The Inhibitory Effect of Ojeoksan on Early and Advanced Atherosclerosis

 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
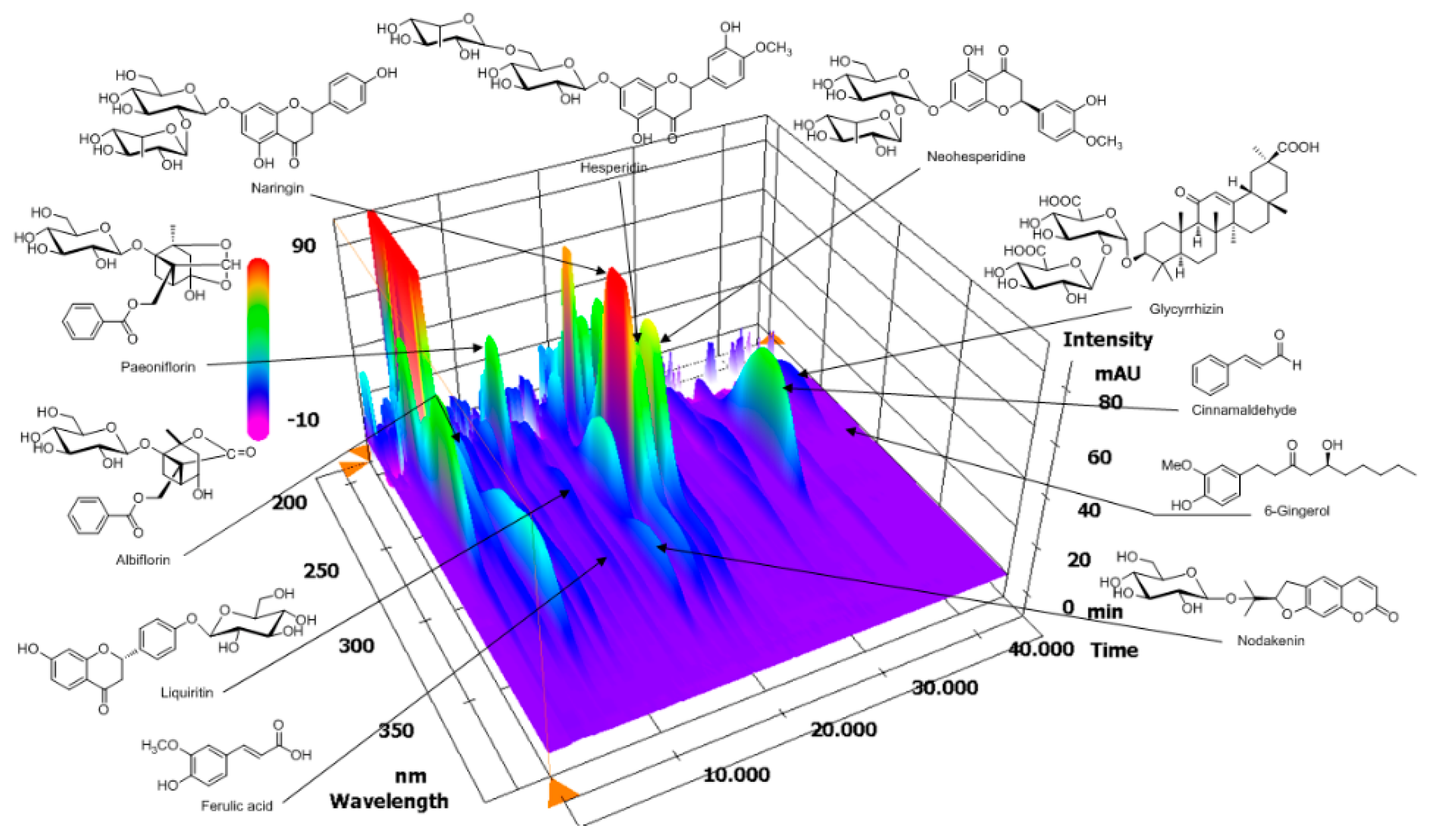
2.1. High-Performance Liquid Chromatography (HPLC) Analysis of OJS
2.2. Experimental Animals
2.3. Cell Cultures
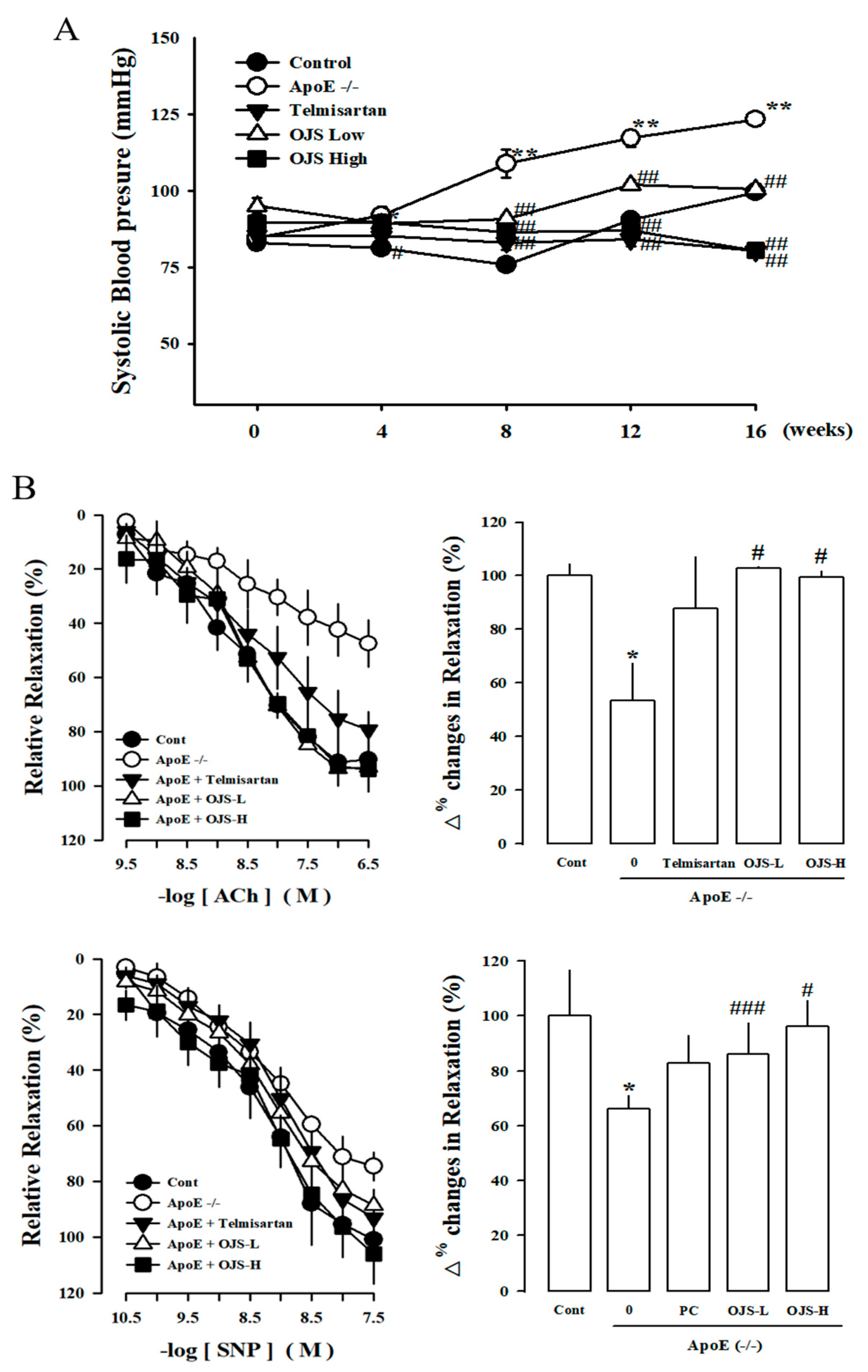
2.4. Measurement of Systolic Blood Pressure
2.5. Plasma Biochemical Analysis
2.6. Preparation of Carotid Artery Samples and Measurement of Vascular Reactivity
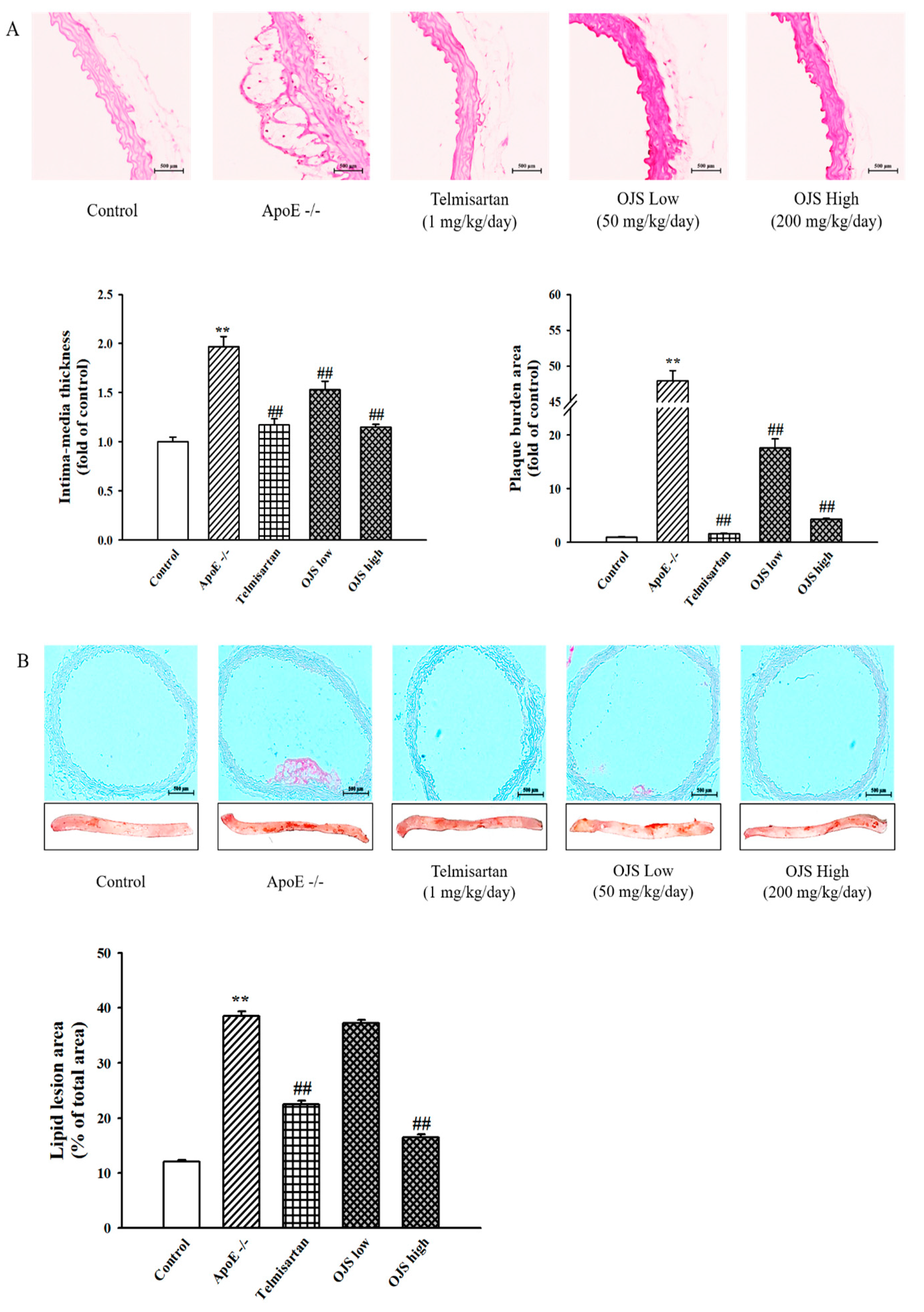
2.7. Histopathological Staining of Aorta
2.8. Measurement of Atherosclerotic Lesions by Oil Red O Staining
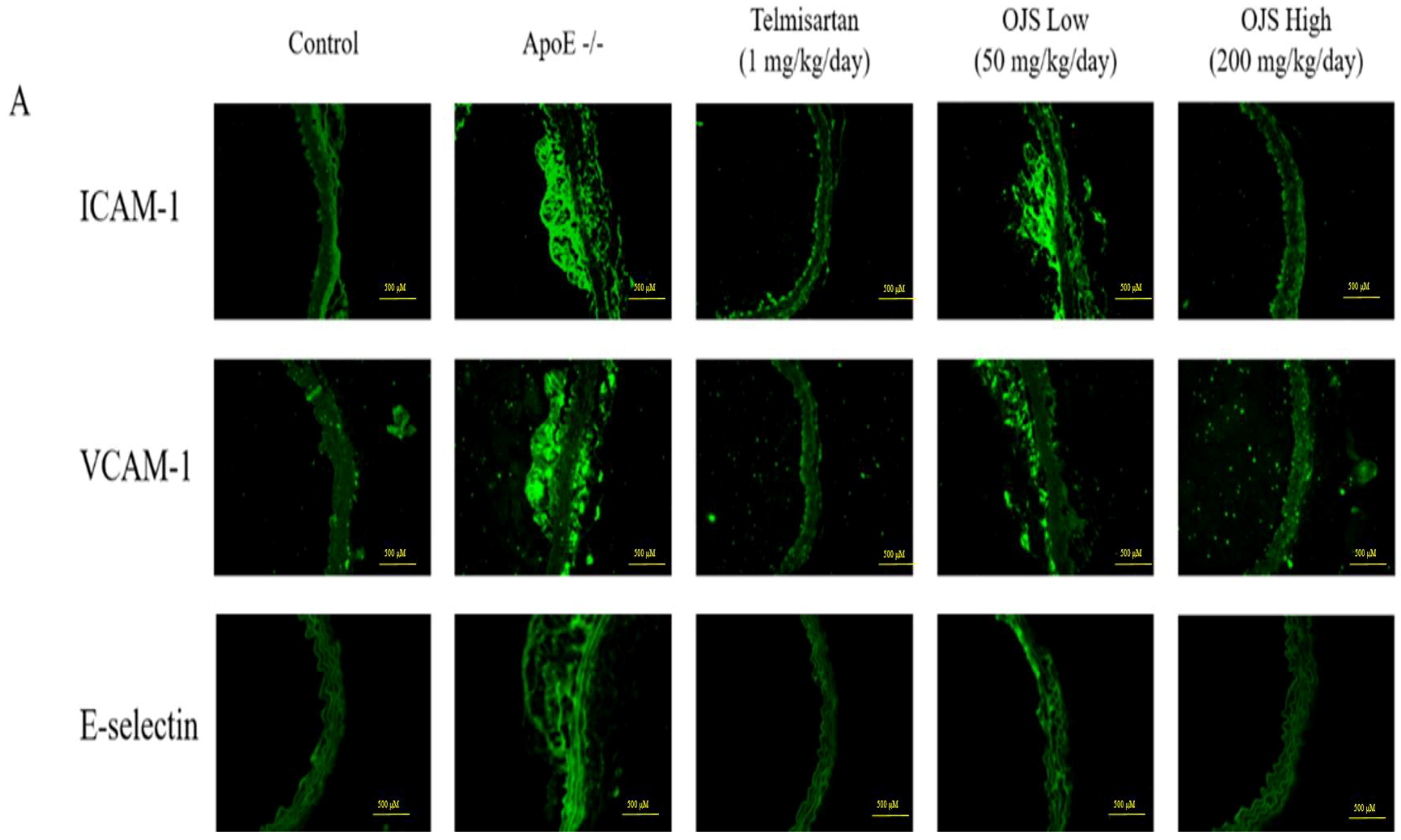
2.9. Immunofluorescence
2.10. Western Blot Analysis
2.11. RNA Preparation and Quantitative Real-Time Reverse Transcription-PCR (Real-Time RT-qPCR)
2.12. Measurement of NO Production Using Griess Reagent System
2.13. Fluorescence Microscopy
2.14. Statistical Analysis
3. Results
3.1. HPLC Analysis of OJS
3.2. The Effect of OJS on Food Intake and Body Weight
3.3. The Effect of OJS on Lipid Parameters in ApoE −/− Mice
3.4. Effect of OJS on Vascular Dysfunction in ApoE −/− Mice
3.5. Effect of OJS on Atherosclerotic Lesions in ApoE −/− Mice
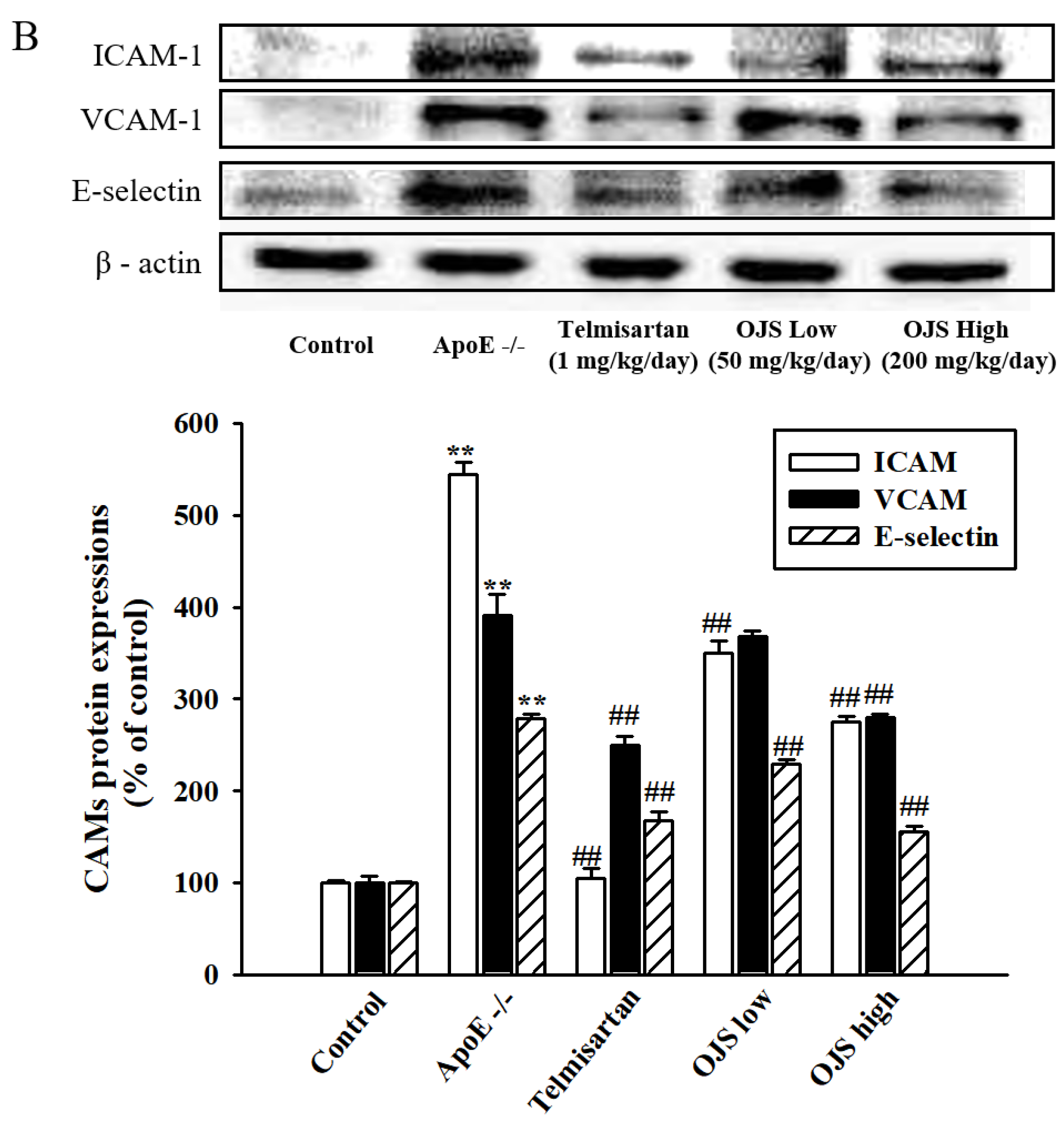
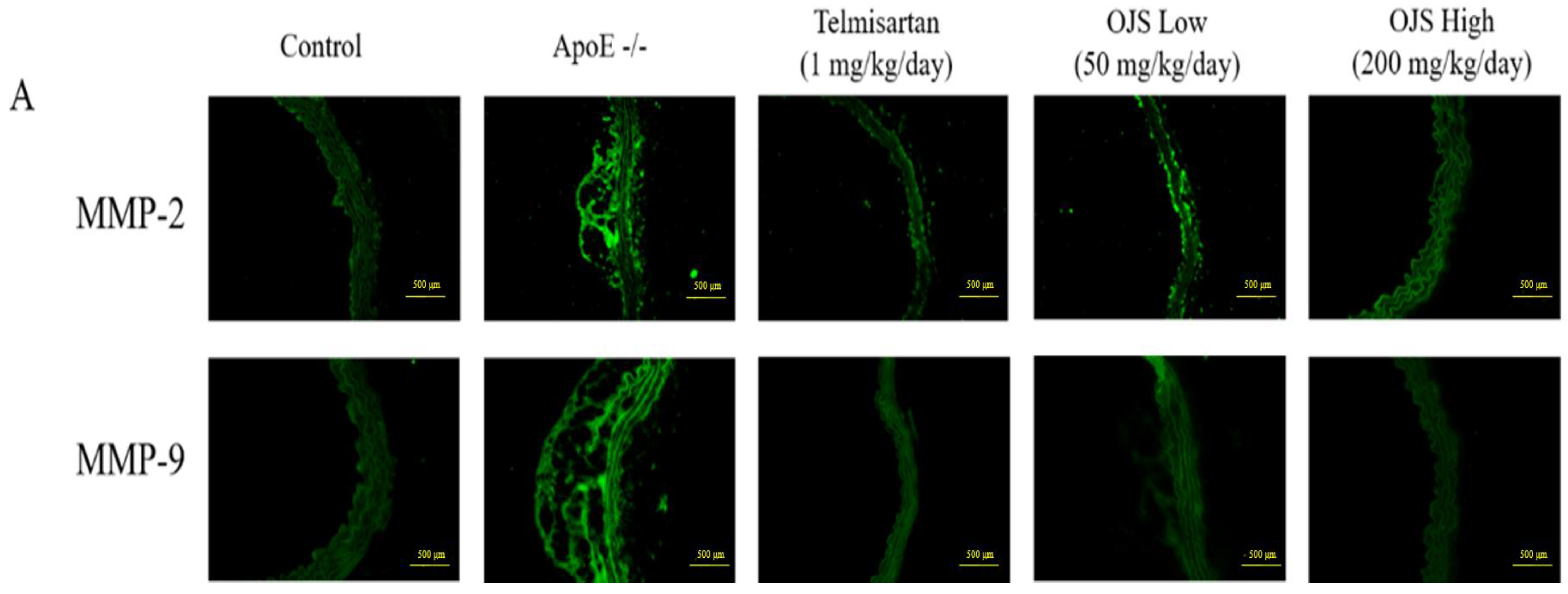
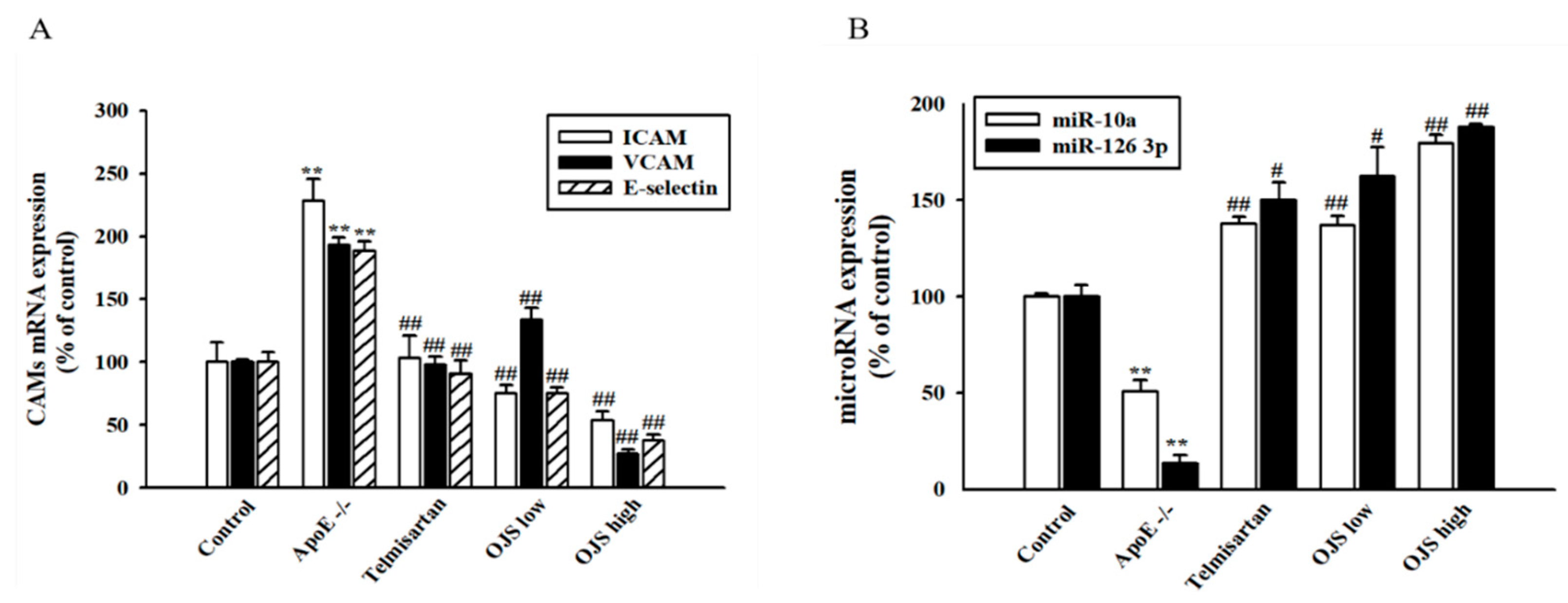
3.6. Effect of OJS on Vascular Inflammation in ApoE −/− Mice
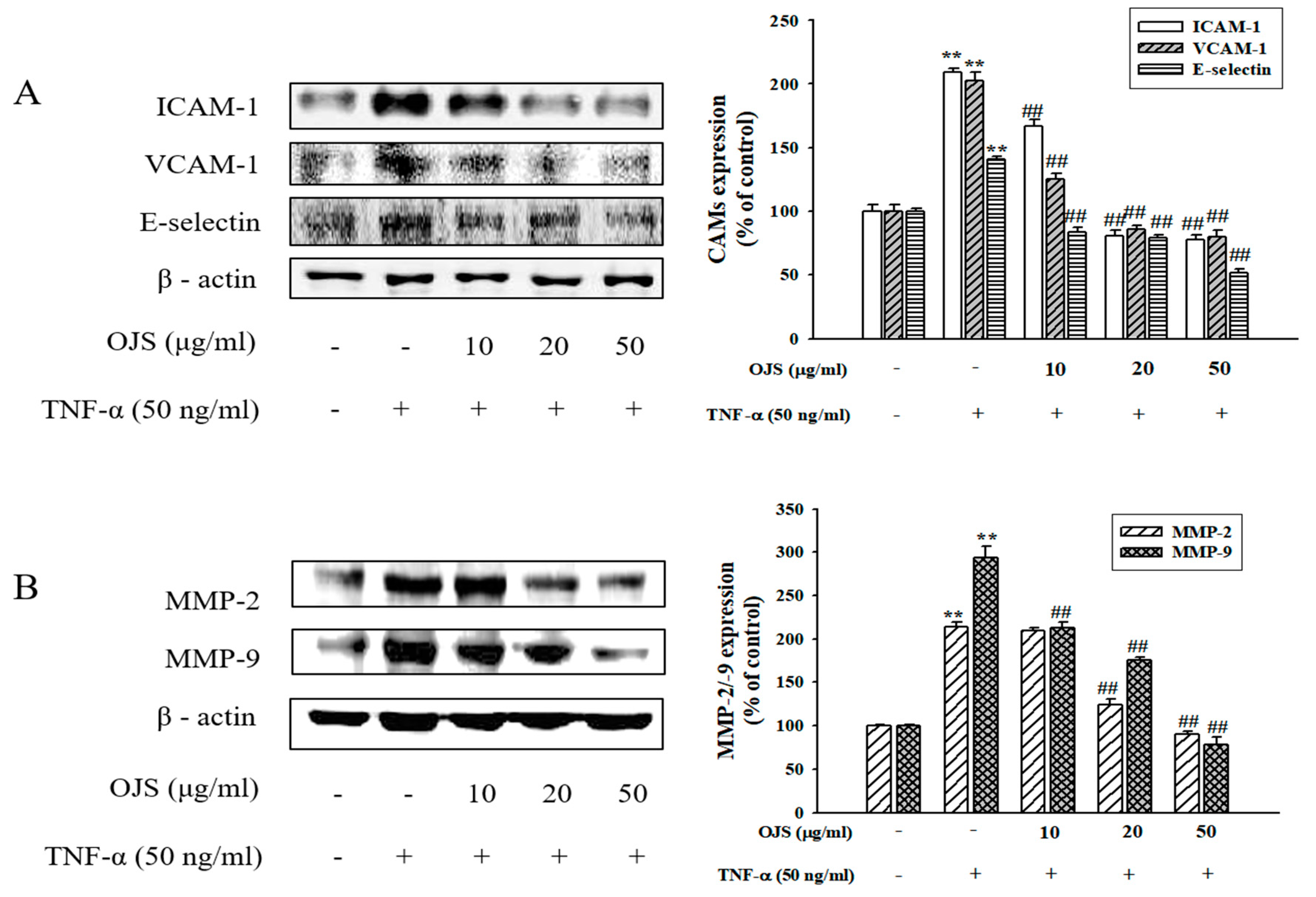
3.7. Effects of OJS on TNF-α-Induced Adhesion Molecules and MMPs Expression in HUVECs
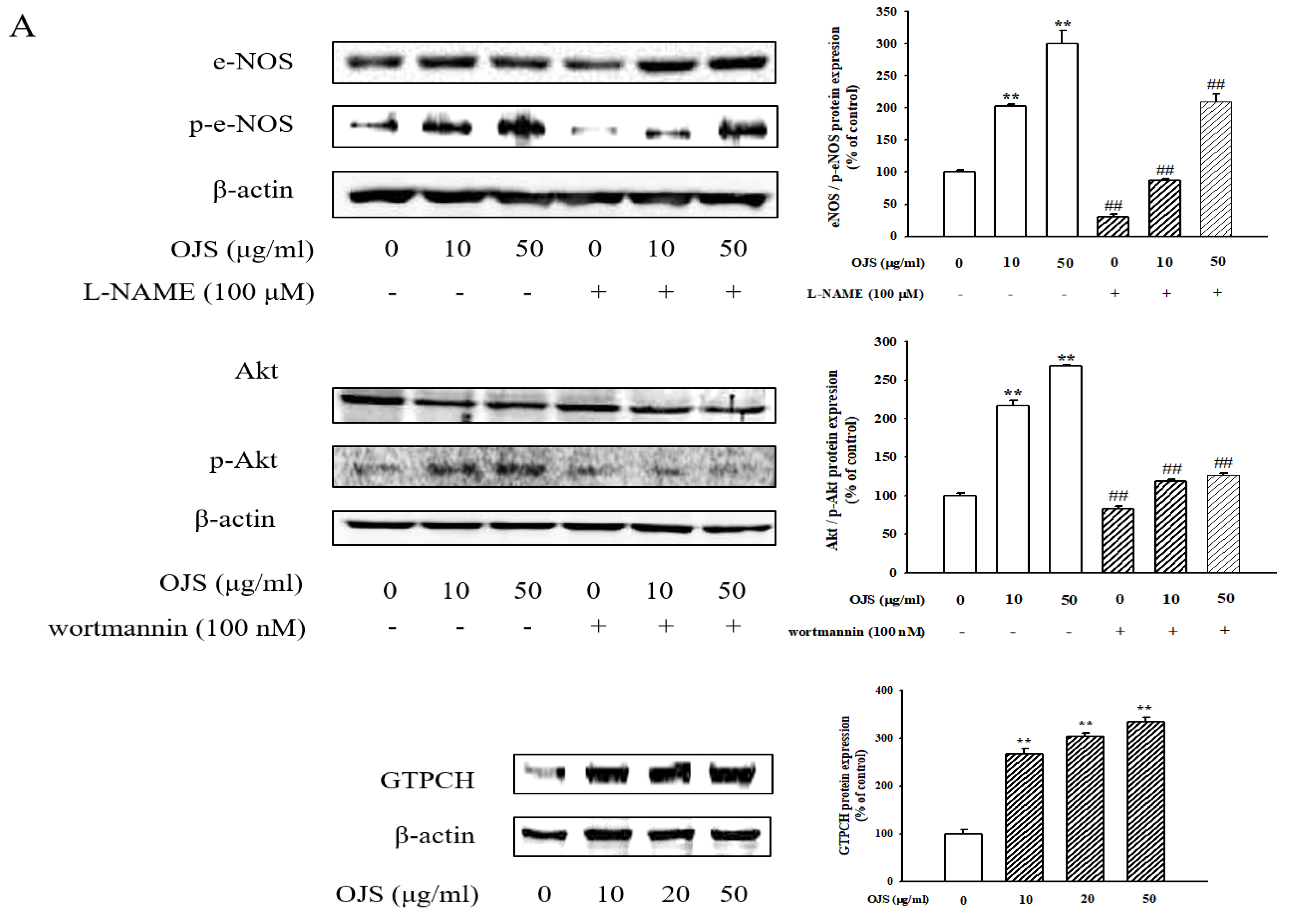
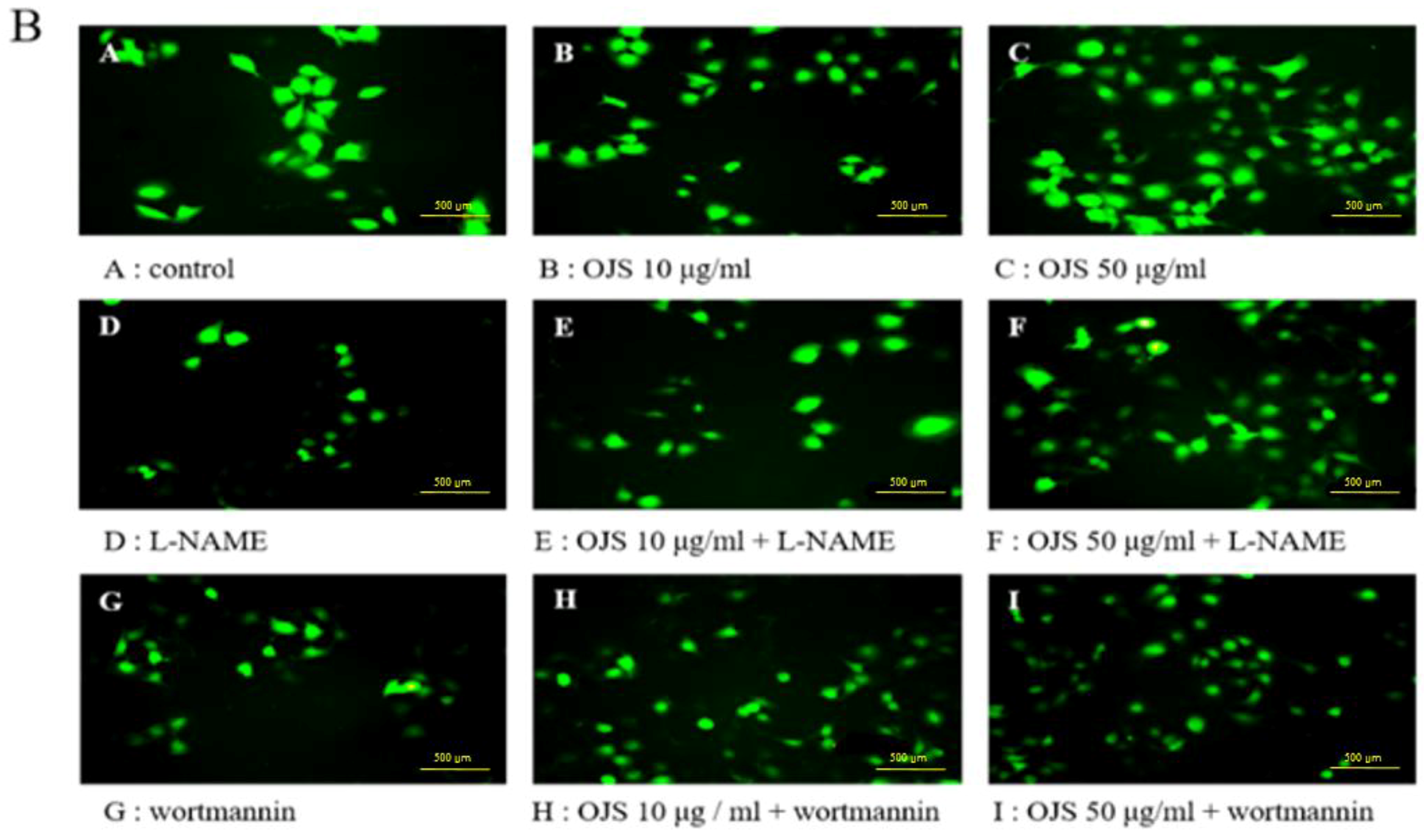
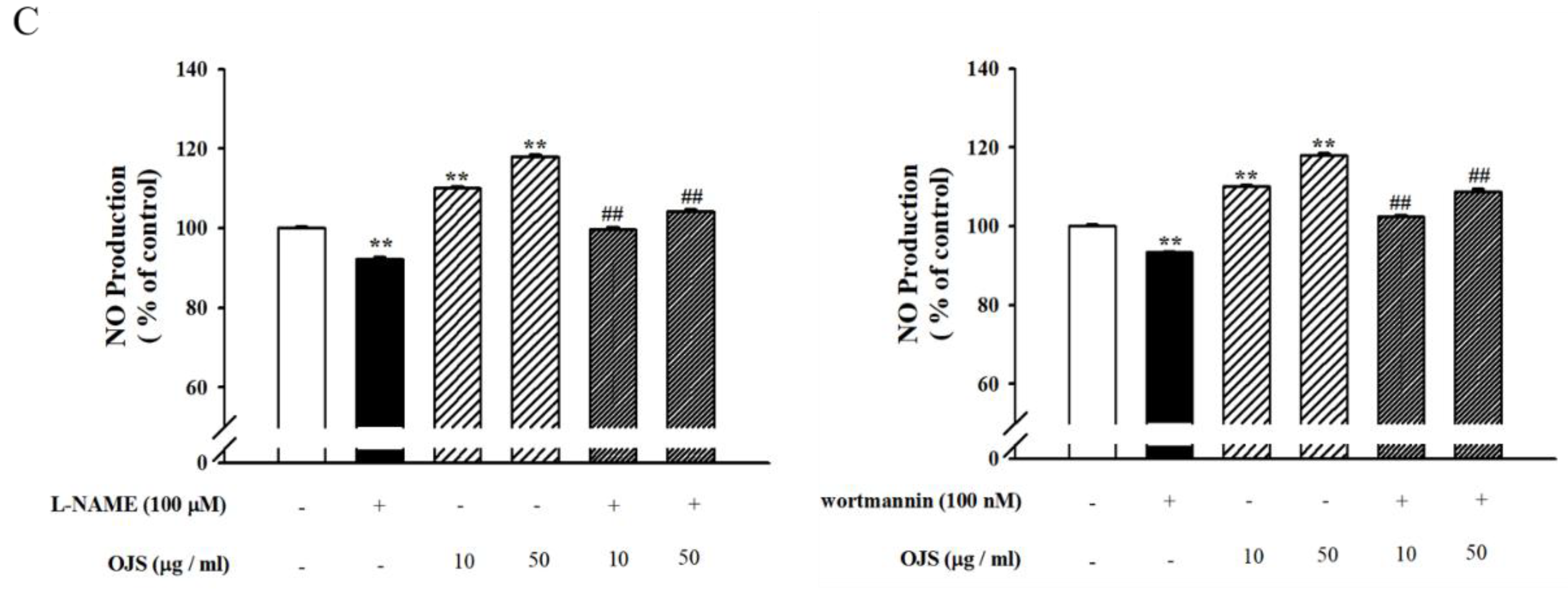
