Bojungikgi-Tang, a Traditional Herbal Formula, Exerts Neuroprotective Effects and Ameliorates Memory Impairments in Alzheimer’s Disease-Like Experimental Models

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. BJIGT and Reagents
2.2. Apparatus and Chromatographic Conditions
2.3. Preparation of Standard Solution
2.4. Preparation of Sample Solutions
2.5. Calibration Curve and Determination of the Limit of Detection (LOD) and Quantification (LOQ)
2.6. Aβ Aggregation Assay
2.7. Cell-Free BACE Activity Assay
2.8. 2,2′-Azino-Bis-(3-Ethylbenzothiazoline-6-Sulfonic Acid) (ABTS) Radical Scavenging Activity
2.9. 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Radical Scavenging Activity
2.10. Cell Viability Assay
2.11. Animals
2.12. Intracerebroventricular (ICV) Injection of Aβ1-42
2.13. Passive Avoidance Test
2.14. Y-Maze Test
2.15. Brain Tissue Preparation
2.16. Western Blot Analysis
2.17. Immunohistochemistry
2.18. Immunofluorescent Staining
2.19. Statistical Analyses
3. Results
3.1. In Vitro Effects of BJIGT on Aβ Aggregation and BACE Activation
3.2. Neuroprotective Effect of BJIGT in H2O2-Damaged HT22 Hippocampal Cells
3.3. Ameliorating Effect of BJIGT on Memory Impairment in Aβ-Injected AD-Like Mouse Model
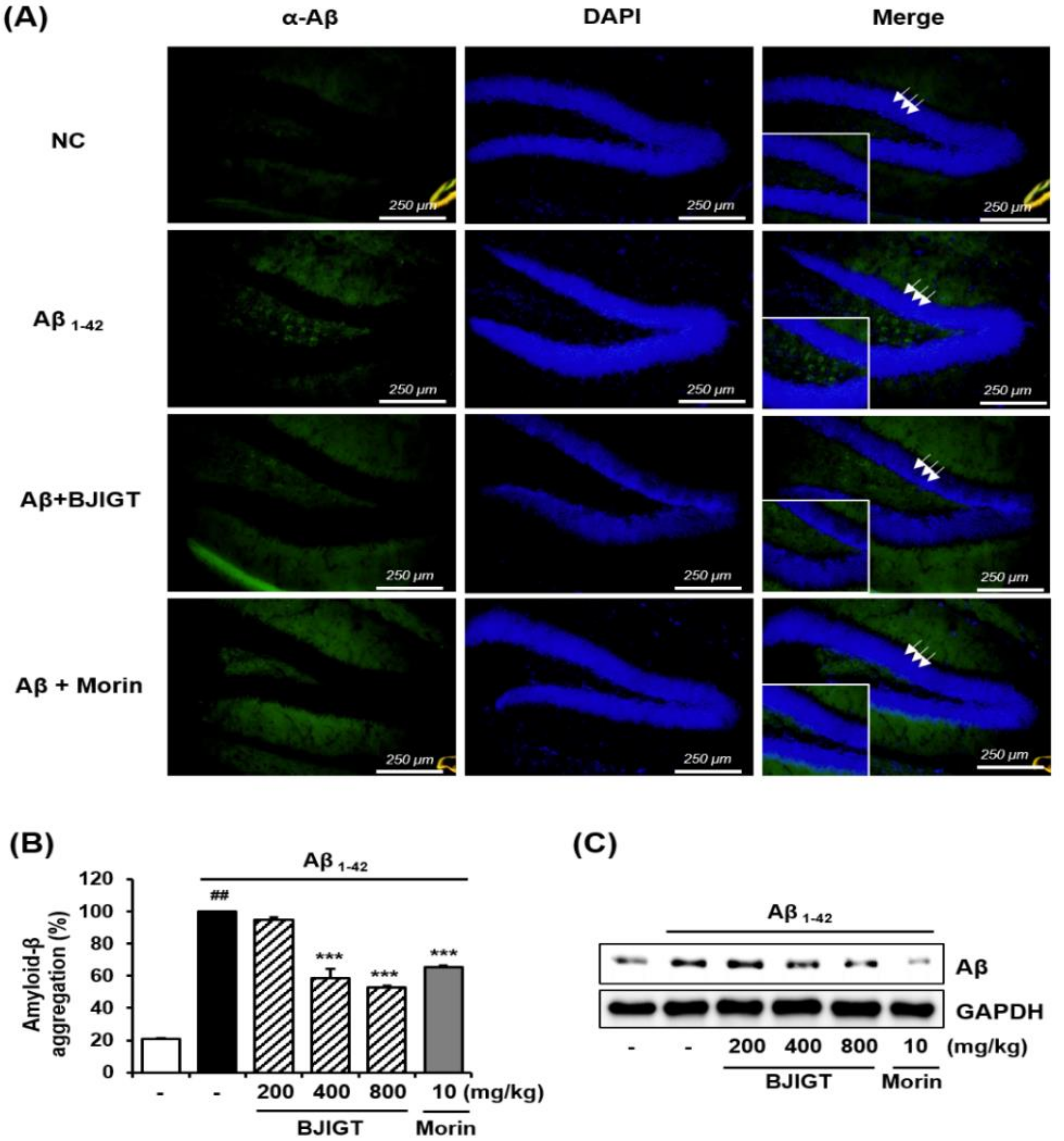
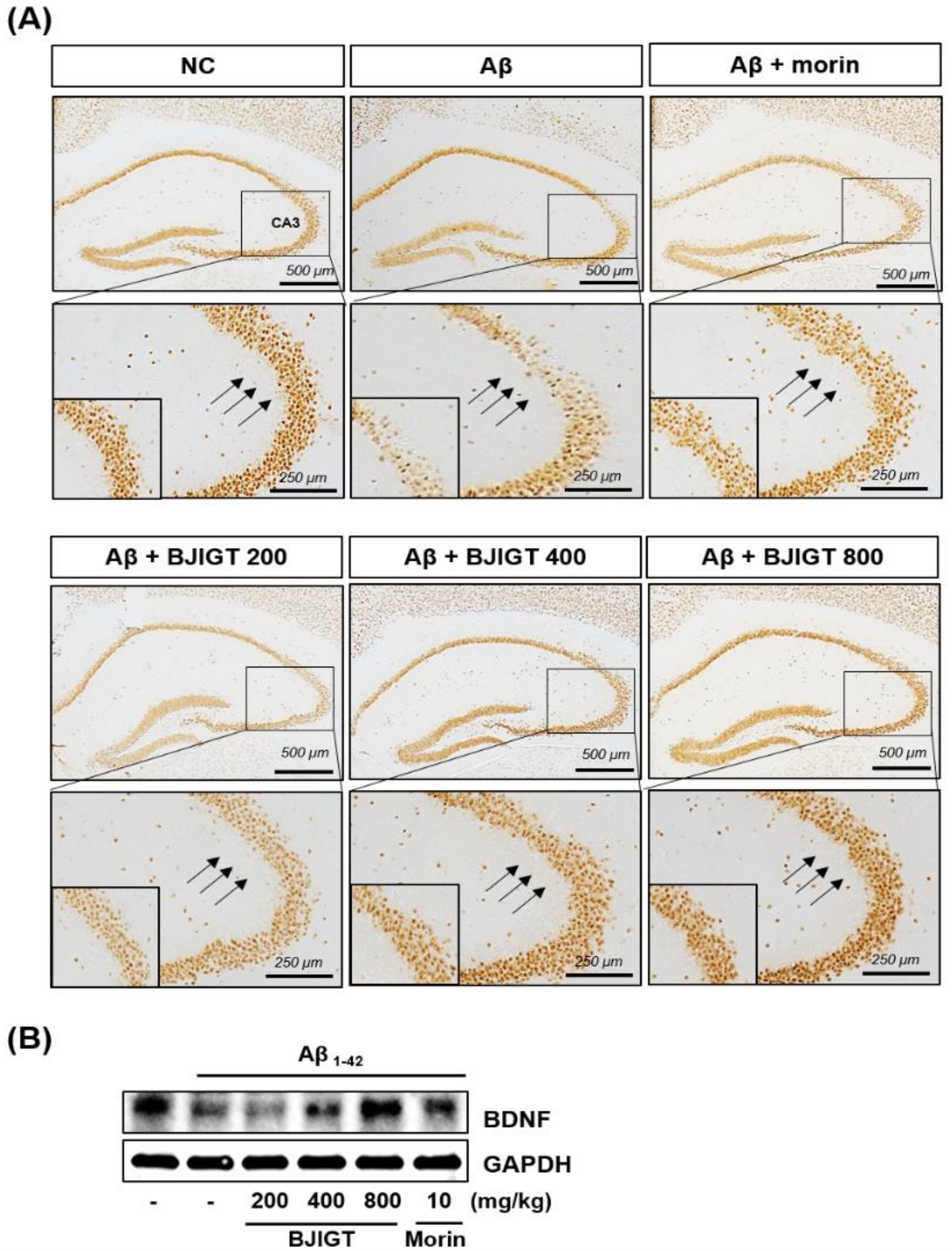
3.4. Effects of BJIGT on Hippocampal Expression of Neuronal Markers and Aβ Accumulation in an Aβ-Injected Ad-Like Mouse Model
3.5. Optimization of HPLC Separation
3.6. Linearity, LOD, and LOQ
