Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model


Abstract
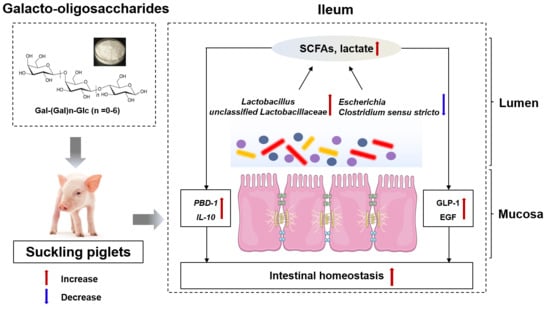
:
1. Introduction
2. Materials and Methods
2.1. Animal Trial
2.2. Microbiota Profiling
2.3. Microbial Metabolites
2.4. RT-PCR
2.5. Endocrine Peptides
2.6. Statistical Analysis
3. Results
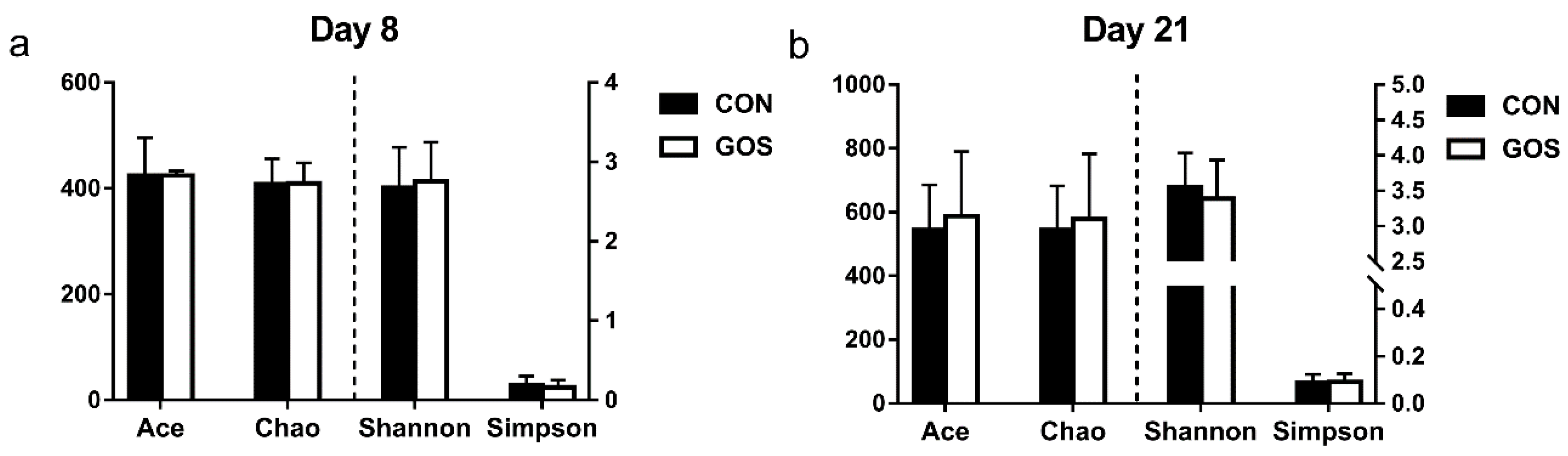
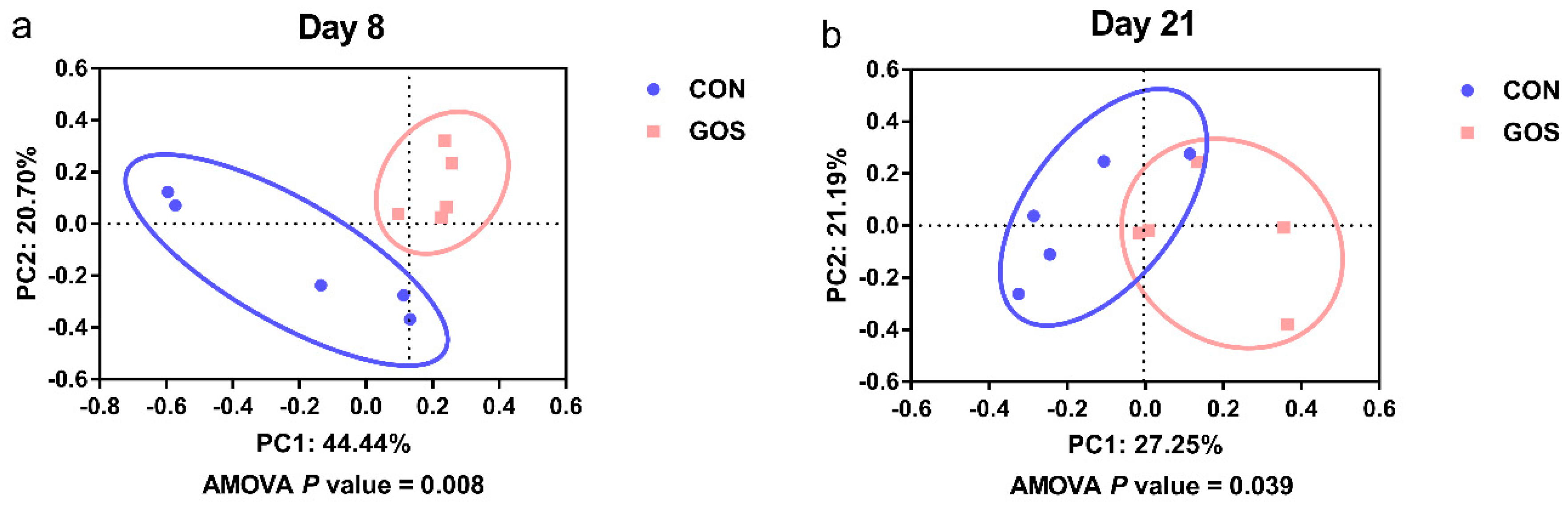
3.1. Effects of the Early-Life GOS Intervention on the Diversity of the Ileal Digesta Microbiota
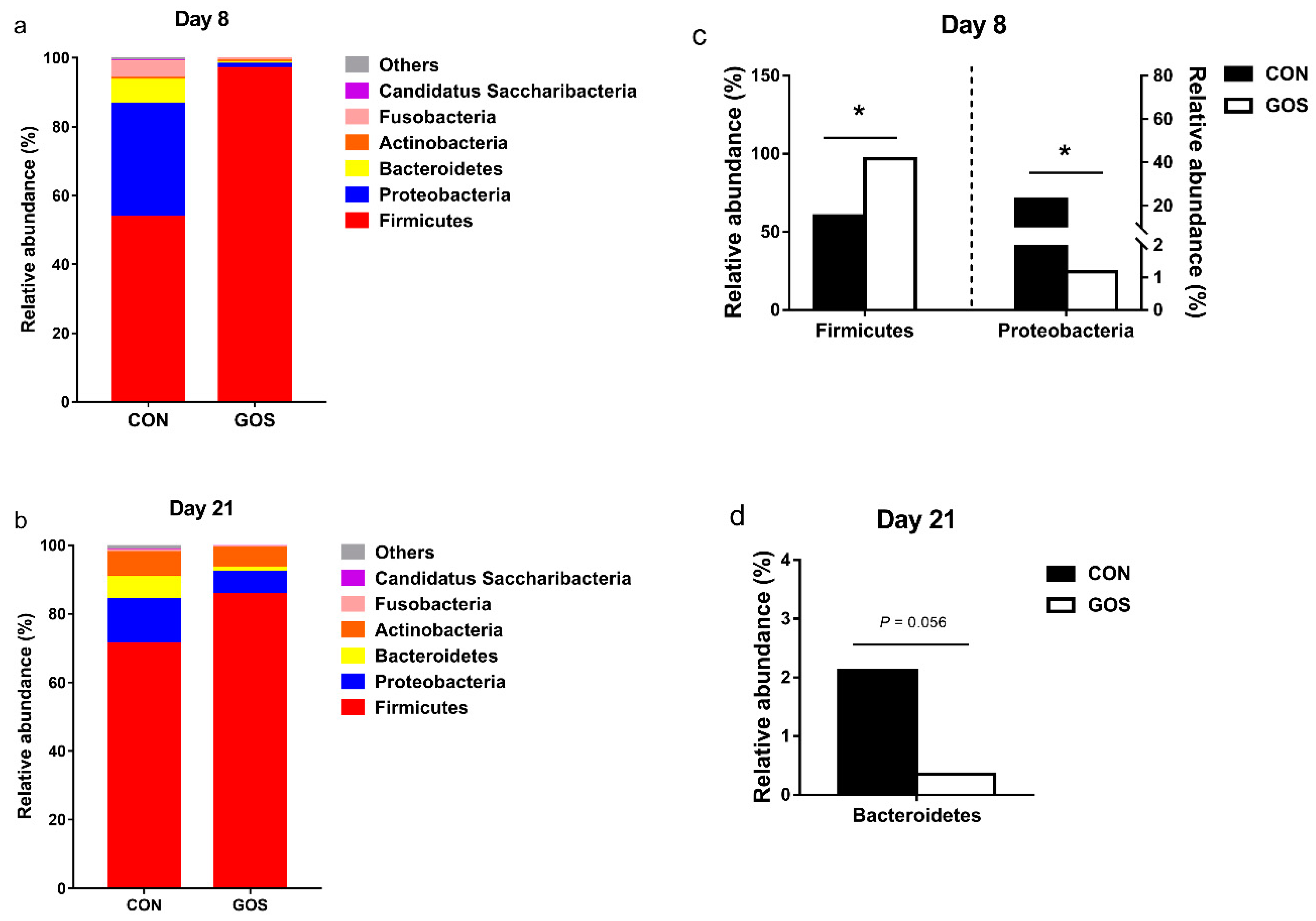
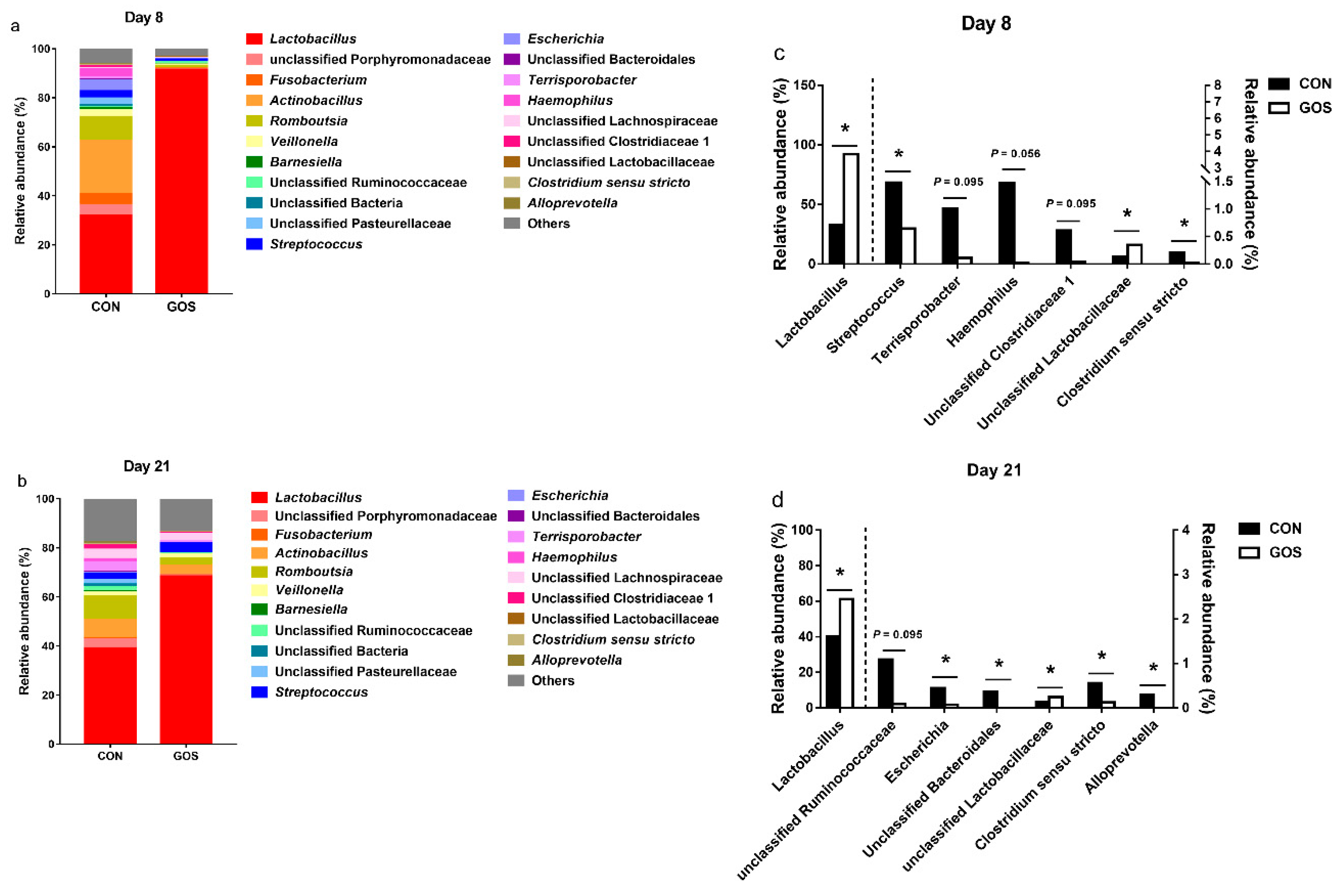
3.2. Effects of the Early-Life GOS Intervention on the Abundance of Ileal Microbiota
3.3. Effects of the Early-Life GOS Intervention on pH Value, SCFAs, and Lactate Concentrations in Ileal Digesta
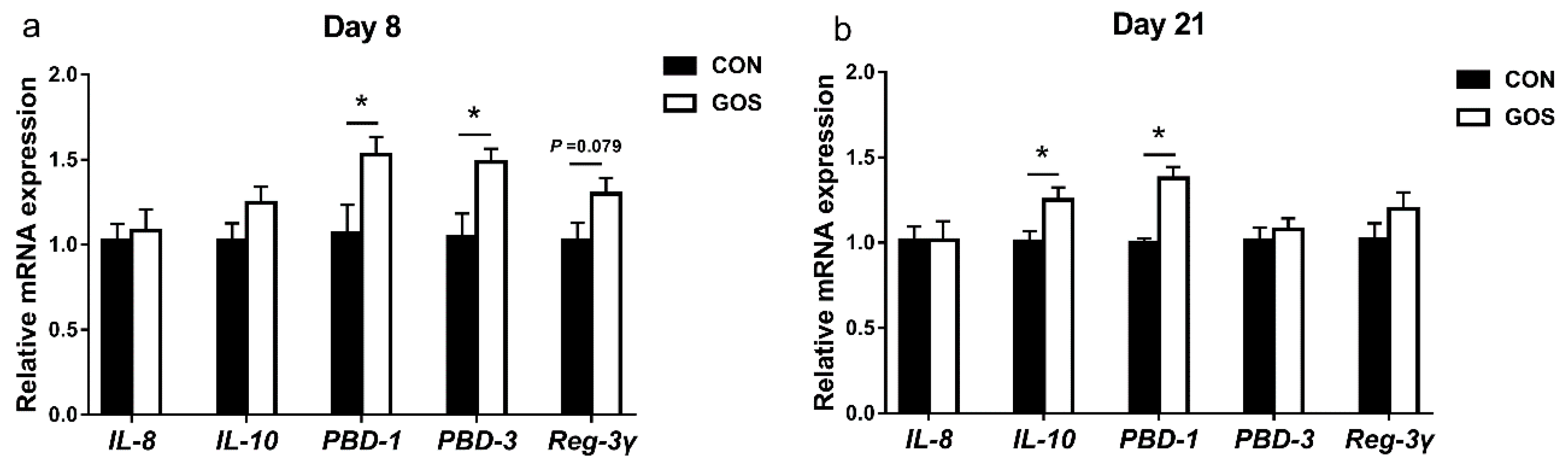
3.4. Effects of the Early-Life GOS Intervention on the mRNA Expression of Cytokines and Antimicrobial Peptide in the Ileal Mucosa
3.5. Effects of the Early-Life GOS Intervention on the Concentration of Endocrine Peptides in the Ileal Mucosa
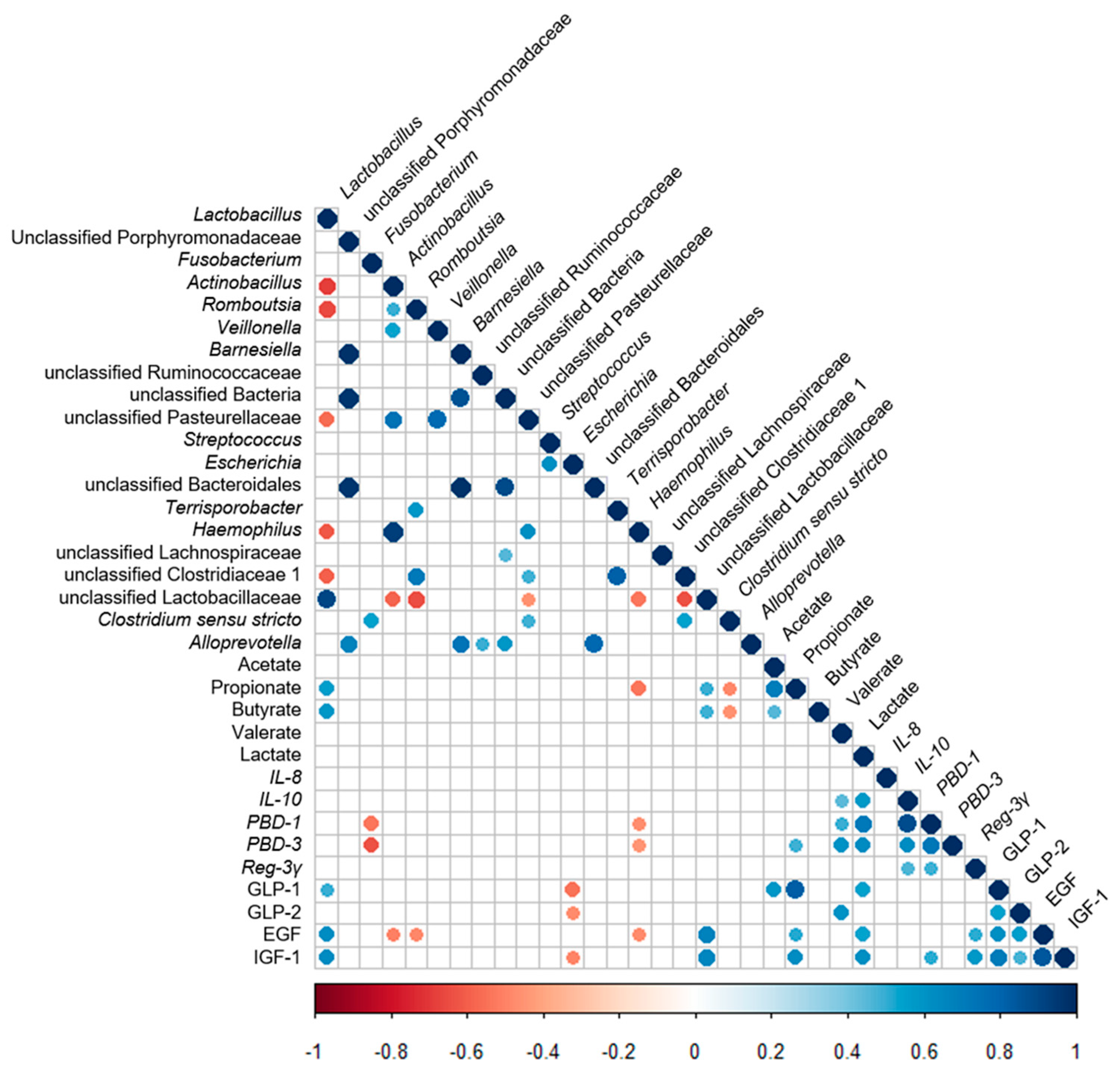
3.6. Correlation Analysis between the Ileal Microbiota and the Metabolites, the Ileal Cytokines’ Expression, Antimicrobial Peptides’ Expression, and Ileal Endocrine Peptides Levels
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martin, R.; Nauta, A.J.; Ben Amor, K.; Knippels, L.M.; Knol, J.; Garssen, J. Early life: Gut microbiota and immune development in infancy. Benef. Microbes 2010, 1, 367–382. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; MacPherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Romick-Rosendale, L.E.; Goodpaster, A.M.; Hanwright, P.J.; Patel, N.B.; Wheeler, E.T.; Chona, D.L.; Kennedy, M.A.; Romick-Rosendale, L.E.; Romick-Rosendale, L.E. NMR-based metabonomics analysis of mouse urine and fecal extracts following oral treatment with the broad-spectrum antibiotic enrofloxacin (Baytril). Magn. Reson. Chem. 2009, 47, S36–S46. [Google Scholar] [CrossRef]
