Anti-Diabetic Effects and Anti-Inflammatory Effects of Laminaria japonica and Hizikia fusiforme in Skeletal Muscle: In Vitro and In Vivo Model



Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Preparations
2.2. Total Polyphenol Contents of LJ and HF Extracts
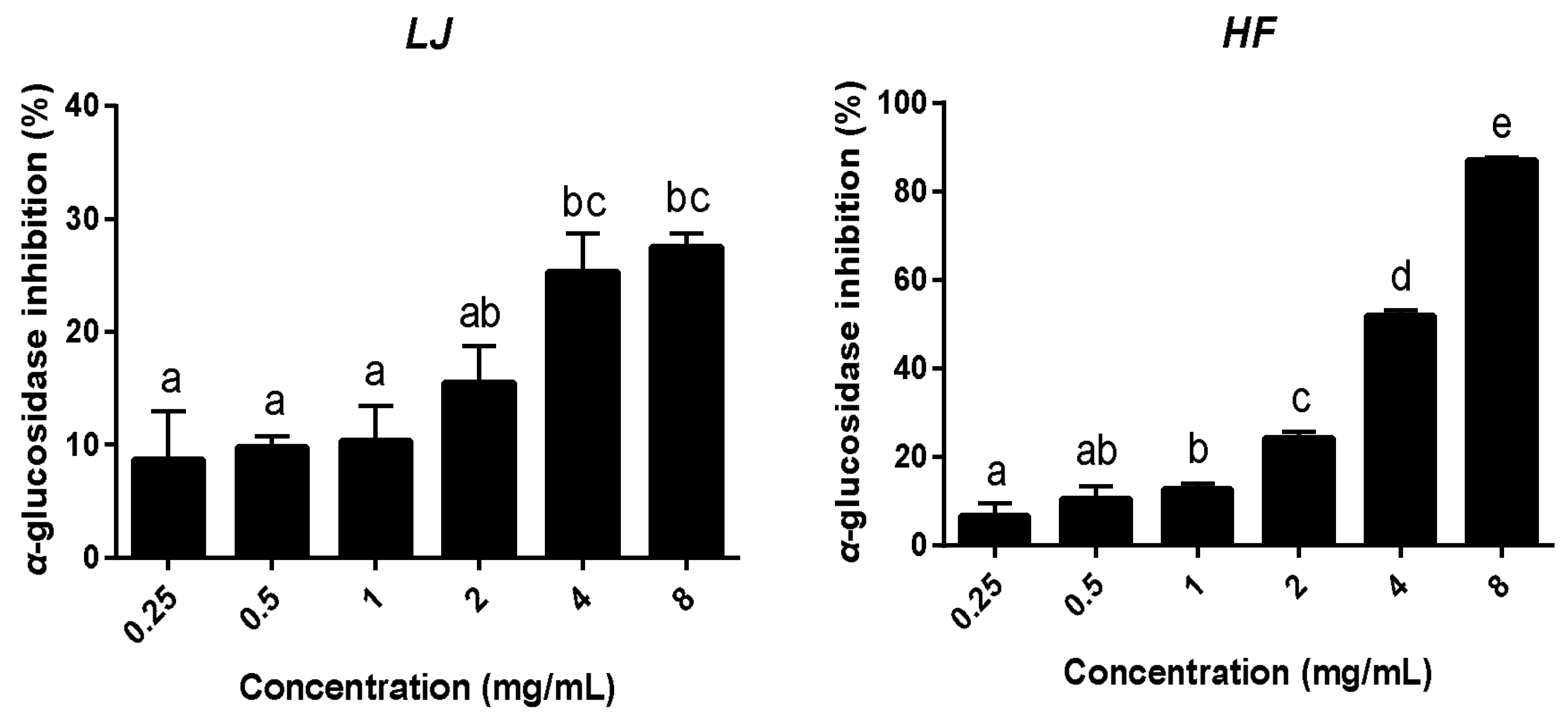
2.3. Alpha-Glucosidase Inhibitory Activities of LJ and HF Extracts
2.4. Animals and Diets
2.5. Cell Culture
2.6. Cell Viability
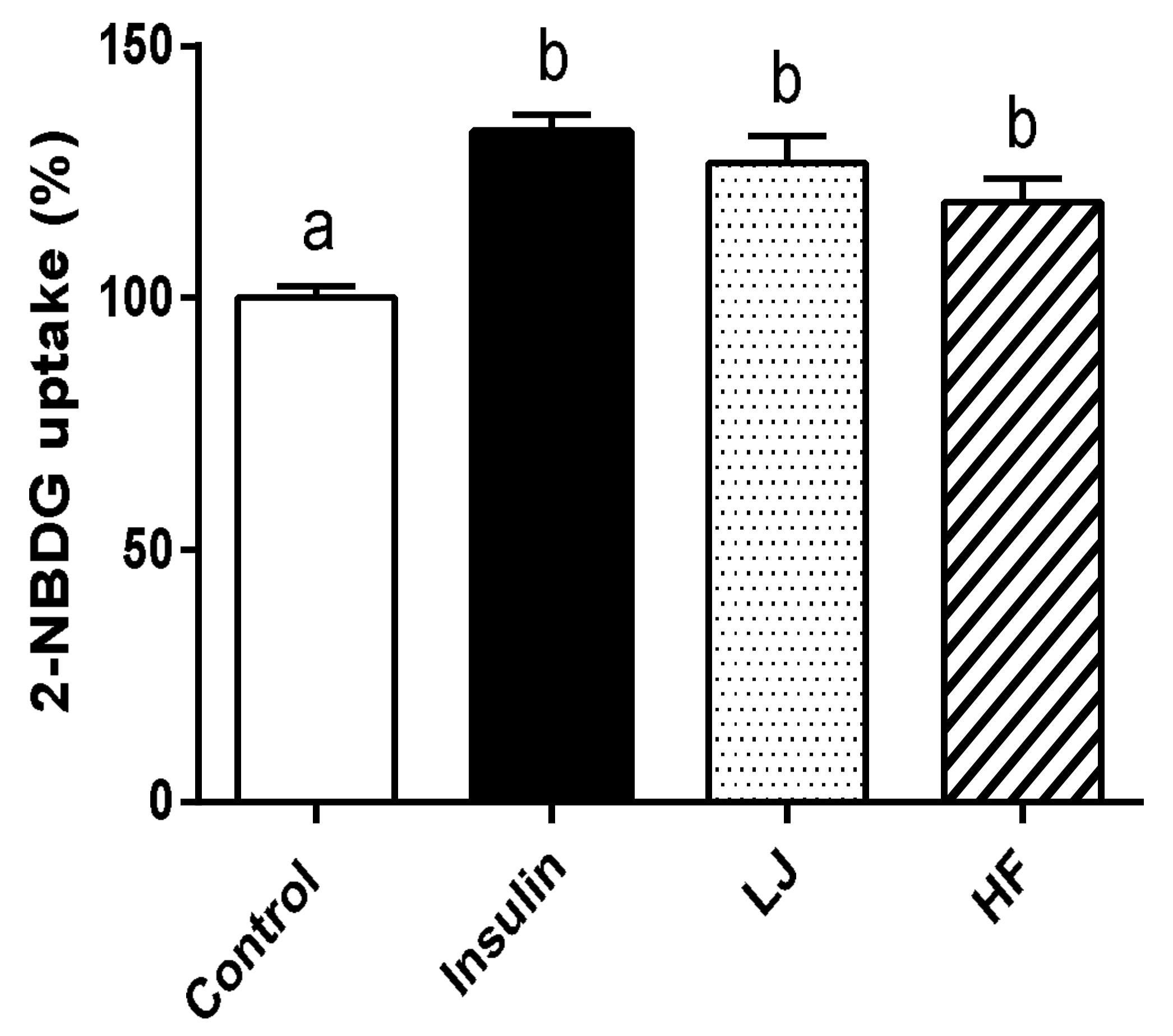
2.7. Measurement Glucose (2-NBDG) Uptake
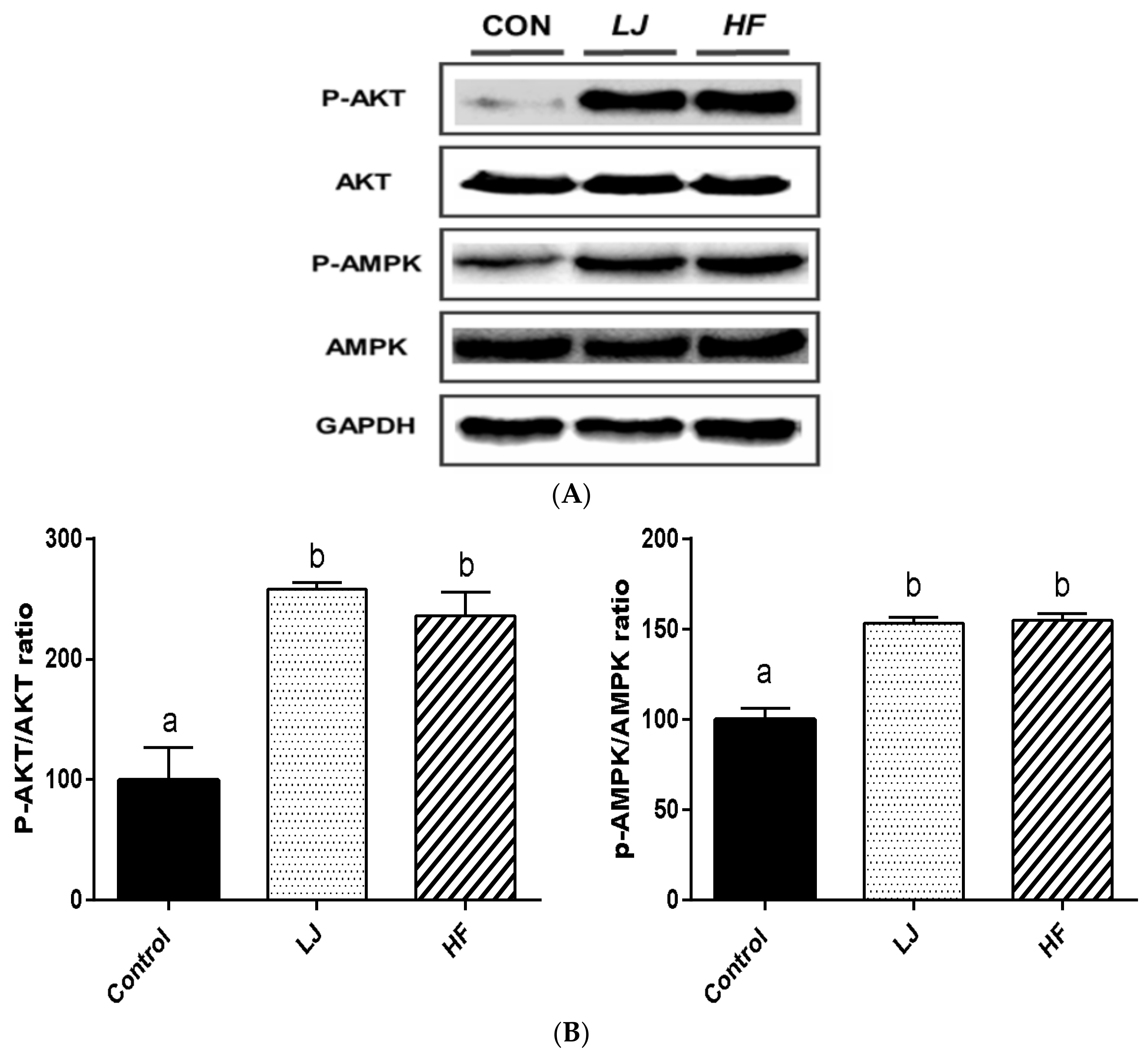
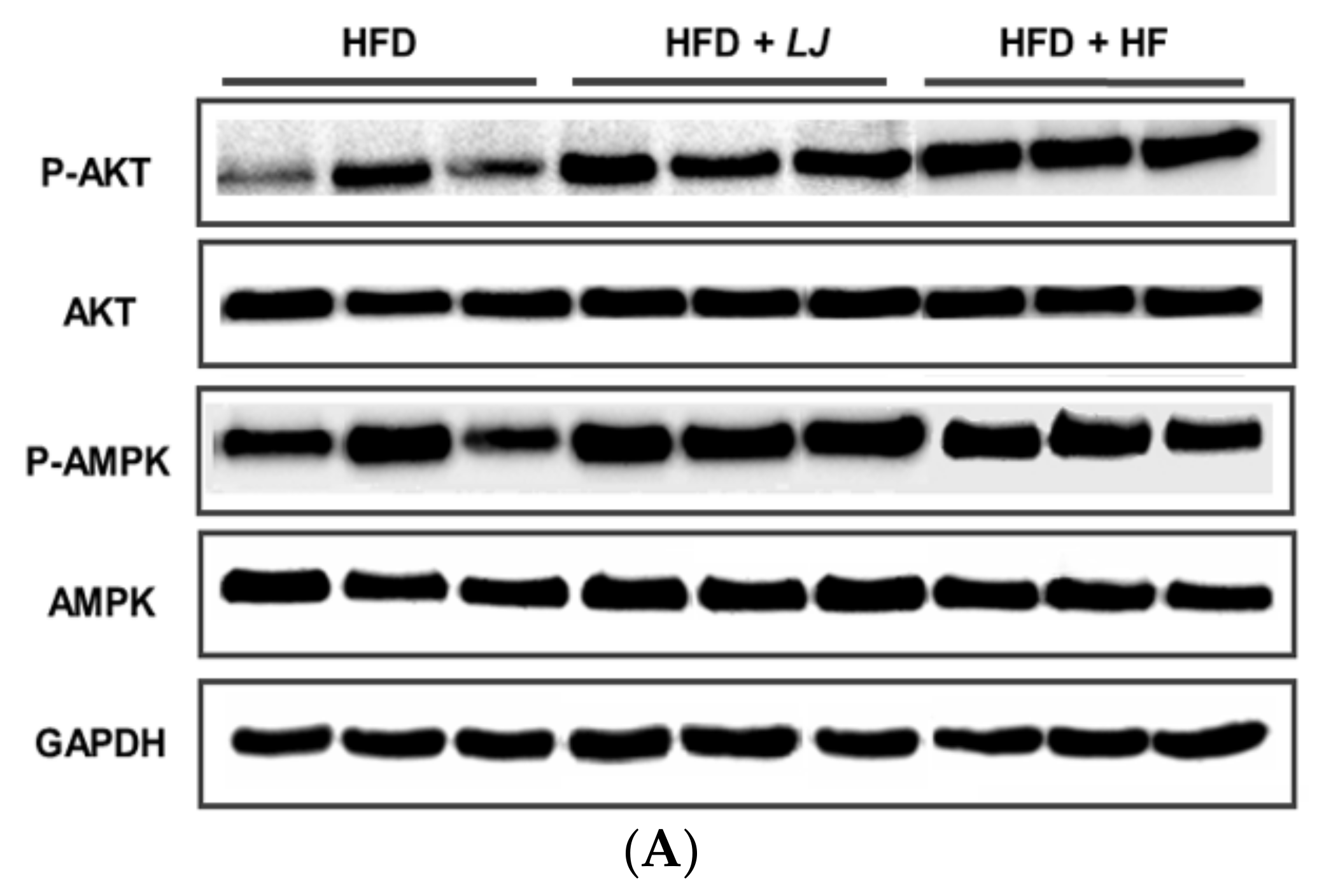
2.8. Western Blotting Analysis
2.9. Inflammation Cytokines Detection by ELISA
2.10. Statistical Analysis
3. Results
3.1. Total Polyphenol Contents and α-Glucosidase Inhibitory Activities of LJ and HF Extracts
3.2. Effects of LJ and HF Extracts on Cell Viability C2C12 Myotubes
3.3. Effects of LJ and HF Extracts on 2-NBDG Uptake in C2C12 Cells
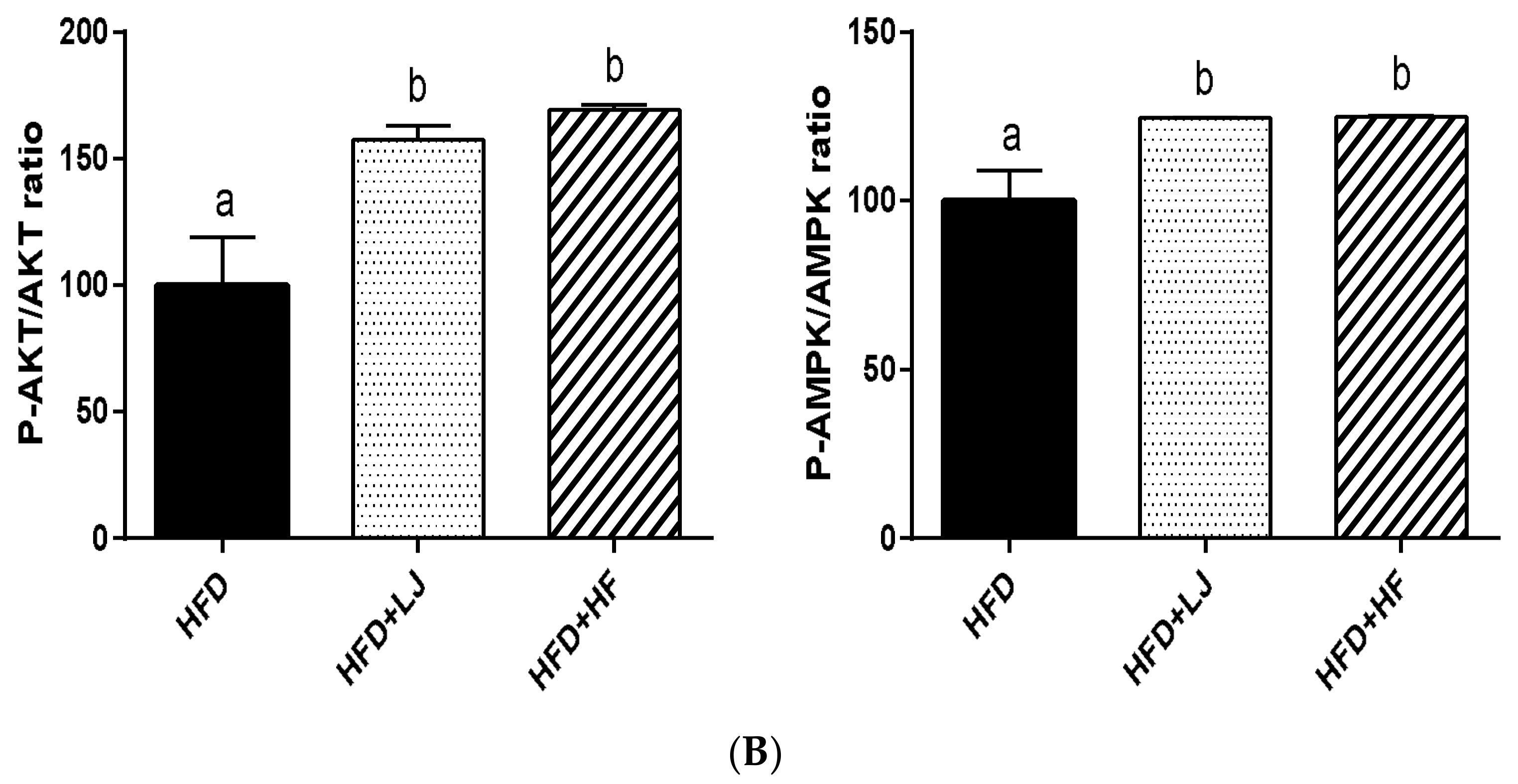
3.4. Effects of LJ and HF on Insulin-Signaling-Involved Proteins in Skeletal Muscle: In Vitro and In Vivo
3.5. Effects of LJ and HF Extracts on Inflammatory Response in C2C12 Muscle Cells Stimulated with LPS
4. Discussion and Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| 2-NBDG | 2-deoxy-2-[(7-nitro-2,1,3-benzoxadizaol-4-yl) amino]-d-glucose |
| Akt | Protein Kinase B |
| AMPK | 5′ adenosine monophosphate-activated protein kinase |
| BMDM | bone marrow derived macrophage |
| DMEM | Dulbecco’s modified eagle medium |
| GAPDH | glyceraldehyde-3-phosphate dehydrogenase |
| HF | Hizikia fusiforme |
| HFD | high fat diet |
