Whole Grain Consumption for the Prevention and Treatment of Breast Cancer


1
State Key Laboratory of Food Nutrition and Safety, Tianjin University of Science & Technology; School of Food Engineering and Biotechnology, Tianjin University of Science & Technology, Tianjin 300457, China
2
China-Canada Joint Lab of Food Nutrition and Health (Beijing), Beijing Technology & Business University (BTBU), Beijing 100048, China
3
Guelph Research and Development Centre, Agriculture and Agri-Food Canada, Guelph, N1G 5C9, Canada
*
Author to whom correspondence should be addressed.
Nutrients 2019, 11(8), 1769; https://doi.org/10.3390/nu11081769
Submission received: 23 June 2019
/
Revised: 28 July 2019
/
Accepted: 30 July 2019
/
Published: 1 August 2019
(This article belongs to the Special Issue Grains and Human Health)
Abstract
:Breast cancer is one of the most common and malignant cancers among females worldwide. Several epidemiological studies have indicated the inverse correlation between the intake of whole grains and the incidence of breast cancer. Whole grains are the most fundamental and important food source of bioactive phytochemicals, which have well-defined roles in the management of each stage of breast carcinogenesis. To better understand the value of whole grains in future prevention and treatment of breast cancer, the effects and possible mechanisms of six different whole grain cereals, which are the most commonly consumed throughout the world, are introduced in the current review. Moreover, the bioactive compounds extracted from whole grains are adequately formulated and the underlying mechanism of action is illustrated. In addition, the present limitations and future perspective of whole grain consumption for breast cancer are also concluded. The objective of this review is to promote the development of nutraceutical and functional food from whole grains and its application for reducing the risk of breast cancer.
1. Introduction
Breast cancer is the most frequently diagnosed cancer among women worldwide and has become an increasing global health issue over the past few decades. The World Cancer Research Fund (WCRF) states that dietary habits have a critically important role in the prevention and causation of cancer [1]. Epidemiological studies have showed that breast cancer is more prevalent in Europe and North America than in Asian countries [2,3]. One of the most important reasons for this may be ascribed to different dietary patterns. There are many evidences that support the inverse relation between whole grain intake and breast cancer risks [3,4,5,6], indicating that whole grains might hold nutraceutical characteristics against breast cancer. As a result, many efforts have been made in the identification and characterization of phytochemicals from whole grains with potential chemo-preventive properties. However, the relationship between whole grains and breast cancer remains not fully understood. Besides, previous research on this topic does not link closely with the current development in cancer prevention and management, such as synergistic application with adjuvant therapy. Therefore, a systematic review over the current research status of preventive effects of whole grains (and its products) against breast cancer is urgently needed.
In view of this, the main objective of this review is to summarize and provide an overview of the chemo-preventive effects of whole grain products and their bioactive compounds against breast cancer and promote their applications for the prevention and management of breast cancer. Considering the fundamental status of whole grains in citizens’ livelihood, the development of whole-grain functional foods is especially important and beneficial for human health. The findings might also be suggestive for guiding policy initiatives and nutritional suggestions for cancer prevention.
2. Breast Cancer
2.1. General Aspect of Breast Cancer
Cancer remains a major cause of mortality and morbidity around the world [7]. Breast cancer is one of the most frequently diagnosed carcinoma in women [1]. According to data of GLOBOCAN 2018, there were 2.1 million newly diagnosed breast cancer cases in 2018, representing a quarter of all new cancer cases in women worldwide [8]. In the United States, there are 268,600 newly diagnosed cases, representing 30% of all new cases in women, and 41,400 deaths, ranking second in the U.S. in 2019 [2,9]. In the meantime, breast cancer death risk ranks the second in all-age U.S. women and first in women between the ages of 20–59 [2]. Moreover, breast cancer is the most frequently diagnosed cancer and the sixth leading cause of death in Chinese women [10,11]. In 2015, there were approximately 270,000 new cases of breast cancer, accounting for 15% of newly diagnosed cancers in women, and 69,500 deaths, making it the leading cause of death in Chinese women younger than the age of 45 [10]. Although, the mortality of breast cancer in North America and Europe has dropped due to advances in early detection and systemic endocrine and targeted therapies. However, the breast cancer toll is still increasing in China [12].
Breast cancer is a complex disease featuring distinct histological, clinical, and molecular phenotypes. A lot of aspects can affect breast cancer development, including intrinsic factors, e.g. family history and genetic mutation, as well as extrinsic factors, e.g. environment, radiation, living styles, and reproduction condition [1,13,14,15,16]. With technology advances in the last decade, researchers have revealed that breast cancer is not just one single disease, but a heterogeneous group of imbalances, and therefore, the responses to different therapeutic treatments vary significantly [17,18]. According to the molecular receptor status (estrogen receptor (ER), progesterone receptor (PR), and human epidermal growth factor receptor-2 (HER-2)) and the gene expression detected by multiple microarray platforms, breast cancer is categorized into various subtypes: luminal A (ER/PR-positive/HER-2-negative), luminal B (ER-positive, PR-positive or negative, and HER-2-positive), basal-like (ER/PR/HER-2-negative), normal breast-like (ER/PR-positive or negative and HER-2-negative), HER-2-enriched, and claudin-low (ER/PR-negative, HER-2-positive, and low expression of claudin) (Table 1) [19,20,21]. Luminal breast cancer (luminal A and luminal B) are featured with positive expression of estrogen (ER) and/or progesterone (PR). Compared with luminal A, luminal B is more aggressive and worse in prognosis. Both subtypes show high expression of luminal-related genes (e.g. ESR1, KRT8) and of ER transcription factors (e.g. GATA3, FOXA1). HER-2-enriched subtype is characterized by positive expression of human epidermal growth factor receptor 2 (HER-2) as well as negative expression of ER. HER-2-enriched breast cancer cells have high expression of genes including HER-2, GRB7, and have higher migration ability compared with luminal cells. Lacking ER, PR, and HER-2, triple-negative breast cancer cells are the most heterogeneous among all intrinsic subtypes of breast cancer. Basal-like subtype is the major one of the basal tumor subtypes for its high expression in basal gene markers such as KRT5, LAMC2, FABP7, etc. Normal basal-like and claudin-low subtype represent the mesenchymal subtype cluster. These subtypes are highly expressed in gene markers featured in highly invasive and aggressive breast cancer phenotypes such as VIM, MMP2/14, COL3A1, CD24(-), and CD44(+).Classification based on gene expression profile is still developing [22,23], which makes characterization of breast cancer even more complex. The distinct subtypes of breast tumor respond differently to treatments, making breast cancer extremely challenging.
Breast cancer mortality has declined significantly in Western countries in the past few decades due to early diagnosis and effective adjuvant therapies [15,24,25]. Currently, for early stage breast cancer, treatment strategies include both local (surgery and radiotherapy) and systemic therapies that combine endocrine, targeted, and chemotherapies. Conventional cytotoxic therapies, including chemotherapy and radiation therapy, are the most common choices for cancer management. However, many women still suffer from relapse and tumor metastasis. The efficacy of conventional chemotherapy and radiotherapy is far from satisfactory due to drug/radiotherapy resistance and side effects. Developments of novel anticancer agents and alternative approaches with high selectivity and therapeutic indices and relatively low toxicity have become a new direction for cancer chemotherapy research and development [26]. There is an urgent call for novel and effective prevention and treatment strategies for breast cancer.
2.2. Molecular Mechanism of Breast Cancer Treatment
2.2.1. Proliferation Inhibition
One major feature of cancer is a dysregulated and aggressive proliferation of the tumor cells. In normal and healthy cells, proliferation is precisely regulated through a balance between the growth and antigrowth signals. However, cancer cells develop the ability to grow uncontrollably, which generate their own growth signals and become insensitive to antigrowth signals. Apoptosis is a programmed cell death process that is continually occurring in normal cells [27]. The caspases and the members of the Bcl-2 family of proteins were involved in modulating cells’ apoptosis [28]. Studies have proven that the mitogen-activated protein kinase (MAPK), the phosphoinositide 3-kinase (PI3K)/Akt, and the epidermal growth factor receptor (EGFR) signal transduction cascades function abnormally in most tumor cases and therefore, they became key targets for targeted therapy-mediated cell death [29]. The PI3K-AKT-mTOR pathway is a central knot in the transduction of extracellular and intracellular growth signals. Inhibition of PI3K pathway signaling reduced cancer cell growth and survival [30].
2.2.2. Immune System Modulation
There is clear evidence that the immune system and inflammation play a critical role in the process of carcinogenesis. An inflammatory microenvironment is an essential component of all tumors [31]. Inflammatory responses are involved in every stage of cancer development and progression, such as initiation, promotion, malignant transformation, invasion, and metastasis [32]. The immune system can eliminate premalignant and transformed cancer cells. However, cancer cells can bypass the immune system through the development of resistant or immunogenic clones. Various immune cells are frequently found accumulated in tumors relative to the surrounding tissue. These immune cells infiltrate tumors and communicate with tumor cells [33]. The important link between inflammation and carcinogenesis is the pro-inflammatory transcription factors. Moreover, the inflammatory mediators such as pro-inflammatory cytokines also stimulate the survival and proliferation of premalignant cells and activate oncogenic transcription factors [34]. Blocking pro-inflammatory cytokines or endotoxin-mediated kinases and transcription factors involved in cancer progression could inhibit inflammation and cancer recurrence.
2.2.3. Targeting Metastasis and Breast Cancer Stem Cells
Emerging studies have revealed that breast cancer stem cells’ (BCSCs) subpopulation and their epithelial-mesenchymal transition (EMT) behavior are major reasons for resistance and migratory potential [35,36,37]. Conventional chemotherapy would increase the percentage of CD44high/CD24low tumor cells in breast cancer patients, which acts as a breast cancer stem cell marker [38]. Mammary cells developed into BCSCs by EMT [36,39,40], a process by which epithelial cells transit into mesenchymal phenotype, allowing them to be free from the primary tumor site and metastasize at distant sites [39]. EMT signaling is involved in the development and maintenance of BCSCs. BCSCs and EMT are involved in a crosslinked signaling network, including TGF-β, Wnt, Notch, NF-κB, and ERK/MAPK pathways [41]. Apart from the intrinsic signal pathway, BCSCs are also regulated by the tumor microenvironment [35], which contains inflammatory cells, BCSCs, fibroblasts, and cytokines. Several inflammatory cytokines, including IL-6 and IL-8, have been reported to regulate breast cancer stem cell self-renewal through the cytokine loop [42]. Developing strategies to interfere with cytokines and their receptors are in progress to target breast cancer stem cells.
