Protection of the Ovine Fetal Gut against Ureaplasma-Induced Chorioamnionitis: A Potential Role for Plant Sterols

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
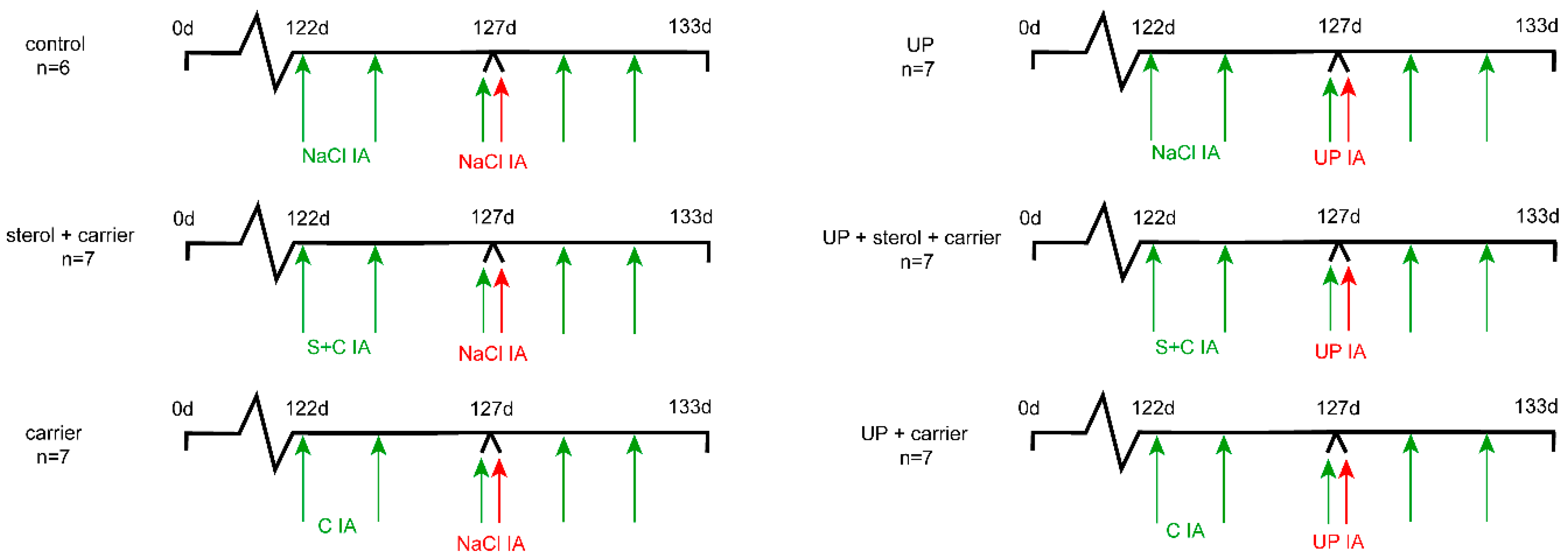
2.1. Experimental Design
2.2. Antibodies
2.3. ELISA
2.4. Immunohistochemistry
2.5. Gas–Liquid Chromatography–Mass Spectrometry (GC-MS)
2.6. Matrix-Assisted Laser Desorption Ionization Mass Spectrometry Imaging
2.7. Statistics
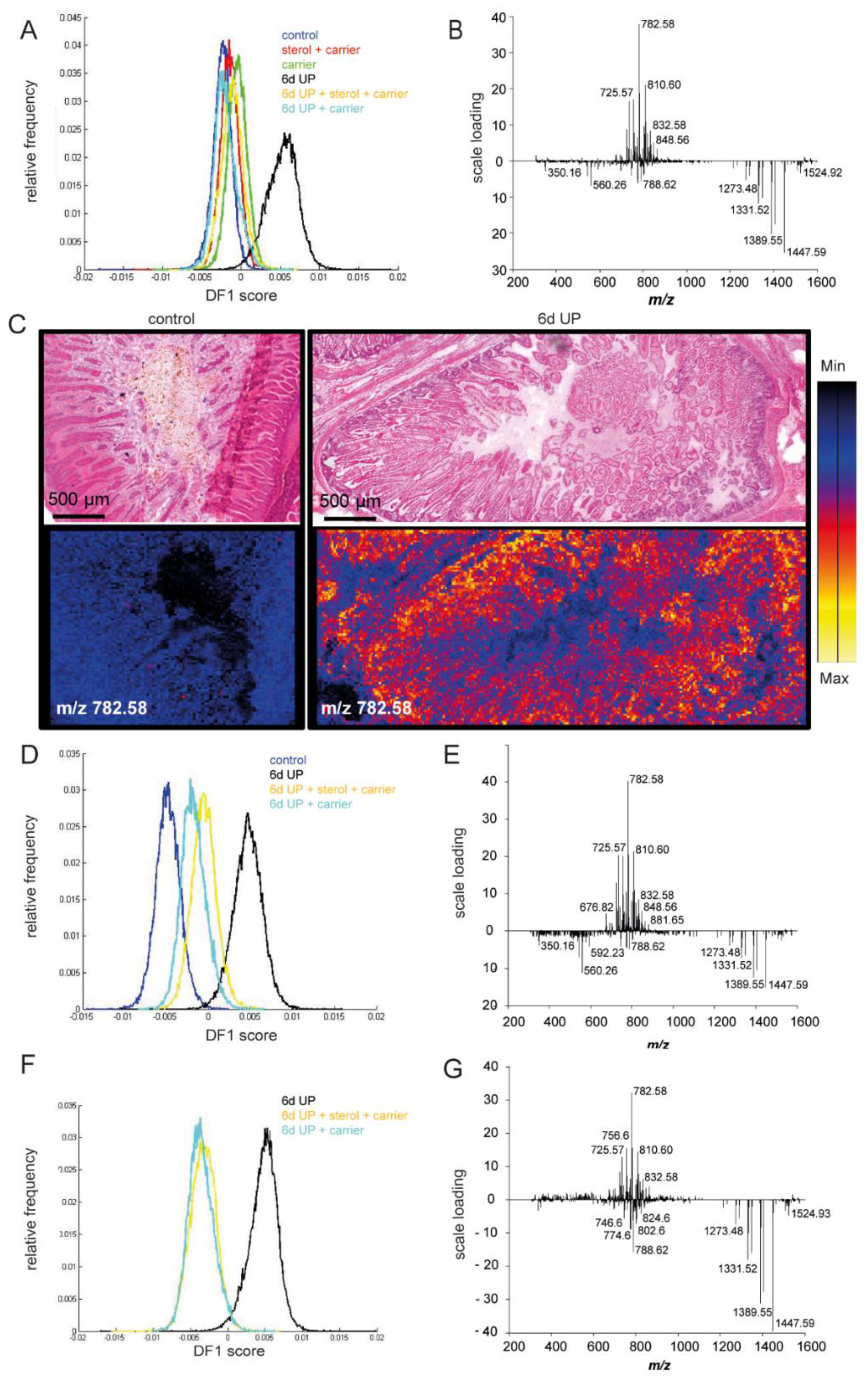
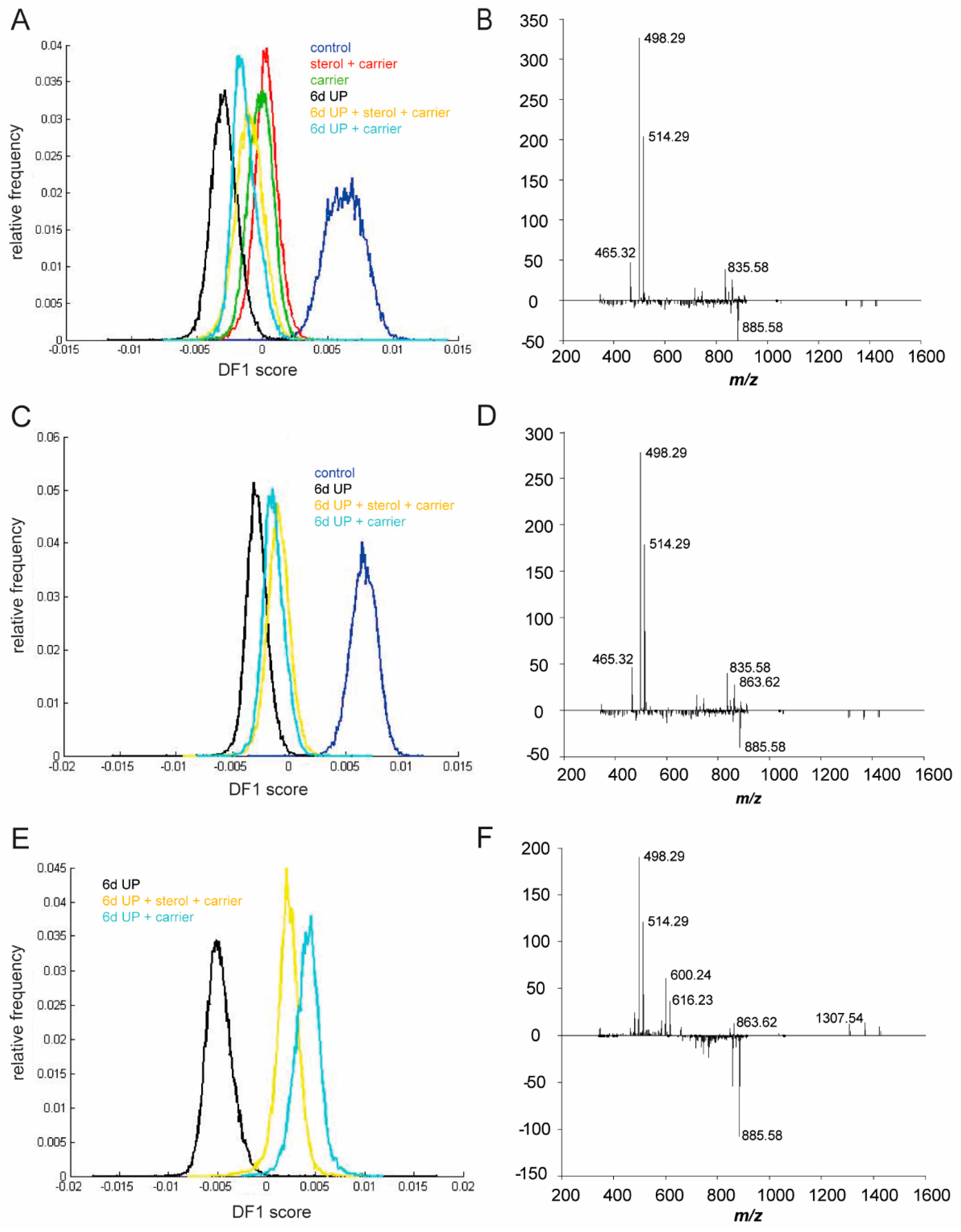
3. Results
3.1. Systemic and Intestinal Inflammation
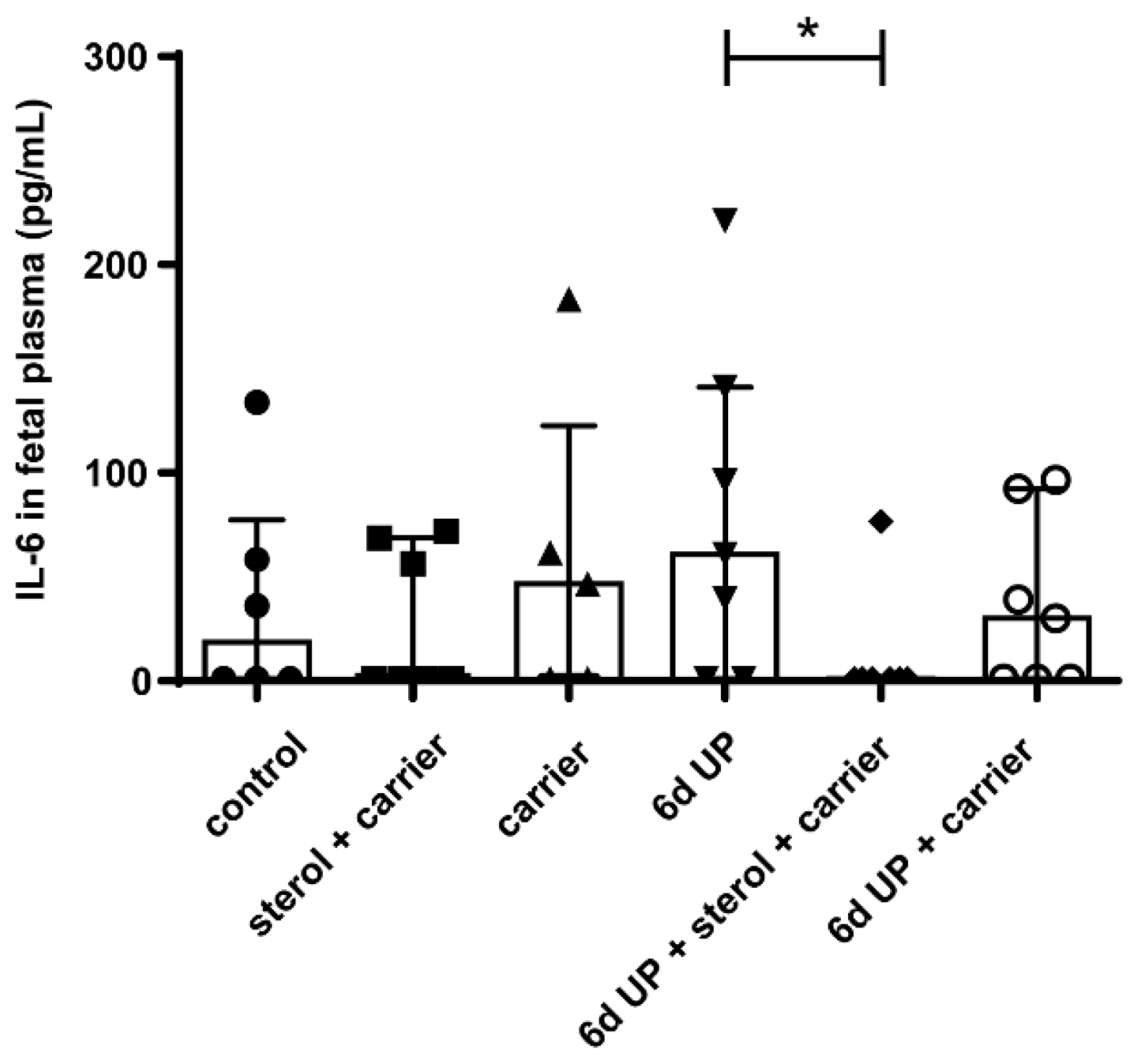
3.1.1. Systemic Inflammation
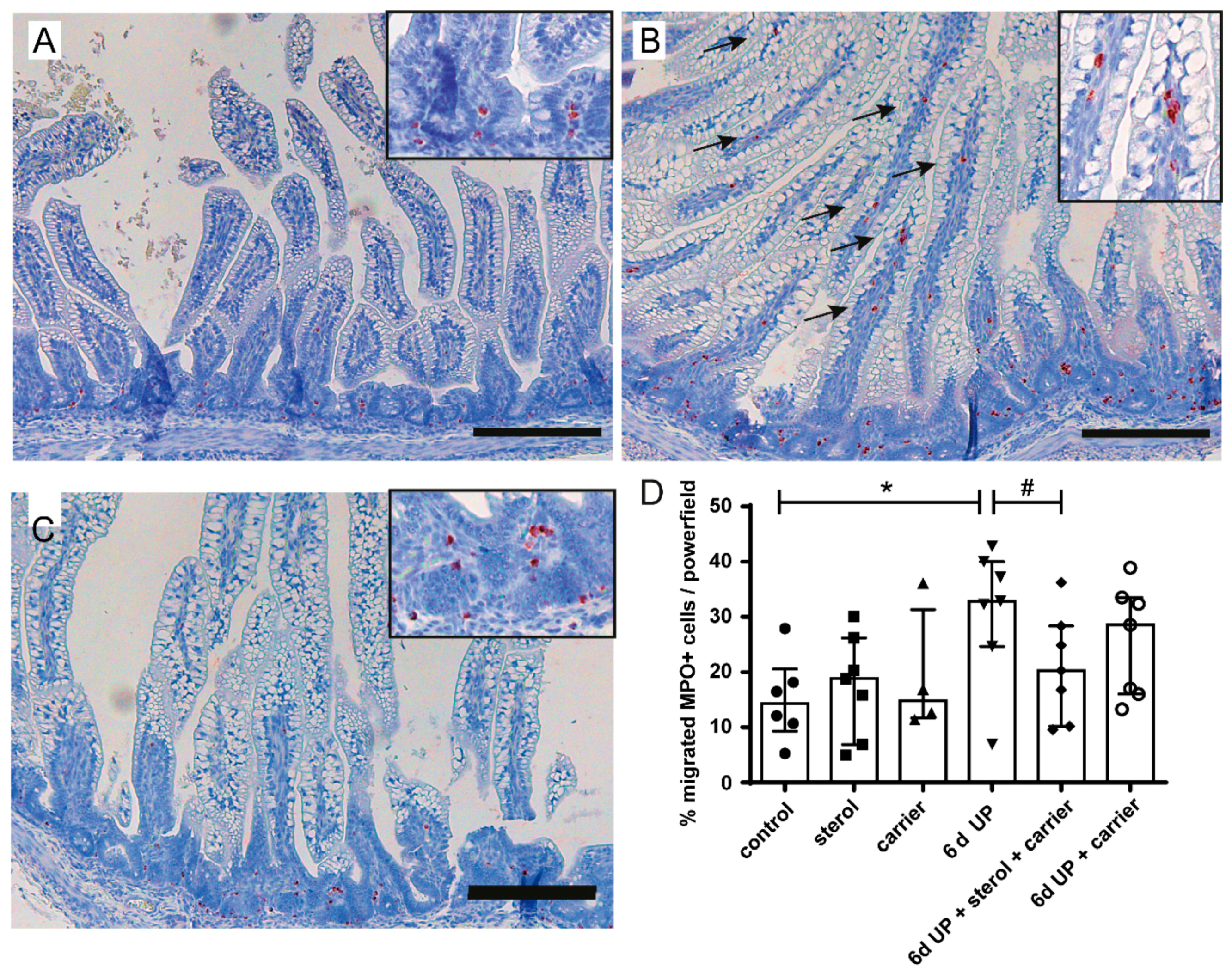
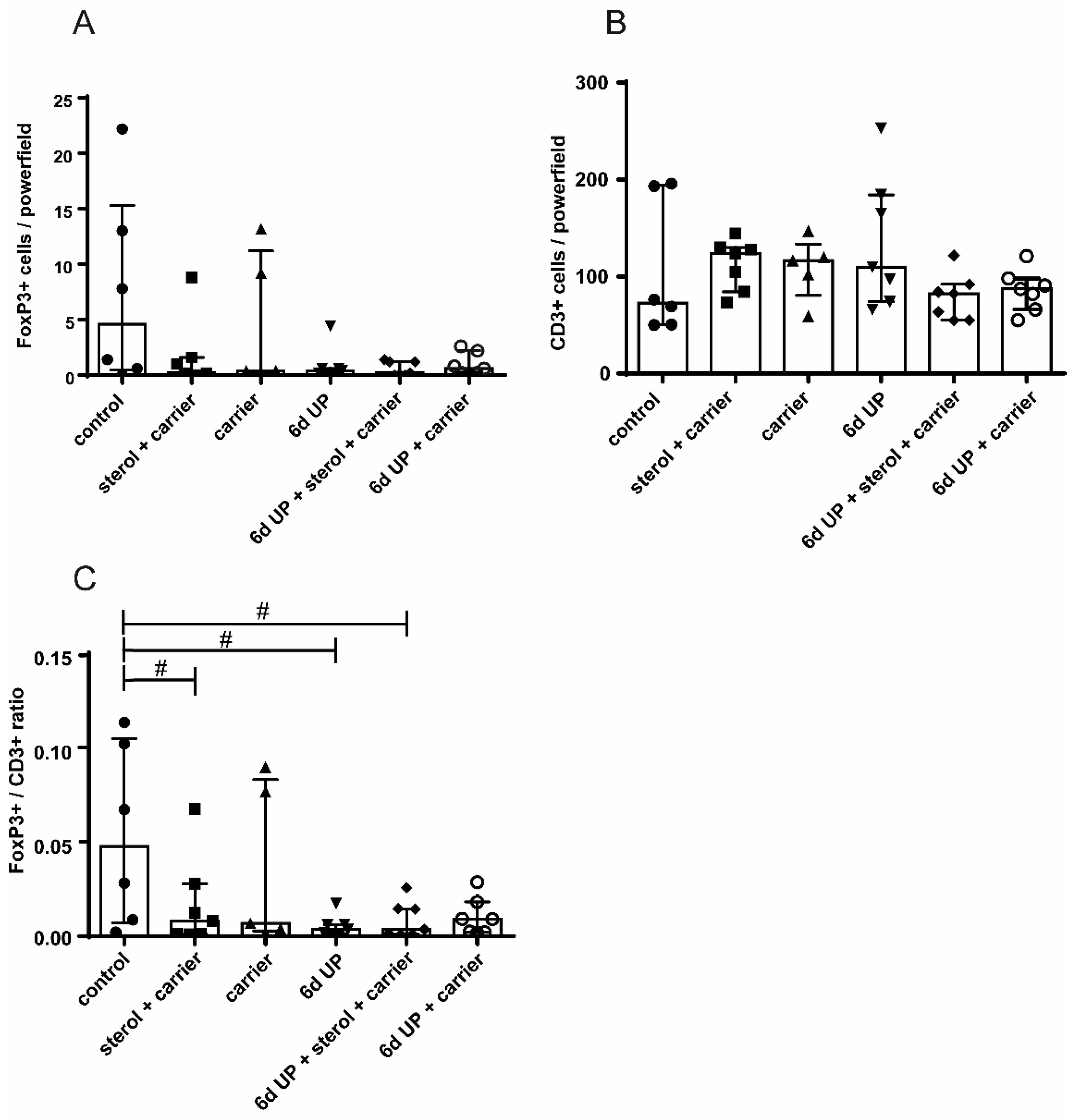
3.1.2. Intestinal inflammation
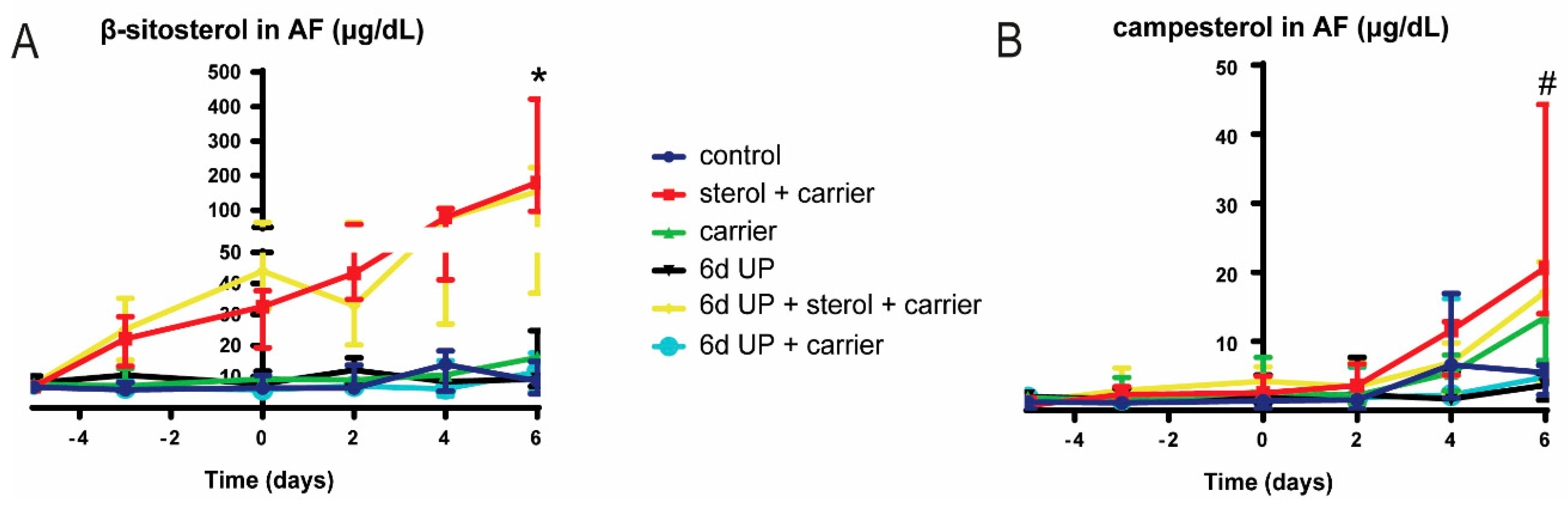
3.2. UP and Sterol Concentrations in Amniotic Fluid
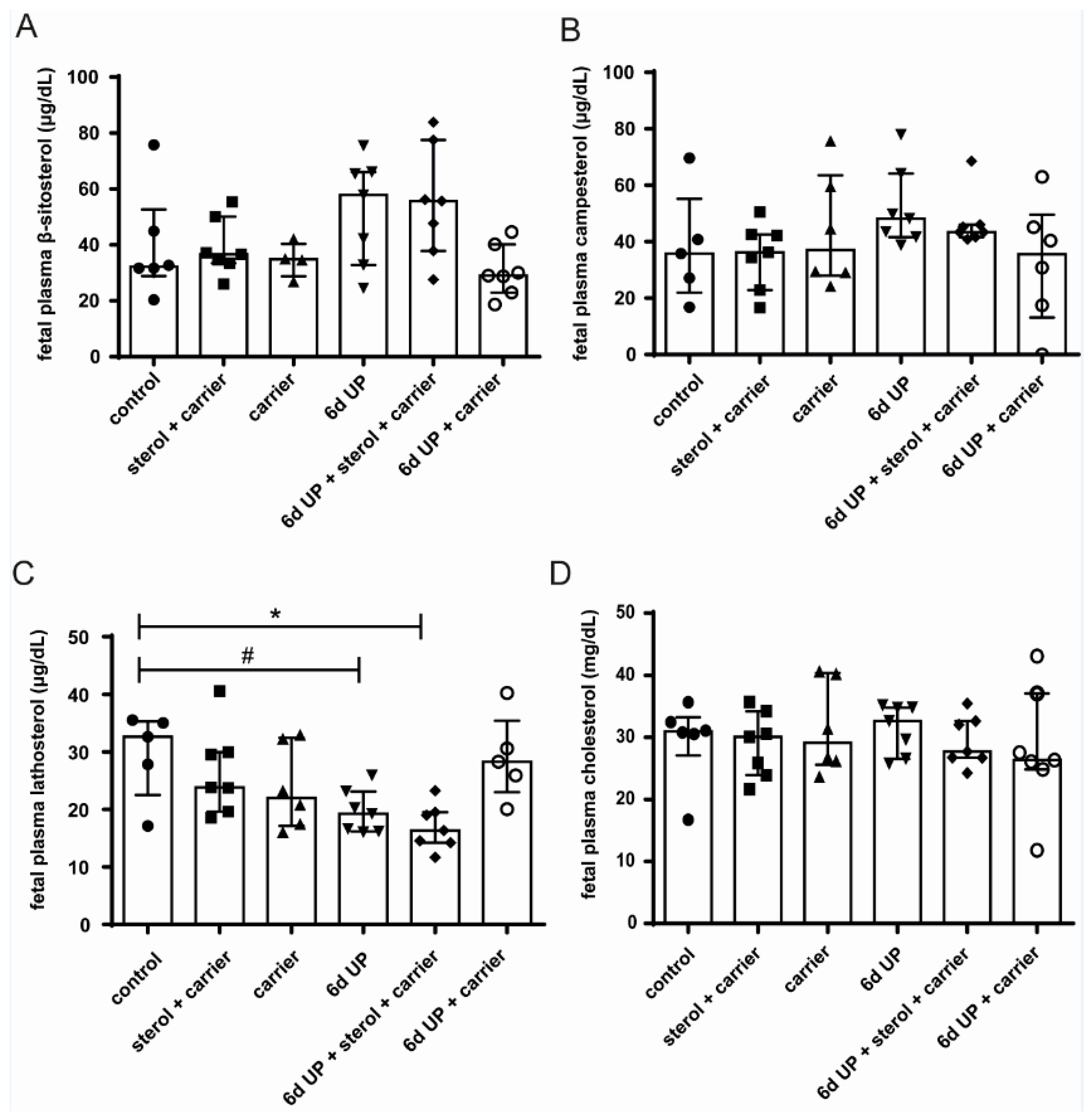
3.3. Sterol Concentrations in Fetal Plasma
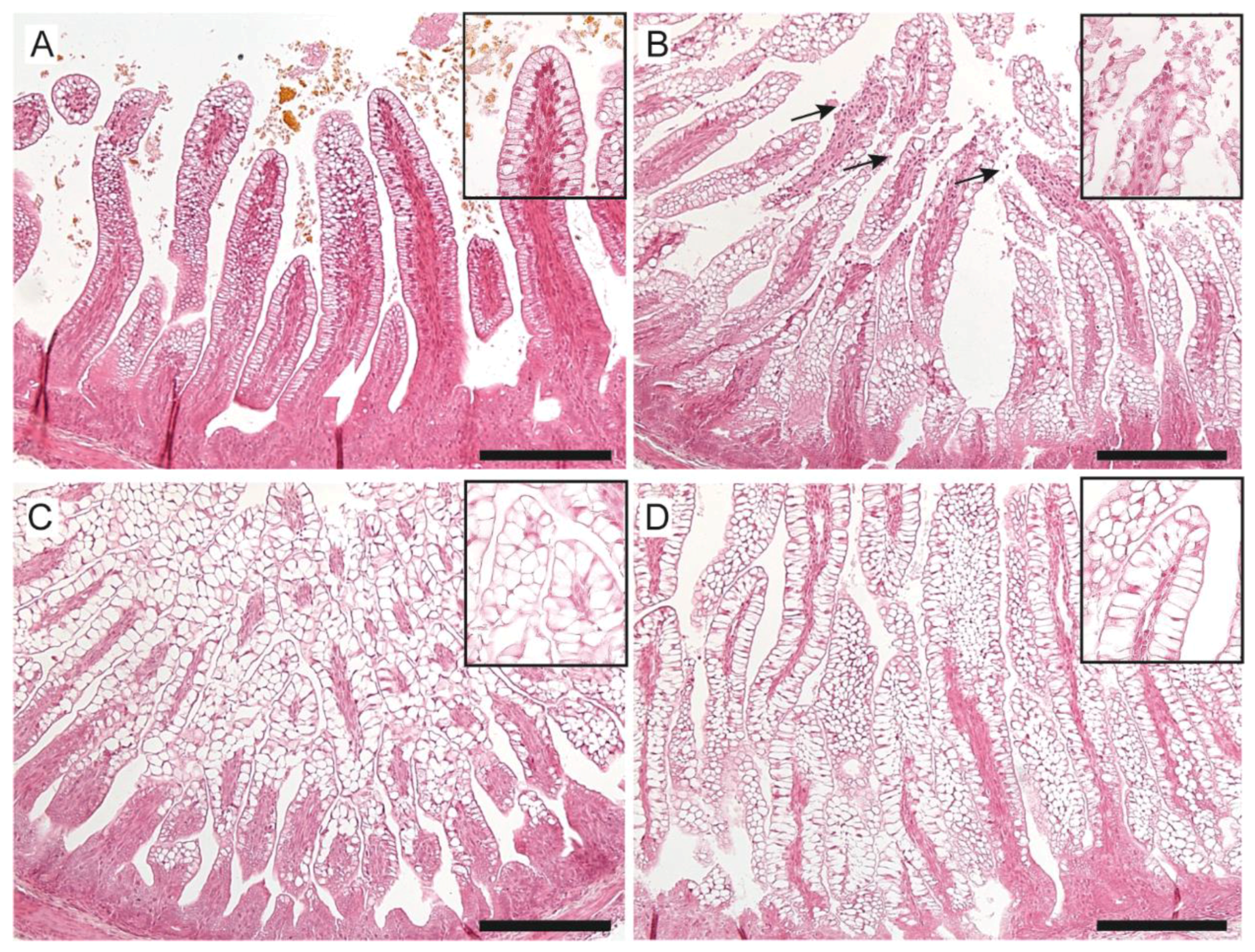
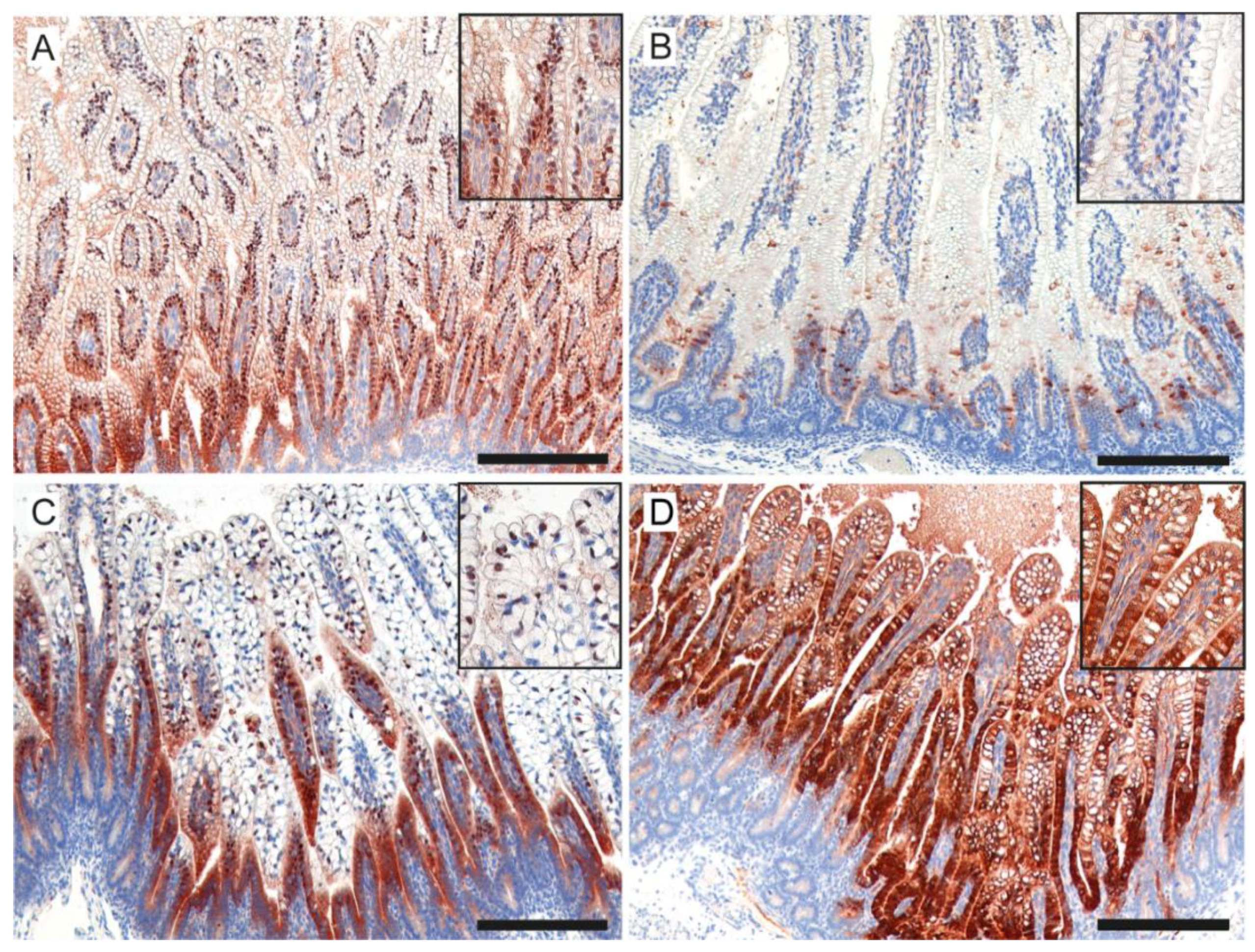
3.4. Intestinal Damage and Gut Maturation
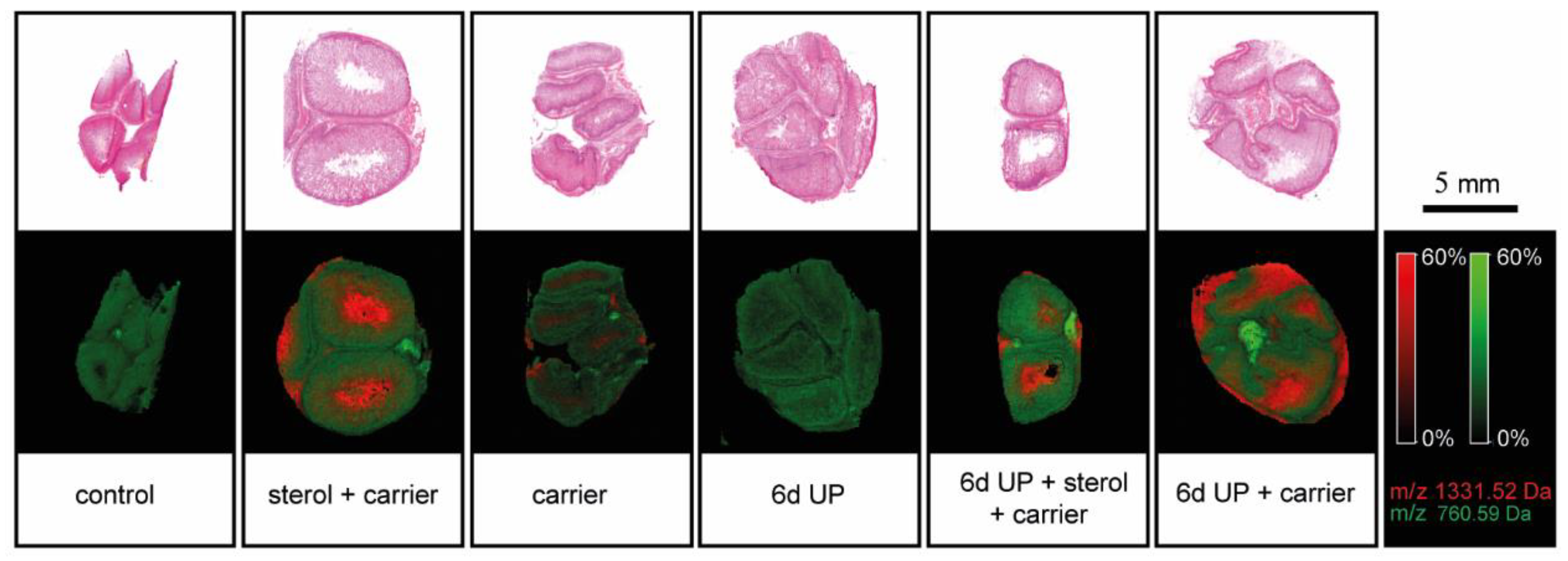
3.5. Changes in Lipid Profiles in the Fetal Gut
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AF | amniotic fluid |
| BA | bile acids |
| CCU | color changing units |
| DA | discriminant analysis |
| FIRS | fetal inflammatory response syndrome |
| FoxP3 | fork head box P3 |
| GA | gestational age |
| GC-MS | gas–liquid chromatography–mass spectrometry |
| H&E | hematoxylin and eosin staining |
| IA | intra-amniotic |
| I-FABP | intestinal fatty acid binding protein |
| IQR | interquartile range |
| LXR | liver X receptor |
| MALDI-MSI | matrix-assisted laser desorption ionization mass spectrometry imaging |
| MPO | myeloperoxidase |
| NEC | necrotizing enterocolitis |
| PC | phosphatidylcholine |
| PCAs | principal component analyses |
| PI | phosphatidylinositol |
| SM | sphingomyelin |
| TCDCA | taurochenodeoxycholic acid |
| TCDA | taurodeoxycholic acid |
| Treg | regulatory T-cells |
| TMB | 3,3′,5,5′-tetramethylbenzidine |
| UP | U. parvum |
References
- Liu, L.; Oza, S.; Hogan, D.; Chu, Y.; Perin, J.; Zhu, J.; E Lawn, J.; Cousens, S.; Mathers, C.; E Black, R. Global, regional, and national causes of under-5 mortality in 2000–15: An updated systematic analysis with implications for the Sustainable Development Goals. Lancet 2016, 388, 3027–3035. [Google Scholar] [CrossRef]
