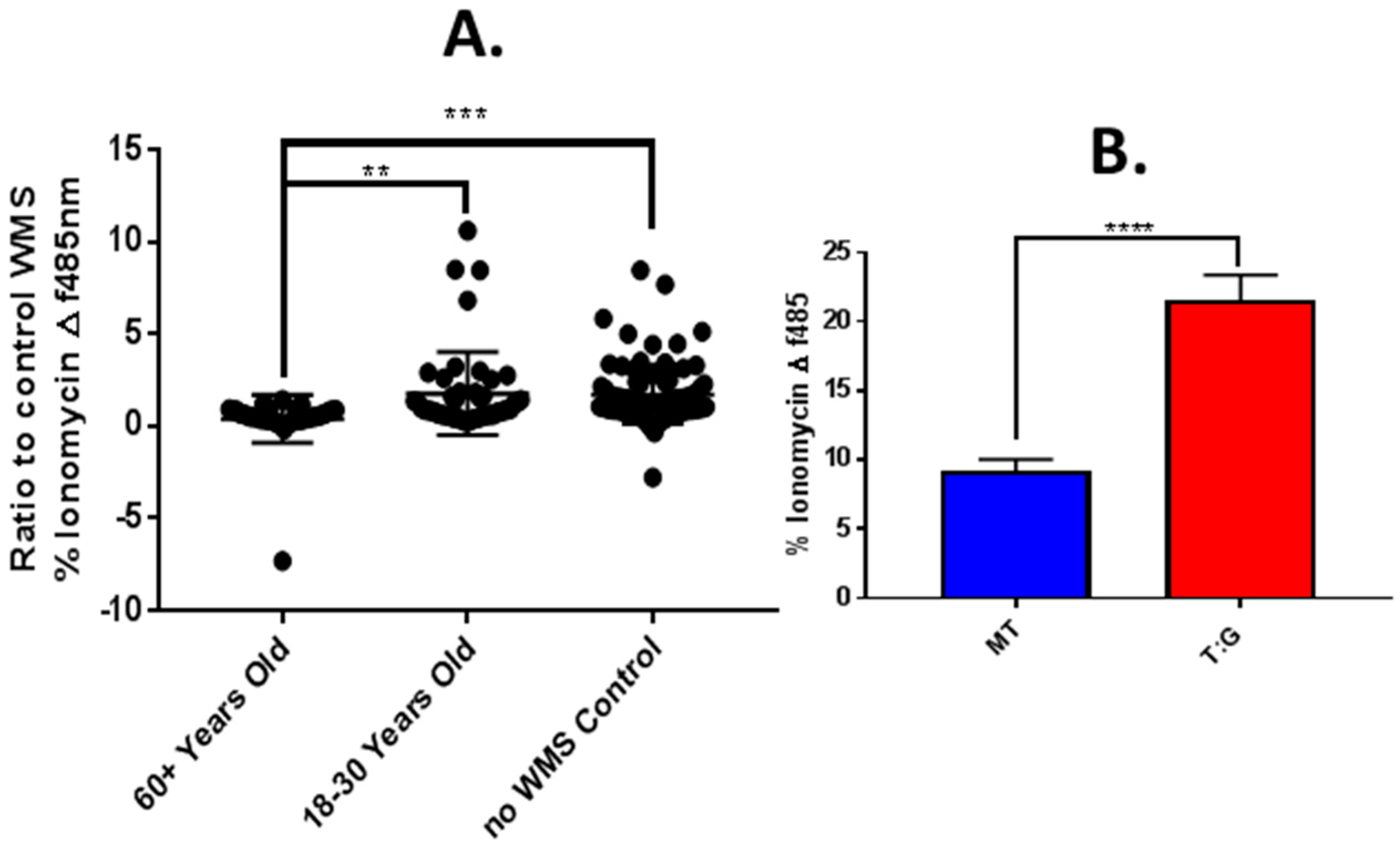
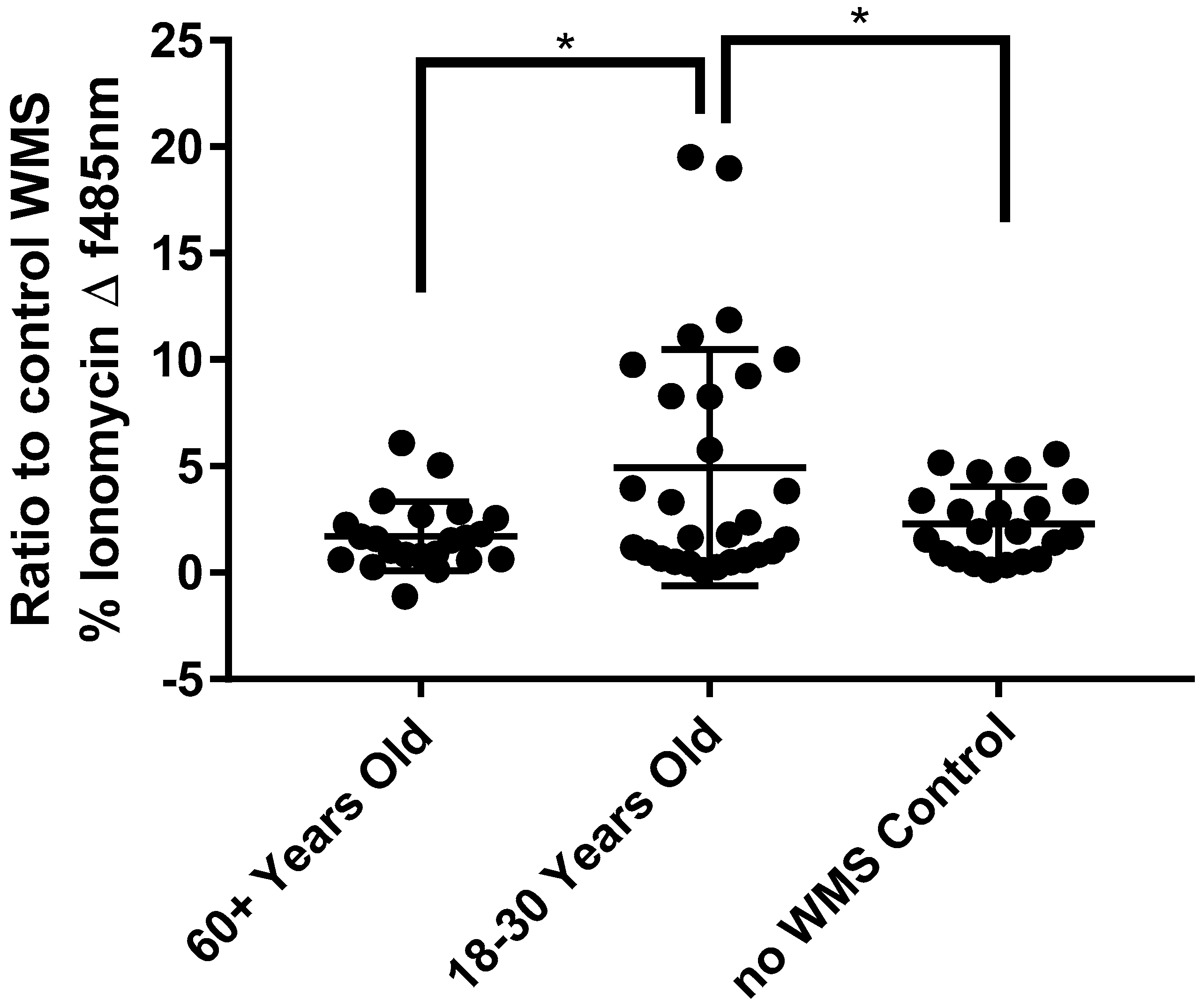
4.5. Modulation of In Vitro Responses to Tastants by Saliva from Older and Younger Adults
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BCA | Bicinchoninic acid |
| MUC1 | mucin type 1 |
| MUC5b | mucin type 5, b |
| MUC7 | mucin type 7 |
| T1R/TAS1R | taste receptor type 1 |
| T2R/TAS2R | taste receptor type 2 |
| WS | whole mouth saliva |
| UWS | un-stimulated whole mouth saliva |
| SWS | stimulated whole mouth saliva |
References
- Yoshinaka, M.; Ikebe, K.; Uota, M.; Ogawa, T.; Okada, T.; Inomata, C.; Takeshita, H.; Mihara, Y.; Gondo, Y.; Masui, Y. Age and sex differences in the taste sensitivity of young adult, young-old and old-old Japanese. Geriatr. Gerontol. Int. 2016, 16, 1281–1288. [Google Scholar] [CrossRef] [PubMed]
- Methven, L.; Allen, V.J.; Withers, C.A.; Gosney, M.A. Ageing and taste. Proc. Nutr. Soc. 2012, 71, 556–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, M.J.; Wu, F.; Guo, Y.; Robledo, L.M.G.; O’Donnell, M.; Sullivan, R.; Yusuf, S. The burden of disease in older people and implications for health policy and practice. Lancet 2015, 385, 549–562. [Google Scholar] [CrossRef]
- Brownie, S. Why are elderly individuals at risk of nutritional deficiency? Int. J. Nurs. Pract. 2006, 12, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, R. Role of Saliva in the Maintenance of Taste Sensitivity. Crit. Rev. Oral Biol. Med. 2000, 11, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Mese, H.; Matsuo, R. Salivary secretion, taste and hyposalivation. J. Oral Rehabil. 2007, 34, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Pushpass, R.-A.G.; Daly, B.; Kelly, C.; Proctor, G.; Carpenter, G.H. Altered salivary flow, protein composition and rheology following taste and TRP stimulation in older adults. Front. Physiol. 2019, 10, 652. [Google Scholar] [CrossRef] [PubMed]
- Cook, S.L.; Bull, S.P.; Methven, L.; Parker, J.K.; Khutoryanskiy, V.V. Mucoadhesion: A food perspective. Food Hydrocoll. 2017, 72, 281–296. [Google Scholar] [CrossRef]
- Gallardo-Escamilla, F.; Kelly, A.; Delahunty, C. Mouthfeel and flavour of fermented whey with added hydrocolloids. Int. Dairy J. 2007, 17, 308–315. [Google Scholar] [CrossRef]
- Cook, S.L.; Woods, S.; Methven, L.; Parker, J.K.; Khutoryanskiy, V.V. Mucoadhesive polysaccharides modulate sodium retention, release and taste perception. Food Chem. 2018, 240, 482–489. [Google Scholar] [CrossRef]
- Gardner, R. Lipophilicity and bitter taste. J. Pharm. Pharmacol. 1978, 30, 531–536. [Google Scholar] [CrossRef] [PubMed]