| IL | interleukin |
| IR | insulin resistance |
| LJ | Laminaria japonica |
| LFD | low fat diet |
| LPS | lipopolysaccarides |
| MTT | 3-(4,5-dimethyl thiazol-2-yl)-2,5-diphenyl tetrazolium bromide |
| SF | Sargassum Fulvellum |
| T2D | type 2 diabetes |
| TNF-α | tumor necrosis factor-α |
| UP | Undaria pinnatifida |
References
- De Luca, C.; Olefsky, J.M. Inflammation and insulin resistance. FEBS Lett. 2008, 582, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. Diagnosis and classification of diabetes mellitus. Diabetes Care 2004, 27, s5–s10. [Google Scholar]
- Hotamisligil, G.S.; Shargill, N.S.; Spiegelman, B.M. Adipose expression of tumor necrosis factor-alpha: Direct role in obesity-linked insulin resistance. Science 1993, 259, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Shoelson, S.E.; Lee, J.; Goldfine, A.B. Inflammation and insulin resistance. J. Clin. Investig. 2006, 116, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Saltiel, A.R.; Kahn, C.R. Insulin signalling and the regulation of glucose and lipid metabolism. Nature 2001, 414, 799–806. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.; Jacot, E.; Jequier, E.; Maeder, E.; Wahren, J.; Felber, J. The effect of insulin on the disposal of intravenous glucose: Results from indirect calorimetry and hepatic and femoral venous catheterization. Diabetes 1981, 30, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. The amp-activated protein kinase pathway—New players upstream and downstream. J. Cell Sci. 2004, 117, 5479–5487. [Google Scholar] [CrossRef] [PubMed]