- Blencowe, H.; Cousens, S.; Oestergaard, M.Z.; Chou, D.; Moller, A.-B.; Narwal, R.; Adler, A.; Garcia, C.V.; Rohde, S.; Say, L.; et al. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: A systematic analysis and implications. Lancet 2012, 379, 2162–2172. [Google Scholar] [CrossRef]
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute Chorioamnionitis and Funisitis: Definition, Pathologic Features, and Clinical Significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfs, T.G.; Jellema, R.K.; Turrisi, G.; Becucci, E.; Buonocore, G.; Kramer, B.W. Inflammation-induced immune suppression of the fetus: A potential link between chorioamnionitis and postnatal early onset sepsis. J. Matern.-Fetal Neonatal Med. 2012, 25, 8–11. [Google Scholar] [CrossRef] [PubMed]
- Wolfs, T.G.A.M.; Buurman, W.A.; Zoer, B.; Moonen, R.M.J.; Derikx, J.P.M.; Thuijls, G.; Villamor, E.; Gantert, M.; Garnier, Y.; Zimmermann, L.J.I.; et al. Endotoxin Induced Chorioamnionitis Prevents Intestinal Development during Gestation in Fetal Sheep. PLoS ONE 2009, 4, e5837. [Google Scholar] [CrossRef]
- Wolfs, T.G.; Kallapur, S.G.; Knox, C.L.; Thuijls, G.; Nitsos, I.; Polglase, G.R.; Collins, J.J.; Kroon, E.; Spierings, J.; Shroyer, N.F.; et al. Antenatal ureaplasma infection impairs development of the fetal ovine gut in an IL-1-dependent manner. Mucosal Immunol. 2013, 6, 547–556. [Google Scholar] [CrossRef]
- Been, J.V.; Lievense, S.; Zimmermann, L.J.; Kramer, B.W.; Wolfs, T.G. Chorioamnionitis as a Risk Factor for Necrotizing Enterocolitis: A Systematic Review and Meta-Analysis. J. Pediatr. 2013, 162, 236–242.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gantert, M.; Been, J.V.; Gavilanes, A.W.; Garnier, Y.; I Zimmermann, L.J.; Kramer, B.W. Chorioamnionitis: A multiorgan disease of the fetus? J. Perinatol. 2010, 30, S21–S30. [Google Scholar] [CrossRef]
- Neu, J.; Walker, W.A. Necrotizing Enterocolitis. N. Engl. J. Med. 2011, 364, 255–264. [Google Scholar] [CrossRef] [Green Version]
- Gotsch, F.; Romero, R.; Kusanovic, J.P.; Mazaki-Tovi, S.; Pineles, B.L.; Erez, O.; Espinoza, J.; Hassan, S.S. The Fetal Inflammatory Response Syndrome. Clin. Obstet. Gynecol. 2007, 50, 652–683. [Google Scholar] [CrossRef]