3. Anti-Breast-Cancer Efficacy of Whole Grains
3.1. General Health Benefits of Whole Grains
Many foods are considered as key elements for cancer prevention, and phytochemicals in whole grains are especially important [1]. Whole grains are the edible seeds of the grass family plants, which are composed of the embryo (or germ), endosperm, starch granules, as well as the outside bran (fiber). Grain plants are highly adaptable to the environment and thus are widely cultivated in different climatic and geographical conditions. Wheat, rice, millet, rye, barley, sorghum, oats, and maize are the most fundamental and important source of food and energy around the world. Grains are rich in carbohydrates, proteins, minerals, and vitamins.
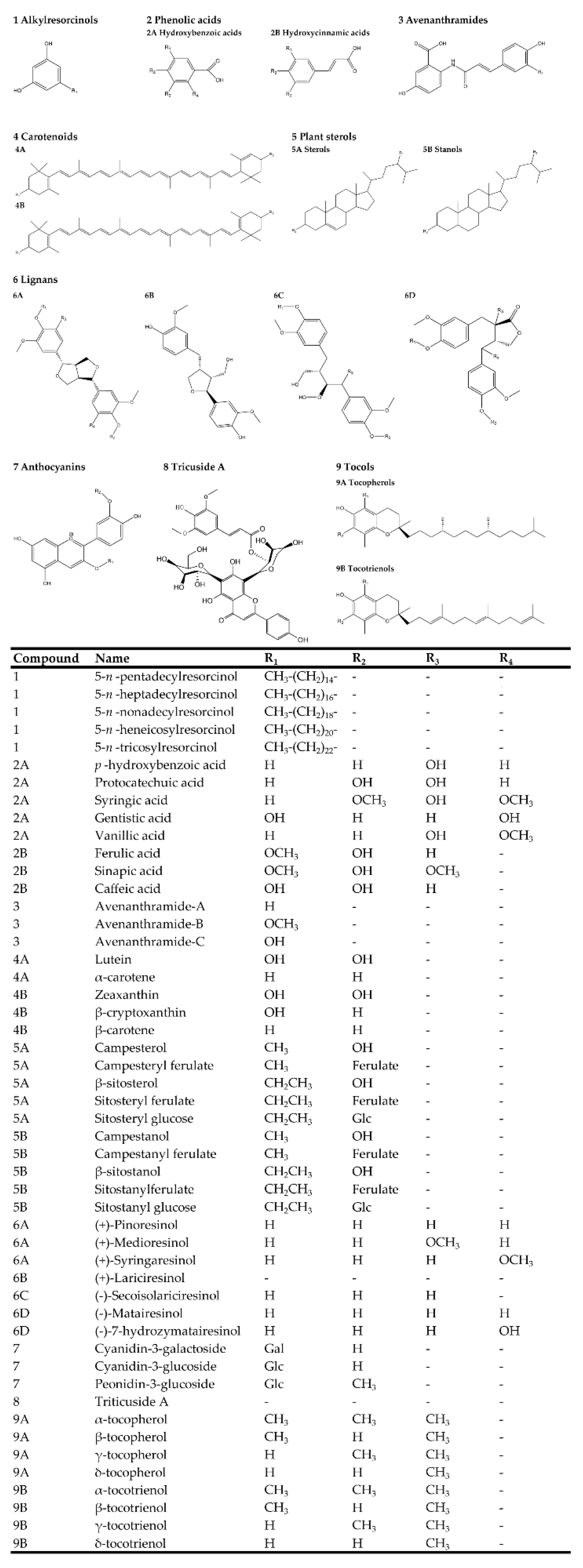
Consumption of whole grains has been proven to decrease the risk of obesity, diabetes, and cardiovascular diseases [43,44,45,46,47,48,49,50,51,52,53,54]. Over the last few decades, accumulating evidence has shown that high intake of whole grain products prevents cancer occurrence [53,55,56,57,58]. These health benefits of whole grains could be attributed to the abundant phytochemicals in different cereal species [59]. The major bioactive phytochemicals found in grains are listed in Figure 1. There has been a huge amount of literature describing the benefits of whole grains including antioxidant, anticancer, anti-inflammatory, as well as illness risk reduction activity [60,61,62,63]. These health benefits are briefly reviewed in Table 2.
3.2. Epidemiological and Clinical Studies of Whole Grain Consumption and Breast Cancer
Clinical epidemiological studies of whole grain consumption and breast cancer have been summarized in Table 3. A recent meta-analysis suggested that a healthy dietary pattern could lower breast cancer risk, especially in postmenopausal, hormone receptor–negative women [6]. Epidemiological studies also suggest that the consumption of whole grains is related to a reduced risk of breast cancer [81]. A recent meta-analysis including four cohort and seven case-control studies suggested that high whole grains intake might have an inverse association with breast cancer risk (RR (relative risks) = 0.84; 95%CI = 0.74–0.96; P = 0.009; I2 = 63.8%). Further stratified analysis indicated that the inverse association was only observed in case-control studies (RR = 0.69; 95%CI = 0.56–0.87; P = 0.001; I2 = 58.2%), but not in cohort studies (RR = 0.96; 95%CI = 0.82–1.14; P = 0.69; I2 = 66.7%) [5]. Two other reviews and meta-analyses of prospective studies correlate dietary fiber intake and reduced breast cancer risk [55]. Data from a more recent cohort analysis of American females suggested an inverse relation between whole grains (or dietary fiber intakes) and breast cancer risks in both adolescent population and adults before menopause [81,82]. Further stratified analysis indicated that higher adult intake of whole grains was associated with lower premenopausal breast cancer risks (RR = 0.82; 95%CI = 0.70–0.97), and that an inverse relation between higher adolescent and early-adulthood intake of whole grains and breast cancer risks (RR = 0.74; 95%CI = 0.56–0.99) existed. Adulthood consumption of brown rice was linked with lower overall risks of breast cancer (2 servings/week: RR = 0.94; 95%CI = 0.89–0.99). A case-controlled study from Greece also suggested that whole grains consumption more than 7 times/week was consistently associated with reduced risk of breast cancer (OR (odds ratios) = 0.49; 95% CI = 0.29-0.82) for women between 44–68 years old [6]. Another case-control study from Iran suggested that the consumption of resistant starch (RS)-containing food, such as whole grain bread, could reduce breast cancer risk (OR = 0.61; 95%CI = 0.37–0.99) for women aged from 25 to 65 years [83]. A 2017 cohort study showed that the consumption of whole grain food may protect against breast cancer risk with a 47% reduction rate (HR (hazard ratios) = 0.53; 95%C I = 0.33–0.86), while consumption of whole grain food had no clear association with other adiposity-related cancers [56]. Dietary intervention with whole grains and related products could be a pragmatic approach for breast cancer prevention and management. Their inherent safety makes whole grain foods an appealing choice for widespread, long-term use in diverse populations.
Although results of most epidemiological studies support an inverse relationship between whole grain (or dietary fiber) consumption and breast cancer risks, more evidences are needed to draw firm conclusions. Controversies still exist. A review of five cohort studies found that there is no solid association between whole grain consumption and breast cancer risks, and there is no statistically significant relationship between whole grain fiber consumption and breast cancer risk in any of the 11 cohort studies conducted in Europe and North America [57]. Among all studies above, four studies announced a significant decrease in cancer risk, yet one study reported a non-significant increase in cancer risk with a heavy intake of whole grains. A possible explanation for this inconsistency in results might be varied study designs and a lack of risk factor adjustments. Moreover, the assessment standard of whole grains (or grain fibers) intake was changed between the different studies [57]. The inability to measure whole-grain intake might lead to diverse, even opposite results from existing evidence. Therefore, more large-scale cohort studies with more standardized whole grain intake assessment methods are needed to confirm the relationship between grains intake and breast cancer risks in the future.
3.3. Whole Grain Phytochemicals and Anti-Breast-Cancer Property
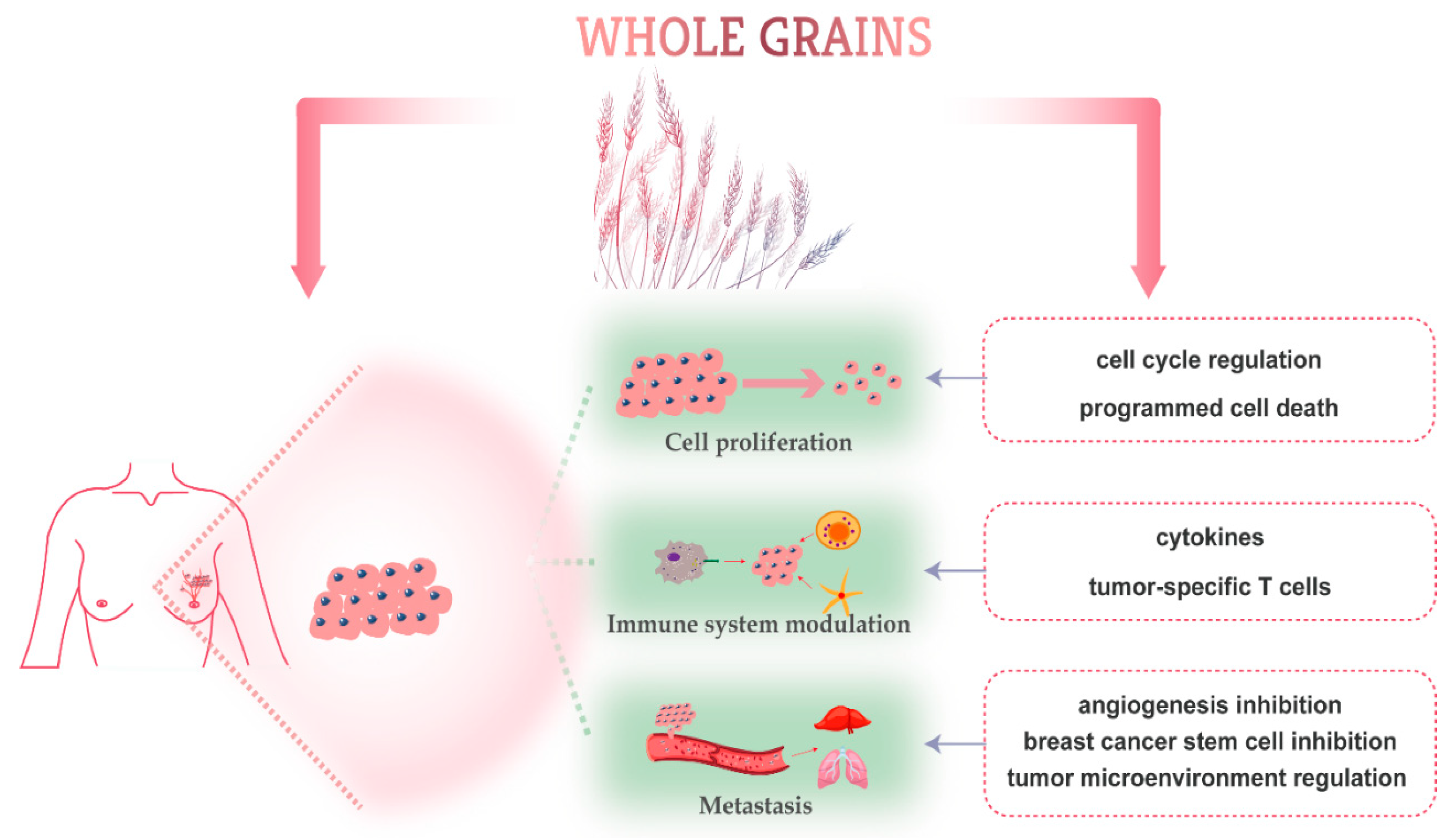
Phytochemicals are non-essential nutrient bioactive components found in plant foods. Whole grains are a rich source of phytochemicals such as phenolic acids, carotenoids, alkylresorcinols (ARs), phytosterols, lignans, anthocyanins, vitamin E member, and polysaccharides (Figure 1) [85]. The anti-cancer activities and potential health benefits can be attributed to the abundant bioactive phytochemicals in whole grains [57]. Whole grains’ dietary fiber function directly in the small intestine and colon, therefore their effects on colorectal carcinogenesis are most intensely studied [52,58,86,87,88]. Unlike dietary fiber, the anticancer activity of some phytochemicals is still needed for future comprehensive investigations [89]. Several experimental studies have shown that bioactive components of whole grains exert anti-breast cancer activity through inhibiting proliferation, modulating immune system, and inhibiting metastasis of breast tumor cells [90]. Below, we will overview some of the most common whole grains and discuss their role in breast cancer prevention (Table 4) and the major breast cancer molecular mechanisms targeted by whole grain-derived phytochemicals (Figure 2).