- Wiet, S.; Miller, G. Does chemical modification of tastants merely enhance their intrinsic taste qualities? Food Chem. 1997, 58, 305–311. [Google Scholar] [CrossRef]
- Brockhoff, A.; Behrens, M.; Massarotti, A.; Appendino, G.; Meyerhof, W. Broad tuning of the human bitter taste receptor hTAS2R46 to various sesquiterpene lactones, clerodane and labdane diterpenoids, strychnine, and denatonium. J. Agric. Food Chem. 2007, 55, 6236–6243. [Google Scholar] [CrossRef] [PubMed]
- Bufe, B.; Hofmann, T.; Krautwurst, D.; Raguse, J.-D.; Meyerhof, W. The human TAS2R16 receptor mediates bitter taste in response to β-glucopyranosides. Nat. Genet. 2002, 32, 397. [Google Scholar] [CrossRef] [PubMed]
- Meyerhof, W.; Batram, C.; Kuhn, C.; Brockhoff, A.; Chudoba, E.; Bufe, B.; Appendino, G.; Behrens, M. The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 2010, 35, 157–170. [Google Scholar] [CrossRef] [PubMed]
- Hershkovich, O.; Nagler, R.M. Biochemical analysis of saliva and taste acuity evaluation in patients with burning mouth syndrome, xerostomia and/or gustatory disturbances. Arch. Oral Biol. 2004, 49, 515–522. [Google Scholar] [CrossRef]
- Temmel, A.F.; Quint, C.; Schickinger-Fischer, B.; Hummel, T. Taste function in xerostomia before and after treatment with a saliva substitute containing carboxymethylcellulose. J. Otolaryngol. 2005, 34, 116. [Google Scholar] [CrossRef]
- Satoh-Kuriwada, S.; Shoji, N.; Kawai, M.; Uneyama, H.; Kaneta, N.; Sasano, T. Hyposalivation strongly influences hypogeusia in the elderly. J. Health Sci. 2009, 55, 689–698. [Google Scholar] [CrossRef]
- Matsuo, R.; Yamauchi, Y.; Morimoto, T. Role of submandibular and sublingual saliva in maintenance of taste sensitivity recorded in the chorda tympani of rats. J. Physiol. 1997, 498, 797–807. [Google Scholar] [CrossRef]
- Nanda, R.; Catalanotto, F.A. Basic biological sciences: Long-term effects of surgical desalivation upon taste acuity, fluid intake, and taste buds in the rat. J. Dent. Res. 1981, 60, 69–76. [Google Scholar] [CrossRef]
- Gurkan, S.; Bradley, R.M. Autonomic control of von Ebner’s lingual salivary glands and implications for taste sensation. Brain Res. 1987, 419, 287–293. [Google Scholar] [CrossRef]
- Gurkan, S.; Bradley, R.M. Secretions of von Ebner’s glands influence responses from taste buds in rat circumvallate papilla. Chem. Senses 1988, 13, 655–661. [Google Scholar] [CrossRef]