- Gylling, H.; Plat, J.; Turley, S.; Ginsberg, H.N.; Ellegård, L.; Jessup, W.; Jones, P.J.; Lütjohann, D.; Maerz, W.; Masana, L.; et al. Plant sterols and plant stanols in the management of dyslipidaemia and prevention of cardiovascular disease. Atherosclerosis 2014, 232, 346–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jong, A.; Plat, J.; Lütjohann, D.; Mensink, R.P. Effects of long-term plant sterol or stanol ester consumption on lipid and lipoprotein metabolism in subjects on statin treatment. Br. J. Nutr. 2008, 100, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Brüll, F.; De Smet, E.; Mensink, R.P.; Vreugdenhil, A.; Kerksiek, A.; Lütjohann, D.; Wesseling, G.; Plat, J. Dietary plant stanol ester consumption improves immune function in asthma patients: Results of a randomized, double-blind clinical trial1. Am. J. Clin. Nutr. 2016, 103, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Velde, A.A.T.; Brüll, F.; Heinsbroek, S.E.M.; Meijer, S.L.; Lütjohann, D.; Vreugdenhil, A.; Plat, J.; Velde, A.T. Effects of Dietary Plant Sterols and Stanol Esters with Low- and High-Fat Diets in Chronic and Acute Models for Experimental Colitis. Nutrients 2015, 7, 8518–8531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldini, R.; Micucci, M.; Cevenini, M.; Fato, R.; Bergamini, C.; Nanni, C.; Cont, M.; Camborata, C.; Spinozzi, S.; Montagnani, M.; et al. Antiinflammatory Effect of Phytosterols in Experimental Murine Colitis Model: Prevention, Induction, Remission Study. PLoS ONE 2014, 9, e108112. [Google Scholar] [CrossRef]
- Lee, I.-A.; Kim, E.-J.; Kim, D.-H. Inhibitory Effect of β-Sitosterol on TNBS-Induced Colitis in Mice. Planta Med. 2012, 78, 896–898. [Google Scholar] [CrossRef]
- Plat, J.; Baumgartner, S.; Vanmierlo, T.; Lutjohann, D.; Calkins, K.L.; Burrin, D.G.; Guthrie, G.; Thijs, C.; Te Velde, A.A.; Vreugdenhil, A.C.E.; et al. Plant-based sterols and stanols in health & disease: “Consequences of human development in a plant-based environment?”. Prog. Lipid Res. 2019. [Google Scholar] [CrossRef]
- Goldenberg, R.L.; Andrews, W.W.; Hauth, J.C. Intrauterine Infection and Preterm Delivery. N. Engl. J. Med. 2000, 342, 1500–1507. [Google Scholar] [CrossRef] [PubMed]