3.3.1. Wheat
Wheat is the most cultivated crop in the world [104]. Wheat grains and their processed products are staple foods and are one of the most important dietary energy sources. Components of germinated wheat flour have been shown to have an inhibitory effect against the human breast cancer cell line MCF-7(ER+) and MDA-MB-231 (TNBC, triple negative) by up-regulating apoptosis of both cell lines [91]. ARs, or 1,3-dihydroxy-5-n-alkylbenzenes, are one major group of phenolic lipids found in whole grain wheat, barley, and rye. ARs can be absorbed and detected in plasma. Thus, ARs can be used as biomarkers for whole grain wheat/rye consumption [105,106,107]. A wide range of bioactive properties of ARs have been reported including antioxidative and anti-carcinoma activities, suppressing adipocyte lipolysis, obesity reduction, and increasing glucose tolerance and insulin sensitivity [64,65,105,108,109,110,111,112,113]. A recent paper reviewed the relation between dietary alkylresorcinols and cancer prevention, concluding that alkylresorcinols are likely to be useful in impeding cancer progression [114]. A series of ARs (C17:0–C25:0) isolated from wheat bran oil showed growth inhibition potential against human colon cancer cell HCT-116 and HT-29 [64], indicating that wheat bran ARs do have an anti-carcinoma ability. There are a few preliminary in vitro studies proving that ARs could inhibit the growth of human breast cancer cell. Five ARs isolated from Homalomena wendlandii Schott(Areacae) inhibited the growth of the breast cancer adenocarcinoma cell line MCF-7 with IC50 values ranging between 8.24–42.17μM [115]. Currently, there have been no direct studies conducted to evaluate the effect of grain-originated ARs on breast cancer, yet the effect is predictable. Besides ARs, triticuside A, a flavone C-glycoside from wheat bran, strongly suppressed the proliferation of human breast cancer cells (MCF-7 and MDA-MB-231) via the mitochondrial apoptosis pathway and the Akt/mTOR signaling pathway [95].
3.3.2. Rice
Rice is a major part of the global food supply, serving as a staple for over 50% of the world’s population. In Asia, nearly half of the grains consumed annually is rice. Unlike white rice with the husk, bran, and germ removed, whole grain rice like red rice and black rice contain higher contents of bioactive phytochemicals, such as phenolics, oryzanol, tocotrienols, and tocopherols that possess various beneficial health activities. Black rice anthocyanins significantly suppressed breast cancer metastasis in several studies [40,71,103]. In a recent study, black rice anthocyanins were shown to inhibit HER-2 breast cancer cell metastasis by suppressing cancer cell growth, migration, and epithelial-mesenchymal transition via the cSrc/FAK/p130Cas signaling pathway [40]. Red mold rice, fermented by Monascus purpureus NTU 803, is a traditional food and folk medicine in East Asia. Extracts of red mold rice exhibited direct cytotoxic and proapoptotic effects on breast cancer cell MCF-7 through activation of Caspase-9 and Caspase-3 of the mitochondria-dependent pathway in a time-dependent manner [116].
A recent study indicated that black rice anthocyanins could suppress HER-2-positive breast cancer cells invasion by targeting the RAS/RAF/MAPK pathway, a pivotal signaling pathway in breast cancer development [71]. Black rice anthocyanins inhibited HER-2+ MDA-MB-453 human breast cancer cell migration and invasion, suppressed the activation of RAF, MEK, and JNK, and downregulated the secretion of MMP-2 and MMP-9. In another study, proanthocyanin, γ-oryzanol, and γ-tocotrienol extracted from red rice fractions were found to have anti-invasion activity against HT1080 and MDA-MB-231 cancer cells by decreasing the expression and activity of matrix metalloproteinase-2 and -9 (MMP-2 and -9) [98]. Black rice anthocyanins also exerted anti-metastasis potential against human breast cancer cells by reducing transplanted tumor growth and inhibiting pulmonary metastasis of breast cancer xenografts and decreasing urokinase-type plasminogen activator (u-PA) activity [103]. Recent studies also suggest that bioactive peptides of food origin have been shown to play important roles in the prevention and treatment of cancer and cardiovascular and infective diseases [117]. Many peptides extracted from whole grains exhibit anticancer potential, however most related studies focus on efficacy over gastrointestinal cancer [118]. In one study, a pentapeptide with a sequence of Glu-Gln-Arg-Pro-Arg, isolated from rice bran, showed antiproliferative characteristics against both MCF-7 and MDA-MB-231 cells [99].
3.3.3. Sorghum
Sorghum is the fifth most economic cereal crop in the world, and the health benefits of its unique phytochemicals have been under intense investigation in recent years [68,119,120]. Various human health benefits associated with sorghum can be attributed to its abundant secondary metabolites such as anthocyanins, phenols, tannins, phytosterols, and policosanols [121]. Anthocyanins extracted from red sorghum bran showed an anti-proliferative effect against MCF-7 cancer cells by inducing apoptosis, as revealed in the formation of apoptosis body and DNA fragmentations [101]. A similar result was found in another study where red-sorghum-bran 3-deoxyanthocyanin inhibited MCF-7 cells viability by upregulating the p53 expression and down regulating the Bcl-2 expression [102]. Sorghum extract suppressed the growth of MDA-MB-231 and MCF-7 cells through inducing the G1 cell cycle phase arrest, down-regulating the STAT5b/IGF-1R and STAT3/VEGF pathways, and inhibiting metastasis in BALB/C nude mice bearing breast cancer xenografts [100]. Results showed that tumor growth was suppressed by sorghum extracts through modulating JAK/STAT pathways and downregulating the expression of angiogenic factors like VEGF, VEGF-R2, and cell cycle regulators like cyclin D, cyclin E, and p-Rb. The same study further showed that sorghum extracts exhibited significantly higher anticancer bioactivity than other grains like wheat, millet, and panicum, indicating that sorghum might serve as an effective and inexpensive edible supplement in cancer management [100]. Sorghum 3-deoxyanthocyanidins were proven to be more cytotoxic to cancer cells than anthocyanidins from other foods [122]. Apart from 3-deoxyanthocyanidins, sorghum tannins also have anticancer potential. In one study, sorghum bran extract, which was rich in tannins, inhibited aromatase activity, a key enzyme in estrogen synthesis and an important target for breast cancer chemotherapy [123]. Further studies are needed for the anticancer effect of sorghum tannins.
3.3.4. Oat
Oats are an outstanding source of soluble dietary fibers such as β-glucans and health-beneficial phytochemicals. Oat β-glucans have been intensely studied against multiple cancer cells [124,125] and in animal models [79]. Oral administration of β-glucans showed an immunomodulatory effect by stimulating peripheral blood monocytes proliferation in advanced breast cancer patients [126]. However, the relation between oat β-glucan consumption and breast cancer still needs to be further explored. Avenacosides, a unique group of steroidal saponins from oats, have drawn growing research attention for their chemopreventive potential against human colon cancer cells [127]. Among all phytochemicals of oats, avenanthramides are uniquely found in oats. Avenanthramides are found to possess a wide range of health benefits [67]. Avenanthramide-C was shown to reduce the viability of MDA-MB-231 breast cancer cells through the induction of sub G1 cell cycle arrest and apoptosis, causing DNA fragmentation and activation of Caspases [94]. Dihydroavenanthramide D, a synthetic analog of avenanthramides, suppressed breast cancer growth by inhibiting MCF-7 cancer cell invasion through the MAPK/NF-κB and MAPK/AP-1 pathway [93].
3.3.5. Other Grain Species
Consumption of other cereals and phytochemicals have also been found to have positive impacts on breast cancer risk reduction and anti-proliferative activities. Barley consumption, for example, has been shown to improve health [45]. In one study, barley exhibited anti-tumor activities in both the rat mammary tumor model and MCF-7 cell line through the induction of cell cycle arrest, pro-apoptosis, and the antiproliferation mechanism [92]. Young barley significantly increased caspase-3 and reduced Ki67 expression in rat tumors [92]. Millet is also an important small-seed grain crop, which is widely cultivated in China and East Asia, but its health benefits are greatly underestimated. Phenolic extract of foxtail millet was found to strongly inhibit the proliferation of MDA-MB-231 breast cancer cells [61]. However, more research needs to be conducted on the specific anti-breast cancer components and their role in chemoprevention.
3.3.6. Synergistic Effects of Whole Grain Phytochemicals and Anti-Breast-Cancer Therapy Agents
Current cancer therapeutic strategies often do not receive good and stable efficacy, due to toxicity, resistance, and tumor metastasis. Chemotherapy is one of the most frequently used treatments in cancer management. However, it is often followed by adverse effects and resistance, which is the major causes for disease recurrence. Some whole grain products and related phytochemicals have shown synergistic effects with chemotherapy, either by sensitizing the tumor cells or by reducing their side effects. Wheat grass juice taken by breast cancer patients under combined chemotherapy (5-fluorouracil, doxorubicin, and cyclophosphamide) was found to have lowered levels of toxicity and reduced chemotherapies dosage, without decreasing their efficacy [128]. Two rice bran components, δ-tocotrienol and ferulic acid, synergistically inhibited the proliferation of human breast cancer cell MCF-7, while ferulic acid alone did not show any anti-proliferative effect [96]. MGN-3/Biobran, arabinoxylan acquired from rice bran, increased the susceptibility of MCF-7 cells and murine metastatic breast cancer cell 4T-1 to paclitaxel by more than 100-fold, resulting in increased DNA damage and apoptosis [97]. MGN-3/Biobran was also reported to increase the susceptibility of human breast cancer cells MCF-7 and HCC70 to daunorubicin by enhancing drug accumulation in cancer cells [129].