3.8. OJS Regulates the Akt/eNOS-NO Pathway in HUVECs
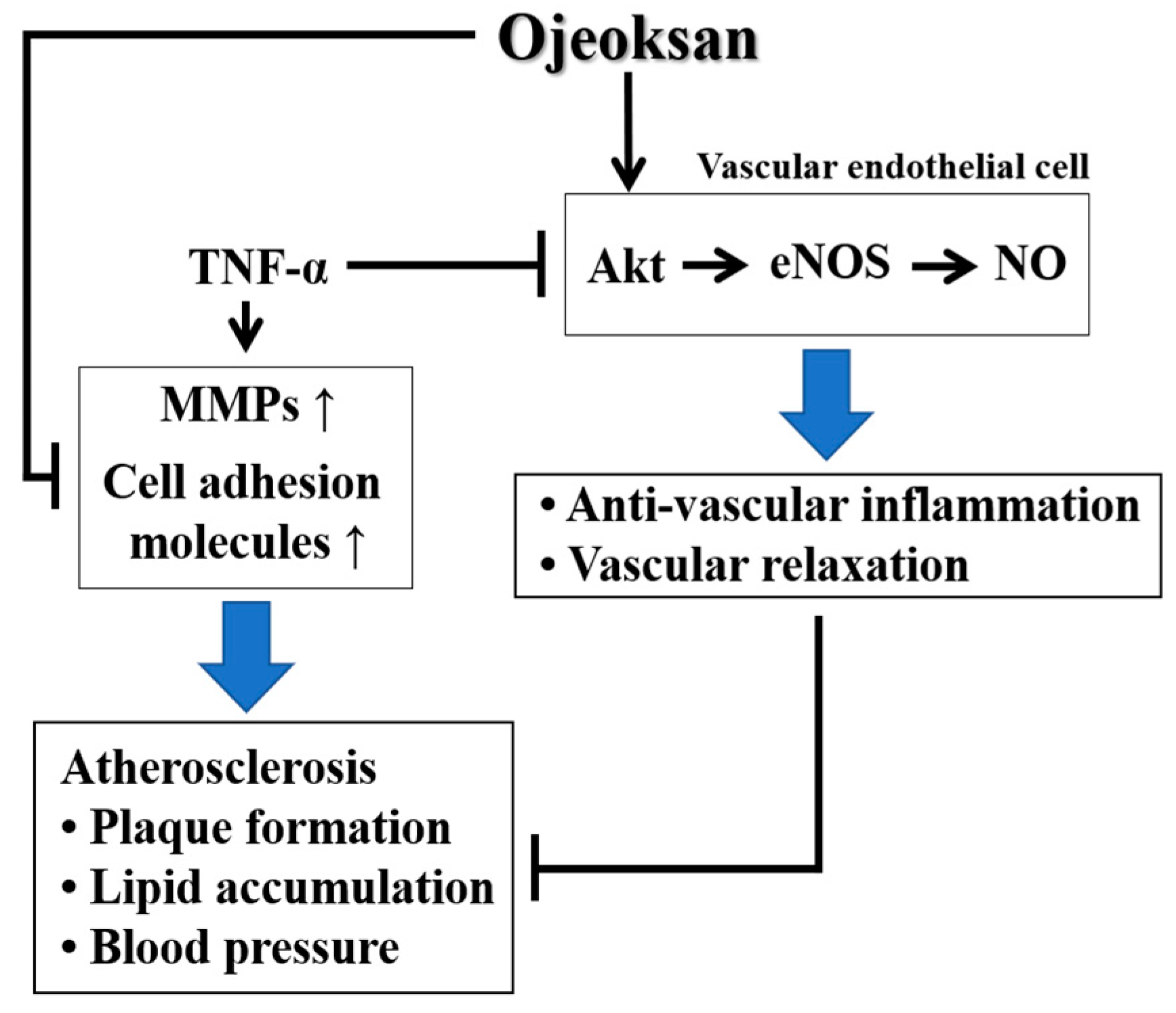
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Tedgui, A.; Mallat, Z. Cytokines in atherosclerosis: Pathogenic and regulatory pathways. Physiol. Rev. 2006, 86, 515–581. [Google Scholar] [CrossRef] [PubMed]
- Libby, P. Inflammation in atherosclerosis. Arterioscler. Thrombosi Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.E.; Greene, L.; Goodfriend, T.L. Insulin resistance and ardiovascular disease. Am. J. Hypertens. 2001, 14, 116–125. [Google Scholar] [CrossRef]
- Blankenberg, S.; Barbaux, S.; Tiret, L. Adhesion molecules and atherosclerosis. Atherosclerosis 2003, 170, 191–203. [Google Scholar] [CrossRef]
- Sana, T.R.; Janatpour, M.J.; Sathe, M.; McEvoy, L.M.; McClanahan, T.K. Microarray analysis of primary endothelial cells challenged with different inflammatory and immune cytokines. Cytokine 2005, 29, 256–269. [Google Scholar] [CrossRef] [PubMed]
- Suarez, Y.; Wang, C.; Manes, T.D.; Pober, J.S. Cutting edge: TNF-induced micrornas regulate TNF-induced expression of e-selectin and intercellular adhesion molecule-1 on human endothelial cells: Feedback control of inflammation. J. Immunol. 2010, 184, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Harris, T.A.; Yamakuchi, M.; Ferlito, M.; Mendell, J.T.; Lowenstein, C.J. Microrna-126 regulates endothelial expression of vascular cell adhesion molecule 1. Proc. Natl. Acad. Sci. USA 2008, 105, 1516–1521. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Shi, C.; Manduchi, E.; Civelek, M.; Davies, P.F. Microrna-10a regulation of proinflammatory phenotype in athero-susceptible endothelium in vivo and in vitro. Proc. Natl. Acad. Sci. USA 2010, 107, 13450–13455. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, M. Biochemistry and molecular cell biology of diabetic complications. Nature 2001, 65, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ho, F.M.; Liu, S.H.; Lin, W.W.; Liau, C.S. Opposite effects of high glucose on MMP-2 and TMIP-2 in human endothelial cells. J. Cell. Biochem. 2007, 101, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.V.; Shahnaz, R.; Diana, N.; Shipeng, Y.; Srikanth, G.; Suresh, C.T. Matrix metalloproteinases in atherosclerosis: Role of nitric oxide, hydrogen sulfide, homocysteine, and polymorphisms. Vasc. Health Risk Manag. 2015, 11, 173–183. [Google Scholar]
- Mariotto, S.; Menegazzi, M.; Suzuki, H. Biochemical aspects of nitric oxide. Curr. Pharm. Des. 2004, 10, 1627–1645. [Google Scholar] [CrossRef] [PubMed]
- Colasanti, M.; Persichini, T. Nitric oside: An inhibitor of NF-kappaB/rel system in glial cells. Brain Res. Bull. 2000, 10, 1627–1645. [Google Scholar]
- Susanne, K.; Philip, W.; Ari, W.; Thomas, M.; Andreas, D. eNOS uncoupling in cardiovascular disease–the role of oxidative stress and inflammation. Curr. Pharm. Des. 2014, 20, 3576–3594. [Google Scholar]
- Mark, J.C.; Ashley, B.H.; Keith, M.C. Dihydrofolate reductase protects endothelial nitric oxide synthase from uncoupling in tetrahydrobiopterin deficiency. Free Radic. Biol. Med. 2011, 50, 1639–1646. [Google Scholar]
- Fulton, D.; Gratton, J.P.; McCabe, T.J.; Fontana, J.; Fujio, Y.; Walsh, K.; Franke, T.F.; Papapetropoulos, A.; Sessa, W.C. Regulation of endothelium-derived nitric oxide production by the protein kinase Akt. Nature 1999, 399, 597–601. [Google Scholar] [CrossRef] [PubMed]