- Viscardi, R.M. Ureaplasma species: Role in neonatal morbidities and outcomes. Arch. Dis. Child.-Fetal Neonatal Ed. 2014, 99, F87–F92. [Google Scholar] [CrossRef] [PubMed]
- Okogbule-Wonodi, A.C.; Gross, G.W.; Sun, C.-C.J.; Agthe, A.G.; Xiao, L.; Waites, K.B.; Viscardi, R.M.; Okogbule-Wonodi, G.W.G.A.C. Necrotizing Enterocolitis is associated with Ureaplasma Colonization in Preterm Infants. Pediatr. Res. 2011, 69, 442–447. [Google Scholar] [CrossRef] [Green Version]
- Kilkenny, C.; Browne, W.; Cuthill, I.C.; Emerson, M.; Altman, D.G. Animal research: Reporting in vivo experiments: The ARRIVE guidelines. Br. J. Pharmacol. 2010, 160, 1577–1579. [Google Scholar] [CrossRef] [Green Version]
- Calpe-Berdiel, L.; Escolà-Gil, J.C.; Blanco-Vaca, F. New insights into the molecular actions of plant sterols and stanols in cholesterol metabolism. Atherosclerosis 2009, 203, 18–31. [Google Scholar] [CrossRef]
- Baardman, M.E.; Erwich, J.J.H.; Berger, R.M.; Hofstra, R.M.; Kerstjens-Frederikse, W.S.; Lütjohann, D.; Plösch, T. The origin of fetal sterols in second-trimester amniotic fluid: Endogenous synthesis or maternal-fetal transport? Am. J. Obstet. Gynecol. 2012, 207, 202.e19–202.e25. [Google Scholar] [CrossRef]
- Amaral, C.; Gallardo, E.; Rodrigues, R.; Leite, R.P.; Quelhas, D.; Tomaz, C.; Cardoso, M. Quantitative analysis of five sterols in amniotic fluid by GC–MS: Application to the diagnosis of cholesterol biosynthesis defects. J. Chromatogr. B 2010, 878, 2130–2136. [Google Scholar] [CrossRef]
- Burke, K.T.; Colvin, P.L.; Myatt, L.; Graf, G.A.; Schroeder, F.; Woollett, L.A. Transport of maternal cholesterol to the fetus is affected by maternal plasma cholesterol concentrations in the Golden Syrian hamster. J. Lipid Res. 2009, 50, 1146–1155. [Google Scholar] [CrossRef] [Green Version]
- Ras, R.T.; Hiemstra, H.; Lin, Y.; Vermeer, M.A.; Duchateau, G.S.; Trautwein, E.A. Consumption of plant sterol-enriched foods and effects on plasma plant sterol concentrations—A meta-analysis of randomized controlled studies. Atherosclerosis 2013, 230, 336–346. [Google Scholar] [CrossRef]