3.7. Determination of the Seven Standard Components of BJIGT
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Burns, A.; Iliffe, S. Dementia. BMJ 2009, 338, b75. [Google Scholar] [CrossRef] [PubMed]
- Kavirajan, H.; Schneider, L.S. Efficacy and adverse effects of cholinesterase inhibitors and memantine in vascular dementia: A meta-analysis of randomised controlled trials. Lancet Neurol. 2007, 6, 782–792. [Google Scholar] [CrossRef]
- Herbology TKAoUPo. Herbal Formula Science; Younglimsa: Seoul, Korea, 1999. [Google Scholar]
- Kao, S.T.; Yeh, C.C.; Hsieh, C.C.; Yang, M.D.; Lee, M.R.; Liu, H.S.; Lin, J.G. The Chinese medicine Bu-Zhong-Yi-Qi-Tang inhibited proliferation of hepatoma cell lines by inducing apoptosis via G0/G1 arrest. Life Sci. 2001, 69, 1485–1496. [Google Scholar] [CrossRef]
- Lee, S.E.; Hong, J.E.; Lee, S.H.; Shin, J.Y.; Ro, S.S. Study on Apoptosis Effect and Mechanism by Bojungikki-tang on Human Cancer Cell Line H460. J. Kor. Orient. Intern Med. 2004, 25, 274–288. [Google Scholar]
- Kuo, C.C.; Chen, J.J.; Tsai, J.Y.; Hsueh, C.T. Effects of Chinese herbal medicine in combination with mitomycin C on gastric cancer cells. Biomark. Res. 2014, 2, 26. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Kita, K.; Sato, C.; Kaneda, A. Hochuekkito (Buzhongyiqitang), a herbal medicine, enhances cisplatininduced apoptosis in HeLa cells. Mol. Med. Rep. 2015, 12, 6215–6220. [Google Scholar] [CrossRef] [PubMed]