- Emmanouil, C.; Elisabeth, D.; Corinna, H.; Ulrich, F.M.; Andreas, M.Z.; Stefanie, D. Oxidized LDL Inhibits Vascular Endothelial Growth Factor–Induced Endothelial Cell Migration by an Inhibitory Effect on the Akt/Endothelial Nitric Oxide Synthase Pathway. Circulation 2001, 103, 2102–2107. [Google Scholar]
- National Health Insurance Corporation, Health Insurance Review & Assessment Service. 2010 National Health Insurance Statistical Yearbook; National Health Insurance Corporation, Health Insurance Review & Assessment Service: Seoul, Korea, 2011; pp. 300–301. [Google Scholar]
- Lee, S.Y.; Park, W.H.; Cha, Y.Y.; Lee, E. Effects of Ojeoksangamibang Extract on the Recovery of Liver Function in CCl4-exposed Rats. J. Korean Med. Rehabil. 2013, 23, 45–53. [Google Scholar]
- Nakashima, Y.; Plump, A.S.; Raines, E.W.; Breslow, J.L.; Ross, R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arterioscler. Thrombosi Vasc. Biol. 1994, 14, 133–140. [Google Scholar] [CrossRef]
- Fukuda, D.; Enomoto, S.; Hirata, Y.; Nagai, R.; Sata, M. The angiotensin receptor blocker, telmisartan, reduces and stabilizes atherosclerosis in ApoE and AT1aR double deficient mice. Biomed. Pharmacother. 2010, 64, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Angeli, V.; Llodra, J.; Rong, J.X.; Satoh, K.; Ishii, S.; Shimizu, T.; Fisher, E.A.; Randolph, G.J. Dyslipidemia associated with atherosclerotic disease systemically alters dendritic cell mobilization. Immunity 2004, 21, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Lauer, R.M.; Lee, J.; Clarke, W.R. Factors Affecting the Relationship between Childhood and Adult Cholesterol Levels: The Muscatine Study. Pediatrics 1988, 8, 309–321. [Google Scholar]