- Moss, T.J.; Knox, C.L.; Kallapur, S.G.; Nitsos, I.; Theodoropoulos, C.; Newnham, J.P.; Ikegami, M.; Jobe, A.H. Experimental amniotic fluid infection in sheep: Effects of Ureaplasma parvum serovars 3 and 6 on preterm or term fetal sheep. Am. J. Obstet. Gynecol. 2008, 198, 122.e1–122.e8. [Google Scholar] [CrossRef]
- Miura, Y.; Payne, M.S.; Keelan, J.A.; Noe, A.; Carter, S.; Watts, R.; Spiller, O.B.; Jobe, A.H.; Kallapur, S.G.; Saito, M.; et al. Maternal Intravenous Treatment with either Azithromycin or Solithromycin Clears Ureaplasma parvum from the Amniotic Fluid in an Ovine Model of Intrauterine Infection. Antimicrob. Agents Chemother. 2014, 58, 5413–5420. [Google Scholar] [CrossRef]
- Dello, S.A.W.G.; Reisinger, K.W.; Van Dam, R.M.; Bemelmans, M.H.A.; Van Kuppevelt, T.H.; Broek, M.A.J.V.D.; Damink, S.W.M.O.; Poeze, M.; Buurman, W.A.; DeJong, C.H.C. Total Intermittent Pringle Maneuver during Liver Resection Can Induce Intestinal Epithelial Cell Damage and Endotoxemia. PLoS ONE 2012, 7, e30539. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Boil. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Barré, F.; Rocha, B.; Dewez, F.; Towers, M.; Murray, P.; Claude, E.; Cillero-Pastor, B.; Heeren, R.; Porta Siegel, T. Faster raster matrix-assisted laser desorption/ionization mass spectrometry imaging of lipids at high lateral resolution. Int. J. Mass Spectrom. 2019, 437, 38–48. [Google Scholar] [CrossRef]
- Willems, M.G.; Ophelders, D.R.; Nikiforou, M.; Jellema, R.K.; Butz, A.; Delhaas, T.; Kramer, B.W.; Wolfs, T.G. Systemic interleukin-2 administration improves lung function and modulates chorioamnionitis-induced pulmonary inflammation in the ovine fetus. Am. J. Physiol. Cell. Mol. Physiol. 2016, 310, L1–L7. [Google Scholar] [CrossRef]
- Jiang, H.; Sidhu, R.; Fujiwara, H.; De Meulder, M.; De Vries, R.; Gong, Y.; Kao, M.; Porter, F.D.; Yanjanin, N.M.; Carillo-Carasco, N.; et al. Development and validation of sensitive LC-MS/MS assays for quantification of HP-β-CD in human plasma and CSF. J. Lipid Res. 2014, 55, 1537–1548. [Google Scholar] [CrossRef] [Green Version]
- Bouic, P.; Clark, A.; Lamprecht, J.; Freestone, M.; Pool, E.; Liebenberg, R.; Kotze, D.; Van Jaarsveld, P. The Effects of B-Sitosterol (BSS) and B-Sitosterol Glucoside (BSSG) Mixture on Selected Immune Parameters of Marathon Runners: Inhibition of Post Marathon Immune Suppression and Inflammation. Int. J. Sports Med. 1999, 20, 258–262. [Google Scholar] [CrossRef]