4. Discussion
In this review, we described literature investigations on cereal bioactivity against breast cancer. We have also overviewed molecular mechanisms of cereal phytochemicals, offering new options for adjuvant therapy development. Whole grain cereals may reduce breast cancer risk via multiple ways. Firstly, whole grains could influence the obesity situation by promoting satiation and satiety, reducing caloric intake, and controlling body adiposity due to its high fiber content, which can lower energy density [130,131]. Abundant fiber content in whole grains could significantly control insulin resistance and insulin-like growth factors expression [56,132]. Therefore, decreasing body adiposity and insulin resistance could contribute to reducing risks of cancer. Moreover, whole grains may also affect breast cancer through regulating hormone levels. Whole grain foods are sources of phytoestrogen-like isoflavones, which could influence hormone levels and activities [57]. Dietary fiber in whole grains may decrease circulating estrogen concentrations by suppressing bacterial β-glucuronidase activity, raising transient time and peristaltic activities in the gut, which inhibits the reabsorption of estrogens in the colon and increases the excretion of estrogens in feces [84]. Phytoestrogens and dietary fiber together would decrease inner estrogen concentrations, inhibit tumor development, and weaken early stage cancer risk marker expression. In addition, many bioactive phytochemicals are uniquely found in commonly consumed whole grains and are highly effective in modulating signaling pathways that are involved in breast cancer occurrence and progression. Thus, the consumption of whole grains containing these micronutrients may be beneficial for breast cancer prevention as hitherto discussed. The potential of whole grain consumption in reducing breast cancer risk is also supported by epidemiological and experimental evidences.
However, several important issues remain to be solved. Firstly, current epidemiological studies on the relationship between whole grain consumption and breast cancer took “whole grains” as the consumption parameter, but did not set different grain species as individual parameters for assessment. Concentration of the bioactive phytochemicals vary significantly among different cereals and growing regions. Future studies should consider using standardized specific species and cultivars with unique bioactive compositions as an experimental factor. The dietary pattern of different geographic regions should also be taken into consideration. Secondly, while many comprehensive studies have evaluated the anti-breast cancer properties of grain extracts including wheat, sorghum, and rice, only a few have closely examined the specific phytochemicals and their mechanisms. Structure activity relationship characteristics of the phytochemicals and their pharmacophoric functionalities should be comprehensively studied. The inability to identify the individual active components is a great obstacle to interpreting the existing results’ correlation between whole grain consumption and breast cancer risk. Thirdly, the new molecular and genetic classification of cancer has provided alternative aspects for the diagnosis and treatment of breast cancer [133,134]. Focus should be placed on the various molecular targets of different breast cancer phenotypes by comprehensively studying the anticancer phytochemicals of whole grains. Moreover, chemo/radio-resistance remains to be one of the major obstacles during metastatic breast cancer treatment. Therefore, the combination of cereal phytochemicals with cytotoxic chemotherapy and/or selective BCSC-target agents could be a novel adjuvant therapy strategy for combating cancer metastasis. In addition, tumor immunotherapy has received increased attention due to its high specificity and low body toxicity [26]. The ability of cereal bioactive components to modulate immune response in breast cancer should be explored in future. More efforts should be placed on the in vivo studies and human intervention trials investigating the preventive effect of whole grains and/or their phytochemicals. Last, but not least, exploiting the synergistic potential of whole grain phytochemicals with chemotherapeutic and targeted therapies should be seriously and comprehensively considered. It is of paramount importance that future research focuses on integrated approaches that combine dietary or nutraceutical interventions with systemic endocrine therapies or chemotherapies.
5. Conclusions
Breast cancer is one of the most pervasive and fatal carcinomas worldwide, which represents a serious threat to women’s health. There is an urgent need for the development and implementation of novel and effective therapeutic strategies that are affordable and accessible. Whole grains account for a large proportion of the dietary structure in developing countries around the world. Epidemiological studies have demonstrated that whole grains (and its products) are associated with reduced risk of breast cancer. Therefore, increasing whole grain consumption in daily dietary structure is a practical strategy for breast cancer prevention.
Whole grains are rich in unique bioactive phytochemicals, which have been proven to be effective at targeting signaling pathways in breast cancer. Utilizing these phytochemicals synergistically with current standard therapy strategies may be a feasible approach for breast cancer treatment. Future studies on the health benefits of whole grains phytochemicals should consider targeting drug resistance and optimizing delivery method.
Author Contributions
M.X. prepared the manuscript and generated the schematic diagrams, J.L. and R.T. conceived the topic and revised the manuscript, Z.W. revised the final version, B.S. and J.W. coordinated the working group and revised the final version. All the authors made significant contributions to this review article.
Acknowledgments
This research was financially supported by the Chinese National Natural Science Foundation (Grant 31701640), Beijing Excellent Talents Funding for Youth Scientist Innovation Team (2016000026833TD01), and Support Project of High-level Teachers in Beijing Municipal Universities (IDHT20180506).
Conflicts of Interest
The authors declare that there are no conflict of interest.
Abbreviations
| BC | breast cancer |
| BCSC | breast cancer stem cells |
| BRCAs | breast cancer growth suppressor proteins |
| EMT | epithelial-mesenchymal transition |
| ER | estrogen receptor |
| ErbB2 | erythroblastic leukemia viral oncogene homolog 2 |
| FAK | focal adhesion kinase |
| Gal | galactose |
| Glc | glucose |
| HER-2 | human epidermal growth factor receptor-2 |
| IGF-1R | insulin-like growth factor 1 receptor |
| MMPs | matrix metallopeptidases |
| PR | progesterone receptor |
| RAF | rapidly accelerated fibrosarcoma |
| MEK | MAPK/Erk kinase |
| JNK | Jun N-terminal kinase |
| u-PA | urokinase-type plasminogen activator |
| SRC | steroid receptor coactivator |
| STAT | signal transducer and activator of transcription |
| VEGF | vascular endothelial growth factor |
| p130Cas | p130 Crk-associated substrate |
References
- World Cancer Research Fund/American Institute for Cancer Research. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; World Cancer Research Fund/American Institute for Cancer Research: Washington, DC, USA, 2018. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2018. CA Cancer J. Clin. 2018, 68, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Dreher, M.L. Dietary Patterns, Whole Plant Foods, Nutrients and Phytochemicals in Breast Cancer Prevention and Management. In Dietary Patterns and Whole Plant Foods in Aging and Disease; Dreher, M.L., Ed.; Humana Press: Cham, Switzerland, 2018; pp. 557–609. [Google Scholar]
- Khan, S.I.; Aumsuwan, P.; Khan, I.A.; Walker, L.A.; Dasmahapatra, A.K. Epigenetic events associated with breast cancer and their prevention by dietary components targeting the epigenome. Chem. Res. Toxicol. 2012, 25, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.J.; Ke, Y.B.; Wu, S.; Huang, S.L.; Li, S.G.; Lv, Z.Q.; Yeoh, E.K.; Lao, X.Q.; Wong, S.; Kim, J.H.; et al. Association between whole grain intake and breast cancer risk: A systematic review and meta-analysis of observational studies. Nutr. J. 2018, 17, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Mourouti, N.; Kontogianni, M.D.; Papavagelis, C.; Psaltopoulou, T.; Kapetanstrataki, M.G.; Plytzanopoulou, P.; Vassilakou, T.; Malamos, N.; Linos, A.; Panagiotakos, D.B. Whole Grain Consumption and Breast Cancer: A Case-Control Study in Women. J. Am. Coll. Nutr. 2016, 35, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, E. Breast cancer in a Renaissance Book of the Dead. Lancet Oncol. 2018, 19, 1023–1024. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.D.; Nogueira, L.; Mariotto, A.B.; Rowland, J.H.; Yabroff, K.R.; Alfano, C.M.; Jemal, A.; Kramer, J.L.; Siegel, R.L. Cancer treatment and survivorship statistics, 2019. CA Cancer J. Clin. 2019. [Google Scholar] [CrossRef]