- Yu, M.; Mu, C.; Zhang, C.; Yang, Y.; Su, Y.; Zhu, W. Marked Response in Microbial Community and Metabolism in the Ileum and Cecum of Suckling Piglets After Early Antibiotics Exposure. Front. Microbiol. 2018, 9, 1166. [Google Scholar] [CrossRef]
- Schokker, D.; Zhang, J.; Zhang, L.-L.; Vastenhouw, S.A.; Heilig, H.G.H.J.; Smidt, H.; Rebel, J.M.J.; Smits, M.A. Early-Life Environmental Variation Affects Intestinal Microbiota and Immune Development in New-Born Piglets. PLoS ONE 2014, 9, 100040. [Google Scholar] [CrossRef]
- Lewis, M.C.; Inman, C.F.; Patel, D.; Schmidt, B.; Mulder, I.; Miller, B.; Gill, B.P.; Pluske, J.; Kelly, D.; Stokes, C.R.; et al. Direct experimental evidence that early-life farm environment influences regulation of immune responses. Pediatr. Allergy Immunol. 2012, 23, 265–269. [Google Scholar] [CrossRef]
- Schachtschneider, K.M.; Yeoman, C.J.; Isaacson, R.E.; White, B.A.; Schook, L.B.; Pieters, M. Modulation of Systemic Immune Responses through Commensal Gastrointestinal Microbiota. PLoS ONE 2013, 8, e53969. [Google Scholar] [CrossRef]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut Immune Maturation Depends on Colonization with a Host-Specific Microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef] [Green Version]
- Konstantinov, S.R.; Awati, A.A.; Williams, B.A.; Miller, B.G.; Jones, P.; Stokes, C.R.; Akkermans, A.D.L.; Smidt, H.; De Vos, W.M. Post-natal development of the porcine microbiota composition and activities. Environ. Microbiol. 2006, 8, 1191–1199. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Zakharova, I.; Dmitrieva, Y. Oligosaccharides in infant formula: More evidence to validate the role of prebiotics. Br. J. Nutr. 2015, 113, 1339–1344. [Google Scholar] [CrossRef]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [Green Version]
- Hernot, D.C.; Boileau, T.W.; Bauer, L.L.; Middelbos, I.S.; Murphy, M.R.; Swanson, K.S.; Fahey, G.C., Jr. In vitro fermentation profiles, gas production rates, and microbiota modulation as affected by certain fructans, galactooligosaccharides, and polydextrose. J. Agric. Food Chem. 2009, 57, 1354–1361. [Google Scholar] [CrossRef]