- Camus, G.; Deby-Dupont, G.; Duchateau, J.; Deby, C.; Pincemail, J.; Lamy, M. Are similar inflammatory factors involved in strenuous exercise and sepsis? Intensiv. Care Med. 1994, 20, 602–610. [Google Scholar] [CrossRef]
- Nashed, B.; HayGlass, K.T.; Moghadasian, M.H.; Yeganeh, B. Antiatherogenic Effects of Dietary Plant Sterols Are Associated with Inhibition of Proinflammatory Cytokine Production in Apo E-KO Mice. J. Nutr. 2005, 135, 2438–2444. [Google Scholar] [CrossRef] [Green Version]
- Wolfs, T.G.; Kallapur, S.G.; Polglase, G.R.; Pillow, J.J.; Nitsos, I.; Newnham, J.P.; Chougnet, C.A.; Kroon, E.; Spierings, J.; Willems, C.H.; et al. IL-1alpha mediated chorioamnionitis induces depletion of FoxP3+ cells and ileal inflammation in the ovine fetal gut. PLoS ONE 2011, 6, e18355. [Google Scholar] [CrossRef]
- De Smet, E.; Mensink, R.P.; Boekschoten, M.V.; De Ridder, R.; Germeraad, W.T.V.; Wolfs, T.G.A.M.; Plat, J. An acute intake of plant stanol esters alters immune-related pathways in the jejunum of healthy volunteers. Br. J. Nutr. 2015, 113, 794–802. [Google Scholar] [CrossRef] [Green Version]
- Woollett, L.; Heubi, J.E. Endotext: Fetal and Neonatal Cholesterol Metabolism; De Groot, L.J., Chrousos, G., Dungan, K., Feingold, K.R., Grossman, A., Hershman, J.M., Koch, C., Korbonits, M., McLachlan, R., New, M., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Furse, S.; De Kroon, A.I.P.M. Phosphatidylcholine’s functions beyond that of a membrane brick. Mol. Membr. Boil. 2015, 32, 117–119. [Google Scholar] [CrossRef] [PubMed]
- Ohanian, J.; Ohanian, V. Sphingolipids in mammalian cell signalling. Cell. Mol. Life Sci. 2001, 58, 2053–2068. [Google Scholar] [CrossRef] [PubMed]
- Payrastre, B.; Missy, K.; Giuriato, S.; Bodin, S.; Plantavid, M.; Gratacap, M.-P. Phosphoinositides: Key players in cell signalling, in time and space. Cell Signal. 2001, 13, 377–387. [Google Scholar] [CrossRef]
- Pike, L.J. Lipid rafts: Bringing order to chaos. J. Lipid Res. 2003, 44, 655–667. [Google Scholar] [CrossRef] [PubMed]