- Tall, A.R. Cholesterol efflux pathways and other potential mechanisms involved in the athero-protective effect of high density lipoproteins. J. Intern. Med. 2008, 263, 256–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Voelkel, N.F.; Tuder, R.M. Hypoxia-induced pulmonary vascular remodeling: A model for what human disease. J. Clin. Investig. 2000, 106, 733–738. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Jeong, T.S.; Kim, D.Y.; Kim, Y.M.; Na, H.J.; Nam, K.H.; Lee, S.B.; Kim, H.C.; Oh, S.R.; Choi, Y.K.; et al. Hematein inhibits atherosclerosis by inhibition of reactive oxygen generation and NF-κB-dependent inflammatory mediators in hyperlipidemic mice. J. Cardiovasc. Pharmacol. 2003, 42, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, H.; Anzai, T.; Morisawa, M. Resveratrol prevents the development of abdominal aortic aneurysm through attenuation of inflammation, oxidative stress, and neovascularization. Atherosclerosis 2011, 217, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Tuttolomondo, A.; Pecoraro, R.; Pinto, A. Studies of selective TNF inhibitors in the treatment of brain injury from stroke and trauma: A review of the evidence to date. Drug Des. Dev. Ther. 2014, 7, 2221–2238. [Google Scholar] [CrossRef] [PubMed]