- Conlon, M.; Bird, A. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2015, 7, 17–44. [Google Scholar] [CrossRef]
- Tzortzis, G. Functional properties of the second generation prebiotic Galacto-oligosaccharide (B-GOS). Agro Food Ind. Hi-Tech 2009, 20, 43–46. [Google Scholar]
- Monteagudo-Mera, A.; Arthur, J.; Jobin, C.; Keku, T.; Bruno-Barcena, J.; Azcarate-Peril, M. High purity galacto-oligosaccharides (GOS) enhance specific Bifidobacterium species and their metabolic activity in the mouse gut microbiome. Benef. Microbes 2016, 7, 247–264. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Walton, G.E.; Claus, S.P.; Tzortzis, G.; Toward, R.E.; Gibson, G.R. Influence of galacto-oligosaccharide mixture (B-GOS) on gut microbiota, immune parameters and metabonomics in elderly persons. Br. J. Nutr. 2015, 114, 586–595. [Google Scholar] [CrossRef]
- Sharma, S.; Puri, S. Prebiotics and lipid metabolism: A review. Altern. Ther. Health Med. 2015, 21, 34–42. [Google Scholar]
- Scholz-Ahrens, K.E.; Adolphi, B.; Rochat, F.; Barclay, D.V.; de Vrese, M.; Açil, Y.; Schrezenmeir, J. Effects of probiotics, prebiotics, and synbiotics on mineral metabolism in ovariectomized rats—Impact of bacterial mass, intestinal absorptive area and reduction of bone turn-over. NFS J. 2016, 3, 41–50. [Google Scholar] [CrossRef]
- Heinritz, S.N.; Mosenthin, R.; Weiss, E. Use of pigs as a potential model for research into dietary modulation of the human gut microbiota. Nutr. Res. Rev. 2013, 26, 191–209. [Google Scholar] [CrossRef] [Green Version]
- Villmones, H.C.; Haug, E.S.; Ulvestad, E.; Grude, N.; Stenstad, T.; Halland, A. Kommedal, Øyvind Species Level Description of the Human Ileal Bacterial Microbiota. Sci. Rep. 2018, 8, 4736. [Google Scholar] [CrossRef]
- Anthony, J.C.; Merriman, T.N.; Heimbach, J.T. 90-Day oral (gavage) study in rats with galactooligosaccharides syrup. Food Chem. Toxicol. 2006, 44, 819–826. [Google Scholar] [CrossRef]