- Gussenhoven, R.; Ophelders, D.R.; Kemp, M.W.; Payne, M.S.; Spiller, O.B.; Beeton, M.L.; Stock, S.J.; Cillero-Pastor, B.; Barré, F.P.; Heeren, R.M.; et al. The paradoxical effects of chronic intra-amniotic Ureaplasma parvum exposure on ovine fetal brain development. Dev. Neurosci. 2017, 39, 472–486. [Google Scholar] [CrossRef]
- Adibhatla, R.M.; Hatcher, J.F. Role of Lipids in Brain Injury and Diseases. Future Lipidol. 2007, 2, 403–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallima, H.; El Ridi, R. Arachidonic acid: Physiological roles and potential health benefits—A review. J. Adv. Res. 2018, 11, 33–41. [Google Scholar] [CrossRef]
- Hofmann, A.F.; Eckmann, L. How bile acids confer gut mucosal protection against bacteria. Proc. Natl. Acad. Sci. USA 2006, 103, 4333–4334. [Google Scholar] [CrossRef] [Green Version]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [Green Version]
- Marin, J.J.G.; Macias, R.I.R.; Briz, O.; Banales, J.M.; Monte, M.J. Bile Acids in Physiology, Pathology and Pharmacology. Drug Metab. 2015, 17, 4–29. [Google Scholar] [CrossRef]
- Bajor, A.; Gillberg, P.-G.; Abrahamsson, H. Bile acids: Short and long term effects in the intestine. Scand. J. Gastroenterol. 2010, 45, 645–664. [Google Scholar] [CrossRef]
- Berseth, C. Gestational evolution of small intestine motility in preterm and term infants. J. Pediatr. 1989, 115, 646–651. [Google Scholar] [CrossRef]
- Niño, D.F.; Sodhi, C.P.; Hackam, D.J. Necrotizing enterocolitis: New insights into pathogenesis and mechanisms. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 590–600. [Google Scholar] [CrossRef] [PubMed]
- Warner, B.B.; Deych, E.; Zhou, Y.; Hall-Moore, C.; Weinstock, G.M.; Sodergren, E.; Shaikh, N.; Hoffmann, J.A.; Linneman, L.A.; Hamvas, A.; et al. Gut bacteria dysbiosis and necrotising enterocolitis in very low birthweight infants: A prospective case-control study. Lancet 2016, 387, 1928–1936. [Google Scholar] [CrossRef]
- Atger, V.M.; Moya, M.D.L.L.; Stoudt, G.W.; Rodrigueza, W.V.; Phillips, M.C.; Rothblat, G.H. Cyclodextrins as catalysts for the removal of cholesterol from macrophage foam cells. J. Clin. Investig. 1997, 99, 773–780. [Google Scholar] [CrossRef]