- De Almeida Pdel, V.; Gregio, A.M.; Machado, M.A.; de Lima, A.A.; Azevedo, L.R. Saliva composition and functions: A comprehensive review. J. Contemp. Dent. Pract. 2008, 9, 72–80. [Google Scholar] [PubMed]
- Pedersen, A.M.L.; Sørensen, C.E.; Proctor, G.; Carpenter, G.; Ekström, J. Salivary secretion in health and disease. J. Oral Rehabil. 2018, 45, 730–746. [Google Scholar] [CrossRef] [PubMed]
- DeSimone, J.A.; Heck, G.L. An analysis of the effects of stimulus transport and membrane charge on the salt, acid and water-response of mammals. Chem. Senses 1980, 5, 295–316. [Google Scholar] [CrossRef]
- Bufe, B.; Breslin, P.A.; Kuhn, C.; Reed, D.R.; Tharp, C.D.; Slack, J.P.; Kim, U.-K.; Drayna, D.; Meyerhof, W. The molecular basis of individual differences in phenylthiocarbamide and propylthiouracil bitterness perception. Curr. Biol. 2005, 15, 322–327. [Google Scholar] [CrossRef]
- Kim, U.; Drayna, D. Genetics of individual differences in bitter taste perception: Lessons from the PTC gene. Clin. Genet. 2005, 67, 275–280. [Google Scholar] [CrossRef]
- Kim, A.J.; Manoharan, V.N.; Crocker, J.C. Swelling-based method for preparing stable, functionalized polymer colloids. J. Am. Chem. Soc. 2005, 127, 1592–1593. [Google Scholar] [CrossRef]
- Cerbino, R.; Cicuta, P. Perspective: Differential dynamic microscopy extracts multi-scale activity in complex fluids and biological systems. J. Chem. Phys. 2017, 147, 110901. [Google Scholar] [CrossRef]
- Feriani, L.; Juenet, M.; Fowler, C.J.; Bruot, N.; Chioccioli, M.; Holland, S.M.; Bryant, C.E.; Cicuta, P. Assessing the Collective Dynamics of Motile Cilia in Cultures of Human Airway Cells by Multiscale DDM. Biophys. J. 2017, 113, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Ployon, S.; Belloir, C.; Bonnotte, A.; Lherminier, J.; Canon, F.; Morzel, M. The membrane-associated MUC1 improves adhesion of salivary MUC5B on buccal cells. Application to development of an in vitro cellular model of oral epithelium. Arch. Oral Biol. 2016, 61, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Ugawa, S.; Yamamura, H.; Imaizumi, Y.; Shimada, S. Functional interaction between T2R taste receptors and G-protein α subunits expressed in taste receptor cells. J. Neurosci. 2003, 23, 7376–7380. [Google Scholar] [CrossRef] [PubMed]
- Greene, T.A.; Alarcon, S.; Thomas, A.; Berdougo, E.; Doranz, B.J.; Breslin, P.A.; Rucker, J.B. Probenecid inhibits the human bitter taste receptor TAS2R16 and suppresses bitter perception of salicin. PLoS ONE 2011, 6, e20123. [Google Scholar] [CrossRef] [PubMed]