- Chen, W.Q.; Zheng, R.S.; Zhang, S.W.; Zeng, H.M.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Strasser-Weippl, K.; Li, J.J.; St Louis, J.; Finkelstein, D.M.; Yu, K.D.; Chen, W.Q.; Shao, Z.M.; Goss, P.E. Breast cancer in China. Lancet Oncol. 2014, 15, 279–289. [Google Scholar] [CrossRef]
- Jia, M.; Zheng, R.; Zhang, S.; Zeng, H.; Zou, X.; Chen, W. Female breast cancer incidence and mortality in 2011, China. J. Thorac. Dis. 2015, 7, 1221–1226. [Google Scholar] [CrossRef]
- Reeves, G.K.; Beral, V.; Green, J.; Gathani, T.; Bull, D. Hormonal therapy for menopause and breast-cancer risk by histological type: A cohort study and meta-analysis. Lancet Oncol. 2006, 7, 910–918. [Google Scholar] [CrossRef]
- Pharoah, P.D.P.; Day, N.E.; Duffy, S.; Easton, D.F.; Ponder, B.A.J. Family history and the risk of breast cancer: A systematic review and meta-analysis. Int. J. Cancer 2015, 71, 800–809. [Google Scholar] [CrossRef]
- Steward, B.W.; Wild, C.P. World Cancer Report 2014; International Agency for Research on Cancer, WHO Press: Lyon, France, 2014. [Google Scholar]
- Ronckers, C.M.; Erdmann, C.A.; Land, C.E. Radiation and breast cancer: A review of current evidence. Breast Cancer Res. 2005, 7, 21–32. [Google Scholar] [CrossRef]
- Rakha, E.A.; Green, A.R. Molecular classification of breast cancer: What the pathologist needs to know. Pathology 2017, 49, 111–119. [Google Scholar] [CrossRef]
- Perou, C.M.; Borresen-Dale, A.L. Systems biology and genomics of breast cancer. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–17. [Google Scholar] [CrossRef]
- Eroles, P.; Bosch, A.; Perez-Fidalgo, J.A.; Lluch, A. Molecular biology in breast cancer: Intrinsic subtypes and signaling pathways. Cancer Treat. Rev. 2012, 38, 698–707. [Google Scholar] [CrossRef]
- Reis-Filho, J.S.; Pusztai, L. Gene expression profiling in breast cancer: Classification, prognostication, and prediction. Lancet 2011, 378, 1812–1823. [Google Scholar] [CrossRef]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [Green Version]
- The Cancer Genome Atlas Network. Comprehensive molecular portraits of human breast tumors. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef]
- Curtis, C.; Shah, S.P.; Chin, S.-F.; Turashvili, G.; Rueda, O.M.; Dunning, M.J.; Speed, D.; Lynch, A.G.; Samarajiwa, S.; Yuan, Y.; et al. The genomic and transcriptomic architecture of 2000 breast tumours reveals novel subgroups. Nature 2012, 486, 346–352. [Google Scholar] [CrossRef]
- Berry, D.A.; Cronin, K.A.; Plevritis, S.K.; Fryback, D.G.; Clarke, L.; Zelen, M.; Mandelblatt, J.S.; Yakovlev, A.Y.; Habbema, J.D.; Feuer, E.J.; et al. Effect of screening and adjuvant therapy on mortality from breast cancer. N. Engl. J. Med. 2005, 353, 1784–1792. [Google Scholar] [CrossRef] [PubMed]
- Torre, L.A.; Siegel, R.L.; Ward, E.M.; Jemal, A. Global Cancer Incidence and Mortality Rates and Trends—An Update. Cancer Epidemiol. Biomark. Prev. 2016, 25, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Nathan, M.R.; Schmid, P. The emerging world of breast cancer immunotherapy. Breast 2018, 37, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Myriam Calabrese, C.; Giorgio, C. Role of Nutraceuticals in Cancer Therapy. J. Food Res. 2014, 3, 18–25. [Google Scholar] [CrossRef]
- Thomadaki, H.; Talieri, M.; Scorilas, A. Treatment of MCF-7 cells with taxol and etoposide induces distinct alterations in the expression of apoptosis-related genes BCL2, BCL2L12, BAX, CASPASE-9 and FAS. Biol. Chem. 2006, 387, 1081–1086. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, A.; Krajewski, K.; Cakar, B.; Ma, C.X. Targeted therapy for breast cancer. Am. J. Pathol. 2013, 183, 1096–1112. [Google Scholar] [CrossRef]
- Hennessy, B.T.; Smith, D.L.; Ram, P.T.; Lu, Y.L.; Mills, G.B. Exploiting the PI3K/AKT pathway for cancer drug discovery. Nat. Rev. Drug Discov. 2005, 4, 988–1004. [Google Scholar] [CrossRef]
- Jiang, X.G.; Shapiro, D.J. The immune system and inflammation in breast cancer. Mol. Cell. Endocrinol. 2014, 382, 673–682. [Google Scholar] [CrossRef]
- Qian, B.Z.; Zhang, H.; Li, J.F.; Yeo, E.J.; Carragher, N.O.; Bresnick, A.R.; Lang, R.A.; Pollard, J.W. Macrophage FLT1 mediated inflammatory response determines breast cancer distal metastasis. Cancer Res. 2016, 76. [Google Scholar] [CrossRef]
- Meyer, M.A.; Baer, J.M.; Knolhoff, B.L.; Nywening, T.M.; Panni, R.Z.; Su, X.; Weilbaecher, K.N.; Hawkins, W.G.; Ma, C.; Fields, R.C.; et al. Breast and pancreatic cancer interrupt IRF8-dependent dendritic cell development to overcome immune surveillance. Nat. Commun. 2018, 9, 1–19. [Google Scholar] [CrossRef]
- Karin, M. NF-kappa B as a Critical Link Between Inflammation and Cancer. Cold Spring Harb. Perspect. Biol. 2009, 1. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Wicha, M.S. Targeting breast cancer stem cells. J. Clin. Oncol. 2010, 28, 4006–4012. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechJournal of Food Researchanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Chuthapisith, S.; Eremin, J.; El-Sheemey, M.; Eremin, O. Breast cancer chemoresistance: Emerging importance of cancer stem cells. Surg. Oncol. 2010, 19, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Li, X.X.; Lewis, M.T.; Huang, J.; Gutierrez, C.; Osborne, C.K.; Wu, M.F.; Hilsenbeck, S.G.; Pavlick, A.; Zhang, X.M.; Chamness, G.C.; et al. Intrinsic resistance of tumorigenic breast cancer cells to chemotherapy. J. Natl. Cancer Inst. 2008, 100, 672–679. [Google Scholar] [CrossRef] [PubMed]
- Morel, A.P.; Lievre, M.; Thomas, C.; Hinkal, G.; Ansieau, S.; Puisieux, A. Generation of Breast Cancer Stem Cells through Epithelial-Mesenchymal Transition. PLoS ONE 2008, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Zhu, Y.F.; Chen, X.Y.; Han, B.; Li, F.; Chen, J.Y.; Peng, X.L.; Luo, L.P.; Chen, W.; Yu, X.P. Black rice-derived anthocyanins inhibit HER-2-positive breast cancer epithelial-mesenchymal transition-mediated metastasis in vitro by suppressing FAK signaling. Int. J. Mol. Med. 2017, 40, 1649–1656. [Google Scholar] [CrossRef]
- Kotiyal, S.; Bhattacharya, S. Breast cancer stem cells, EMT and therapeutic targets. Biochem. Biophys. Res. Commun. 2014, 453, 112–116. [Google Scholar] [CrossRef]
- Korkaya, H.; Paulson, A.; Charafe-Jauffret, E.; Ginestier, C.; Brown, M.; Dutcher, J.; Clouthier, S.G.; Wicha, M.S. Regulation of mammary stem/progenitor cells by PTEN/Akt/beta-catenin signaling. PLoS Biol. 2009, 7, e1000121. [Google Scholar] [CrossRef]
- Hallmans, G.; Zhang, J.-X.; Lundin, E.; Stattin, P.; Johansson, A.; Johansson, I.; Hultén, K.; Winkvist, A.; Lenner, P.; Åman, P.; et al. Rye, lignans and human health. Proc. Nutr. Soc. 2007, 62, 193–199. [Google Scholar] [CrossRef]
- Chanson-Rolle, A.; Meynier, A.; Aubin, F.; Lappi, J.; Poutanen, K.; Vinoy, S.; Braesco, V. Systematic Review and Meta-Analysis of Human Studies to Support a Quantitative Recommendation for Whole Grain Intake in Relation to Type 2 Diabetes. PLoS ONE 2015, 10, e0131377. [Google Scholar] [CrossRef]
- Idehen, E.; Tang, Y.; Sang, S. Bioactive phytochemicals in barley. J. Food Drug Anal. 2017, 25, 148–161. [Google Scholar] [CrossRef]
- Chandra, D.; Chandra, S.; Pallavi; Sharma, A.K. Review of Finger millet (Eleusine coracana (L.) Gaertn): A power house of health benefiting nutrients. Food Sci. Hum. Wellness 2016, 5, 149–155. [Google Scholar] [CrossRef]
- Shahidi, F.; Chandrasekara, A. Millet grain phenolics and their role in disease risk reduction and health promotion: A review. J. Funct. Foods 2013, 5, 570–581. [Google Scholar] [CrossRef]
- Seo, C.-R.; Yi, B.; Oh, S.; Kwon, S.-M.; Kim, S.; Song, N.-J.; Cho, J.Y.; Park, K.-M.; Ahn, J.-Y.; Hong, J.-W.; et al. Aqueous extracts of hulled barley containing coumaric acid and ferulic acid inhibit adipogenesis in vitro and obesity in vivo. J. Funct. Foods 2015, 12, 208–218. [Google Scholar] [CrossRef]
- Luthria, D.L.; Lu, Y.; John, K.M.M. Bioactive phytochemicals in wheat: Extraction, analysis, processing, and functional properties. J. Funct. Foods 2015, 18, 910–925. [Google Scholar] [CrossRef]
- Koh-Banerjee, P.; Rimm, E.B. Whole grain consumption and weight gain: A review of the epidemiological evidence, potential mechanisms and opportunities for future research. Proc. Nutr. Soc. 2003, 62, 25–29. [Google Scholar] [CrossRef]
- Montonen, J.; Knekt, P.; Järvinen, R.; Aromaa, A.; Reunanen, A. Whole-grain and fiber intake and the incidence of type 2 diabetes. Am. J. Clin. Nutr. 2003, 77, 622–629. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.C.; Tong, X.; Xu, J.Y.; Han, S.F.; Wan, Z.X.; Qin, J.B.; Qin, L.Q. Whole-grain intake and total, cardiovascular, and cancer mortality: A systematic review and meta-analysis of prospective studies. Am. J. Clin. Nutr. 2016, 104, 164–172. [Google Scholar] [CrossRef]
- Zhang, B.; Zhao, Q.; Guo, W.; Bao, W.; Wang, X. Association of whole grain intake with all-cause, cardiovascular, and cancer mortality: A systematic review and dose–response meta-analysis from prospective cohort studies. Eur. J. Clin. Nutr. 2017, 72, 57–65. [Google Scholar] [CrossRef]