- Diamant, M.; Tushuizen, M.E. The metabolic syndrome and endothelial dysfunction: Common highway to type 2 diabetes and CVD. Curr. Diab. Rep. 2006, 6, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Stroes, E.; Kastelein, J.; Cosentino, F. Tetrahydrobiopterin restores endothelial function in hypercholesterolemia. J. Clin. Investig. 1997, 99, 41–46. [Google Scholar] [CrossRef] [PubMed]
- Chuaiphichai, S.; McNeill, E.; Douglas, G.; Crabtree, J.M.; Bendall, K.J.; Hale, A.B.; Alp, J.N.; Channon, M.K. Cell autonomous role of endothelial GTP Cyclohydrolase 1 and tetrahydrobiopterin in blood pressure regulation. Hypertension 2014, 64, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Mark, J.C.; Amy, L.T.; Yasir, A.; Nicholas, W.; Ashley, B.H.; Shijie, C.; Keith, M.C.; Nicholas, J.A. Quantitative regulation of intracellular indothelial nitric-oxide synthase (eNOS) coupling by both tetrahydrobiopterin-eNOS stoichiometry and biopterin redox status insights from cells with the-regulated GTP cyclohydrolase I expression. J. Biol. Chem. 2008, 284, 1134–1144. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Latin Name | Scientific Name | Amount (g) | Origin |
|---|---|---|---|
| Atractylodis Rhizoma | Atractylodes lancea DC | 7.5 | China |
| Ephedrae Herba | Ephedra sinica Stapf | 3.7 | China |
| Citri unshii Percarpium | Citrus reticulata Blanco | 3.7 | Jeju, Korea |
| Magnoliae Cortex | Magnolia officinalis Rehder & E.H. Wilson | 3.0 | China |
| Platycodi Radix | Platycodon grandiflorus (Jacq.) A. DC | 3.0 | Yeongcheon, Korea |
| Aurantii Fructus Immaturus | Citrus aurantium L | 3.0 | China |
| Angelicae Gigantis Radix | Angelica gigas Nakai | 3.0 | Pyeongchang, Korea |
| Zingiberis Rhizoma | Zingiber officinale Roscoe | 3.0 | Yeongcheon, Korea |