- Navazesh, M.; Mulligan, R.A.; Kipnis, V.; Denny, P.A.; Denny, P.C. Comparison of whole saliva flow rates and mucin concentrations in healthy Caucasian young and aged adults. J. Dent. Res. 1992, 71, 1275–1278. [Google Scholar] [CrossRef]
- Lumley, T.; Diehr, P.; Emerson, S.; Chen, L. The importance of the normality assumption in large public health data sets. Annu. Rev. Public Health 2002, 23, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Denny, P.C.; Denny, P.A.; Klauser, D.K.; Hong, S.H.; Navazesh, M.; Tabak, L.A. Age-related changes in mucins from human whole saliva. J. Dent. Res. 1991, 70, 1320–1327. [Google Scholar] [CrossRef] [PubMed]
- Baughan, L.; Robertello, F.; Sarrett, D.; Denny, P.; Denny, P. Salivary mucin as related to oral Streptococcus mutans in elderly people. Mol. Oral Microbiol. 2000, 15, 10–14. [Google Scholar] [CrossRef]
- Chaudhury, N.M.A.; Proctor, G.B.; Karlsson, N.G.; Carpenter, G.H.; Flowers, S.A. Reduced Mucin-7 (Muc7) Sialylation and Altered Saliva Rheology in Sjögren’s Syndrome Associated Oral Dryness. Mol. Cell. Proteom. 2016, 15, 1048–1059. [Google Scholar] [CrossRef]
- Rousseau, K.; Wickstrom, C.; Whitehouse, D.B.; Carlstedt, I.; Swallow, D.M. New monoclonal antibodies to non-glycosylated domains of the secreted mucins MUC5B and MUC7. Hybrid. Hybridomics 2003, 22, 293–299. [Google Scholar] [CrossRef]
- Chang, W.-I.; Chang, J.-Y.; Kim, Y.-Y.; Lee, G.; Kho, H.-S. MUC1 expression in the oral mucosal epithelial cells of the elderly. Arch. Oral Biol. 2011, 56, 885–890. [Google Scholar] [CrossRef]
- Malkki, Y.; Heinio, R.; Autio, K. Influence of oat gum, guar gum and carboxymethyl cellulose on the perception of sweetness and flavor. Food Hydrocoll. 1993, 6, 525–532. [Google Scholar] [CrossRef]
- Larhed, A.W.; Artursson, P.; Gråsjö, J.; Björk, E. Diffusion of drugs in native and purified gastrointestinal mucus. J. Pharm. Sci. 1997, 86, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Norris, D.A.; Sinko, P.J. Effect of size, surface charge, and hydrophobicity on the translocation of polystyrene microspheres through gastrointestinal mucin. J. Appl. Polym. Sci. 1997, 63, 1481–1492. [Google Scholar] [CrossRef]
- Matthes, I.; Nimmerfall, F.; Sucker, H. Mucus models for investigation of intestinal absorption mechanisms. 2. Mechanisms of drug interactions with intestinal mucus. Die Pharm. 1992, 47, 609–613. [Google Scholar]
- Schiffman, S.S.; Gatlin, L.A.; Frey, A.E.; Heiman, S.A.; Stagner, W.C.; Cooper, D.C. Taste perception of bitter compounds in young and elderly persons: Relation to lipophilicity of bitter compounds. Neurobiol. Aging 1994, 15, 743–750. [Google Scholar] [CrossRef]