- Matsuki, T.; Tajima, S.; Hara, T.; Yahagi, K.; Ogawa, E.; Kodama, H. Infant formula with galacto-oligosaccharides (OM55N) stimulates the growth of indigenous bifidobacteria in healthy term infants. Benef. Microbes 2016, 7, 453–461. [Google Scholar] [CrossRef]
- Pan, X.-D.; Chen, F.-Q.; Wu, T.-X.; Tang, H.-G.; Zhao, Z.-Y. Prebiotic oligosaccharides change the concentrations of short-chain fatty acids and the microbial population of mouse bowel. J. Zhejiang Univ. Sci. B 2009, 10, 258. [Google Scholar] [CrossRef]
- Zoetendal, E.G.; Akkermans, A.D.L.; De Vos, W.M. Temperature Gradient Gel Electrophoresis Analysis of 16S rRNA from Human Fecal Samples Reveals Stable and Host-Specific Communities of Active Bacteria. Appl. Environ. Microbiol. 1998, 64, 3854–3859. [Google Scholar] [Green Version]
- Shi, C.; Zhu, Y.; Niu, Q.; Wang, J.; Wang, J.; Zhu, W. The Changes of Colonic Bacterial Composition and Bacterial Metabolism Induced by an Early Food Introduction in a Neonatal Porcine Model. Curr. Microbiol. 2018, 75, 745–751. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tian, S.; Yu, H.; Wang, J.; Zhu, W. Response of Colonic Mucosa-Associated Microbiota Composition, Mucosal Immune Homeostasis, and Barrier Function to Early Life Galactooligosaccharides Intervention in Suckling Piglets. J. Agric. Food Chem. 2018, 67, 578–588. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tian, S.; Wang, J.; Yu, H.; Wang, J.; Zhu, W. Effects of galacto-oligosaccharides on growth and gut function of newborn suckling piglets. J. Anim. Sci. Biotechnol. 2018, 9, 75. [Google Scholar] [CrossRef]
- Cheng, W.; Lu, J.; Lin, W.; Wei, X.; Li, H.; Zhao, X.; Jiang, A.; Yuan, J. Effects of a galacto-oligosaccharide-rich diet on fecal microbiota and metabolite profiles in mice. Food Funct. 2018, 9, 1612–1620. [Google Scholar] [CrossRef]