- Yu, N.; Xiong, Y.; Wang, C. Bu-Zhong-Yi-Qi Decoction, the Water Extract of Chinese Traditional Herbal Medicine, Enhances Cisplatin Cytotoxicity in A549/DDP Cells through Induction of Apoptosis and Autophagy. Biomed. Res. Int. 2017, 2017, 3692797. [Google Scholar] [CrossRef]
- Ishimitsu, R.; Nishimura, H.; Kawauchi, H.; Kawakita, T.; Yoshikai, Y. Dichotomous effect of a traditional Japanese medicine, bu-zhong-yi-qi-tang on allergic asthma in mice. Int. Immunopharmacol. 2001, 1, 857–865. [Google Scholar] [CrossRef]
- Jang, S.I.; Kim, H.J.; Kim, Y.J.; Pae, H.O.; Chung, H.T.; Yun, Y.G.; Jeong, O.S.; Kim, Y.C. Bojungikgitang Inhibits LPS Plus Interferon−gamma-induced inflammatory mediators in RAW 264.7 macrophages. Herb. Formula. Sci. 2003, 11, 115–128. [Google Scholar]
- Ryu, J.H.; Kim, M.S.; Hwang, Y.S.; Yook, C.S. Anxiolytic Effects of the Three Kinds of Traditional Chinese Medicine, Shin - Ki - Hwan, Bo - Jung - Ik - Ki - Tang, and Sa - Mul - Tang, Using the Elevated Plus - maze Test. Biomol. Ther. 2001, 9, 125–130. [Google Scholar]
- Tajima, S.; Bando, M.; Yamasawa, H.; Ohno, S.; Moriyama, H.; Terada, M.; Takada, T.; Suzuki, E.; Gejyo, F.; Sugiyama, Y. Preventive effect of hochu-ekki-to, a Japanese herbal medicine, on bleomycin-induced lung injury in mice. Respirology 2007, 12, 814–822. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.J.; Park, M.Y. Cases Report about Vascular Dementia Treated with Bojungikki-tang-gamibang. Kor. J. Orient. Physiol. Pathol. 2011, 25, 144–148. [Google Scholar]
- Park, K.; Kim, H.J.; Son, J.Y.; Kwak, M.A.; Kim, S.M.; Kim, D.J.; Byun, J.S. Case of Vascular Dementia Treated with Bojungikki-tang-gamibang. Kor. J. Orient. Physiol. Pathol. 2008, 22, 943–947. [Google Scholar]