- Lim, L.-Y.; Go, M.-L. Caffeine and nicotinamide enhances the aqueous solubility of the antimalarial agent halofantrine. Eur. J. Pharm. Sci. 2000, 10, 17–28. [Google Scholar] [CrossRef]
- Rasool, A.A.; Hussain, A.A.; Dittert, L.W. Solubility enhancement of some water-insoluble drugs in the presence of nicotinamide and related compounds. J. Pharm. Sci. 1991, 80, 387–393. [Google Scholar] [CrossRef]
- Sharma, B.; Paul, S. Role of caffeine as an inhibitor in aggregation of hydrophobic molecules: A molecular dynamics simulation study. J. Mol. Liq. 2016, 224, 930–939. [Google Scholar] [CrossRef]
- Van der Reijden, W.; Veerman, E.; Nieuw Amerongen, A. Shear rate dependent viscoelastic behavior of human glandular salivas. Biorheology 1993, 30, 141–152. [Google Scholar] [CrossRef]
- Van der Reijden, W.A.; Veerman, E.C.; Nieuw Amerongen, A.V. Rheological properties of commercially available polysaccharides with potential use in saliva substitutes. Biorheology 1994, 31, 631–642. [Google Scholar] [CrossRef]
- Sajewicz, E. Effect of saliva viscosity on tribological behaviour of tooth enamel. Tribol. Int. 2009, 42, 327–332. [Google Scholar] [CrossRef]
- Sánchez, E.R.B.; Honores, M.J.C. Effect of orthodontic fixed appliances on salivary flow and viscosity. Rev. Mex. Ortod. 2015, 3, 186–190. [Google Scholar] [CrossRef]
- Rantonen, P.J.; Meurman, J.H. Viscosity of whole saliva. Acta Odontol. Scand. 1998, 56, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Gittings, S.; Turnbull, N.; Henry, B.; Roberts, C.J.; Gershkovich, P. Characterisation of human saliva as a platform for oral dissolution medium development. Eur. J. Pharm. Biopharm. 2015, 91, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ligtenberg, A.; Liem, E.; Brand, H.; Veerman, E. The effect of exercise on salivary viscosity. Diagnostics 2016, 6, 40. [Google Scholar] [CrossRef] [PubMed]
- Briedis, D.; Moutrie, M.; Balmer, R. A study of the shear viscosity of human whole saliva. Rheol. Acta 1980, 19, 365–374. [Google Scholar] [CrossRef]
- Martin, D.S.; Forstner, M.B.; Käs, J.A. Apparent subdiffusion inherent to single particle tracking. Biophys. J. 2002, 83, 2109–2117. [Google Scholar] [CrossRef]
- Jaedicke, K.M.; Taylor, J.J.; Preshaw, P.M. Validation and quality control of ELISAs for the use with human saliva samples. J. Immunol. Methods 2012, 377, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Schipper, R.G.; Silletti, E.; Vingerhoeds, M.H. Saliva as research material: Biochemical, physicochemical and practical aspects. Arch. Oral Biol. 2007, 52, 1114–1135. [Google Scholar] [CrossRef]