- Yagasaki, K. Anti-diabetic phytochemicals that promote GLUT4 translocation via AMPK signaling in muscle cells. Nutr. Aging 2014, 2, 35–44. [Google Scholar]
- Park, S.H.; Lee, S.J.; Jeon, M.; Kim, S.-Y.; Mun, O.-J.; Kim, M.; Kong, C.-S.; Lee, D.-G.; Yu, K.H.; Kim, Y.Y.; et al. Evaluation of biological activities of fermented hizikia fusiformis extracts. J. Life Sci. 2014, 24, 304–310. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Murakami-Funayama, K.; Miyashita, K. Anti-obesity and anti-diabetic effects of fucoxanthin on diet-induced obesity conditions in a murine model. Mol. Med. Rep. 2009, 2, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Jeon, Y.-J. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lahaye, M. Marine algae as sources of fibres: Determination of soluble and insoluble dietary fibre contents in some ‘sea vegetables’. J. Sci. Food Agric. 1991, 54, 587–594. [Google Scholar] [CrossRef]
- Lee, S.-H.; Park, M.-H.; Heo, S.-J.; Kang, S.-M.; Ko, S.-C.; Han, J.-S.; Jeon, Y.-J. Dieckol isolated from Ecklonia cava inhibits α-glucosidase and α-amylase in vitro and alleviates postprandial hyperglycemia in streptozotocin-induced diabetic mice. Food Chem. Toxicol. 2010, 48, 2633–2637. [Google Scholar] [CrossRef] [PubMed]
- Murugan, A.C.; Karim, M.R.; Yusoff, M.B.M.; Tan, S.H.; Asras, M.F.B.F.; Rashid, S.S. New insights into seaweed polyphenols on glucose homeostasis. Pharm. Biol. 2015, 53, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Oh, J.H.; Kim, J.; Lee, Y. Anti-inflammatory and anti-diabetic effects of brown seaweeds in high-fat diet-induced obese mice. Nutr. Res. Pract. 2016, 10, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.-J.; Lee, Y.-S.; Ryu, B.-H. Antitumor effect and immunology activity of seaweeds toward sarcoma-180. Korean J. Fish. Aquat. Sci. 1990, 23, 345–352. [Google Scholar]