- Fardet, A. New hypotheses for the health-protective mechanisms of whole-grain cereals: What is beyond fibre? Nutr. Res. Rev. 2010, 23, 65–134. [Google Scholar] [CrossRef]
- Aune, D.; Chan, D.S.; Greenwood, D.C.; Vieira, A.R.; Rosenblatt, D.A.; Vieira, R.; Norat, T. Dietary fiber and breast cancer risk: A systematic review and meta-analysis of prospective studies. Ann. Oncol. 2012, 23, 1394–1402. [Google Scholar] [CrossRef]
- Makarem, N.; Bandera, E.V.; Lin, Y.; McKeown, N.M.; Hayes, R.B.; Parekh, N. Associations of Whole and Refined Grain Intakes with Adiposity-Related Cancer Risk in the Framingham Offspring Cohort (1991–2013). Nutr. Cancer 2018, 70, 776–786. [Google Scholar] [CrossRef]
- Makarem, N.; Nicholson, J.M.; Bandera, E.V.; McKeown, N.M.; Parekh, N. Consumption of whole grains and cereal fiber in relation to cancer risk: A systematic review of longitudinal studies. Nutr. Rev. 2016, 74, 353–373. [Google Scholar] [CrossRef]
- Tayyem, R.F.; Bawadi, H.A.; Shehadah, I.; Agraib, L.M.; Al-Awwad, N.J.; Heath, D.D.; Bani-Hani, K.E. Consumption of Whole Grains, Refined Cereals, and Legumes and Its Association With Colorectal Cancer Among Jordanians. Integr. Cancer Ther. 2016, 15, 318–325. [Google Scholar] [CrossRef]
- Okarter, N.; Liu, R.H. Health benefits of whole grain phytochemicals. Crit. Rev. Food Sci. Nutr. 2010, 50, 193–208. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Zhang, L.Z.; Liu, R.H. Phenolic and carotenoid profiles and antiproliferative activity of foxtail millet. Food Chem. 2015, 174, 495–501. [Google Scholar] [CrossRef]
- Abdal Dayem, A.; Choi, H.Y.; Yang, G.M.; Kim, K.; Saha, S.K.; Cho, S.G. The Anti-Cancer Effect of Polyphenols against Breast Cancer and Cancer Stem Cells: Molecular Mechanisms. Nutrients 2016, 8, e581. [Google Scholar] [CrossRef]
- Crascì, L.; Lauro, M.R.; Puglisi, G.; Panico, A. Natural antioxidant polyphenols on inflammation management: Anti-glycation activity vs metalloproteinases inhibition. Crit. Rev. Food Sci. Nutr. 2018, 58, 893–904. [Google Scholar] [CrossRef]
- Zhu, Y.; Conklin, D.R.; Chen, H.; Wang, L.; Sang, S. 5-alk(en)ylresorcinols as the major active components in wheat bran inhibit human colon cancer cell growth. Bioorg. Med. Chem. 2011, 19, 3973–3982. [Google Scholar] [CrossRef]
- Agil, R.; Patterson, Z.R.; Mackay, H.; Abizaid, A.; Hosseinian, F. Triticale Bran Alkylresorcinols Enhance Resistance to Oxidative Stress in Mice Fed a High-Fat Diet. Foods 2016, 5, e5–e21. [Google Scholar] [CrossRef]
- Martínez-Villaluenga, C.; Peñas, E. Health benefits of oat: Current evidence and molecular mechanisms. Curr. Opin. Food Sci. 2017, 14, 26–31. [Google Scholar] [CrossRef]
- Sang, S.; Chu, Y. Whole grain oats, more than just a fiber: Role of unique phytochemicals. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- De Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef]
- Chen, X. Functional Food-Related Bioactive Compounds Effect of Sorghum Phenolics on Cancer Cells. Ph.D. Dissertation, Kansas State University, Manhattan, KS, USA, 2017. [Google Scholar]
- Pintha, K.; Yodkeeree, S.; Limtrakul, P. Proanthocyanidin in Red Rice Inhibits MDA-MB-231 Breast Cancer Cell Invasion via the Expression Control of Invasive Proteins. Biol. Pharm. Bull. 2015, 38, 571–581. [Google Scholar] [CrossRef]
- Chen, X.Y.; Zhou, J.; Luo, L.P.; Han, B.; Li, F.; Chen, J.Y.; Zhu, Y.F.; Chen, W.; Yu, X.P. Black Rice Anthocyanins Suppress Metastasis of Breast Cancer Cells by Targeting RAS/RAF/MAPK Pathway. Biomed. Res. Int. 2015, 1–11. [Google Scholar] [CrossRef]
- Peterson, J.; Dwyer, J.; Adlercreutz, H.; Scalbert, A.; Jacques, P.; McCullough, M.L. Dietary lignans: Physiology and potential for cardiovascular disease risk reduction. Nutr. Rev. 2010, 68, 571–603. [Google Scholar] [CrossRef]
- Imran, M.; Ahmad, N.; Anjum, F.M.; Khan, M.K.; Mushtaq, Z.; Nadeem, M.; Hussain, S. Potential protective properties of flax lignan secoisolariciresinol diglucoside. Nutr. J. 2015, 14, 71. [Google Scholar] [CrossRef]
- Awika, J.M. Sorghum Flavonoids: Unusual Compounds with Promising Implications for Health. In Advances in Cereal Science: Implications to Food Processing and Health Promotion; Awika, J.M., Piironen, V., Bean, S., Eds.; American Chemical Society: Washington, DC, USA, 2011; Volume 1089, pp. 171–200. [Google Scholar]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, e1600852. [Google Scholar] [CrossRef]
- Luyen, B.T.; Thao, N.P.; Tai, B.H.; Lim, J.Y.; Ki, H.H.; Kim, D.K.; Lee, Y.M.; Kim, Y.H. Chemical constituents of Triticum aestivum and their effects on adipogenic differentiation of 3T3-L1 preadipocytes. Arch. Pharm. Res. 2015, 38, 1011–1018. [Google Scholar] [CrossRef]
- Hallikainen, M.A.; Sarkkinen, E.S.; Uusitupa, M.I. Plant stanol esters affect serum cholesterol concentrations of hypercholesterolemic men and women in a dose-dependent manner. J. Nutr. 2000, 130, 767–776. [Google Scholar] [CrossRef]
- Pi-Sunyer, X. Do glycemic index, glycemic load, and fiber play a role in insulin sensitivity, disposition index, and type 2 diabetes? Diabetes Care 2005, 28, 2978–2979. [Google Scholar] [CrossRef]
- Shen, R.-L.; Wang, Z.; Dong, J.-L.; Xiang, Q.-S.; Liu, Y.-Q. Effects of oat soluble and insoluble β-glucan on 1,2-dimethylhydrazine-induced early colon carcinogenesis in mice. Food Agric. Immunol. 2016, 27, 657–666. [Google Scholar] [CrossRef]
- Upadhyay, J.; Misra, K. Towards the interaction mechanism of tocopherols and tocotrienols (vitamin E) with selected metabolizing enzymes. Bioinformation 2009, 3, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Farvid, M.S.; Eliassen, A.H.; Cho, E.; Liao, X.; Chen, W.Y.; Willett, W.C. Dietary Fiber Intake in Young Adults and Breast Cancer Risk. Pediatrics 2016, 137, e20151226. [Google Scholar] [CrossRef] [Green Version]
- Farvid, M.S.; Cho, E.; Eliassen, A.H.; Chen, W.Y.; Willett, W.C. Lifetime grain consumption and breast cancer risk. Breast Cancer Res. Treat. 2016, 159, 335–345. [Google Scholar] [CrossRef]
- Tajaddini, A.; Pourzand, A.; Sanaat, Z.; Pirouzpanah, S. Dietary Resistant Starch Contained Foods and Breast Cancer Risk: A Case-Control Study in Northwest of Iran. Asian Pac. J. Cancer Prev. 2015, 16, 4185–4192. [Google Scholar] [CrossRef]
- Dong, J.Y.; He, K.; Wang, P.; Qin, L.Q. Dietary fiber intake and risk of breast cancer: A meta-analysis of prospective cohort studies. Am. J. Clin. Nutr. 2011, 94, 900–905. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Song, M.; Wu, K.; Meyerhardt, J.A.; Ogino, S.; Wang, M.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Fiber intake and survival after colorectal cancer diagnosis. JAMA Oncol. 2018, 4, 71–79. [Google Scholar] [CrossRef]
- Song, M.; Garrett, W.S.; Chan, A.T. Nutrients, foods, and colorectal cancer prevention. Gastroenterology 2015, 148, 1244–1260. [Google Scholar] [CrossRef]
- Kyro, C.; Olsen, A.; Landberg, R.; Skeie, G.; Loft, S.; Aman, P.; Leenders, M.; Dik, V.K.; Siersema, P.D.; Pischon, T.; et al. Plasma Alkylresorcinols, Biomarkers of Whole-Grain Wheat and Rye Intake, and Incidence of Colorectal Cancer. J. Natl. Cancer Inst. 2014, 106, djt352. [Google Scholar] [CrossRef]
- Andersson, A.A.M.; Dimberg, L.; Åman, P.; Landberg, R. Recent findings on certain bioactive components in whole grain wheat and rye. J. Cereal Sci. 2014, 59, 294–311. [Google Scholar] [CrossRef]
- Li, Y.; Li, S.; Meng, X.; Gan, R.Y.; Zhang, J.J.; Li, H.B. Dietary Natural Products for Prevention and Treatment of Breast Cancer. Nutrients 2017, 9, 728. [Google Scholar] [CrossRef]
- Cho, K.; Lee, C.W.; Ohm, J.-B. In Vitro Study on Effect of Germinated Wheat on Human Breast Cancer Cells. Cereal Chem. 2016, 93, 647–649. [Google Scholar] [CrossRef]
- Kubatka, P.; Kello, M.; Kajo, K.; Kruzliak, P.; Vybohova, D.; Smejkal, K.; Marsik, P.; Zulli, A.; Gonciova, G.; Mojzis, J.; et al. Young Barley Indicates Antitumor Effects in Experimental Breast Cancer In Vivo and In Vitro. Nutr. Cancer 2016, 68, 611–621. [Google Scholar] [CrossRef]
- Lee, Y.R.; Noh, E.M.; Oh, H.J.; Hur, H.; Kim, J.M.; Han, J.H.; Hwang, J.K.; Park, B.H.; Park, J.W.; Youn, H.J.; et al. Dihydroavenanthramide D inhibits human breast cancer cell invasion through suppression of MMP-9 expression. Biochem. Biophys. Res. Commun. 2011, 405, 552–557. [Google Scholar] [CrossRef]
- Hastings, J.; Kenealey, J. Avenanthramide-C reduces the viability of MDA-MB-231 breast cancer cells through an apoptotic mechanism. Cancer Cell Int. 2017, 17, 93–105. [Google Scholar] [CrossRef]
- Shan, Y.; Cheng, Y.; Zhang, Y.; Guan, F.Q.; Sun, H.; Ren, X.C.; Chen, Y.; Feng, X.; Yang, J.M. Triticuside A, a dietary flavonoid, inhibits proliferation of human breast cancer cells via inducing apoptosis. Nutr. Cancer 2013, 65, 891–899. [Google Scholar] [CrossRef]
- Eitsuka, T.; Tatewaki, N.; Nishida, H.; Kurata, T.; Nakagawa, K.; Miyazawa, T. Synergistic inhibition of cancer cell proliferation with a combination of delta-tocotrienol and ferulic acid. Biochem. Biophys. Res. Commun. 2014, 453, 606–611. [Google Scholar] [CrossRef]