| Paeoniae Radix | Paeonia lactiflora Pall | 3.0 | Hwasun, Korea |
| Poria Sclerotium | Wolfiporia extensa | 3.0 | Yeongcheon, Korea |
| Angelicae Dahuricae Radix | Angelica dahurica (Hoffm.) Benth. & Hook.f. ex Franch. & Sav | 2.6 | Yeongcheon, Korea |
| Cnidii Rhizoma | Ligusticum officinale (Makino) Kitag | 2.6 | Yeongcheon, Korea |
| Pinelliae Tuber | Pinellia ternata (Thunb.) Ten. ex Breitenb | 2.6 | China |
| Cinnamomi Cortex | Cinnamomum cassia (L.) J. Presl | 2.6 | Vietnam |
| Glycyrrhizae Radix et Rhizoma | Glycyrrhiza uralensis Fisch | 2.2 | China |
| Zingiberis Rhizoma recens | Zingiber officinale Roscoe | 3.7 | Hanam, Korea |
| Allii Fistulosi Bulbus | Allium fistulosum L | 3.7 | Hanam, Korea |
| Product | D12450B | D12079B | ||
|---|---|---|---|---|
| gm % | kcal % | gm % | kcal % | |
| Protein | 19.2 | 20 | 20 | 17 |
| Carbohydrate | 67.3 | 70 | 50 | 43 |
| Fat | 4.3 | 10 | 21 | 40 |
| Total | 100 | 100 | ||
| kcal/gm | 3.85 | 4.7 | ||
| Ingredient | gm | kcal | gm | kcal |
| Casein, 80 Mesh | 200 | 195 | ||
| dl-Methionine | 3 | |||
| l-Cystine | 3 | |||
| Corn Starch | 315 | 50 | ||
| Maltodextrin 10 | 35 | 100 | ||
| Sucrose | 350 | 341 | ||
| cellulose | 50 | 50 | ||
| Milk Fat, Anhydrous | 200 | |||
| Corn oil | 10 | |||
| Soybean oil | 25 | |||
| Lard | 20 | |||
| Mineral Mix S10026 | 10 | 35 | ||
| Dicalcium Phosphate | 13 | |||
| Calcium Carbonate | 5.5 | 4 | ||
| Potassium Citrate, 1 H2O | 16.5 | |||
| Vitamin Mix V10001 | 10 | 10 | ||
| Choline Bitartrate | 2 | 2 | ||
| Cholesterol, USP | 0.05 | 1.5 | ||
| FD&C Yellow Dye #5 | ||||
| Total | 1055.05 | 4057 | 1001.54 | 4686 |