- Kang, C.; Jin, Y.B.; Lee, H.; Cha, M.; Sohn, E.-T.; Moon, J.; Park, C.; Chun, S.; Jung, E.-S.; Hong, J.-S.; et al. Brown alga Ecklonia cava attenuates type 1 diabetes by activating ampk and akt signaling pathways. Food Chem. Toxicol. 2010, 48, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.-C.; Wijesinghe, W.; Lee, S.-H.; Kang, S.-M.; Ko, S.-C.; Yang, X.; Kang, N.; Jeon, B.-T.; Kim, J.; Lee, D.-H.; et al. Dieckol isolated from brown seaweed Ecklonia cava attenuates type ІІ diabetes in db/db mouse model. Food Chem. Toxicol. 2013, 53, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.-A.; Lee, S.-H.; Lee, J.-H.; Kang, N.; Oh, J.-Y.; Ahn, G.; Ko, S.C.; Fernando, S.P.; Kim, S.-Y.; Park, S.-J.; et al. A marine algal polyphenol, dieckol, attenuates blood glucose levels by akt pathway in alloxan induced hyperglycemia zebrafish model. RSC Adv. 2016, 6, 78570–78575. [Google Scholar] [CrossRef]
- Kang, S.-M.; Lee, S.-H.; Heo, S.-J.; Kim, K.-N.; Jeon, Y.-J. Evaluation of antioxidant properties of a new compound, pyrogallol-phloroglucinol-6, 6′-bieckol isolated from brown algae, Ecklonia cava. Nutr. Res. Pract. 2011, 5, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Folin, O.; Denis, W. On phosphotungstic-phosphomolybdic compounds as color reagents. J. Biol. Chem. 1912, 12, 239–243. [Google Scholar]