- Ghoneum, M.; El-Din, N.K.B.; Ali, D.A.; El-Dein, M.A. Modified Arabinoxylan from Rice Bran, MGN-3Biobran Sensitizes Metastatic Breast Cancer Cells to Paclitaxel In Vitro. Anticancer Res. 2014, 34, 81–88. [Google Scholar]
- Pintha, K.; Yodkeeree, S.; Pitchakarn, P.; Limtrakul, P. Anti-invasive Activity against Cancer Cells of Phytochemicals in Red Jasmine Rice (Oryza sativa L.). Asian Pac. J. Cancer Prev. 2014, 15, 4601–4607. [Google Scholar] [CrossRef]
- Kannan, A.; Hettiarachchy, N.S.; Lay, J.O.; Liyanage, R. Human cancer cell proliferation inhibition by a pentapeptide isolated and characterized from rice bran. Peptides 2010, 31, 1629–1634. [Google Scholar] [CrossRef]
- Park, J.H.; Darvin, P.; Lim, E.J.; Joung, Y.H.; Hong, D.Y.; Park, E.U.; Park, S.H.; Choi, S.K.; Moon, E.S.; Cho, B.W.; et al. Hwanggeumchal sorghum induces cell cycle arrest, and suppresses tumor growth and metastasis through Jak2/STAT pathways in breast cancer xenografts. PLoS ONE 2012, 7, e40531. [Google Scholar] [CrossRef]
- Suganyadevi, P.; Saravanakumar, K.M.; Mohandas, S. Evaluation of Antiproliferative Activity of Red Sorghum Bran Anthocyanin on a Human Breast Cancer Cell Line (MCF-7). Int. J. Breast Cancer 2011, 2011, 891481. [Google Scholar] [CrossRef]
- Suganyadevi, P.; Saravanakumar, K.M.; Mohandas, S. The antiproliferative activity of 3-deoxyanthocyanins extracted from red sorghum (Sorghum bicolor) bran through P53-dependent and Bcl-2 gene expression in breast cancer cell line. Life Sci. 2013, 92, 379–382. [Google Scholar] [CrossRef]
- Luo, L.P.; Han, B.; Yu, X.P.; Chen, X.Y.; Zhou, J.; Chen, W.; Zhu, Y.F.; Peng, X.L.; Zou, Q.; Li, S.Y. Anti-metastasis Activity of Black Rice Anthocyanins Against Breast Cancer: Analyses Using an ErbB2 Positive Breast Cancer Cell Line and Tumoral Xenograft Model. Asian Pac. J. Cancer Prev. 2014, 15, 6219–6225. [Google Scholar] [CrossRef] [Green Version]
- Bonjean, A.P.; Angus, W.J. The World Wheat Book: A History of Wheat Breeding; Lavoisier Publishing: Paris, France, 2001; p. 1131. [Google Scholar]
- Landberg, R.; Marklund, M.; Kamal-Eldin, A.; Aman, P. An update on alkylresorcinols—Occurrence, bioavailability, bioactivity and utility as biomarkers. J. Funct. Foods 2014, 7, 77–89. [Google Scholar] [CrossRef]
- Biskup, I.; Kyro, C.; Marklund, M.; Olsen, A.; van Dam, R.M.; Tjonneland, A.; Overvad, K.; Lindahl, B.; Johansson, I.; Landberg, R. Plasma alkylresorcinols, biomarkers of whole-grain wheat and rye intake, and risk of type 2 diabetes in Scandinavian men and women. Am. J. Clin. Nutr. 2016, 104, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Kyro, C.; Olsen, A.; Bueno-de-Mesquita, H.B.; Skeie, G.; Loft, S.; Aman, P.; Leenders, M.; Dik, V.K.; Siersema, P.D.; Pischon, T.; et al. Plasma alkylresorcinol concentrations, biomarkers of whole-grain wheat and rye intake, in the European Prospective Investigation into Cancer and Nutrition (EPIC) cohort. Br. J. Nutr. 2014, 111, 1881–1890. [Google Scholar] [CrossRef] [Green Version]
- Gliwa, J.; Gunenc, A.; Ames, N.; Willmore, W.G.; Hosseinian, F.S. Antioxidant activity of alkylresorcinols from rye bran and their protective effects on cell viability of PC-12 AC cells. J. Agric. Food Chem. 2011, 59, 11473–11482. [Google Scholar] [CrossRef]
- Zhu, Y.; Soroka, D.N.; Sang, S. Synthesis and inhibitory activities against colon cancer cell growth and proteasome of alkylresorcinols. J. Agric. Food Chem. 2012, 60, 8624–8631. [Google Scholar] [CrossRef]
- Andersson, U.; Dey, E.S.; Holm, C.; Degerman, E. Rye bran alkylresorcinols suppress adipocyte lipolysis and hormone-sensitive lipase activity. Mol. Nutr. Food Res. 2011, 55 (Suppl. 2), S290–S293. [Google Scholar] [CrossRef] [PubMed]
- Oishi, K.; Yamamoto, S.; Itoh, N.; Nakao, R.; Yasumoto, Y.; Tanaka, K.; Kikuchi, Y.; Fukudome, S.-I.; Okita, K.; Takano-Ishikawa, Y. Wheat Alkylresorcinols Suppress High-Fat, High-Sucrose Diet-Induced Obesity and Glucose Intolerance by Increasing Insulin Sensitivity and Cholesterol Excretion in Male Mice. J. Nutr. 2015, 145, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Winter, K.M.; Stevenson, L.; Morris, C.; Leach, D.N. Wheat bran lipophilic compounds with in vitro anticancer effects. Food Chem. 2012, 130, 156–164. [Google Scholar] [CrossRef]
- Liu, J.; Hao, Y.; Wang, Z.; Ni, F.; Wang, Y.; Gong, L.; Sun, B.; Wang, J. Identification, Quantification, and Anti-inflammatory Activity of 5- n-Alkylresorcinols from 21 Different Wheat Varieties. J. Agric. Food Chem. 2018, 66, 9241–9247. [Google Scholar] [CrossRef] [PubMed]
- Kruk, J.; Aboul-Enein, B.; Bernstein, J.; Marchlewicz, M. Dietary alkylresorcinols and cancer prevention: A systematic review. Eur. Food Res. Technol. 2017, 243, 1693–1710. [Google Scholar] [CrossRef]
- Sanchez, L.A.; Olmedo, D.; Luis Lopez-Perez, J.; Williams, T.D.; Gupta, M.P. Two New Alkylresorcinols from Homalomena wendlandii and Their Cytotoxic Activity. Nat. Prod. Commun. 2012, 7, 1043–1046. [Google Scholar] [CrossRef]
- Lee, C.I.; Lee, C.L.; Hwang, J.F.; Lee, Y.H.; Wang, J.J. Monascus-fermented red mold rice exhibits cytotoxic effect and induces apoptosis on human breast cancer cells. Appl. Microbiol. Biotechnol. 2013, 97, 1269–1278. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Colletti, A. Potential role of bioactive peptides in prevention and treatment of chronic diseases: A narrative review. Br. J. Pharmacol. 2017, 174, 1378–1394. [Google Scholar] [CrossRef]
- Ortiz-Martinez, M.; Winkler, R.; Garcia-Lara, S. Preventive and therapeutic potential of peptides from cereals against cancer. J. Proteomics 2014, 111, 165–183. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W. Sorghum phytochemicals and their potential impact on human health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef]
- Patil, J.V. Millets and Sorghum: Biology and Genetic Improvement; Wiley-Blackwell Publishing: Hoboken, NJ, USA, 2016. [Google Scholar]
- Yang, L.; Browning, J.D.; Awika, J.M. Sorghum 3-Deoxyanthocyanins Possess Strong Phase II Enzyme Inducer Activity and Cancer Cell Growth Inhibition Properties. J. Agric. Food Chem. 2009, 57, 1797–1804. [Google Scholar] [CrossRef]
- Shih, C.H.; Siu, S.O.; Ng, R.; Wong, E.; Chiu, L.C.M.; Chu, I.K.; Lo, C. Quantitative analysis of anticancer 3-deoxyanthocyanidins in infected sorghum seedlings. J. Agric. Food Chem. 2007, 55, 254–259. [Google Scholar] [CrossRef]
- Hargrove, J.L.; Greenspan, P.; Hartle, D.K.; Dowd, C. Inhibition of Aromatase and alpha-Amylase by Flavonoids and Proanthocyanidins from Sorghum bicolor Bran Extracts. J. Med. Food 2011, 14, 799–807. [Google Scholar] [CrossRef]
- Choromanska, A.; Kulbacka, J.; Rembialkowska, N.; Pilat, J.; Oledzki, R.; Harasym, J.; Saczko, J. Anticancer properties of low molecular weight oat beta-glucan—An in vitro study. Int. J. Biol. Macromol. 2015, 80, 23–28. [Google Scholar] [CrossRef]
- Boffetta, P.; Thies, F.; Kris-Etherton, P. Epidemiological studies of oats consumption and risk of cancer and overall mortality. Br. J. Nutr. 2014, 112 (Suppl. 2), S14–S18. [Google Scholar] [CrossRef] [Green Version]
- Demir, G.; Klein, H.O.; Mandel-Molinas, N.; Tuzuner, N. Beta glucan induces proliferation and activation of monocytes in peripheral blood of patients with advanced breast cancer. Int. Immunopharmacol. 2007, 7, 113–116. [Google Scholar] [CrossRef]
- Yang, J.; Wang, P.; Wu, W.; Zhao, Y.; Idehen, E.; Sang, S. Steroidal Saponins in Oat Bran. J. Agric. Food Chem. 2016, 64, 1549–1556. [Google Scholar] [CrossRef]
- Bar-Sela, G.; Tsalic, M.; Fried, G.; Goldberg, H. Wheat grass juice may improve hematological toxicity related to chemotherapy in breast cancer patients: A pilot study. Nutr. Cancer 2007, 58, 43–48. [Google Scholar] [CrossRef]
- Gollapudi, S.; Ghoneum, M. MGN-3/Biobran, modified arabinoxylan from rice bran, sensitizes human breast cancer cells to chemotherapeutic agent, daunorubicin. Cancer Detect. Prev. 2008, 32, 1–6. [Google Scholar] [CrossRef]
- Serra-Majem, L.; Bautista-Castano, I. Relationship between bread and obesity. Br. J. Nutr. 2015, 113 (Suppl. 2), S29–S35. [Google Scholar] [CrossRef]
- Cioffi, I.; Ibrugger, S.; Bache, J.; Thomassen, M.T.; Contaldo, F.; Pasanisi, F.; Kristensen, M. Effects on satiation, satiety and food intake of wholegrain and refined grain pasta. Appetite 2016, 107, 152–158. [Google Scholar] [CrossRef]
- Ullrich, I.H.; Albrink, M.J. The effect of dietary fiber and other factors on insulin response: Role in obesity. J. Environ. Pathol. Toxicol. Oncol. 1985, 5, 137–155. [Google Scholar]
- Hutter, C.; Zenklusen, J.C. The Cancer Genome Atlas: Creating Lasting Value beyond Its Data. Cell 2018, 173, 283–285. [Google Scholar] [CrossRef]
- Hoadley, K.A.; Yau, C.; Hinoue, T.; Wolf, D.M.; Lazar, A.J.; Drill, E.; Shen, R.; Taylor, A.M.; Cherniack, A.D.; Thorsson, V.; et al. Cell-of-Origin Patterns Dominate the Molecular Classification of 10,000 Tumors from 33 Types of Cancer. Cell 2018, 173, 291–304.e296. [Google Scholar] [CrossRef]
Figure 1.
Major bioactive phytochemicals derived from whole grains.

Figure 2.