- Franklin, K.B.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates; Academic press: New York, NY, USA, 2008. [Google Scholar]
- Murphy, M.P.; LeVine, H. 3rd: Alzheimer’s disease and the amyloid-beta peptide. Journal of Alzheimer’s disease. J. Alzheimers Dis. 2010, 19, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Vassar, R. The beta-secretase, BACE: A prime drug target for Alzheimer’s disease. J. Mol Neurosci. 2001, 17, 157–170. [Google Scholar] [CrossRef]
- Huang, W.J.; Zhang, X.; Chen, W.W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef]
- Wolf, H.K.; Buslei, R.; Schmidt-Kastner, R.; Schmidt-Kastner, P.K.; Pietsch, T.; Wiestler, O.D.; Blumcke, I. NeuN: A useful neuronal marker for diagnostic histopathology. J. Histochem Cytochem. 1996, 44, 1167–1171. [Google Scholar] [CrossRef]
- Platenik, J.; Fisar, Z.; Buchal, R.; Jirak, R.; Kitzlerova, E.; Zverova, M.; Raboch, J. GSK3beta, CREB, and BDNF in peripheral blood of patients with Alzheimer’s disease and depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 50, 83–93. [Google Scholar] [CrossRef]
- Prince, M.; Wimo, A.; Guerchet, M.; Ali, G.C.; Wu, Y.T.; Prina, M. World Alzheimer Report 2015: The Global Impact of Dementia: An Analysis of Prevalence, Incidence, Cost and Trends. 2015. Available online: https://www.alz.co.uk/sites/default/files/conf2016/pl12-martin-prince-the-global-impact-of-dementia.pdf (accessed on 1 November 2018).