- Watanabe, J.; Kawabata, J.; Kurihara, H.; Niki, R. Isolation and identification of α-glucosidase inhibitors from Tochu-cha (Eucommia ulmoides). Biosci. Biotechnol. Biochem. 1997, 61, 177–178. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Kim, M.H.; Ahn, J.H.; Lee, S.J.; Lee, J.H.; Eum, W.S.; Choi, S.Y.; Kwon, H.Y. The stimulatory effect of essential fatty acids on glucose uptake involves both akt and ampk activation in C2C12 skeletal muscle cells. Korean J. Physiol. Pharmacol. 2014, 18, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Shulman, G.I.; Rothman, D.L.; Jue, T.; Stein, P.; DeFronzo, R.A.; Shulman, R.G. Quantitation of muscle glycogen synthesis in normal subjects and subjects with non-insulin-dependent diabetes by 13C nuclear magnetic resonance spectroscopy. N. Engl. J. Med. 1990, 322, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-S.; SUH, J.H.; SUH, K.-H. Preparation of antibacterial agent from seaweed extract and its antibacterial effect. Korean J. Fish. Aquat. Sci. 2000, 33, 32–37. [Google Scholar]
- Zhang, J.; Tiller, C.; Shen, J.; Wang, C.; Girouard, G.S.; Dennis, D.; Barrow, C.J.; Miao, M.; Ewart, H.S. Antidiabetic properties of polysaccharide-and polyphenolic-enriched fractions from the brown seaweed Ascophyllum nodosum. Can. J. Physiol. Pharmacol. 2007, 85, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Alpha-glucosidase inhibitors. Endocrinol. Metab. Clin. N. Am. 1997, 26, 539–551. [Google Scholar] [CrossRef]