Schematic overview of molecular-targeted mechanisms of phytochemicals derived from whole grains against breast cancer.
Figure 2.
Schematic overview of molecular-targeted mechanisms of phytochemicals derived from whole grains against breast cancer.


{kind=link}
{kind=link}
| Molecular Subtype | IHC Marker (ER/PR/HER2) | Frequency (%) | Proliferation Cluster | Gene Markers | Histologic Grade | Prognosis |
|---|---|---|---|---|---|---|
| Luminal A | ER+ PR+ HER-2- | 50–60 | Low | ESR1, GATA3, KRT8, XBP1, FOXA1, TFF3, CCND1, LIV1 | Low | Excellent |
| Luminal B | ER+ PR+/- HER-2+ | 10–20 | High | ESR1, GATA3, KRT8, XBP1, FOXA1, TFF3, SQLE, LAPTM4B | Intermediate/High | Intermediate/Bad |
| Basal-like | ER- PR-/+ HER-2-/+ | 10–20 | High | KRT5, CDH3, ID4, FABP7, KRT17, LAMC2, TRIM29 | High | Bad |
| HER2-enriched | ER-/+ PR-/+ HER-2+ | 10–15 | High | ERBB2, GRB7 | High | Bad |
| Normal breast-like | ER+/- PR+/- HER-2- | 5–10 | Low | VIM, MMP2/14, COL3A1, TIMP1, CD36, FABP4, ITGA7 | Low | Intermediate |
| Claudin-low | ER- PR- HER-2-/+ | 12–14 | High | CD24(-), CD44(+) | High | Bad |
IHC: immunohistochemistry; ER: estrogen receptor; PR: progesterone receptor; +: positive; –: negative; +/-: predominantly positive; -/+: predominantly negative.
Table 2.
The potential health benefits of bioactive phytochemicals extracted from whole grains.
| Bioactive Phytochemicals | Major Sources | Potential Health Benefits | References |
|---|---|---|---|
| Alkylresorcinols | Wheat, rye | Cancer prevention; obesity reduction | [64,65] |
| Avenanthramide | Oat | Neutralizing free radicals, cancer prevention | [66,67] |
| Phenolics | |||
| Anthocyanins | Barley, rice, sorghum | Neutralizing free radicals; inflammatory inhibition; cancer prevention | [40,45,68,69,70,71] |
| Lignans | Wheat, rye | Cancer prevention; hormone modulation; reducing the risk of cardiovascular disease | [72,73] |
| Flavones | Rye, barley, sorghum | Neutralizing free radicals; cancer prevention. | [45,68,69,74] |
| Tannins | Barley, sorghum | Improve urinary tract health; reducing risk of cardiovascular disease | [45,68] |
| Carotenoids | |||
| α-carotene/β-carotene | Wheat, barley, millet | Neutralizing free radicals; reducing heart disease risks | [61,75] |
| Phytosterols | |||
| Sterols | Wheat, barley, oat | Lowering blood cholesterol levels; reducing lipid accumulation; cancer prevention; reducing cardiovascular disease risks | [66,76,77] |
| Stanols | Wheat, maize, barley | Lowering blood cholesterol levels; reducing lipid accumulation; cancer prevention; reducing cardiovascular disease risks | [66,76,77] |
| Non-starchy Polysaccharide | |||
| Insoluble dietary fiber | Wheat | Cancer prevention; lowering plasma cholesterol; reducing insulin resistance level | [75,78] |
| β-Glucans | Oat, barley | Reducing the risk of cardiovascular disease; lowering the level of low-density lipoprotein and total cholesterol, cancer prevention | [45,66,67,79] |
| Tocols | |||
| Tocopherols | Barley, oat | Inhibiting lipid peroxidation; reducing the risk of cardiovascular disease; reducing stroke risks | [66,80] |
| Tocotrienols | Barley, oat | Inhibiting lipid peroxidation; reducing the risk of cardiovascular disease; reducing stroke risks | [66,80] |
Table 3.
Epidemiological and clinical evidences of breast cancer and whole grain intakes.
| Natural Product (diet) | Study Type | Case/Participants | OR/RR (95%CI) | Conclusion | Reference |
|---|---|---|---|---|---|
| Whole grain | Meta-analysis of cohort and case-control studies | 11,589/131,151 (4 cohort and 7 case-control studies) | Summary RR: 0.84 (0.74–0.96, P= 0.009, I2 = 63.8%) | High intake of whole grains might be inversely associated with reduced breast cancer risks, but the inverse association was only observed in case-control not cohort studies. | [5] |
| Cereal dietary fiber | Meta-analysis of perspective studies | 14,694/502,082 (six prospective studies) | Summary RR: 0.96 (0.90–1.02, I2 = 5%) | Cereal dietary fibers have an inverse association with breast cancer risk. | [55] |
| Dietary fiber | Meta-analysis of perspective studies | 16,848/712,195 (10 prospective cohort studies) | Summary RR: 0.89 (0.83–0.96, I2 = 0%) | There was a significant inverse dose-response association between dietary fiber intake and breast cancer risk. | [84] |
| Whole grain bread | Case-controlled study | 306/309 | OR:0.61 (0.37–0.99) | Resistant starch containing foods (whole grain wheat bread) may reduce breast cancer risk. | [83] |
| Whole grains | Case-controlled study | 250/250 | OR:0.49 (0.29–0.82) | Whole grain consumption more than 7 times/week was associated with reduced risk of breast cancer. | [6] |
| Dietary Fiber | Prospective cohort study | 2833/90534 (Follow-up: 20 years) | RR: 0.84(0.70–1.01; Ptrend = 0.04) | Higher fiber intakes during adolescence and early adulthood could reduce breast cancer risk. | [81] |
| Whole grain contained food | Prospective cohort study | 3235/90516 (Follow-up: 22 years) | RR: 0.82(0.70–0.97; Ptrend = 0.03) | High whole grain food intake may be associated with lower breast cancer risk before menopause. | [82] |
| Whole and refined grain food | Prospective cohort study | 124/3184 (Follow-up: 22 years) | HR: 0.53(0.33–0.86) | Higher consumption of whole grain food may protect against breast cancer, with 47% lower breast cancer risk. | [56] |
OR: Odds ratios; RR: Relative Risks; HR: hazard ratios; CI: confidence intervals.
Table 4.
Summary of health benefits of whole grains (and/or its components) on breast cancer.
| Source | Constituents | Study Model (Cell Lines/Animal) | Mechanism | Reference |
|---|---|---|---|---|
| in vitro | ||||
| Wheat | Germinated wheat flour | Human breast cancer ER-positive MCF-7& TNBC MDA-MB-231 | Up-regulation of apoptosis | [91] |
| Barley | Young barley and its methanolic extract | Human breast cancer MCF-7 | Up-regulation of apoptosis, through lower metabolic activity, inhibition of proliferation, and cell cycle arrest in S phase | [92] |
| Foxtail millet | Total phenolic extracts | Human breast cancer MDA-MB-231 | Proliferation inhibition | [61] |
| Synthetic analog of oat avenanthramide | Dihydroavenanthramide D | Human breast cancer MCF-7 | Cancer cell invasion inhibition through the down regulation of MMP-9 activity and suppression of MAPK/NF-κB and MAPK/AP-1 pathway | [93] |
| Oat | Avenanthramide-C | Human breast cancer MDA-MB-231 | Activation of apoptosis and caspases activity, positive annexin V staining and cell cycle arrest in sub G1 indicating DNA fragmentation | [94] |
| Wheat bran | Triticuside A | Human breast cancer MCF-7& MDA-MB-231 | Activation of mitochondrial apoptosis pathway and Akt/mTOR signaling pathway, with downregulation of Mcl-1 and Bcl-2 and increase of cleavage of caspases-3, -7, -9, and PARP. Level of phospho-Akt and its downstream targets, mTOR, and P70 S6 kinase are also decreased | [95] |
| Rice bran | δ-Tocotrienol and Ferulic acid | Human breast cancer MCF-7 | δ-tocotrienol and ferulic acid co-use synergistically inhibit cancer cell proliferation and induced cell arrest in the G1 phase | [96] |
| Rice bran | Arabinoxylan | Human breast cancer MCF-7; murine metastatic breast cancer 4T-1 | Arabinoxylan increased the susceptibility of both types of cancer cells to paclitaxel by causing DNA damage, enhancing apoptosis, and inhibiting cell proliferation | [97] |
| Red rice bran | crude ethanolic extract of red rice bran | Human breast cancer MDA-MB-231 | Decreased the secretion and activity of MMP-2 and MMP-9 reducing cells invasion | [98] |
| Rice bran | Glu-Gln-Arg-Pro-Arg | Human breast cancer MCF-7& MDA-MB-231 | Anti-proliferation activity | [99] |
| Sorghum | Total sorghum extracts | Human breast cancer MCF-7, MDA-MB-231 & HER-2+/ER-SKBR-3 | G1 phase arrest Down-regulation of the STAT5/IGF-1R and STAT3/VEGF pathway | [100] |
| Red sorghum bran | Anthocyanins | Human breast cancer MCF-7 | Anti-proliferation activity | [101] |
| Red sorghum bran | 3-Deoxyanthocyanins | Human breast cancer MCF-7 | Anti-proliferation activity P53 gene up-regulation; bcl-2 gene down-regulation | [102] |
| in vivo | ||||
| Sorghum (Hwanggeumchal sorghum) | Total sorghum extracts | BALB/c nude mice | Breast cancer tumor suppression; down-regulation of STAT5b/IGF-1R and STAT3/VEGF signal pathways; breast-to-lung metastasis blockage | [100] |
| Black rice | Anthocyanins | BALB/c nude mice | Decreased activity of urokinase-type plasminogen activator (u-PA), and reduced transplanted tumor growth and inhibited pulmonary | [103] |
| Barley | Young barley | Sprague-Dawley female rats | Decrease in tumor incidence and average tumor volume; Caspase-3/caspase-7 increased; Ki67 decreased | [92] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xie, M.; Liu, J.; Tsao, R.; Wang, Z.; Sun, B.; Wang, J. Whole Grain Consumption for the Prevention and Treatment of Breast Cancer. Nutrients 2019, 11, 1769. https://doi.org/10.3390/nu11081769
AMA Style
Xie M, Liu J, Tsao R, Wang Z, Sun B, Wang J. Whole Grain Consumption for the Prevention and Treatment of Breast Cancer. Nutrients. 2019; 11(8):1769. https://doi.org/10.3390/nu11081769
Chicago/Turabian StyleXie, Mingsi, Jie Liu, Rong Tsao, Ziyuan Wang, Baoguo Sun, and Jing Wang. 2019. "Whole Grain Consumption for the Prevention and Treatment of Breast Cancer" Nutrients 11, no. 8: 1769. https://doi.org/10.3390/nu11081769
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.