- Wang, L.; Hu, L.; Yan, S.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Effects of different oligosaccharides at various dosages on the composition of gut microbiota and short-chain fatty acids in mice with constipation. Food Funct. 2017, 8, 1966–1978. [Google Scholar] [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimarăes, V.; Sokol, H.; Doré, J.; Corthier, G.; Furet, J.-P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Angelis, M.; Calasso, M.; Cavallo, N.; Di Cagno, R.; Gobbetti, M. Functional proteomics within the genus Lactobacillus. Proteomics 2016, 16, 946–962. [Google Scholar] [CrossRef]
- Valeriano, V.; Balolong, M.; Kang, D. Probiotic roles of Lactobacillus sp. in swine: Insights from gut microbiota. J. Appl. Microbiol. 2017, 122, 554–567. [Google Scholar] [CrossRef]
- Kenny, M.; Smidt, H.; Mengheri, E.; Miller, B. Probiotics—Do they have a role in the pig industry? Animal 2011, 5, 462–470. [Google Scholar] [CrossRef]
- Yang, Y.; Zhao, X.; Le, M.H.A.; Zijlstra, R.T.; Gänzle, M.G. Reutericyclin producing Lactobacillus reuteri modulates development of fecal microbiota in weanling pigs. Front. Microbiol. 2015, 6, 762. [Google Scholar] [CrossRef] [Green Version]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef]
- Li, K.; Xiao, Y.; Chen, J.; Chen, J.; He, X.; Yang, H. Microbial composition in different gut locations of weaning piglets receiving antibiotics. Asian Australas. J. Anim. Sci. 2017, 30, 78. [Google Scholar] [CrossRef]
- Downes, J.; Dewhirst, F.E.; Tanner, A.C.; Wade, W.G. Description of Alloprevotella rava gen. nov., sp. nov., isolated from the human oral cavity, and reclassification of Prevotella tannerae Moore et al. 1994 as Alloprevotella tannerae gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2013, 63, 1214. [Google Scholar] [CrossRef]
- Ren, W.; Chen, S.; Zhang, L.; Liu, G.; Hussain, T.; Hao, X.; Yin, J.; Duan, J.; Tan, B.; Wu, G.; et al. Interferon Tau Affects Mouse Intestinal Microbiota and Expression of IL-17. Mediat. Inflamm. 2016, 2016, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci. Rep. 2017, 7, 43412. [Google Scholar] [CrossRef]
- Zhou, L.; Fang, L.; Sun, Y.; Su, Y.; Zhu, W. Effects of the dietary protein level on the microbial composition and metabolomic profile in the hindgut of the pig. Anaerobe 2016, 38, 61–69. [Google Scholar] [CrossRef]