- Krentz, A.J.; Bailey, C.J. Oral antidiabetic agents: Current role in type 2 diabetes mellitus. Drugs 2005, 65, 385–411. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Hao, J.; Wang, W.; Han, Z.; Li, G.; Zhang, L.; Zhao, X.; Yu, G. Insulin sensitizing effects of oligomannuronate-chromium (III) complexes in C2C12 skeletal muscle cells. PLoS ONE 2011, 6, e24598. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.; Liu, M.; Zheng, L.; Guo, Y.; Lin, X. Α-glucosidase inhibition and the in vivo hypoglycemic effect of butyl-isobutyl-phthalate derived from the Laminaria japonica rhizoid. Phytother. Res. 2010, 24, 1588–1591. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, H.; Ando, J.; Hatano, M.; Kawabata, J. Sulfoquinovosyldiacylglycerol as an α-glucosidase inhibitor. Bioorg. Med. Chem. Lett. 1995, 5, 1241–1244. [Google Scholar] [CrossRef]
- Oboh, G.; Ademiluyi, A.O.; Akinyemi, A.J.; Henle, T.; Saliu, J.A.; Schwarzenbolz, U. Inhibitory effect of polyphenol-rich extracts of jute leaf (Corchorus olitorius) on key enzyme linked to type 2 diabetes (α-amylase and α-glucosidase) and hypertension (angiotensin I converting) in vitro. J. Funct. Foods 2012, 4, 450–458. [Google Scholar] [CrossRef]
- Sakamoto, K.; Aschenbach, W.G.; Hirshman, M.F.; Goodyear, L.J. Akt signaling in skeletal muscle: Regulation by exercise and passive stretch. Am. J. Physiol.-Endocrinol. Metab. 2003, 285, E1081–E1088. [Google Scholar] [CrossRef] [PubMed]
- Pessin, J.E.; Saltiel, A.R. Signaling pathways in insulin action: Molecular targets of insulin resistance. J. Clin. Investig. 2000, 106, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.C.; Zierath, J.R. Amp-activated protein kinase signaling in metabolic regulation. J. Clin. Investig. 2006, 116, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Qian, Z.-J.; Ryu, B.; Lee, S.-H.; Kim, M.