| Food Intake (g) | Body Weight (g) | T-Cho (mg/dL) | TG (mg/dL) | LDL/VLDL-Cho (mg/dL) | HDL-Cho (mg/dL) | Atherogenic Index | |
|---|---|---|---|---|---|---|---|
| Control | 3.90 ± 0.12 | 29.26 ± 0.92 | 64.69 ± 2.89 | 78.71 ± 1.25 | 27.51 ± 3.31 | 46.91 ± 1.04 | 0.38 ± 0.06 |
| ApoE −/− | 3.82 ± 0.42 | 33.62 ± 1.32 ** | 271.78 ± 9.79 ** | 122.30 ± 1.90 ** | 128.58 ± 5.03 ** | 20.02 ± 1.10 ** | 12.69 ± 1.12 ** |
| Telmisartan | 4.14 ± 0.46 | 35.68 ± 0.86 | 100.52 ± 1.12 ## | 90.28 ± 1.38 ## | 87.20 ± 1.26 ## | 32.96 ± 1.82 ## | 2.07 ± 1.12 ## |
| OJS low | 3.97 ± 0.07 | 37.14 ± 0.41 | 179.55 ± 2.42 ## | 104.28 ± 0.76 ## | 107.20 ± 0.82 # | 22.62 ± 1.70 | 7.01 ± 0.54 ## |
| OJS high | 3.72 ± 0.23 | 35.48 ± 1.25 # | 122.73 ± 4.84 ## | 90.06 ± 1.46 ## | 79.43 ± 2.88 ## | 30.21 ± 2.23 # | 3.11 ± 0.33 ## |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, B.H.; Seo, C.S.; Yoon, J.J.; Kim, H.Y.; Ahn, Y.M.; Eun, S.Y.; Hong, M.H.; Lee, J.G.; Shin, H.K.; Lee, H.S.; et al. The Inhibitory Effect of Ojeoksan on Early and Advanced Atherosclerosis. Nutrients 2018, 10, 1256. https://doi.org/10.3390/nu10091256
Han BH, Seo CS, Yoon JJ, Kim HY, Ahn YM, Eun SY, Hong MH, Lee JG, Shin HK, Lee HS, et al. The Inhibitory Effect of Ojeoksan on Early and Advanced Atherosclerosis. Nutrients. 2018; 10(9):1256. https://doi.org/10.3390/nu10091256
Chicago/Turabian StyleHan, Byung Hyuk, Chang Seob Seo, Jung Joo Yoon, Hye Yoom Kim, You Mee Ahn, So Young Eun, Mi Hyeon Hong, Jae Geon Lee, Hyeun Kyoo Shin, Ho Sub Lee, and et al. 2018. "The Inhibitory Effect of Ojeoksan on Early and Advanced Atherosclerosis" Nutrients 10, no. 9: 1256. https://doi.org/10.3390/nu10091256