- Bovera, F.; Marono, S.; Di Meo, C.; Piccolo, G.; Iannaccone, F.; Nizza, A. Effect of mannanoligosaccharides supplementation on caecal microbial activity of rabbits. Animal 2010, 4, 1522–1527. [Google Scholar] [CrossRef] [Green Version]
- Suiryanrayna, M.V.A.N.; Ramana, J.V. A review of the effects of dietary organic acids fed to swine. J. Anim. Sci. Biotechnol. 2015, 6, 215. [Google Scholar] [CrossRef]
- Liu, B.; Wang, W.; Zhu, X.; Sun, X.; Xiao, J.; Li, D.; Cui, Y.; Wang, C.; Shi, Y. Response of Gut Microbiota to Dietary Fiber and Metabolic Interaction With SCFAs in Piglets. Front. Microbiol. 2018, 9, 2344. [Google Scholar] [CrossRef] [Green Version]
- Borrelli, L.; Coretti, L.; Dipineto, L.; Bovera, F.; Menna, F.; Chiariotti, L.; Nizza, A.; Lembo, F.; Fioretti, A. Insect-based diet, a promising nutritional source, modulates gut microbiota composition and SCFAs production in laying hens. Sci. Rep. 2017, 7, 16269. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef]
- Dubé, P.E.; Brubaker, P.L. Frontiers in glucagon-like peptide-2: Multiple actions, multiple mediators. Am. J. Physiol. Metab. 2007, 293, E460–E465. [Google Scholar] [CrossRef]
- Everard, A.; Cani, P.D. Gut microbiota and GLP-1. Rev. Endocr. Metab. Disord. 2014, 15, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Rowland, K.J.; Trivedi, S.; Lee, D.; Wan, K.; Kulkarni, R.N.; Holzenberger, M.; Brubaker, P.L. Loss of Glucagon-Like Peptide-2-Induced Proliferation Following Intestinal Epithelial Insulin-Like Growth Factor-1–Receptor Deletion. Gastroenterology 2011, 141, 2166–2175. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Everard, A.; Duparc, T. Gut microbiota, enteroendocrine functions and metabolism. Curr. Opin. Pharmacol. 2013, 13, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, J.; Hasegawa, S.; Kasubuchi, M.; Ichimura, A.; Nakajima, A.; Kimura, I. Nutritional Signaling via Free Fatty Acid Receptors. Int. J. Mol. Sci. 2016, 17, 450. [Google Scholar] [CrossRef] [PubMed]