- Derrien, M.; van Passel, M.W.; van de Bovenkamp, J.H.; Schipper, R.; de Vos, W.; Dekker, J. Mucin-bacterial interactions in the human oral cavity and digestive tract. Gut Microbes 2010, 1, 254–268. [Google Scholar] [CrossRef] [Green Version]
- Ito, F.; Yamada, S.; Mizuno, Y.; Sugihara, N.; Chen, L. Correlation between viscosity and sialic acid content of whole human saliva. Aichi-Gakuin Dent. Sci. 1988, 1, 21–27. [Google Scholar] [PubMed]
- Coles, J.M.; Chang, D.P.; Zauscher, S. Molecular mechanisms of aqueous boundary lubrication by mucinous glycoproteins. Curr. Opin. Colloid Interface Sci. 2010, 15, 406–416. [Google Scholar] [CrossRef]
- Milton, J.C.; Hill-Smith, I.; Jackson, S.H. Prescribing for older people. BMJ 2008, 336, 606–609. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligo Name | 5′–3′ Sequence | Expected Product Size (Base Pairs) | Melting Temperature (°C) |
|---|---|---|---|
| Gα16Gust44 forward | CCT GGT TCA AAA GCA CAT CCG | 254 | 68.6 |
| Gα16Gust44 reverse | TTG GGT GTC AGT AGC ACA GGT | 64.6 | |
| YWHAZ forward | ACT TTT GGT ACA TTG TGG CTT CAA | 94 | 65.3 |
| YWHAZ reverse | CCG CCA GGA CAA ACC AGT AT | 66.0 | |
| TAS2R7 forward | GGA TTC TAC TGG GGT GCG TGG T | 260 | 70.8 |
| TAS2R7 reverse | ATA GTC CGC TTA CGT CGA GTC AC | 65.4 | |
| TAS2R43 forward | ATC TGG GCA GTG ATC AAC CA | 156 | 65.6 |
| TAS2R43 reverse | TAG CAA AGG CCC CAA CAA CA | 67.5 | |
| TAS2R38 forward | AGG CCC ACA TTA AAG CCC TC | 204 | 60.03 |
| TAS2R38 reverse | CAG CTC TCC TCA ACT TGG CA | 59.96 | |
| TAS2R10 forward | GTG TAG TGG AAG GCA TCT TCA | 296 | 61.7 |
| TAS2R10 reverse | GCT GGT GGC AAA CCA CAT AC | 65.3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pushpass, R.-A.G.; Pellicciotta, N.; Kelly, C.; Proctor, G.; Carpenter, G.H. Reduced Salivary Mucin Binding and Glycosylation in Older Adults Influences Taste in an In Vitro Cell Model. Nutrients 2019, 11, 2280. https://doi.org/10.3390/nu11102280
Pushpass R-AG, Pellicciotta N, Kelly C, Proctor G, Carpenter GH. Reduced Salivary Mucin Binding and Glycosylation in Older Adults Influences Taste in an In Vitro Cell Model. Nutrients. 2019; 11(10):2280. https://doi.org/10.3390/nu11102280
Chicago/Turabian StylePushpass, Rose-Anna G., Nicola Pellicciotta, Charles Kelly, Gordon Proctor, and Guy H. Carpenter. 2019. "Reduced Salivary Mucin Binding and Glycosylation in Older Adults Influences Taste in an In Vitro Cell Model" Nutrients 11, no. 10: 2280. https://doi.org/10.3390/nu11102280