- Mohammadzadeh Honarvar, N.; Saedisomeolia, A.; Abdolahi, M.; Shayeganrad, A.; Taheri Sangsari, G.; Hassanzadeh Rad, B.; Muench, G. Molecular Anti-inflammatory Mechanisms of Retinoids and Carotenoids in Alzheimer’s Disease: A Review of Current Evidence. J. Mol. Neurosci. 2017, 61, 289–304. [Google Scholar] [CrossRef]
- McGeer, E.G.; McGeer, P.L. Clinically tested drugs for Alzheimer’s disease. Expert. Opin. Investig. Drugs. 2003, 12, 1143–1151. [Google Scholar] [CrossRef]
- Glenner, G.G.; Wong, C.W. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–890. [Google Scholar] [CrossRef]
- Vassar, R.; Bennett, B.D.; Babu-Khan, S.; Kahn, S.; Mendiaz, E.A.; Denis, P.; Teplow, D.B.; Ross, S.; Amarante, P.; Loeloff, R.; et al. Beta-secretase cleavage of Alzheimer’s amyloid precursor protein by the transmembrane aspartic protease BACE. Science 1999, 286, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.G.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Biophys. Acta. 2014, 1842, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Lauderback, C.M. Lipid peroxidation and protein oxidation in Alzheimer’s disease brain: Potential causes and consequences involving amyloid beta-peptide-associated free radical oxidative stress. Free Radic. Biol. Med. 2002, 32, 1050–1060. [Google Scholar] [CrossRef]
- Sultana, R.; Boyd-Kimball, D.; Poon, H.F.; Cai, J.; Pierce, W.M.; Klein, J.B.; Merchant, M.; Markesbery, W.R.; Butterfield, D.A. Redox proteomics identification of oxidized proteins in Alzheimer’s disease hippocampus and cerebellum: An approach to understand pathological and biochemical alterations in AD. Neurobiol. Aging 2006, 27, 1564–1576. [Google Scholar] [CrossRef] [PubMed]
- Butterfield, D.A.; Yatin, S.M.; Varadarajan, S.; Koppal, T. Amyloid beta-peptide-associated free radical oxidative stress, neurotoxicity, and Alzheimer’s disease. Methods Enzymol. 1999, 309, 746–768. [Google Scholar] [PubMed]
- Pal, A.; Badyal, R.K.; Vasishta, R.K.; Attri, S.V.; Thapa, B.R.; Prasad, R. Biochemical, histological, and memory impairment effects of chronic copper toxicity: A model for non-Wilsonian brain copper toxicosis in Wistar rat. Biol. Trace Elem. Res. 2013, 153, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.R.; Lin, H.C.; Su, M.H. Reversal by aqueous extracts of Cistanche tubulosa from behavioral deficits in Alzheimer’s disease-like rat model: Relevance for amyloid deposition and central neurotransmitter function. BMC Complement. Altern. Med. 2014, 14, 202. [Google Scholar] [CrossRef]
- Gold, C.A.; Budson, A.E. Memory loss in Alzheimer’s disease: Implications for development of therapeutics. Expert Rev. Neurother. 2008, 8, 1879–1891. [Google Scholar] [CrossRef]
- Papuć, E.; Kurys-Denis, E.; Krupski, W.; Tatara, M.; Rejdak, K. Can Antibodies against Glial Derived Antigens be Early Biomarkers of Hippocampal Demyelination and Memory Loss in Alzheimer’s Disease? J. Alzheimers Dis. 2015, 48, 115–121. [Google Scholar] [CrossRef]