- Nøhr, M.K.; Pedersen, M.H.; Gille, A.; Egerod, K.L.; Engelstoft, M.S.; Husted, A.S.; Sichlau, R.M.; Grunddal, K.V.; Poulsen, S.S.; Han, S.; et al. GPR41/FFAR3 and GPR43/FFAR2 as Cosensors for Short-Chain Fatty Acids in Enteroendocrine Cells vs FFAR3 in Enteric Neurons and FFAR2 in Enteric Leukocytes. Endocrinology 2013, 154, 3552–3564. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, G.H.; Agerberth, B.; Kai-Larsen, Y.; Kai-Larsen, Y. A review of the innate immune defence of the human foetus and newborn, with the emphasis on antimicrobial peptides. Acta Paediatr. 2014, 103, 1000–1008. [Google Scholar]
- Alizadeh, A.; Akbari, P.; Difilippo, E.; Schols, H.; Ulfman, L.; Schoterman, M.; Garssen, J.; Fink-Gremmels, J.; Braber, S. The piglet as a model for studying dietary components in infant diets: Effects of galacto-oligosaccharides on intestinal functions. Br. J. Nutr. 2016, 115, 605–618. [Google Scholar] [CrossRef]
- Veldhuizen, E.J.; Rijnders, M.; Claassen, E.A.; Van Dijk, A.; Haagsman, H.P. Porcine β-defensin 2 displays broad antimicrobial activity against pathogenic intestinal bacteria. Mol. Immunol. 2008, 45, 386–394. [Google Scholar] [CrossRef]
- De Witte, C.; Flahou, B.; Ducatelle, R.; Smet, A.; De Bruyne, E.; Cnockaert, M.; Taminiau, B.; Daube, G.; Vandamme, P.; Haesebrouck, F. Detection, isolation and characterization of Fusobacterium gastrosuis sp. nov. colonizing the stomach of pigs. Syst. Appl. Microbiol. 2017, 40, 42–50. [Google Scholar] [CrossRef]
- Cardines, R.; Daprai, L.; Torresani, E.; Garlaschi, M.L.; Cerquetti, M.; Giufrè, M. Genital carriage of the genus Haemophilus in pregnancy: Species distribution and antibiotic susceptibility. J. Med. Microbiol. 2015, 64, 724–730. [Google Scholar] [CrossRef]
- Watkins, L.R.; Maier, S.F.; Goehler, L.E. Immune activation: The role of pro-inflammatory cytokines in inflammation, illness responses and pathological pain states. Pain 1995, 63, 289–302. [Google Scholar] [CrossRef]
- Opal, S.M.; Depalo, V.A. Anti-inflammatory cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | CON | GOS | P-Value |