-M.; Kim, S.-K. Chemical components and its antioxidant properties in vitro: An edible marine brown alga, Ecklonia cava. Bioorg. Med. Chem. 2009, 17, 1963–1973. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-X.; Wijesekara, I.; Li, Y.; Kim, S.-K. Phlorotannins as bioactive agents from brown algae. Process Biochem. 2011, 46, 2219–2224. [Google Scholar] [CrossRef]
- Nieto-Vazquez, I.; Fernández-Veledo, S.; de Alvaro, C.; Lorenzo, M. Dual role of interleukin-6 in regulating insulin sensitivity in murine skeletal muscle. Diabetes 2008, 57, 3211–3221. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.; Febbraio, M. Interleukin-6 and insulin sensitivity: Friend or foe? Diabetologia 2004, 47, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Steensberg, A.; Keller, C.; Starkie, R.L.; Osada, T.; Febbraio, M.A.; Pedersen, B.K. IL-6 and TNF-alpha expression in, and release from, contracting human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E1272–E1278. [Google Scholar] [CrossRef] [PubMed]
- Starkie, R.; Ostrowski, S.R.; Jauffred, S.; Febbraio, M.; Pedersen, B.K. Exercise and IL-6 infusion inhibit endotoxin-induced TNF-α production in humans. FASEB J. 2003, 17, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Haddad, F.; Zaldivar, F.; Cooper, D.M.; Adams, G.R. IL-6-induced skeletal muscle atrophy. J. Appl. Physiol. 2005, 98, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Higashimori, T.; Park, S.-Y.; Choi, H.; Dong, J.; Kim, Y.-J.; Noh, H.-L.; Cho, Y.-R.; Cline, G.; Kim, Y.