- Liu, S.M.; Cogny, A.; Kockx, M.; Dean, R.T.; Gaus, K.; Jessup, W.; Kritharides, L. Cyclodextrins differentially mobilize free and esterified cholesterol from primary human foam cell macrophages. J. Lipid Res. 2003, 44, 1156–1166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmer, S.; Grebe, A.; Bakke, S.S.; Bode, N.; Halvorsen, B.; Ulas, T.; Skjelland, M.; De Nardo, D.; Labzin, L.I.; Kerksiek, A.; et al. Cyclodextrin promotes atherosclerosis regression via macrophage reprogramming. Sci. Transl. Med. 2016, 8, 333ra50. [Google Scholar] [CrossRef] [PubMed]
- Plat, J.; Nichols, J.A.; Mensink, R.P. Plant sterols and stanols: Effects on mixed micellar composition and LXR (target gene) activation. J. Lipid Res. 2005, 46, 2468–2476. [Google Scholar] [CrossRef] [PubMed]
- Jakobsson, T.; Vedin, L.-L.; Hassan, T.; Venteclef, N.; Greco, D.; D’Amato, M.; Treuter, E.; Gustafsson, J.-A.; Steffensen, K.R. The oxysterol receptor LXRβ protects against DSS- and TNBS-induced colitis in mice. Mucosal Immunol. 2014, 7, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Andersen, V.; Christensen, J.; Ernst, A.; Jacobsen, B.A.; Tjonneland, A.; Krarup, H.B.; Vogel, U. Polymorphisms in NF-κB, PXR, LXR, PPARγ and risk of inflammatory bowel disease. World J. Gastroenterol. 2011, 17, 197–206. [Google Scholar] [CrossRef]











© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
van Gorp, C.; de Lange, I.H.; Spiller, O.B.; Dewez, F.; Cillero Pastor, B.; Heeren, R.M.A.; Kessels, L.; Kloosterboer, N.; van Gemert, W.G.; Beeton, M.L.; et al. Protection of the Ovine Fetal Gut against Ureaplasma-Induced Chorioamnionitis: A Potential Role for Plant Sterols. Nutrients 2019, 11, 968. https://doi.org/10.3390/nu11050968
van Gorp C, de Lange IH, Spiller OB, Dewez F, Cillero Pastor B, Heeren RMA, Kessels L, Kloosterboer N, van Gemert WG, Beeton ML, et al. Protection of the Ovine Fetal Gut against Ureaplasma-Induced Chorioamnionitis: A Potential Role for Plant Sterols. Nutrients. 2019; 11(5):968. https://doi.org/10.3390/nu11050968
Chicago/Turabian Stylevan Gorp, Charlotte, Ilse H. de Lange, Owen B. Spiller, Frédéric Dewez, Berta Cillero Pastor, Ron M. A. Heeren, Lilian Kessels, Nico Kloosterboer, Wim G. van Gemert, Michael L. Beeton, and et al. 2019. "Protection of the Ovine Fetal Gut against Ureaplasma-Induced Chorioamnionitis: A Potential Role for Plant Sterols" Nutrients 11, no. 5: 968. https://doi.org/10.3390/nu11050968