|---|---|---|---|
| Day 8 | |||
| pH value | 7.05 ± 0.07 | 6.57 ± 0.11 | 0.009 |
| Acetate (μmol/g digesta) | 3.73 ± 0.49 | 10.09 ± 3.21 | 0.099 |
| Propionate (μmol/g digesta) | 0.83 ± 0.57 | 2.59 ± 0.29 | 0.000 |
| Butyrate (μmol/g digesta) | 0.59 ± 0.10 | 1.84 ± 0.36 | 0.011 |
| Valerate (μmol/g digesta) | 0.16 ± 0.02 | 0.25 ± 0.02 | 0.008 |
| Lactate (μmol/g digesta) | 15.15 ± 0.55 | 18.54 ± 0.66 | 0.003 |
| Day 21 | |||
| pH value | 6.94 ± 0.07 | 7.05 ± 0.11 | 0.406 |
| Acetate (μmol/g digesta) | 9.43 ± 1.88 | 9.96 ± 1.85 | 0.847 |
| Propionate (μmol/g digesta) | 2.18 ± 0.47 | 2.18 ± 0.35 | 0.996 |
| Butyrate (μmol/g digesta) | 0.73 ± 0.08 | 1.13 ± 0.14 | 0.041 |
| Valerate (μumol/g digesta) | 0.29 ± 0.05 | 0.30 ± 0.06 | 0.939 |
| Lactate (μmol/g digesta) | 29.53 ± 1.40 | 30.28 ± 1.20 | 0.693 |
| Item b | CON | GOS | P-Value |
|---|---|---|---|
| Day 8 | |||
| GLP-1 (pg/g) | 1.16 ± 0.10 | 1.51 ± 0.10 | 0.033 |
| GLP-2 (pg/g) | 2.25 ± 0.23 | 2.66 ± 0.14 | 0.155 |
| IGF-1 (ug/g) | 4.38 ± 0.57 | 5.37 ± 0.30 | 0.156 |
| EGF (ng/g) | 114.63 ± 10.05 | 153.99 ± 15.50 | 0.059 |
| Day 21 | |||
| GLP-1 (pg/g) | 1.47 ± 0.13 | 1.69 ± 0.08 | 0.163 |
| GLP-2 (pg/g) | 2.83 ± 0.17 | 3.06 ± 0.26 | 0.463 |
| IGF-1 (ug/g) | 5.24 ± 0.33 | 6.15 ± 0.60 | 0.216 |
| EGF (ng/g) | 154.19 ± 8.90 | 167.18 ± 15.84 | 0.491 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, S.; Wang, J.; Yu, H.; Wang, J.; Zhu, W. Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model. Nutrients 2019, 11, 1753. https://doi.org/10.3390/nu11081753
Tian S, Wang J, Yu H, Wang J, Zhu W. Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model. Nutrients. 2019; 11(8):1753. https://doi.org/10.3390/nu11081753
Chicago/Turabian StyleTian, Shiyi, Jue Wang, Hu Yu, Jing Wang, and Weiyun Zhu. 2019. "Changes in Ileal Microbial Composition and Microbial Metabolism by an Early-Life Galacto-Oligosaccharides Intervention in a Neonatal Porcine Model" Nutrients 11, no. 8: 1753. https://doi.org/10.3390/nu11081753