- Lee, M.Y.; Shin, I.S.; Jeon, W.Y.; Seo, C.S.; Ha, H.; Huh, J.I.; Shin, H.K. Protective effect of Bojungikki-tang, a traditional herbal formula, against alcohol-induced gastric injury in rats. J. Ethnopharmacol. 2012, 142, 346–453. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.A.; Jang, S.; Jun, J.H.; Lee, M.S.; Lee, E.; Kim, N.; Lee, D.H. Herbal medicine (Bojungikki-tang) for allergic rhinitis: A protocol for a systematic review of controlled trials. Medicine 2018, 97, e9551. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.Y.; Shin, C.H.; Shin, T.Y.; Lee, E.J.; Kim, H.M. Effect of bojungikki-tang on lipopolysaccharide-induced cytokine production from peripheral blood mononuclear cells of chronic fatigue syndrome patients. Immunopharmacol. Immunotoxicol. 2003, 25, 491–501. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Jeong, J.S.; Kwon, H.J.; Hong, S.P. Quantification of isoflavonoids and triterpene saponins in Astragali Radix, the root of Astragalus membranaceus, via reverse-phase high-performance liquid chromatography coupled with integrated pulsed amperometric detection. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2017, 1070, 76–81. [Google Scholar] [CrossRef] [PubMed]
- He, J.M.; Zhang, Y.Z.; Luo, J.P.; Zhang, W.J.; Mu, Q. Variation of Ginsenosides in Ginseng of Different Ages. Nat. Prod. Commun. 2016, 11, 739–740. [Google Scholar] [PubMed]
- Chen, L.G.; Jan, Y.S.; Tsai, P.W.; Norimoto, H.; Michihara, S.; Murayama, C.; Wang, C.C. Anti-inflammatory and Antinociceptive Constituents of Atractylodes japonica Koidzumi. J. Agric. Food Chem. 2016, 64, 2254–2262. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Ye, M. Chemical analysis of the Chinese herbal medicine Gan-Cao (licorice). J. Chromatogr. A. 2009, 1216, 1954–1969. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.Y.; Kim, H.M.; Lee, K.H.; Kim, K.Y.; Huang, D.S.; Kim, J.H.; Seong, R.S. Quantitative analysis of marker compounds in Angelica gigas, Angelica sinensis, and Angelica acutiloba by HPLC/DAD. Chem. Pharm. Bull. 2015, 63, 504–511. [Google Scholar] [CrossRef]
- Xia, J.; Kotani, A.; Hakamata, H.; Kusu, F. Determination of hesperidin in Pericarpium Citri Reticulatae by semi-micro HPLC with electrochemical detection. J. Pharm. Biomed. Anal. 2006, 41, 1401–1405. [Google Scholar] [CrossRef]
- Sakai, S.; Kawamata, H.; Kogure, T.; Mantani, N.; Terasawa, K.; Umatake, M.; Ochiai, H. Inhibitory effect of ferulic acid and isoferulic acid on the production of macrophage inflammatory protein-2 in response to respiratory syncytial virus infection in RAW264.7 cells. Mediators Inflamm. 1999, 8, 173–175. [Google Scholar] [CrossRef]
- Yang, F.; Dong, X.; Yin, X.; Wang, W.; You, L.; Ni, J. Radix Bupleuri: A Review of Traditional Uses, Botany, Phytochemistry, Pharmacology, and Toxicology. Biomed. Res. Int. 2017, 2017, 7597596. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Wang, W.; Guo, H.; Zhou, D. Antidepressant-like effect of liquiritin from Glycyrrhiza uralensis in chronic variable stress induced depression model rats. Behav. Brain. Res. 2008, 194, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Hu, X.; Zhao, Z.; Liu, P.; Hu, Y.; Zhou, J.; Wang, Z.; Guo, D.; Guo, H. Antidepressant-like effects of liquiritin and isoliquiritin from Glycyrrhiza uralensis in the forced swimming test and tail suspension test in mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 1179–1184. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.A.; Wang, J.L.; Liu, R.T.; Ren, J.P.; Wen, L.Q.; Chen, X.J.; Bian, G.X. Liquiritin potentiate neurite outgrowth induced by nerve growth factor in PC12 cells. Cytotech 2009, 60, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Tang, Y.; Wu, A.; Liu, T.; Dai, X.; Zheng, Q.; Wang, Z.B. Neuroprotective effect of liquiritin against focal cerebral ischemia/reperfusion in mice via its antioxidant and antiapoptosis properties. J. Asian Nat. Prod. Res. 2010, 12, 1051–1060. [Google Scholar] [CrossRef] [PubMed]