-B.; et al. Differential effects of interleukin-6 and-10 on skeletal muscle and liver insulin action in vivo. Diabetes 2004, 53, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Van Exel, E.; Gussekloo, J.; de Craen, A.J.; Frölich, M.; Bootsma-van der Wiel, A.; Westendorp, R.G. Low production capacity of interleukin-10 associates with the metabolic syndrome and type 2 diabetes. Diabetes 2002, 51, 1088–1092. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Total Polyphenol Contents (μg Gallic Acid/mL) |
|---|---|
| LJ | 2.084 ± 0.01 (1) |
| HF | 3.215 ± 0.08 |
| Sample | TNF-α Level (pg/mL) | IL-6 Level (pg/mL) | IL-10 Level (pg/mL) |
|---|---|---|---|
| LPS | 30.68 ± 4.68 (1),(2),b | 62.13 ± 2.22 a | 0.28 ± 0.57 a |
| LPS + LJ | 15.03 ± 4.46 a | 433.15 ± 2.64 c | 5.61 ± 1.07 c |
| LPS + HF | 14.82 ± 3.09 a | 106.00 ± 6.75 b | 3.77 ± 1.50 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, S.-y.; Kim, E.; Kang, I.; Lee, M.; Lee, Y. Anti-Diabetic Effects and Anti-Inflammatory Effects of Laminaria japonica and Hizikia fusiforme in Skeletal Muscle: In Vitro and In Vivo Model. Nutrients 2018, 10, 491. https://doi.org/10.3390/nu10040491
Kang S-y, Kim E, Kang I, Lee M, Lee Y. Anti-Diabetic Effects and Anti-Inflammatory Effects of Laminaria japonica and Hizikia fusiforme in Skeletal Muscle: In Vitro and In Vivo Model. Nutrients. 2018; 10(4):491. https://doi.org/10.3390/nu10040491
Chicago/Turabian StyleKang, Sae-ym, Eunyoung Kim, Inhae Kang, Myoungsook Lee, and Yunkyoung Lee. 2018. "Anti-Diabetic Effects and Anti-Inflammatory Effects of Laminaria japonica and Hizikia fusiforme in Skeletal Muscle: In Vitro and In Vivo Model" Nutrients 10, no. 4: 491. https://doi.org/10.3390/nu10040491