- Song, J.H.; Lee, J.W.; Shim, B.; Lee, C.Y.; Choi, S.; Kang, C.; Sohn, N.W.; Shin, J.W. Glycyrrhizin Alleviates Neuroinflammation and Memory Deficit Induced by Systemic Lipopolysaccharide Treatment in Mice. Molecules 2013, 18, 15788–15803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Li, W.; Jung, S.W.; Lee, Y.W.; Kim, Y.H. Protective effects of decursin and decursinol angelate against amyloid β-protein-induced oxidative stress in the PC12 cell line: The role of Nrf2 and antioxidant enzymes. Biosci. Biotechnol. Biochem. 2011, 75, 434–442. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Linear Range (μg/mL) | Regression Equation (y = ax+b) a | Correlation Coefficient (r2) | LOD b (μg/mL) | LOQ c (μg/mL) | |
|---|---|---|---|---|---|---|
| Slope (a) | Intercept (b) | |||||
| Liquiritin apioside | 1.56–100 | 14986 | 3467.2 | 1.0000 | 0.228 | 0.692 |
| Liquiritin | 0.39–25 | 18335 | 1582.3 | 1.0000 | 0.052 | 0.157 |
| Nodakenin | 0.78–50 | 24593 | 3093.5 | 0.9999 | 0.189 | 0.574 |
| Hesperidin | 1.56–100 | 17953 | 1857.5 | 0.9999 | 0.441 | 1.336 |
| Glycyrrhizin | 6.25–400 | 5195.6 | 1474.1 | 1.0000 | 0.797 | 2.415 |
| Decursin | 0.78–50 | 32417 | 10204 | 0.9997 | 0.221 | 0.669 |
| Decursinol angelate | 0.39–25 | 35549 | 4903.6 | 0.9997 | 0.108 | 0.328 |
| Compound | Content (mg/g) |
|---|---|
| Liquiritin apioside | 0.400 ± 0.001 |
| Liquiritin | 0.110 ± 0.000 |
| Nodakenin | 0.158 ± 0.000 |
| Hesperidin | 0.487 ± 0.001 |
| Glycyrrhizin | 1.766 ± 0.003 |
| Decursin | 0.136 ± 0.000 |
| Decursinol angelate | 0.109 ± 0.001 |
| Herbal Medicine | Scientific Name of Plant Source | Family | Origin |
|---|---|---|---|
| Astragali Radix | Astragalus membranaceus Bunge | Fabaceae | South Korea |
| Ginseng Radix | Panax ginseng C. A. Meyer | Araliaceae | South Korea |
| Atractylodis Rhizoma Alba | Atractylodes japonica Koidz. | Compositae | China |
| Glycyrrhizae Radix et Rhizoma | Glycyrrhiza uralensis Fischer | Fabaceae | China |
| Angelicae Gigantis Radix | Angelica gigas Nakai | Apiaceae | South Korea |
| Citri Unshius Pericarpium | Citrus reticulata Blanco | Rutaceae | South Korea |
| Cimicifugae Rhizoma | Cimicifuga heracleifolia Kom. | Ranunculaceae | China |
| Bupleuri Radix | Bupleurum falcatum L. | Apiaceae | China |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, H.-S.; Kim, Y.J.; Sohn, E.; Yoon, J.; Kim, B.-Y.; Jeong, S.-J. Bojungikgi-Tang, a Traditional Herbal Formula, Exerts Neuroprotective Effects and Ameliorates Memory Impairments in Alzheimer’s Disease-Like Experimental Models. Nutrients 2018, 10, 1952. https://doi.org/10.3390/nu10121952
Lim H-S, Kim YJ, Sohn E, Yoon J, Kim B-Y, Jeong S-J. Bojungikgi-Tang, a Traditional Herbal Formula, Exerts Neuroprotective Effects and Ameliorates Memory Impairments in Alzheimer’s Disease-Like Experimental Models. Nutrients. 2018; 10(12):1952. https://doi.org/10.3390/nu10121952
Chicago/Turabian StyleLim, Hye-Sun, Yu Jin Kim, Eunjin Sohn, Jiyeon Yoon, Bu-Yeo Kim, and Soo-Jin Jeong. 2018. "Bojungikgi-Tang, a Traditional Herbal Formula, Exerts Neuroprotective Effects and Ameliorates Memory Impairments in Alzheimer’s Disease-Like Experimental Models" Nutrients 10, no. 12: 1952. https://doi.org/10.3390/nu10121952